

Вопросы ихтиологии, 2020, T. 60, № 1, стр. 106-110
Ископаемая зубатка (Anarhichadidae) из миоценовых отложений острова Сахалин
М. В. Назаркин 1, *, В. В. Платонов 1
1 Зоологический институт РАН − ЗИН
Санкт-Петербург, Россия
* E-mail: m_nazarkin@mail.ru
Поступила в редакцию 27.06.2019
После доработки 09.07.2019
Принята к публикации 09.07.2019
Аннотация
В отложениях среднего−позднего миоцена глубоководной свиты кураси Юго-Западного Сахалина обнаружен разрушенный скелет головы зубатки Anarhichadidae gen. et sp. indet. Это наиболее древняя находка ископаемого представителя семейства. Характер озубления нижней челюсти и сошника миоценовой рыбы сходен с таковыми современного атлантического вида – пятнистой зубатки Anarhichas minor.
К семейству зубаток (Zoarcoidei: Anarhichadidae) относятся два рода и пять видов современных рыб, широко распространённых в северных частях Тихого и Атлантического океанов, а также в Арктических морях (Макушок, 1958; Барсуков, 1959; Barsukov, 1986; Mecklenburg, 2003; Mecklenburg et al., 2016). Взрослые зубатки ведут донный образ жизни на разных глубинах. Тихоокеанские виды населяют прибрежные мелководные биотопы с крупными камнями и зарослями растительности, атлантические предпочитают в основном большие глубины – до 1700 м (Андрияшев, 1954; Барсуков, 1959; Mecklenburg et al., 2002, 2016). Для части видов отмечены суточные вертикальные, а также протяжённые нерестовые и нагульные миграции (Mecklenburg et al., 2016). Молодь некоторых зубаток пелагическая и часто встречается вдали от берегов в открытом море (Андрияшев, 1954; Линдберг, Красюкова, 1975), в то время как у других видов она придерживается больших глубин (Барсуков, 1959).
Главной морфологической особенностью зубаток является строение их зубной системы, приспособленной для питания организмами с твёрдым наружным скелетом (Андрияшев, 1954; Макушок, 1958). Зубы на челюстях, сошнике и нёбных костях этих рыб дифференцированы на клыковидные, конические и бугорковидные; лобные кости сращены между собой; заглазничная часть крыши черепа у большинства видов сильно сужена; парасфеноид очень высокий за счёт мощного киля, поддерживающего мышцы нёбной дуги (Макушок, 1958).
С древних времён и по настоящее время зубатки являются важным объектом прибрежного рыболовства, поэтому их кости нередко находят в археологических памятниках, оставленных прибрежным населением северных широт (Назаркин, Лебединцев, 1993; Bradley, Spiess, 1994; Edvardsson et al., 2004; Spiess, 2011). Существует единственная достоверная находка представителя семейства дочетвертичного возраста: передний челюстной зуб из плиоценовых отложений формации Coralline Crag в Великобритании был описан как принадлежащий современному виду Anarhichas lupus (Newton, 1891).
Дизартикулированный скелет головы зубатки был недавно обнаружен нами в отложениях среднего−позднего миоцена курасийской свиты Сахалина. Богатый комплекс ископаемых остатков из этой свиты включает ~ 33 видов рыб (Назаркин, 2018), бóльшая часть которых является представителями мезопелагического сообщества с доминированием миктофид (Myctophidae) и глубоководных корюшек (Bathylaginae). Остатки зубатки в этом комплексе найдены впервые и представлены единственным и сильно дизартикулированным экземпляром. На настоящий момент это самая древняя ископаемая зубатка и первая находка доплиоценового представителя семейства. В современных морях северо-западной части Тихого океана обитает только один вид семейства – дальневосточная зубатка A. orientalis, распространённый от Японского моря на юге до зал. Аляска, Чукотского и Бофортова морей на севере, а к востоку до канадской Арктики (Андрияшев, 1954; Барсуков, 1959; Линдберг, Красюкова, 1975; Шейко, Фёдоров, 2000; Фёдоров и др., 2003; Mecklenburg et al., 2016).
МАТЕРИАЛ И МЕТОДИКА
Экземпляр миоценовой зубатки вместе с противоотпечатком депонирован в палеонтологической секции ихтиологической коллекции ЗИН РАН (ЗИН 457п). Очистку образца проводили иглами под стереомикроскопом. Измерения выполнены штангенциркулем с точностью до 0.1 мм. Стандартная длина (SL) восстановлена по длине зубной кости через прямую пропорцию относительно современной сеголетки A. minor. Рисунок сделан на основе цифровых фотографий обеих частей образца. В качестве сравнительного материала изучено по 1 экз. молоди (сеголеток) каждого вида зубаток: A. denticulatus Krøyer, ЗИН 56481, SL 92 мм; A. lupus Linnaeus, ЗИН 52483, SL 116 мм; A. minor Olafsen, ЗИН 35520, SL 104 мм; A. orientalis Pallas, ЗИН 53000, SL 130 мм; Anarrhichthys ocellatus Ayres, ЗИН 47427, SL 497 мм.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Anarhichadidae gen. et sp. indet.
Описание. Экземпляр представляет собой разрушенный скелет головы и шести передних туловищных позвонков. Длина фрагмента до заднего края последнего позвонка составляет 31.9 мм (рис. 1а, 1б). Лобные кости, по-видимому, срослись между собой; позади орбиты заметны их треугольные латеральные выросты и невысокий медиальный гребень (рис. 1в). В отличие от большинства современных зубаток frontalia данного экземпляра не сужаются позади орбиты. Позади frontalia различимы supraoccipitale и крупное pteroticum, снабжённое длинным продольным гребнем. Ниже переднего края frontalia имеется костный элемент, представляющий собой, по-видимому, фрагмент ethmoidale laterale. Под задней частью лобных костей видны sphenotici правой и левой сторон. Basioccipitale высокое, латерально трапециевидное, с мелко иззубренным вентральным краем. Parasphenoideum латерально плоское, очень высокое, с сильно развитыми восходящими отростками; его задний конец приострён. К его антевентральному краю прочно присоединено крупное vomer; при этом никаких швов между этими костями не заметно, и потому форма сошника не может быть распознана. На переднем крае vomer имеются три довольно крупных тупоконических зуба, а также различимы два углубления – места прикрепления более мелких зубов. Ещё один крупный тупоконический зуб, отделённый, вероятно, от сошника, находится между парасфеноидом и нёбной костью. Два мощных давящих зуба (моляра) также отделены от сошника и лежат в непосредственной близости от него спереди и сзади.
Рис. 1.
Anarhichadidae gen. et sp. indet., ЗИН 457п: а – отпечаток, б − противоотпечаток, в – прорисовка образца; aar – anguloarticulare, boc – basiocciptale, d – dentale, ecp – ectopterygoideum, el – ethmoidale laterale, fr – frontale, hm – hyomandibulare, pal – palatinum, pop – praeoperculum, prs – parasphenoideum, ptr – pteroticum, q – quadratum, soc – supraoccipitale, sph – sphenoticum, sy – symplecticum. Масштаб: 5 мм.
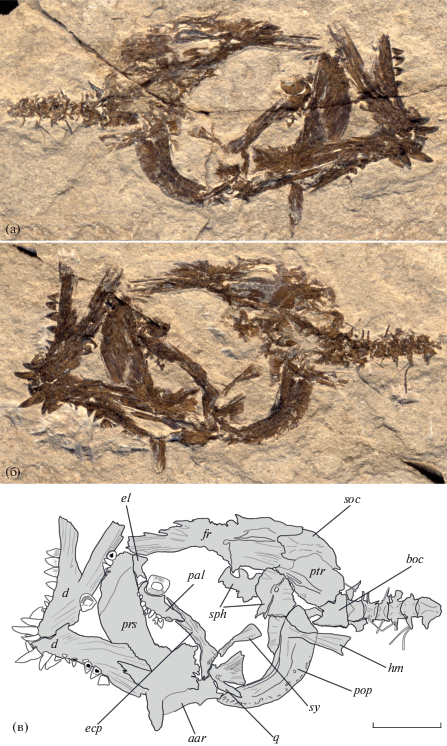
Hyomandibulare прямое и высокое. Praeoperculum полулунное, с тупым углом между ветвями и несколько расширенным верхним краем. Перед предкрышкой расположены вееровидное quadratum и палочковидное symplecticum. Ectopterygoideum довольно длинное, проксимально широкое, плавно сужающееся дистально. Palatinum в 1.5 раза короче ectopterygoideum, высокое; на его вентральной поверхности имеется не менее восьми тупоконических некрупных зубов, расположенных, вероятно, в один ряд. Нёбные зубы мельче сошниковых и нижнечелюстных зубов.
Dentale высокое, компактное, с умеренно скошенным назад симфизным краем и невысоким восходящим отростком. Имеется, по крайней мере, два ряда нижнечелюстных зубов. В наружном ряду в передней части челюсти по четыре высоких конических клыковидных зуба. Зубы внутреннего ряда, общим числом не менее восьми, также конические, довольно широкие в основании, увеличиваются в размерах спереди назад и, по-видимому, слегка сжаты латерально. В нижней части наиболее задних нижнечелюстных зубов заметна очень тонкая вертикальная струйчатость. Моляровидные зубы в нижней челюсти, по-видимому, отсутствуют. Фрагмент anguloarticulare, частично перекрытый парасфеноидом, распознаётся ниже quadratum. Сохранность этой кости не позволяет различить особенности её строения.
Сохранившиеся позвонки высокие, симметричные, латерально почти квадратные; лишь у 6-го позвонка длина заметно превышает высоту. С 4-го позвонка заметны сильно выступающие парапофизы. Обломки нескольких пар плевральных рёбер также видны в задней части образца; при этом рёбра 4-го и 5-го позвонков прикреплены к соответствующим парапофизам.
Измерения. Длину нёбного и сошникового рядов зубов не удаётся измерить точно; видимая часть нёбного ряда (3.4 мм) несколько больше сошникового (2.7 мм). Длина frontale 13.2 мм; длина dentale 10.4 мм; длина нижней челюсти ~ 14.6 мм; высота praeoperculum 9.3 мм; наибольшее расстояние от переднего края vomer до заднего края parasphenoideum 13.6 мм; высота наибольшего клыковидного зуба нижней челюсти 1.9 мм; высота заднего зуба нижней челюсти 1.0 мм; диаметр наиболее крупного сошникового моляра 1.8 мм; высота наибольшего нёбного зуба 0.7 мм. Восстановленная длина (SL) ископаемой рыбы составляет ~ 100 мм. Молодь современных зубаток достигает таких размеров в течение первого года жизни.
Сравнительные замечания. Уникальное строение зубной системы с дифференциацией на клыковидные, конические и давящие моляровидные зубы свидетельствует о принадлежности ископаемой рыбы к семейству зубаток. Кости головы всех современных видов семейства имеют довольно сходные пропорции и очертания, что связано с пищевой специализацией этих рыб (Макушок, 1958; Барсуков, 1959). Небольшие видовые и родовые особенности современных рыб большей частью не могут быть распознаны на ископаемом материале. Все кости, очертания которых хорошо различимы у миоценового экземпляра, имеют форму, сходную с таковой у современных рыб. Однако относительно большая ширина frontalia позади орбиты и, соответственно, менее сжатый латерально краниальный отдел черепа миоценовой формы более соответствуют строению этого отдела у видов, распространённых в бассейне Северной Атлантики – синей зубатки A. denticulatus и в меньшей степени пятнистой зубатки A. minor (Андрияшев, 1954. Рис. 110 ; Барсуков, 1959. Табл. XIX ).
Число и форма зубов миоценовой зубатки демонстрируют значительное сходство с таковыми у A. minor. У молоди современного вида, как и у ископаемой формы, клыковидные зубы нижней челюсти высокие и хорошо выраженные; задние зубы расположены в один ряд, относительно высокие, тупоконические, слегка сжаты латерально и обладают слабой струйчатостью в основании (рис. 2). Число зубов, расположенных позади передних клыков, у современного вида шесть–восемь, что совпадает с этим числом у миоценовой формы. Сошниковые зубы у молодых рыб современного вида образуют неровный ряд из четырёх тупоконических зубов, к которым сзади примыкает один крупный моляр. Наконец, длина нёбного ряда зубов у обеих зубаток почти равна длине сошникового ряда.
Рис. 2.
Озубление Anarhichas minor, ЗИН 35520, SL 104 мм; зубы: dt – нижнечелюстные, pmt – верхнечелюстные, vt – сошниковые.
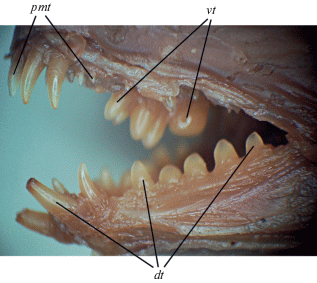
От синей зубатки A. denticulatus миоценовая форма отличается длинным рядом сошниковых зубов (в несколько раз короче нёбного у современного вида), хорошо выраженными клыковидными (слабо дифференцированы у современного), а также более тупыми (не остроконическими) задними зубами нижней челюсти. От полосатой A. lupus миоценовая зубатка отличается более высокими клыками и более острыми зубами в задней части нижней челюсти, а также отсутствием в ней моляров; кроме того, у молоди современного вида на сошнике практически все зубы давящего типа, а тупоконические зубы отсутствуют. От обоих тихоокеанских видов – восточной A. orientalis и угревидной Anarrhichthys ocellatus зубаток − миоценовая форма отличается меньшим числом задних зубов нижней челюсти, расположенных в один ряд (против не менее 12 зубов в два ряда); отсутствием нижнечелюстных моляров; меньшим числом зубов на сошнике (8 против не менее 10).
Анализ молекулярно-генетических данных определяет время появления семейства Anarhichadide началом позднего миоцена, 10.2 (8.4–12.3) млн лет назад (Радченко, 2016, 2017). Поскольку накопление осадков курасийской свиты происходило в среднем и позднем миоцене в пределах серравалия–тортона (Гладенков и др., 2002), т.е. 7.2–13.8 млн лет назад, находка зубатки в её слоях не противоречит этому выводу. Являясь на настоящий момент самой древней, данная находка намекает на тихоокеанское происхождение семейства. Некоторое морфологическое сходство миоценовой тихоокеанской зубатки с атлантическими видами может быть свидетельством того, что эти последние являются вытесненными на периферию ареала реликтами. Всё же делать определённые выводы на основе этой единственной находки преждевременно.
Список литературы
Андрияшев А.П. 1954. Рыбы северных морей СССР. М.; Л.: Изд-во АН СССР, 566 с.
Барсуков В.В. 1959. Фауна СССР. Рыбы. Т. V. Вып. 5. Сем. Зубаток (Anarhichadidae). М.; Л.: Изд-во АН СССР, 196 с.
Гладенков Ю.Б., Баженова О.К., Гречин В.И. и др. 2002. Кайнозой Сахалина и его нефтегазоносность. М.: ГЕОС, 223 с.
Линдберг Г.У., Красюкова З.В. 1975. Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Ч. 4. М.; Л.: Наука, 463 с.
Макушок В.М. 1958. Морфологические основы стихеевых и близких к ним семейств рыб (Stichaeidae, Blennioidei, Pisces) // Тр. ЗИН АН СССР. Т. 25. С. 4–129.
Назаркин М.В. 2018. Особенности миоценовой ихтиофауны Дальнего Востока России // Матер. Междунар. науч. конф., посвящ. памяти В.Л. Контримавичуса “Биологические проблемы Севера”. Магадан: Изд-во ИБПС ДВО РАН. С. 445–447.
Назаркин М.В., Лебединцев А.И. 1993. Остатки рыб из древних стоянок северного побережья Охотского моря // Вопр. ихтиологии. Т. 33. Вып. 4. С. 589–592.
Радченко О.А. 2016. Время эволюции бельдюговидных рыб подотряда Zoarcoidei (Perciformes) по данным об изменчивости ДНК // Там же. Т. 56. № 4. С. 440−452.https://doi.org/10.7868/S0042875216040160
Радченко О.А. 2017. Молекулярная систематика и филогения бельдюговидных рыб. М.: ГЕОС, 384 с.
Федоров В.В., Черешнев И А., Назаркин М.В. и др. 2003. Каталог морских и пресноводных рыб северной части Охотского моря. Владивосток: Дальнаука, 204 с.
Шейко Б.А., Федоров В.В. 2000. Рыбообразные и рыбы // Каталог позвоночных Камчатки и сопредельных морских акваторий. Петропавловск-Камчатский: Камчат. печат. двор. С. 7–69.
Barsukov V.V. 1986. Anarhichadidae // Fishes of the North-eastern Atlantic and the Mediterranian. V. III / Eds. Whitehead P.J.P. et al. Paris: UNESCO. P. 1113–1116.
Bradley J.W., Spiess A.E. 1994. Two shell middens on Indian neck, Wellfleet, Massachusetts: the excavation of Fred A. Luce // Bull. Massachusetts Archaeolog. Soc. V. 55. № 2. P. 45–60.
Edvardsson R., Perdikaris S., McGovern T.H. et al. 2004. Coping with hard times in NW Iceland: zooarchaeology, history, and landscape archaeology at Finnbogastadir in the 18-th century // Archaeologia Islandica. V. 3. P. 20–47.
Mecklenburg C.W. 2003. Family Anarhichadidae Bonaparte 1846 – wolffishes // Calif. Acad. Sci. Annotated checklists of fishes. № 10. 6 p.
Mecklenburg C.W., Mecklenburg T.A., Thorsteinson L.K. 2002. Fishes of Alaska. Bethesda, Maryland: Amer. Fish. Soc., 1037 p.
Mecklenburg, C.W., Mecklenburg T.A., Sheiko B.A., Steinke D. 2016. Pacific Arctic marine fishes. Akureyri, Iceland: CAFF, 377 p.
Newton E.T. 1891. The Vertebrata of the Pliocene deposits of Britain. London: Mem. Geol. Survey UK., 137 p.
Spiess A.E. 2011. Cape Cod National Seashore Archaeological Survey: faunal analysis // Chapters in the Archaeology of Cape Cod. V. IV. Faunal analysis and metallurgical analysis from the Cape Cod National Seashore Archaeological Survey / Ed. McManamon F.P. Wellfleet, MA: Nat. Park Service. P. 13–61.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии