

Вопросы ихтиологии, 2020, T. 60, № 1, стр. 26-39
Новые данные по ихтиофауне прибрежья Южного Вьетнама
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН – ИПЭЭ РАН
Москва, Россия
2 Институт океанологии им. П.П. Ширшова РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 03.06.2019
После доработки 10.06.2019
Принята к публикации 04.07.2019
Аннотация
Сообщено о новых находках девяти видов рыб из семейств Congridae, Gonorhynchidae, Scorpaenidae, Serranidae, Nemipteridae и Tetraodontidae, из которых семь видов впервые указываются для фауны страны, а два вида (Chaunax breviradius и Torquigener gloerfelti) ранее были известны только по экземплярам типовой серии. Уточнена морфологическая изменчивость рассматриваемых видов, для T. gloerfelti впервые описана онтогенетическая изменчивость признаков.
Морская ихтиофауна прибрежья Вьетнама всё ещё изучена очень слабо. Достаточно сказать, что в компилятивном списке морских рыб этой страны (Nguyen, Tran, 1994; Nguyen, 1999) было указано лишь около 1600 видов, тогда как в списке рыб Южно-Китайского моря, составленном приблизительно в то же время, отмечено 3365 видов (Randall, Liem, 2000), большинство из которых можно ожидать и во вьетнамских водах. Наряду с сообщениями о новых находках в последние десятилетия из прибрежных вод Вьетнама нередки и описания новых для науки видов (Fricke et al., 2019). Помимо задач зоогеографии и систематики выявление видового состава и особенностей распространения рыб в прибрежных морских водах Вьетнама необходимо для разработки мер охраны и рационального использования морских биоресурсов. Однако пока что эти работы проводятся всё ещё недостаточно активно.
В ходе траловой съёмки, проводившейся в зал. Нячанг и Ванфонг в 2005–2010 гг., и анализа уловов местного промысла в прибрежных водах Южного Вьетнама и прилегающих островов от зал. Ванфонг до Сиамского зал. (2005–2013 гг.) мною была собрана обширная коллекция, включающая многочисленные новые для ихтиофауны страны находки и ряд новых для науки видов, частью уже публиковавшиеся в различных журнальных статьях. Настоящее сообщение продолжает серию работ по результатам обработки вьетнамской коллекции и посвящено девяти видам рыб из шести семейств, из которых семь видов были впервые обнаружены в водах Вьетнама. Некоторые из этих видов (Chaunax breviradius, Torquigener gloerfelti) до сих пор были известны науке только по их типовым сериям.
МАТЕРИАЛ И МЕТОДИКА
Методика изучения соответствует общепринятой для конкретных групп (Hubbs, Lagler, 1958; Eschmeyer et al., 1973; Caruso, 1989; Nakabo, 2002). Координаты точек сбора даются так, как они указаны на этикетках. В работе использованы следующие сокращения: D (ID, IID), A, P, V, С – соответственно спинной (первый и второй), анальный, грудные, брюшные и хвостовой плавники; sp. br – число жаберных тычинок в наружном ряду на 1-й дуге, LL – число чешуй/пор боковой линии, squ – число поперечных рядов чешуй от жаберного отверстия до основания С, SL – стандартная длина, TL – полная длина, n – число экземпляров, ЗИН – Зоологический институт РАН, Санкт-Петербург; ИО РАН – Институт океанологии РАН, Москва.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
(рис. 1а)
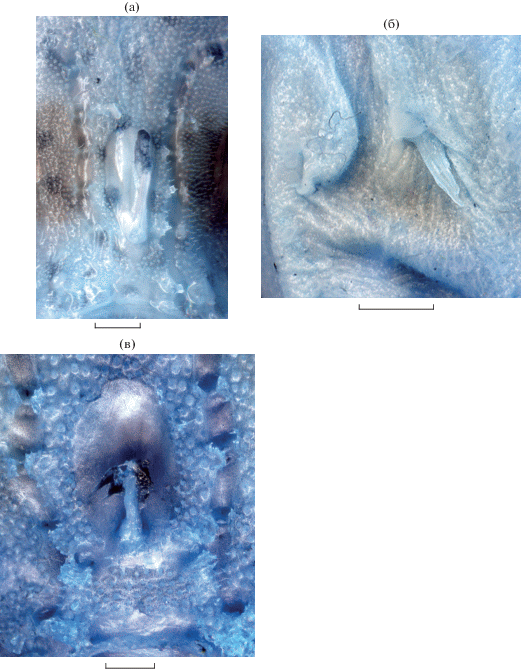
Рис. 1.
Conger philippinus TL 220 мм (а), Gonorhynchus abbreviatus SL 164 мм (б), Chaunax breviradius SL 63 мм (в, г – вид сверху и сбоку), C. abei SL 126 мм (д), общий вид.

Материал. ИО РАН б/№, 2 экз. TL 165–220 мм, Вьетнам, зал. Нячанг, уловы местного промысла, весна–лето 2007 г., коллектор А.М. Прокофьев.
Описание. Начало D расположено заметно позади конца прижатого к телу P, промежуток между ними составляет 71.4–72.7% длины Р. Преанальных пор 35–37, в промежутке между вертикалями начала Р и А 31–32 поры. Супраорбитальных пор две или три (две крупные этмоидные поры, у экз. TL 165 мм присутствует одна маленькая пора в промежутке между задней этмоидной порой и задней ноздрей); супратемпоральная пора одна, инфраорбитальных пор шесть (последняя расположена на линии угла рта), преоперкуло-мандибулярных пор девять. Длина интермаксиллярного зубного пятна больше ширины, максиллярные зубы однорядные, на нижней челюсти в присимфизной четверти её длины имеется дополнительный внутренний ряд зубов. Лучей Р 14. Окраска фиксированных рыб светлая, кремово-коричневая, белеющая на вентральной стороне, плавники светлые, почти белые, дистальный край С и прилегающих участков D и А черноватый.
Некоторые измерения. В % TL: длина головы 11.8–13.3, длина рыла 2.7–3.3, горизонтальный диаметр глаза 2.1–2.4, ширина межглазничного промежутка 2.3–3.0, длина ротовой щели 3.6–4.9, длина Р 4.2–5.0, расстояние от конца Р до начала D 3.0–3.6, высота тела на уровне начала А 5.0–5.5, предорсальное и преанальное расстояния соответственно 19.6–21.8 и 35.5–36.4.
Замечания. Вид был описан от Филиппин и недавно указан из вод Тайваня и Вьетнама (Kanazawa, 1958; Smith, Ho, 2018). Во Вьетнаме указан с рынков города Ханой (вероятно, пойман в Тонкинском зал.) и городов Фантьет и Муйнэ (зал. Фантьет). Для зал. Нячанг указывается впервые. Изученные экземпляры хорошо соответствуют имеющимся описаниям, за исключением неожиданной изменчивости числа супраорбитальных пор (две или три против четырёх, только две передние (этмоидные) поры хорошо развиты) и несколько меньшего числа лучей Р (14 против 15 или 16). Эти различия отражают не более чем индивидуальную изменчивость.
(рис. 1б)
Материал. ИО РАН б/№, 22 экз. SL 30–48 мм и 1 экз. SL 164 мм, Вьетнам, зал. Нячанг и Ванфонг, сборы 2005–2007 гг., коллектор А.М. Прокофьев.
Описание. D 10–11, A 7–8, P 10–11, V 8; C I + + 9 + 8 + I; squ 160–170. Рыло длинное, заострённое; глаз округлый, умеренно крупный, затянут прозрачной кожей; рот маленький, нижний, поперечный; жаберные перепонки широко сращены с истмусом, жаберное отверстие не переходит на вентральную поверхность головы. Дистальные концы нижней губы не образуют разветвлённого усика. Начало D расположено позади вертикали прикрепления V; V не достигают ануса; P прикрепляются вблизи вентрального края тела; C вырезан, его лопасти равной длины, закруглены. Чешуя очень мелкая, с серией мелких шипиков по свободному краю, за счёт которых рыба сильно шероховатая на ощупь; покрывает все тело и голову, включая всё рыло, нижнюю поверхность головы, в том числе нижнюю поверхность ветвей нижней челюсти, жаберную перепонку и истмус, горло, грудь и брюхо; заходит на лучи всех плавников. С внутренней стороны у верхнего края основания P и у наружного края основания V отходит длинная, быстро сужающаяся назад и заострённая на вершине аксиллярная лопасть; длина грудной и брюшной аксиллярных лопастей одинакова и составляет 7.3%SL при SL 164 мм.
Некоторые измерения (по экз. SL 164 мм). В % SL: длина головы 24.4, максимальная и минимальная высота тела соответственно 12.2 и 4.9, длина хвостового стебля 12.2; предорсальное, преанальное, превентральное, пектовентральное и вентроанальное расстояния соответственно 69.5, 82.3, 67.1, 42.1 и 15.2; длина оснований D и А соответственно 6.7 и 4.9, высота D и А соответственно 11.6 и 9.8, длина P, V, наибольших и наименьших ветвистых лучей С соответственно 18.3, 11.0, 10.4 и 7.3; расстояние от центра ануса до начала А 2.4. В % длины головы: длина рыла 37.5, горизонтальный диаметр глаза 21.3, ширина межглазничного промежутка 20.0, преоральная длина 20.0, ширина рта 7.5, длина усика 10.0.
Окраска фиксированных рыб коричневая сверху, становящаяся розовато-бежевой ниже средней линии тела и белой на брюхе. Усик белый, самый его кончик чёрный. Ротожаберная полость, края жаберных отверстий, анус, парные плавники, дистальные концы D и C, а также задний край A чёрные; в основании C нерезкое тёмное пятно; проксимальная часть C, базальная часть D и бóльшая часть A светлые. Окраска тем ярче и контрастнее, чем мельче размер рыб.
Замечания. Отсутствие раздвоенных усиковидных окончаний дистальных отделов нижней губы и меньшее число лучей V (восемь против девяти у других видов рода) являются характерными признаками данного вида (Grande, 1999). Ранее был известен из Японского и Восточно-Китайского морей на юг до Тайваньского пролива и из тихоокеанских вод Японии; в Южно-Китайском море отмечался только в его северной части у берегов Тайваня (Grande, 1999; Shao et al., 2008). Для вод Вьетнама ранее указан не был (Nguyen H., Nguyen N., 1994), однако, по моим данным, должен быть здесь нередок. В зал. Нячанг и Ванфонг молодь этого вида SL < 50 мм ловилась при тралениях на глубинах 40–90 м, однако взрослые рыбы на столь малых глубинах, по-видимому, исключительно редки. Мною был обнаружен единственный экземпляр SL 164 мм, по-видимому, происходящий с глубин < 100 м, так как он встречен в уловах на рынке Бэ (Cho Cua Be), где разгружаются малые суда, промышляющие на таких глубинах.
(рис. 1в, 1г, 2а, 2б)
Материал. Всего 43 экз. из Вьетнама: ИО РАН б/№, 14 экз. SL 36–63 мм, г. Нячанг, рыбный рынок Бэ, 24.05–19.06.2005 г.; ИО РАН б/№, 5 экз. SL 28–44 мм, 12°40′000–12°34′970 с.ш., 109°30′050–109°30′013 в.д., 96–119 м, трал № 1, 30.05–01.06.2007 г.; ИО РАН б/№, 6 экз. SL 24–52 мм, 12º40′950″–12°36′924 с.ш., 109°29′752–109°30′002 в.д., 97–119 м, трал № 5, 30.05–01.06.2007 г.; ИО РАН б/№, 1 экз. SL 34 мм, 12°27′987–12°28′000 с. ш., 109°29′035–109°26′283 в.д., 38–51 м, трал № 8, 30.05–01.06.2007 г.; ИО РАН б/№, 4 экз. SL 28–45 мм, 12°40′019–12°37′463 с.ш., 109°30′414–109°30′402 в.д., 97–119 м, трал № 1, 12.05.2007 г.; ИО РАН б/№, 5 экз. SL 30–50 мм, 12°40′036–12°37′466 с.ш., 109°30′256–109°30′307 в.д., 128–150 м, трал № 1, 17.05.2007 г.; ЗИН № 55995, 2 экз. SL 30–40 мм, 12°18′000–12°16′736 с.ш., 109°25′497–109º25′485 в.д., 55–68 м, трал № 1, 06.06.2006 г.; ЗИН № 55996, 6 экз. SL 28–45 мм, 12°10′000–12°06′985 с.ш., 109°26′535–109°25′992 в.д., 91–95 м, трал № 1, 31.05.2006 г. Коллектор А.М. Прокофьев.
Сравнительный материал. Chaunax abei Le Danois, 1978: ИО РАН б/№, 1 экз. SL 126 мм (рис. 1д, 2в), Филиппины, 27.09.1971 г. D I + I + I + + 11, A 7, P 12, V 2/3, C 8; невромастов в надглазничном, верхнечелюстно-подглазничном, нижнечелюстно-пекторальном и туловищном каналах соответственно 11, 12, 26 и 36/38; верхне- и нижнепредкрышечных невромастов соответственно три и четыре.
Описание (рис. 1в, 1г). D I + I + I + 11, A (6)7, P 10–12, V 0–3, C 8 (ветвистых шесть). Невромастов в надглазничном канале 10–11, в верхнечелюстно-подглазничном – 12, в нижнечелюстно-пекторальном – 21–23, в канале туловищной боковой линии – 30–35 (последние один–три невромаста на хвостовом плавнике); кроме того, два–три верхнепредкрышечных и три–четыре нижнепредкрышечных невромаста и два невромаста в нижнеподбородочной комиссуре.
Иллициальная ямка (рис. 2а) очень мелкая, плоская, светлая, сужена кпереди и сильно вытянута продольно (наибольшая ширина в 2.4–4.0 раза меньше длины), так что в некоторых случаях её ширина едва превышает толщину иллициума. Иллициум светлый, его длина колеблется от двух третей до почти всей длины иллициальной ямки. Эска мягкая, узко-коническая, спереди, по бокам и на вершине покрыта тонкими, довольно длинными тёмными волосками. По краям иллициальной ямки нет кожистых волосовидных придатков. Рот верхний, нижняя челюсть выступает, конец верхней челюсти угловато-закруглённый, не достигает уровня переднего края глаза. Зубы в челюстях мелкие, острые, конические, на praemaxillare – полоской, более широкой в симфизной части, на dentale – в три ряда; поперечный ряд мелких острых зубов на vomer и такие же мелкие зубы на palatinum. Кожа сплошь густо покрыта бугорками, оканчивающимися шипиками, гораздо более длинными и грубыми на верхней и боковых поверхностях головы и туловища, хвостовом стебле и вентральной поверхности нижней челюсти, переходящими на лучи С; лучи D, лучи и перепонка P в мелких шипиках; все шипики простые. Чешуи вокруг невромастов с тремя шипами на приподнятом выступе у более крупных рыб, у рыб SL < 40 мм – с единственным тонким шипиком. На нижней челюсти и по ходу сейсмосенсорных каналов сидят мелкие волосовидные кожные придатки. Жаберное отверстие поровидное, со складчатыми краями, расположено заметно позади основания Р. Туловищный канал LL резко изгибается книзу за жаберным отверстием. Основание Р длинное, образует “локоть”, скрытый в коже. V нередко отсутствуют, когда имеются – очень малы, сидят в пазухах, образованных кожей брюха, и только у 1 экз. хорошо развиты с обеих сторон тела (менее 10% SL). Часто (в том числе у наибольших из имеющихся экземпляров) с одной стороны V развит, а с другой сильно редуцирован (рис. 2б). Третья колючка D от очень короткой до хорошо выраженной. Перед началом мягкой части D из мышц выступают два невральных отростка позвонков. Начало мягкой части D впереди вертикали жаберного отверстия. Срединные лучи С самые длинные.
Рис. 2.
Chaunax breviradius (а – иллициальная ямка и эска, б – асимметричное развитие V на разных сторонах тела) и C. abei (в – иллициальная ямка и эска). Масштаб: 2 мм.
Некоторые измерения. В % SL ( n = 7, SL 47–63 мм11): расстояние от вершины головы до центра жаберного отверстия 51.7–64.5 (71.2?), ширина головы между наружными краями sphenotica 19.0–19.1 (20.9, 21.3), длина и минимальная высота хвостового стебля 11.1–19.1 и 7.7–8.5, расстояние от вершины головы до основания верхнего луча Р 43.1–54.8 (61.5), предорсальное (до 1-го соединенного луча D), преанальное и превентральное (если есть V) расстояния соответственно (43.1?, 46.0) 51.9–54.8 (59.6?), (66.7?) 69.0–73.1 и 35.5–39.7 (40.4), длина грудного и хвостового плавников соответственно 17.7–23.4 и 37.4–30.8 (34.0), расстояние от симфиза praemaxillaria до переднего края глаза 7.9–10.6, горизонтальный диаметр глаза 7.9–10.6, ширина межглазничного промежутка 6.9–8.5, длина верхней челюсти (16.1) 19.0–21.3, длина иллициума (5.2) 7.1–8.9, длина и максимальная ширина иллициальной ямки соответственно 7.8–9.7 и 2.0–4.0.
Окраска фиксированных рыб светлая, желтоватая или кремовая (при жизни – красновато-розовая); низ головы, грудь и брюхо белые. На верху и боках головы и туловища и на лучах Р многочисленные тёмные (при жизни – зеленоватые) пятна, без светлого ободка вокруг них; поверхность между пятнами с мелкими точечными меланофорами, как правило, не различимыми невооруженным глазом. Пятнистость мало варьирует у изученных экземпляров, за исключением самых мелких рыб (SL 24–34 мм), у которых пятна на голове и теле более размыты, отдельные точечные меланофоры между пятнами выражены гораздо сильнее и формируют подобие сетчатого рисунка. Лучи D и С с неясными тёмными пестринами. Ротожаберная полость светлая. Подкожная пигментация представлена разрозненными скоплениями меланофоров на затылке и на вентролатеральной поверхности между основанием Р и началом А; единичные меланофоры присутствуют также под и за глазом. В основании С у самых мелких рыб один–два точечных меланофора, с ростом рыб редуцирующихся.
Сравнительные замечания. В моих сборах из зал. Нячанг и Ванфонг хаунаксы этого вида представлены исключительно молодью, взрослых рыб собрать не удалось. Крупные экземпляры хаунаксов, которые встречались в уловах близнецовых тралов мористее зал. Нячанг и Ванфонг, разгружавшихся в порту в бухте Няфу (Cang Ca Vinh Luong), относились к другому виду – C. fimbriatus Hilgendorf, 1879. Изученные рыбы хорошо соответствуют первоописанию C. breviradius, однако длина третьей свободной колючки D, слабое развитие которой считалось диагностическим признаком данного вида (Le Danois, 1978), у изученных рыб сильно варьирует. C. breviradius очень сходен с C. abei, отличаясь от последнего деталями строения иллициальной ямки и эски: у C. abei ямка более широкая (ширина 1.5–2.4 раза в длине), округло-овальная и более углублённая, тогда как у C. breviradius она совсем мелкая и более удлинённая (рис. 2а, 2в; см. также: Le Danois, 1978. Figs. 2, 3). Эска у C. abei с ярко-чёрными, довольно широкими кожистыми придатками в виде оборок, тогда как у C. breviradius они тонкие, волосовидные и не столь интенсивно пигментированы. Чётким отличием C. breviradius от других видов является гораздо меньшее число невромастов в нижнечелюстно-пекторальном канале (21–23 против 25–27 у C. abei).
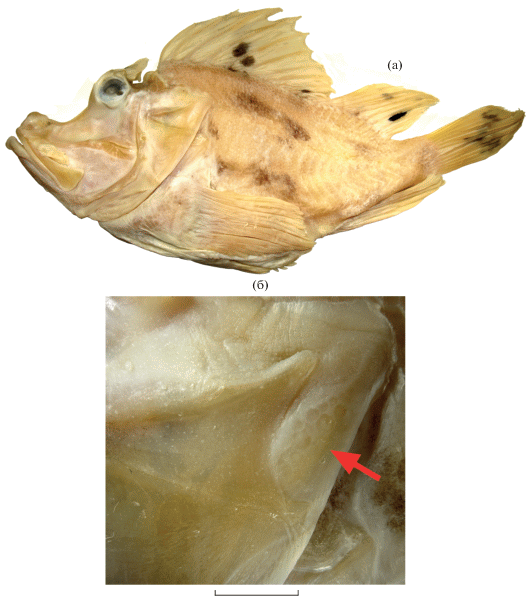
Рис. 3.
Rhinopias eschmeyeri SL 145 мм: а – общий вид, б – чешуя на жаберной крышке (→). Масштаб: 5 мм.
C. breviradius до сих пор был известен только по типовой серии, собранной в водах Филиппин на глубинах 180–510 м (Le Danois, 1978; Ho et al., 2015). В моих сборах из Вьетнама молодь этого вида нередко встречалась при тралениях на глубинах от 38 до 150 м. Нельзя сказать, что глубина обитания этого вида увеличивается с ростом, так как три паратипа этого вида SL 28.6–40.0 мм были добыты на глубине 415–510 м, а наибольший известный экземпляр (голотип) SL 110 мм – на глубине 182–200 м. Вероятно, как и другие глубоководные рыбы, отмечавшиеся в мористой части зал. Нячанг и Ванфонг на аномально малых глубинах (Prokofiev, 2008), молодь хаунаксов поднимается на такие глубины “подсасыванием” глубинных вод по узким каньонам, многочисленным в районе работ.
(рис. 3)
Материал. ИО РАН б/№, 1 экз. SL 145 мм (рис. 3а), Вьетнам, зал. Фантьет (Cho Ca Con Cha), 28–29.04.2009 г., коллектор А.М. Прокофьев.
Описание. D XII + 10, A III + 6, P 16, V I + 5, C 7 + 6 (ветвистых 6 + 5); sp. br 6 + 1 + 13, LL 22, squ ~60. Голова и тело высокие, сильно сжатые с боков; глазницы сильно приподняты над дорсальным контуром головы, дорсальный контур с глубокими выемками перед и позади орбиты. Шипы на голове в виде тупых бугорков с округлыми вершинами, надглазничный гребень оканчивается плоским треугольным шипом, нухальные гребни и шипы валикообразные, преоперкулярные шипы в виде едва заметных бугорков, числом пять. В челюстях мелкие однородные зубы полосками, такие же зубы в узком V-образном пятне на сошнике, нёбные кости без зубов. Все жаберные тычинки бугорковидные, ложножабра хорошо развита, состоит из 20 элементов, нет щели за 4-й жаберной дугой. По крупной плоской мочке расположено в начальной части надглазничного гребня и у заднего (третьего) лакримального шипа; мочки на голове, теле и плавниках отсутствуют, за исключением очень мелких и немногочисленных нитевидных на колючих лучах D и единичных длинных нитевидных у отдельных пор LL. D с глубокой выемкой у 11-й колючки, без сильно удлинённых колючих лучей, его четвертая колючка едва короче третьей, 10-я и 11-я заметно изогнуты. Последний луч D и А не соединены перепонкой с хвостовым стеблем, самый внутренний луч V соединён перепонкой с телом. Все мягкие лучи D, A и V и 2-й–7-й лучи Р ветвистые. Чешуя циклоидная, её дистальный край волнистый, особенно у более передних чешуй, выступы которых сужены наподобие ктений. Грудь и брюхо, кроме вершины истмуса, покрыты чешуей. Голова без чешуи, за исключением небольшого участка чешуй у заднего края жаберного отверстия между вершинами оперкулярных шипов (рис. 3б).
Некоторые измерения. В % SL: длина головы 46.2, максимальная и минимальная высота тела соответственно 48.3 и 11.0, длина хвостового стебля 12.4, длина рыла 18.6, горизонтальный диаметр глаза 6.9, ширина межглазничного промежутка 5.9, длина верхней челюсти 20.7; длина Р, колючего луча V и С соответственно 34.5, 13.8 и 29.0; длина наибольшего (третьего) ветвистого луча V 22.8, длина колючек D с 1-й по 4-ю соответственно 16.2, 22.1, 26.9 и 26.2; длина колючек А с 1-й по 3-ю соответственно 9.7, 15.9 и 17.2; длина наибольшего ветвистого луча D и А соответственно 21.4 и 23.5. В % длины головы: длина рыла 40.3, горизонтальный диаметр глаза 14.9, ширина межглазничного промежутка 12.7.
Окраска фиксированной рыбы светлая (при жизни – жёлто-розовая), с черновато-серыми, неопределённой формы пятнами на боках тела; передний край надглазничных мочек с участком чёрного пигмента в основании. На межлучевой перепонке между 4-й–5-й, 5-й–6-й и 9-й–10-й колючими и 7-м–8-м мягкими лучами D по округлому чёрному пятну, между 2-м–3-м, 3-м–4-м и 5-м–6-м мягкими лучами D присутствуют размытые участки сероватого пигмента, посередине C проходит нерезкая зигзагообразная чёрно-серая поперечная полоса.
Замечания. Вид был известен по 9 экз. из вод Маскаренских и Сейшельских о-вов, Японии, Вьетнама, Северной и Западной Австралии, во Вьетнаме был найден 1 экз. в зал. Нячанг (сборы П. Фурмануа (P. Fourmanoir) (Motomura, Johnson, 2006)). Описанный экземпляр происходит из зал. Фантьет, его нахождение свидетельствует о широком распространении вида в прибрежных водах Южного Вьетнама. Признаки нового экземпляра хорошо согласуются с таковыми у ранее описанных рыб, за исключением наличия у него чешуй у заднего края жаберной крышки между вершинами оперкулярных шипов (рис. 3б). Ранее диагностическим признаком R. eschmeyeri считалось отсутствие чешуи в этой области (Motomura, Johnson, 2006).
(рис. 4a)
Рис. 4.
Общий вид Plectranthias longimanus SL 22 мм (а), P. japonicus SL 100 мм (б) и Pentapodus setosus SL 165 мм (в).

Материал. ИО РАН б/№, 1 экз. SL 22 мм, Вьетнам, зал. Нячанг, Хон-Нок, глубина 25 м, на горгонариях, 28.11.2005 г.
Описание. ID X, IID i + 13, A III + 7, P 13, V I + + 5, C I + 8 + 7 + I; sp. br (развитые) 4 + 1 + 9 (длинные и тонкие); squ 26. Maxillare расширено сзади, достигает вертикали заднего края глаза. В челюстях мелкие щетинковидные зубы полосками, на симфизе praemaxillare пара увеличенных клыковидных зубов; на сошнике и нёбных костях мелкие зубы. Язык сужается кпереди, без зубов. Передняя ноздря трубковидная. Чешуя на верху головы доходит до уровня задних краёв глаз; щёки и жаберная крышка покрыты чешуёй; на щеке три ряда чешуй; задняя пластинка maxillare без чешуи. Praeoperculum с грубо зазубренными угловой частью и дорсальной ветвью и двумя направленными вперёд шипами на вентральной ветви; три шипа на operculum; три сильных шипа на interoperculum и два на suboperculum. Спинные плавники полностью разобщены; 1-я колючка ID короткая, 2-я незначительно короче 3-й, 3-я и 4-я самые длинные, далее назад длина колючих лучей постепенно уменьшается, последние две колючки ID очень малы; 1-й (неветвистый) луч IID гибкий и членистый на вершине. Вторая колючка А значительно толще и длиннее третьей. Р длинные, значительно заходят за начало А; все лучи Р неветвистые, членистые; нет кожной перемычки между самым верхним лучом Р и телом. Дистальный край С закруглён. Боковая линия неполная, в передней части круто изгибается кверху и идет вблизи дорсального края тела, оканчиваясь немного позади уровня начала IID, содержит 15 прободённых чешуй.
Некоторые измерения. В % SL: длина головы 40.9, максимальная и минимальная высота тела 34.1 и 15.9; длина P, V и С соответственно 36.4, 22.7 и 22.7.
В % длины головы: длина рыла 22.2, горизонтальный диаметр глаза 22.2, ширина межглазничного промежутка 5.6, длина верхней челюсти 44.4.
Основной фон окраски светлый, с вертикальными тёмными полосками под глазом, косыми или горизонтальными полосками и пятнами на рыле, на верху головы и в её заглазничной части. Около шести нечётких поперечных полос на теле, частично распадающихся на отдельные пятна (при жизни были зеленоватыми); под концом основания IID тёмное пятнышко, около четырёх тёмных пятен вдоль основания А и два супротивных тёмных пятна в основании лучей С. У живой рыбы радужина глаз была с радиальными красноватыми полосками, исчезнувшими после фиксации. Все плавники не окрашены. Ротожаберная полость светлая.
Замечания. Вид широко распространён в Индо-Вест-Пацифике от побережья Восточной и Южной Африки до Новой Каледонии, Микронезии и Южной Японии (Randall, 1980; Heemstra, Randall, 2009). В Южно-Китайском море вид ранее был отмечен у берегов Тайваня (Chen, Shao, 2002). Для фауны Вьетнама указывается впервые.
Род Plectranthias Bleeker, 1873 ранее вообще не был указан в списке рыб Вьетнама (Nguyen et al., 1995), однако по результатам моих сборов в зал. Нячанг и Ванфонг здесь встречаются, по меньшей мере, два вида этого рода. Помимо P. longimanus, характерного для коралловых рифов, в уловах близнецовых тралов мористее зал. Нячанг и Ванфонг отмечен P. japonicus (Steindachner, 1883) (рис. 4б), связанный с мягкими грунтами. Судя по траловым уловам, этот вид держится группами или небольшими стайками и обычно попадается как прилов при промысле ящероголовых рыб (Synodidae) и нитепёров (Nemipteridae).
(рис. 4в)
Материал. ИО РАН б/№, 3 экз. SL 96–165 мм, Вьетнам, зал. Нячанг, 12°08′000–12°05′585 с.ш., 109°13′600–109°12′735 в.д., глубина 7–19 м, трал № 1 (10.12.2005 г.) и № 3 (13.12.2005 г.), коллектор А.М. Прокофьев.
Описание. D X + 9½, A III + 7½, P 16 (ii + 14), V I + 5; LL 46–48, между LL и началом D 3½ чешуи; sp. br 4 + 1 + 4 (короткие, утолщённые на концах). Чешуя на верху головы почти доходит до уровня задней пары ноздрей; рыло, lacrimale и свободный край praeoperculum голые. На щеке пять рядов чешуй. Рыло приострённое. Челюсти равной длины, рот мал, конечный, его задний край не достигает вертикали переднего края глаза. На симфизе praemaxillaria с каждой стороны по два клыковидных зуба, задняя пара таких зубов заметно крупнее передней и соизмерима с небольшим, загнутым назад клыком у симфиза dentalia (по одному с каждой стороны); боковые зубы мелкие, на нижней челюсти несколько укрупняются кзади; на сошнике группа мелких зубов. Суборбитального шипа нет. Свободный край lacrimale и praeoperculum волнистый; по краю praeoperculum многочисленные тонкие ребрышки, не оканчивающиеся шипами. D единый, без отчётливой выемки; первая колючка A мала, а вторая явно короче третьей, но все три колючки примерно одинаковой толщины. Имеются хорошо развитые грудная и брюшная аксиллярные чешуи. C глубоко вырезан, его верхняя лопасть вытянута в очень длинный и тонкий филамент, нижняя обычного строения.
Некоторые измерения. В % SL: длина головы 24.0–26.1, максимальная и минимальная высота тела соответственно 28.1–28.5 и 9.1–10.4. В % длины головы: длина рыла 32.6–43.5, горизонтальный диаметр глаза 23.3–30.4, ширина межглазничного промежутка 37.2–43.5.
Окраска фиксированных рыб зелёно-пурпурно-серая (при жизни иридирующая пурпурно-тёмно-зелёная), более тёмная у спины, книзу светлеющая; на боках салатово-зелёная иридирующая полоса; нечёткая тёмная полоса на жаберной крышке и вдоль боков тела, а у экз. SL 96 мм на жаберной крышке еще небольшое тёмное пятнышко; в основании С крупное треугольное тёмное пятно. На рыле две тёмные полосы: верхняя от вершины рыла до переднего края глаза, а нижняя от переднего края глаза к верхнему краю переднего конца верхней челюсти. Все плавники не окрашены; хвостовая нить тёмная. Ротожаберная полость светлая.
Замечания. Вид ранее был известен из Сиамского залива, внутренних морей Индонезии и от Филиппин (Russell, 1990). Для фауны Вьетнама отмечается впервые. До сих пор в списках ихтиофауны Вьетнама представители данного рода указаны не были (Nguyen et al., 1995; Russell, 1990), хотя P. setosus является объектом местного промысла в зал. Нячанг, где этот вид отмечен в южной части залива (от мыса Муй-Дам-Ба и южнее) вблизи участков каменистых и коралловых рифов; здесь он попадался в тралы постоянно, но в небольшом количестве в зимний сезон.
(рис. 5, 6)
Рис. 5.
Torquigener gloerfelti (а–в – SL 121 мм; г, д – SL 57 мм), вид: а, г –сверху; б, д –сбоку; в – снизу.

Рис. 6.
Детали строения Torquigener gloerfelti: а – папиллы вдоль LL в задней части тела, б – текстура кожи дорсальной поверхности хвостового стебля, в – перибранхиальные папиллы. Масштаб: 5 мм.

Материал. Везде ИО РАН б/№, Вьетнам, зал. Нячанг, коллектор А.М. Прокофьев: 1 экз. SL 73 мм, 12°14′723–12°14′962 с.ш., 109°19′095–109°16′939 в.д., 7–10 м, трал № 2, 17.06.2006 г.; 1 экз. SL 70 мм, 12°02′959–12°05′056 с.ш., 109°13′183–109°13′095 в.д., 10–18 м, трал № 1, 19.06.2006 г.; 1 экз. SL 57 мм, 12°02′480–12°06′253 с.ш., 109°12′583–109°13′067 в.д., 10–18 м, тралы № 1 и 2, 23.06.2006 г.; 1 экз. SL 46 мм, 12°15′960–12°18′518 с.ш., 109°16′501–109°16′503 в.д., 15–18.6 м, трал № 3, 27.04.2007 г.; 1 экз. SL 121 мм, 12°06′983–12°04′753 с.ш., 109°1′027–109°13′104 в.д., 18–18.8 м, трал № 3, 04.05.2007 г.
Описание22. D ii + 7, A ii + 5, P ii + 13 (ii + 13–14) (самый верхний луч очень короткий), C I + 3 + + 3 + I. Общий вид взрослой и молодой рыбы показан на рис. 5. Тело удлинённое, невысокое, в поперечном сечении закруглено дорсально и уплощено вентрально, плавно понижается к основанию С; хвостовой стебель уплощён дорсально и вентрально. Вентролатеральная складка хорошо развита от подбородка до основания С. Глаз приращён к коже только вдоль дорсального края, его нижний край расположен ближе к нижнему краю носового органа, чем к горизонтали вершины рыла. Носовой орган в виде небольшого мешка, открывающегося спереди и сзади, полностью занимающего углублённую округлую ямку, вдвое ближе к переднему краю глаза, чем к вершине рыла. Губы папильчатые; вершина подбородочного выступа в тонких продольных папильчатых складках, замещающихся хаотично сидящими папиллами на его верхней поверхности. Кожа верха рыла и межглазничного промежутка в тонких продольных бороздках, отграничивающих продольные складки, которые становятся более грубыми и извилистыми на дорсальной поверхности туловища и в верхней половине боков между вертикалями начал Р и А; бока туловища и хвостового стебля ниже этих бороздок несут нерезкие узкие поперечные складочки. Верх и бока туловища позади глаз и до уровня начала D несут многочисленные плоские33 ланцетовидные, закруглённые на вершинах папиллы, внутри содержащие шипик. Эти папиллы очень маленькие и редкие на боках головы под глазом и на дорсальной поверхности головы между ноздрями и в межглазье; кпереди от ноздрей совершенно отсутствуют. В поперечном ряду между верхними боковыми линиями соседних сторон, сразу за комиссурой, их соединяющей, 8 (8–10) шипиков. Позади вертикали начала D обособленных папилл нет, но вдоль верхней боковой линии прослеживаются удлинённые, хорошо отграниченные кожные валики, возможно, представляющие собой видоизменённые вросшие папиллы (рис. 6а). Продольные кожные складки на дорсальной поверхности хвостового стебля не доходят до основания С (примерно на четверть длины постдорсального расстояния: рис. 6б). На вентральной стороне шипики, заключённые в папиллы, сзади немного не доходят до ануса, спереди начинаются от основания подбородочного выступа. В поперечном ряду между основаниями Р 17 (18–19) шипиков. По переднему краю жаберного отверстия 8/9 (7–10, обычно 8) плоских, часто изогнутых папилл, содержащих жёсткий шипик (рис. 6в). У рыбы SL 121 мм перибранхиальные папиллы развиты только в нижней половине переднего края жаберного отверстия, у более мелких рыб этот признак варьирует, и папиллы могут присутствовать на всем протяжении переднего края (SL 46 и 70 мм). Число, размер и расположение перибранхиальных папилл могут варьировать на разных сторонах одной и той же рыбы. Начало А расположено на вертикали середины основания D, 1–2-й ветвистые лучи в этих плавниках наибольшие; вершина D оттянута и приострена, вершина А закруглена. Р с практически прямым дистальным краем и закруглённым задненижним углом, его 1–2-й ветвистые лучи самые длинные. Дистальный край С практически усечённый.
Рис. 7.
Torquigener brevipinnis (а, в, г – SL 90 мм; б – SL 70 мм), вид: а – сверху, б, в – сбоку (в – раздувшийся экземпляр); г – перибранхиальные папиллы. Масштаб: 5 мм.

Некоторые измерения. В % SL: длина головы 33.1 (32.9–37.0), длина рыла 12.4 (12.3–13.7), горизонтальный диаметр глаза 8.7 (9.7–10.9), ширина межглазничного промежутка (по мягким тканям) 15.3 (12.9–15.8), длина жаберной щели 9.1 (9.4–10.5), ширина рта 9.1 (9.7–10.9), максимальная и минимальная высота тела соответственно 22.3 (21.7–28.1) и 7.4 (6.5–7.9), максимальная ширина тела 22.3 (29.0–32.6), длина и минимальная ширина хвостового стебля соответственно 27.3 (24.3–26.3) и 5.0 (4.3–5.4), предорсальное и преанальное расстояние соответственно 63.6 (64.3–67.4) и 66.1 (67.1–70.7), длина грудного и хвостового плавников соответственно 15.7 (15.2–17.5) и 20.7 (22.8–25.0), высота спинного и анального плавника соответственно 19.8 (17.1–17.5) и 14.9 (14.0–14.1).
Окраска фиксированных рыб. Верх тёмный, коричнево-серый, низ светлый, между вентролатеральными складками чисто-белый. На дорсальной поверхности развит неяркий, умеренно густой чёрно-коричневый крап; бока головы и туловища в немногочисленных простых округлых (под глазом часто овальных, изредка V-образных) пятнах, наиболее крупных на боках тела в промежутке между концом прижатого к телу Р и основанием С (диаметр самых крупных пятен равен половине горизонтального диаметра глаза). В основании С узкая поперечная тёмная полоска. С дистально зачернён; в остальном плавники не окрашены. Передний край подбородочного выступа и губы белые, вершины шипиконосных папилл выделяются светлыми кончиками.
Онтогенетическая изменчивость. У молоди SL 46–73 мм подбородочный выступ слабо намечен, тогда как у экз. SL 121 мм он сильно развит, лопастевидной формы (рис. 5б, 5д). Перибранхиальные шипики у молоди развиты на большем протяжении переднего края жаберной щели, их концы могут далеко выступать из папиллы, вершина D не столь оттянута.
Окраска с ростом изменяется мало, у мелких рыб более отчётливый мелкий крап на дорсальной и латеральной стороне тела, а чёрная пигментация дистального края С занимает несколько бóльшую площадь.
Замечания. Вид был известен по 4 экз. типовой серии, собранным у южного побережья о. Ява и у о-вов Бали и Сумба (Индонезия) (Hardy, 1984). Для фауны Вьетнама и Южно-Китайского моря отмечается впервые. Изученные экземпляры хорошо согласуются с первоописанием, более широкий диапазон колебания некоторых счётных и пластических признаков может быть объяснён индивидуальной изменчивостью и тем, что типовая серия представлена в целом более крупными особями (SL 94–154 мм).
Помимо T. gloerfelti в сборах из зал. Нячанг представлен другой вид этого рода – T. brevipinnis (Regan, 1902) (рис. 7). Изученные мною 4 экз. SL 60–90 мм характеризуются следующими особенностями: под глазом три или четыре (тогда задние две соединяются друг с другом по направлению к глазу) узкие субвертикальные светлые полоски и инвертированная U-образная светлая полоска между глазом и жаберным отверстием; на боках сплошная тёмная (при жизни жёлтая) продольная полоса на границе тёмной и светлой окраски соответственно дорсальной и вентральной поверхностей, под ней проходит ещё одна подобная, но более размытая и нечёткая полоса (рис. 7а–7в); 5–9 перибранхиальных папилл, расположенных вдоль всего переднего края жаберного отверстия (рис. 7г), 20–22 шипика в поперечном ряду между основаниями Р; горизонтальный диаметр глаза и длина хвостового стебля составляют соответственно 9.2–10.0 и 21.5–23.7% SL (10.0–10.8 и 4.2–4.6 раза). Эти признаки полностью соответствуют характеристике данного вида, приводимой Харди (Hardy, 1984). Однако необходимо отметить, что в литературе встречаются разночтения при описании признаков, по которым различают близкие виды T. brevipinnis и T. hypselogeneion (Bleeker, 1852). По сведениям Ямады (Yamada, 2002), меньшее число полос на щеке (три–четыре против пяти) и сплошная (против разбитой на отдельные секции) латеральная полоса отличают T. hypselogeneion от T. brevipinnis. Однако, по данным Харди (Hardy, 1983, 1984), эти виды по указанным признакам не различимы, а характеристики, даваемые Ямадой T. brevipinnis, на самом деле соответствуют признакам другого вида этого комплекса – T. flavimaculosus Hardy et Randall, 1983, известного из западной части Индийского океана (Hardy, Randall, 1983). Я принимаю взгляды Харди, так как они основаны на переизучении типового материала. Возможно, в водах Японии встречается неописанный вид комплекса “hypselogeneion”, отождествляемый Ямадой с T. brevipinnis. Для фауны Вьетнама T. brevipinnis до сих пор не был указан. Повсеместно в водах Вьетнама ранее был отмечен T. hypselogeneion (Nguyen, 1999), но, вероятно, вьетнамские авторы не отличали эти два вида. Оба вида могут встречаться совместно, но в моих сборах из зал. Нячанг, Ванфонг и Фантьет экземпляры, подходящие под переописание T. hypselogeneion у Харди (Hardy, 1983), не найдены.
Список литературы
Caruso J.H. 1989. Systematics and distribution of Atlantic chaunacid anglerfishes (Pisces: Lophiiformes) // Copeia. № 1. P. 153–165.
Chen J.-P., Shao K.-T. 2002. Plectranthias sheni, a new species and P. kamii, a new record of anthiine fishes (Perciformes: Serranidae) from Taiwan // Zool. Stud. V. 41. № 1. P. 63–68.
Eschmeyer W.N., Hirosaki Y., Abe T. 1973. Two new species of the scorpionfish genus Rhinopias, with comments on related genera and species // Proc. Calif. Acad. Sci. Ser. 4. V. 39. № 16. P. 285–310.
Fricke R., Eschmeyer W.N., van der Laan R. 2019. Catalog of Fishes: Genera, Species, References. Available. (http:// researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Version 06/2019)
Grande T. 1999. Revision of the genus Gonorynchus Scopoli, 1777 (Teleostei: Ostariophysi) // Copeia. № 2. P. 453–469.
Hardy G.S. 1983. The status of Torquigener hypselogeneion (Bleeker) (Tetraodontiformes: Tetraodontidae) and some related species, including a new species from Hawaii // Pac. Sci. V. 37. № 1. P. 65–74.
Hardy G.S. 1984. Redescription of the pufferfish Torquigener brevipinnis (Regan) (Tetraodontiformes: Tetraodontidae), with description of a new species of Torquigener from Indonesia // Ibid. V. 38. № 2. P. 127–133.
Hardy G.S., Randall J.E. 1983. Description of a new species of pufferfish (Tetraodontiformes: Tetraodontidae) from the Red Sea and adjacent waters // Israel J. Zool. V. 32. P. 13–20.
Heemstra P.C., Randall J.E. 2009. A review of the anthiine fish genus Plectranthias (Perciformes: Serranidae) of the Western Indian Ocean, with description of a new species, and a key to the species // Smithiana Bull. № 10. P. 3–17.
Ho H.-C., Kawai T., Satria F. 2015. Species of the anglerfish genus Chaunax from Indonesia, with descriptions of two new species (Lophiiformes: Chaunacidae) // Raffles Bull. Zool. V. 63. P. 301–308.
Hubbs C.L., Lagler K.F. 1958. Fishes of the Great Lakes region // Bull. Cranbrook Inst. Sci.№ 26. 213 p.
Kanazawa R.H. 1958. A revision of the eels of the genus Conger with descriptions of four new species // Proc. US. Natl. Mus. V. 108. № 3400. P. 219–267.
Le Danois Y. 1978. Description de deux nouvelles especes de Chaunacidae (Pisces pediculati) // Cybium. Ser. 3. № 4. P. 87–93.
Motomura H., Johnson J.W. 2006. Validity of the poorly known scorpionfish, Rhinopias eschmeyeri, with redescriptions of R. frondosa and R. aphanes (Scorpaeniformes: Scorpaenidae) // Copeia. № 3. P. 500– 515.
Nakabo T. 2002. Introduction to Ichthyology // Fishes of Japan with pictorial keys to the species. V. 1 / Ed. Nakabo T. Tokyo: Tokai Univ. Press. P. xxi–xlii.
Nguyen H.P. 1999. Checklist of marine fishes in Vietnam. V. 5. Hochiminh: Agricult. Publ. House, 305 p.
Nguyen H.P., Nguyen N.T. 1994. Checklist of marine fishes in Vietnam. V. 2. Hanoi: Publ. House “Science and Technics”, 269 p.
Nguyen H.P., Tran H.L. 1994. Checklist of marine fishes in Vietnam. V. 1. Hanoi: Publ. House “Science and Technics”, 115 p.
Nguyen H.P., Le T.P., Nguyen N.T. et al. 1995. Checklist of marine fishes in Vietnam. V. 3. Hanoi: Publ. House “Science and Technics”, 606 p.
Prokofiev A.M. 2008. A new species of Platygobiopsis from Vietnam (Teleostei: Perciformes: Gobiidae) // J. Ichthyol. V. 48. № 10. P. 853–859.
Randall J.E. 1980. Revision of the fish genus Plectranthias (Serranidae: Anthiinae) with descriptions of 13 new species // Micronesica. V. 16. № 1. P. 101–187.
Randall J.E., Liem K.K.P. 2000. A checklist of the fishes of the South China Sea // Raffles Bull. Zool. Suppl. № 8. P. 569–667.
Russell B.C. 1990. FAO species catalogue. V. 12. Nemipterid fishes of the World. Rome: FAO, 148 p., VIII pls.
Shao K.-T., Ho H.-C., Lin P.-L. et al. 2008. A checklist of the fishes of southern Taiwan, northern South China Sea // Raffles Bull. Zool. Suppl. 19. P. 233–271.
Smith D.G., Ho H.-C. 2018. Review of the congrid eel genus Conger (Anguilliformes: Congridae) in Taiwan // Zootaxa. V. 4454. № 1. P. 168–185.
Yamada U. 2002. 351. Tetraodontidae puffers // Fishes of Japan with pictorial keys to the species. V. 2 / Ed. Nakabo T. Tokyo: Tokai Univ. Press. P. 1418–1431.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии