

Вопросы ихтиологии, 2020, T. 60, № 2, стр. 137-164
Морфофункциональные особенности висцерального аппарата рубиново-пепельной рыбы-попугая Scarus rubroviolaceus (Scaridae)
Е. С. Громова 1, В. В. Махотин 1, *
1 Московский государственный университет
Москва, Россия
* E-mail: vmakhotin@mail.ru
Поступила в редакцию 31.01.2019
После доработки 25.03.2019
Принята к публикации 04.04.2019
Аннотация
Исследованы детали строения некоторых мускулов, соединительнотканных образований и костных структур висцерального аппарата рубиново-пепельной рыбы-попугая Scarus rubroviolaceus. В нижней челюсти обнаружен замочный механизм, который ограничивает степень приведения anguloarticulare относительно суспензориума. Обсуждается воздействие, оказываемое со стороны многочисленных подразделений m. adductor mandibulae, на элементы передних челюстей в процессе закрывания рта рыбы.
Исследование анатомии висцерального аппарата видов костистых рыб (Teleostei), потребляющих небольшой спектр жертв, позволяет раскрыть морфологическую основу их уникальных способов добывания корма. Отряд окунеобразных (Perciformes) открывает значительные возможности для изучения структрурных адаптаций челюстного аппарата его специализированных представителей, а большое число видов Perciformes позволяет провести сравнение этих приспособлений с подобными образованиями у ряда генерализованных форм данного отряда (Nelson, 2006).
Настоящая работа является продолжением нашего исследования (Громова, Махотин, 2019), посвящённого строению и функционированию спланхнокраниума рыб-попугаев (Scaridae), в котором дана подробная характеристика представителей этого семейства, в том числе особенностей их питания. Робустные передние челюсти являются характерным признаком этих рыб. Прочное крепление контрлатеральных praemaxillare и dentale друг к другу создаёт монолитную конструкцию внешне единой верхней и нижней челюстей, которые напоминают клюв попугая. В сравнении с другими видами коралловых рыб попугаевые обладают одной из самых высоких скоростей питания (Bonaldo et al., 2014), что, по-видимому, служит компенсацией их не слишком питательному рациону. По характеру укусов поверхности рифа, на которой располагаются пищевые объекты попугаевых (бентосные водоросли или коралловые полипы) (Bruggemann et al., 1996; Ong, Holland, 2010), среди этих рыб выделяют соскребателей и копателей (Bellwood, Choat, 1990; Bonaldo et al., 2011, 2014). Соскребатели выполняют скобление субстрата при помощи более частых и высокоамплитудных, но менее силовых укусов в сравнении с более глубокими, мощными и редкими укусами видов-копателей, которые в процессе питания захватывают в ротовую полость значительную долю осадочной породы рифа (CaCO3) (Bellwood, 1996). В связи с использованием разных способов добычи пищи представители этих двух групп попугаевых оставляют на поверхности, с которой собирают корм, царапины разной формы и размера (Francini-Filho et al., 2008; Alwany et al., 2009; Bonaldo, Bellwood, 2009). Эти следы отражают разную степень вовлечения в исполнение укуса верхней и нижней челюстей. Кроме того, у соскребателей и копателей были обнаружены определённые различия в строении гипертрофированного m. adductor mandibulae, костей передних челюстей и суспензориума, что послужило морфологическим основанием для функционального разделения данных групп по способу питания (Bellwood, 1985; Bellwood, Choat, 1990). Вместе с тем некоторые виды попугаевых из группы соскребателей, в том числе рубиново-пепельная рыба-попугай Scarus rubroviolaceus, в процессе кормодобывания могут использовать способ сбора пищи копателей (Bellwood, Choat, 1990).
В литературе можно найти небольшое количество исследований скелета и мышечной системы головы определённых видов попугаевых (Randall, Nelson, 1979; Tedman, 1980a, 1980b; Bellwood, 1985, 1994; Clements, Bellwood, 1988; Bellwood, Choat, 1990; Alfaro, Westneat, 1999; Konow et al., 2008; Mikami, 2013; Nanami, 2016; Melgarejo-Damian et al., 2018). Однако эти работы, на наш взгляд, нуждаются в дополнении и уточнении содержащихся в них морфологических данных, благодаря чему открывается широкий доступ для анализа функциональных возможностей структур висцерального аппарата представителей Scaridae.
Цель работы – подробно исследовать строение нейрокраниума, нижней челюсти, а также некоторых мышц и соединительнотканных структур аппарата питания рубиново-пепельной рыбы-попугая для анализа функционального значения сочетания различных анатомических признаков в процессе кормодобывания рыбы. Описаны новые признаки строения составляющих спланхнокраниума, которые могут служить морфологической основой для использования данным видом-соскребателем стратегии сбора корма с поверхности субстрата по модели копателей.
МАТЕРИАЛ И МЕТОДИКА
В работе использовали половозрелых особей рубиново-пепельной рыбы-попугая на стадии онтогенеза IP (Bruggemann et al., 1996; Howard et al., 2013): шесть самок стандартной длиной (SL) 23−42 см и двух самцов 25 и 30 см. Для исследования мускулов и соединительнотканных образований аппарата питания изготовили три спиртовых и два сухих препарата голов по традиционной методике (Ромейс, 1953). Для анализа движений спланхнокраниума рыбы использовали три свежих, очищенных до скелета препарата голов с сохранением связок и мускулов. Препараты исследовали при помощи стереомикроскопа МБС-1. Рисунки выполнены на основе цифровых цветных фотографий препаратов (камера Panasonic DMC-FZ8). Фотографии обрабатывали в программе Adobe Photoshop CS2, создавая по ним точные контурные рисунки, которые затем корректировали, сравнивая с исходным объектом.
Для описания мускулов и костных составляющих головы рыбы использованы термины, применяемые при изучении элементов скелетно-мышечной системы висцерального аппарата попугаевых (Tedman, 1980a, 1980b; Bellwood, 1985, 1994; Clements, Bellwood, 1988; Bellwood, Choat, 1990), а также других представителей губановидных (Labroidei) (Rognes, 1971; Goedel, 1974; Liem, Osse, 1975; Barel et al., 1976; Anker, 1978; Liem, 1979; Stiassny, 1981a). В некоторых исследованиях из последней категории также рассматриваются сухожилия и связки (Hasselt, 1978, 1979; Stiassny, 1981b, 1990). В работу включены обозначения ветвей черепно-мозговых нервов, которые заимствованы из описаний периферической нервной системы различных представителей Teleostei (Gierse, 1904; Manigk, 1933; Maheshwari, 1965; Saxena, 1969; Springer, Freihofer, 1976; Harrison, 1981; Nakae, Sasaki, 2006, 2007). Также вводятся некоторые термины для указания деталей структур мышечной, костной и соединительнотканной систем висцерального аппарата рубиново-пепельной рыбы-попугая: collis et valliculum vomerii, folium epioticus, m. epaxialis dorsalis, 1 et 2, manubrium maxillare, marsupium oris, membrana adductores, hyomandibularis 1, 2 et 3, pr. aculeus prooticum, pr. clavis anguloarticulare, pr. praeopercularis, рr. m. levator operculi, pr. levator pectoralis et pr. intercalare pteroticum, spina medialis et pr. clavis anguloarticulare, suffusorium dentale, tectum et cavum claustrum quadratum.
В тексте использованы следующие сокращения: m. – мускул (musculus), lig. – связка (ligamentum), t. – сухожилие (tendo), ap. – апоневроз (aponeurosis), pr. – отросток (processus), cr. – гребень (crista), f. – отверстие (foramen), c. – хрящ (catrilago), fen. – окно (fenestra), cap. – головка (caput), a. – артерия (arteria), v. – вена (vena), n. – нерв (nervus), r. – ветвь нерва (ramus).
РЕЗУЛЬТАТЫ
Особенности строения нейрокраниума
Этмоидальный отдел нейрокраниума рубиново-пепельной рыбы-попугая достигает значительного размера относительно других подразделений черепа. При взгляде сбоку заметно, что данная часть нейрокраниума формирует пологий скат, который служит основанием для переднезадних перемещений pr. ascendens praemaxillare небольшой протяжённости, имеющих место в ходе функционирования максиллярного аппарата.
При взгляде сверху mesethmoideum (рис. 1) − округлая кость, граничащая с vomer спереди, с ectoethmoideum – сбоку и с frontale – сзади. Для каждого из pr. ascendens praemaxillare дорсальная поверхность mesethmoideum и vomer формируют вытянутые канавки, разделённые сагиттальным cr. ethmoidalis, вследствие чего верхняя поверхность этмоидального отдела оказывается вогнутой относительно позади лежащих frontale. Cr. ethmoidalis не доходит до заднего края mesethmoideum. Между каудальным краем левой и правой половин округлого mesethmoideum и frontale соответствующей стороны головы имеется по треугольному отверстию (fen. ethmoidale). На данном участке нейрокраниума передние края frontale формируют правильный полукруг.
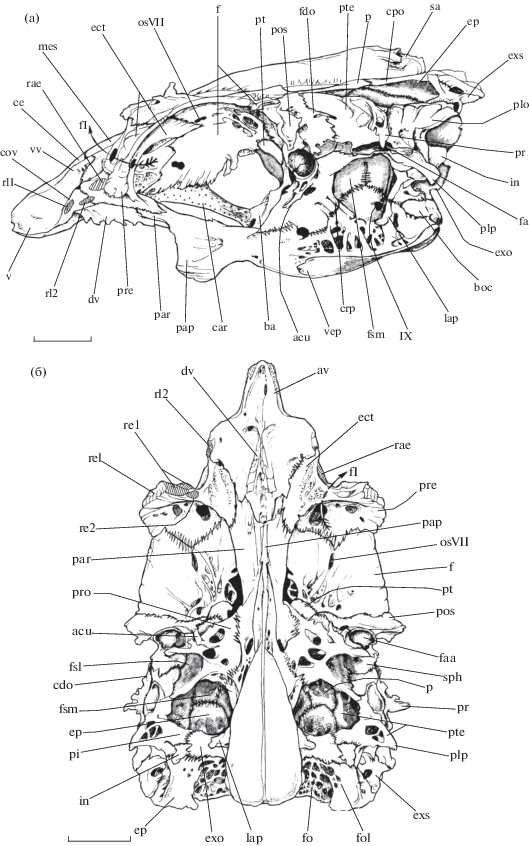
Рис. 1.
Нейрокраниум рубиново-пепельной рыбы-попугая Scarus rubroviolaceus, вид: а − сбоку, б − снизу, в – сверху; IX – n. glossopharyngeus (IX), acu − pr.
aculeus prooticum, av − vomer pars anterior (передний отдел сошника), ba – basisphenoideum,
boc – basioccipitale, car – cartilago, cdo − cr. dilatator operculi, ce − cr. ethmoidalis,
сov − collis vomeri (возвышение сошника), cpo − cr. posttemporalis, crp − cr. prooticus,
dv − dens vomeri, ect − ectoethmoideum, ep − epioticum, epe − regio articulatio epioticum
et pr. epioticus posttemporale, exo – exoccipitale, exs − extrascapulare, f – frontale,
fI – f. n. olfactorius (I), fa − fossa cap. articularis posterior hyomandibulare,
faa − fossa cap. articularis anterior hyomandibulare, fdo − fossa dilatator operculi,
fe − fen. ethmoidale, fl − fen. lateralis, fo − folium epioticus (лепесток epioticum),
fol − fossa occipitalis lateralis, fom − fossa occipitalis medialis, fp − fossa posttemporale,
fsl − fossa subtemporalis lateralis, fsm − fossa subtemporalis medialis, in – intercalare,
lap − cr. lateralis parasphenoideum, mes – mesethmoideum, osVII − foramen r. ophthalmicus
superficialis (VII), p – parietale, pap − pr. anterior parasphenoideum, par – parasphenoideum,
pi − pr. intercalare, plo − рr. m. levator operculi, plp − pr. levator pectoralis,
pos − pr. postorbitalis, pr − pr. praeopercularis, pre − pr. preorbitalis, pro – prooticum,
pt – pterosphenoideum, pte − pteroticum, rae − regio articulatio ectoethmoideum et
palatinum, re1 − regio lig. ectoethmoideum-palatinum 1, re2 − regio lig. ectoethmoideum-palatinum
2, rel − regio articulatio ectoethmoideum et lacrimale, rl1 − regio lig. ethmopalatinum
1, rl2 – regio lig. ethmopalatinum 2, sa − cr. sagittalis, sph − sphenoticum, su −
supraoccipitale, v − vomer, vep − cr. ventralis parasphenoideum, vv − valliculum vomeri
(валики сошника); ( ) − мускулатура, (
) − мускулатура, ( ) − регулярная соединительная ткань, (
) − регулярная соединительная ткань, ( ) − нерегулярная соединительная ткань, (
) − нерегулярная соединительная ткань, ( ) − хрящ, (
) − хрящ, ( ) – жир. Масштаб здесь и на рис. 3−6: 1 см.
) – жир. Масштаб здесь и на рис. 3−6: 1 см.
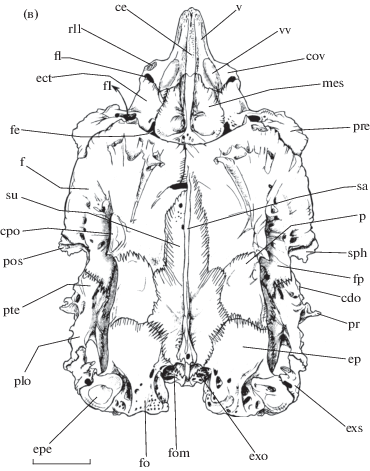
Ectoethmoideum (рис. 1) прободено крупным f. olfactorius для выхода из глазницы n. olfactorius (I), a. orbito-nasalis и v. orbito-nasalis. На границе ectoethmoideum и frontale имеется выходное f. r. ophthalmicus superficialis (VII). Крыловидная латеральная часть кости формирует pr. praeorbitalis, нижнебоковая сторона которого служит для прочного крепления lacrimale. Чуть ростральнее областей крепления lig. ectoethmoideum-palatinum 1 и 2 кость образует небольшую овальную плоскую площадку, покрытую при жизни рыбы тонким слоем хряща для причленения palatinum. Перед этой площадкой на границе ectoethmoideum и vomer имеется маленькая ямка, которая обозначает участок крепления lig. ethmopalatinum 2. Задняя часть ectoethmoideum формирует конусообразное углубление переднего миодома.
Разросшееся крупное vomer (рис. 1) имеет поверхность со сложным рельефом. По обе стороны от канавок pr. ascendens praemaxillare кость образует вытянутые валики (valliculum vomeri), контролирующие устойчивость движения рострального хряща относительно этмоидального отдела нейрокраниума. Выпуклая область vomer (collis vomeri) латеральнее и ниже valliculum vomeri служит местом расположения мениска 1 и крепления lig. ethmopalatinum 1. Впереди valliculum vomeri каждой стороны головы сливаются друг с другом и с сr. ethmoidalis и формируют ориентированный в сагиттальной плоскости и уплощённый с боков передний отдел vomer, контактирующий с lig. maxillare-praemaxillare anterior. Вентральная поверхность кости вогнута. На уровне collis vomeri на ней имеется маленький зубчатый гребень, проходящий сагиттально; по-видимому, это редуцированный ряд зубов vomer.
Основная часть пространства глазницы рыбы образована за счёт frontale (рис. 1): каждая из этих костей обеих сторон головы формирует мощную вентрально ориентированную пластину, остальная доля медиальной стенки глазницы образована соединительнотканной плёнкой. Задняя доля дорсальной поверхности кости участвует в формировании начальной области продольного cr. posttemporalis, медиально ограничивающего fossa posttemporale, а также переднего отдела cr. dilatator operculi – сверху ограничивающего fossa dilatator operculi. Маленькое pterosphenoideum (рис. 1а, 1б) образует латеральный край обширного орбитального отверстия, ведущего в полость нейрокраниума, в которой располагается головной мозг. Внушительный вентральный отросток basisphenoideum (рис. 1а) формирует мощную сагиттальную перегородку заднего миодома.
Слуховой отдел включает две обширные впадины − fossa subtemporalis lateralis и fossa subtemporalis medialis, внутренняя поверхность которых служит для отхождения волокон m. levator externus 4 и m. levator posterior (Громова, Махотин, 2019). Впадины отделены друг от друга костной перегородкой.
Компактное sphenoticum (рис. 1) образует треугольный pr. postorbitalis, а снизу – верхнюю половину округлой сочленовной впадины крупного размера, ориентированной дорсовентрально, для создания сустава с cap. articularis anterior hyomandibulare. Впадина данного сустава открыта в латеральном направлении. Её задняя сторона представлена выпуклой каудально костной перегородкой, вдающейся в пространство fossa subtemporalis lateralis. Sphenoticum образует около 50% вогнутой латерально поверхности fossa dilatator operculi и формирует срединную область cr. dilatator operculi. На участке перехода выдающегося вбок pr. postorbitalis в область fossa dilatator operculi наружный рельеф кости представлен небольшими гребнями и рёбрышками.
Разросшееся prooticum (рис. 1а, 1б) участвует в формировании рострального края переднего отверстия и наружной стенки заднего миодома. Верхняя часть кости образует нижнюю половину сочленовной впадины для cap. articularis anterior hyomandibulare. Сразу же ниже этой впадины prooticum формирует иглообразный отросток (pr. aculeus prooticum), который ориентирован вниз и латерально. Позади кость включается в создание основной доли (до 60%) внутренней поверхности fossa subtemporalis lateralis. Задняя сторона fossa subtemporalis lateralis находится в составе костной перегородки, отделяющей эту впадину от позади лежащей fossa subtemporalis medialis. По наружной стороне prooticum спускается невысокий cr. prooticus, ориентированный вниз и назад. Далее он устремляется на поверхность parasphenoideum и служит передней границей пространства fossa subtemporalis medialis.
Pteroticum (рис. 1) несёт узкую суставную впадину, ориентированную рострокаудально, для сочленения с cap. articularis posterior hyomandibulare. Впадина данного сустава открыта в латеровентральном направлении. Передненижняя часть pteroticum завершает образование fossa subtemporalis lateralis, а также включается в создание перегородки между fossa subtemporalis lateralis и fossa subtemporalis medialis. Дорсальная сторона кости сильно вогнута и образует большую часть вентральной поверхности fossa posttemporale, в формировании которой также участвуют frontale, parietale и epioticum. Pteroticum завершает создание cr. dilatator operculi и fossa dilatator operculi, наружный рельеф задней области которой составлен небольшими костными рёбрышками.
В составе каудальной половины кости можно выделить четыре отростка: pr. praeopercularis, рr. m. levator operculi, pr. levator pectoralis и pr. intercalare. Сr. dilatator operculi сзади включается в состав треугольного pr. praeopercularis, чьё остриё коротким lig. praeopercularis связано с дорсальным концом praeoperculum. Рr. praeopercularis ориентирован вниз и латерально. Прямым продолжением сr. dilatator operculi каудально служит верхний край рr. m. levator operculi, который направлен назад. Латеральная поверхность рr. m. levator operculi несёт несколько хорошо дифференцированных гребней, ориентированных дорсовентрально. От вентральной области рr. m. levator operculi вниз и назад устремляется треугольный pr. levator pectoralis, при взгляде сбоку имеющий палочковидную форму. Основание pr. levator pectoralis несёт маленькую платформу, которая служит местом прикрепления заднего края hyomandibulare при помощи прочных соединительнотканных волокон. Задненижняя часть pteroticum обособляется в виде толстого pr. intercalare, сочленяющегося с одноимённой костью и exoccipitale. Этот отросток принимает участие в образовании задней стенки fossa subtemporalis medialis, которая одновременно является перегородкой между этой впадиной и fossa occipitalis lateralis. Extrascapulare (рис. 1) у данного вида включено в состав нейрокраниума.
Epioticum (рис. 1) завершает формирование cr. posttemporalis, чей задний участок служит местом контакта кости с extrascapulare. Таким образом pteroticum, extrascapulare и epioticum создают замкнутое костное кольцо, канал которого является отверстием для проникновения m. epaxialis anterior 1 в пространство fossa posttemporale. Медиальнее этой площадки толщина epioticum значительно уменьшается, создавая тонкий, сильно перфорированный костный “лепесток”. Последний вместе с латеральной, более утолщённой частью кости составляет порцию внутренней поверхности и крыши fossa occipitalis lateralis.
Каудальная сторона затылочного отдела нейрокраниума несёт две впадины. Fossa occipitalis lateralis используется для начала волокон задней части m. levator posterior, а сильно перфорированная поверхность fossa occipitalis medialis является областью прикрепления m. epaxialis.
Латеральная доля exoccipitale (рис. 1) служит продолжением pr. intercalare pteroticum. От нижнего края “колпачка” intercalare по поверхности exoccipitale нисходит cr. lateralis exoccipitale. Exoccipitale завершает создание обширной fossa subtemporalis medialis, образованной помимо данной кости ещё шестью составляющими: prooticum, pteroticum, epioticum, parasphenoideum, parietale и supraoccipitale. Каудальная сторона exoccipitale формирует основную долю углублений fossa occipitalis lateralis и fossa occipitalis medialis.
Supraoccipitale (рис. 1а, 1в) формирует cr. sagittalis, тянунущийся вдоль большей части дорсальной поверхности нейрокраниума. Впереди cr. sagittalis практически достигает ростральной области контакта frontale контрлатеральных сторон головы рыбы. Задняя часть supraoccipitale образует верхнюю долю поверхности fossa occipitalis medialis.
Вентральная часть нейрокраниума составлена parasphenoideum (рис. 1а, 1б), протягивающимся от этмоидального до затылочного отделов. В глазничном отделе parasphenoideum формирует широкий уплощённый с боков pr. anterior. Через короткую перетяжку pr. anterior соединён со сжатым в сагиттальной плоскости невысоким cr. ventralis parasphenoideum. Приблизительно на уровне нахождения fossa subtemporalis lateralis по бокам от cr. ventralis формируется поперечно ориентированная сочленовная поверхность для образования сустава с верхней глоточной челюстью (по: Андрияшев, 1945а, 1945б, 1948а). В области затылочного отдела кость несёт выраженный cr. lateralis parasphenoideum, вдоль задней поверхности которого нисходит пучок ветвей n. vagus (X). Cr. lateralis снизу ограничивает пространство fossa subtemporalis medialis.
Строение нижнечелюстного сустава: quadratum и anguloarticulare
Поверхность челюстного сустава со стороны quadratum (рис. 2–5) ориентирована приблизительно под углом 90° относительно остального тела кости. Передняя головка quadratum имеет два отростка, взаимно переходящих друг в друга, каждый из которых несёт по мыщелку, совместно с anguloarticulare образующих челюстной сустав.
Рис. 2.
Устройство нижнечелюстного сустава рубиново-пепельной рыбы-попугая Scarus rubroviolaceus: а – сочленовная головка quadratum, вид спереди; б, в − anguloarticulare (б − вид сзади, в – с внутренней стороны); ac − apex pr. clavis, acc − apex cavum claustrum (острие полости замка), cc − cavum claustrum (полость замка), cco − planum convexus pr. clavis, cla − pr. clavis, cq − corpus quadratum (тело quadratum), cv – cavum ventralis, da − pr. dorsolateralis ascendens, dq − planum lateralis portio dorsalis quadratum, i − isthmus, man − planum medialis anguloarticulare, oc − os coronomeckeli, pcm − planum concavus medialis, pda − planum lateralis portio dorsalis аnguloarticulare, plr − planum articularis, pm − pr. medialis, pmq − planum medialis quadratum, pop − praeoperculum, pq − planum quadratus, pva − planum lateralis portio ventralis аnguloarticulare, re − retroarticulare, rv − ramus verticalis quadratum, sm − spina medialis, sul − sulcus, te – tectum (козырёк), vde − pr. ventromedialis descendens, vq − planum lateralis portio ventralis quadratum. Масштаб: 3 мм.
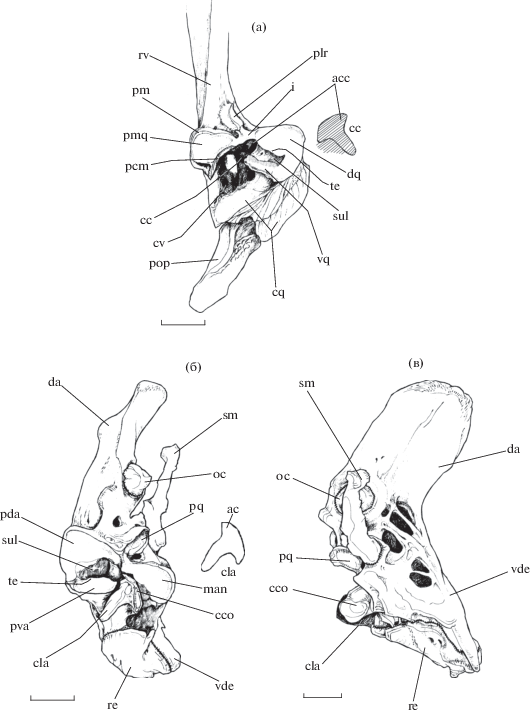
Рис. 3.
Некоторые мускулы, соединительнотканные структуры и нервы висцерального аппарата рубиново-пепельной рыбы-попугая Scarus rubroviolaceus, вид сбоку: а − после снятия окологлазничных костей; б – operculum и глазное яблоко изъяты, волокна m. adductor mandibulae частично удалены. A1a − portio A1a, А1а + А2 − stratum superficialis А1а + А2, A1б-1, 2, 3 − subdivisio № 1, 2, 3 portio A1б, aA2 − ap. portio A2, A3 – portio A3, aap – m. adductor arcus palatini, abs − m. abductor superficialis, ahh − mm. adductores hyohyoidei, ao – m. adductor operculi, aon − a. orbito-nasalis, bVII – r. buccalis (VII), baVII − r. buccalis accessoius (VII), cd − pr. coronoidalis dentale, cl – cleithrum, сmo − paries caudalis marsupium oris (задняя стенка кармана рта (marsupium oris)), crq − cr. quadratum, dA1a − pars dorsalis ap. portio A1a, da − pr. dorsolateralis ascendens, di − пальцевидный отросток (pr. digitatus) palatinum, do − m. dilatator operculi, ea1 − m. epaxialis anterior 1, emo − paries externa marsupium oris (наружная стенка кармана рта (marsupium oris)), epd − m. epaxialis dorsalis, epx − m. epaxialis, exs − extrascapulare, h – hyomandibulare, hVII − r. hyomandibularis (VII), hyVII – r. hyohyoidei (VII), imo − paries interna marsupium oris (внутренняя стенка кармана рта (marsupium oris)), ina − lig. integumentum anterior, inp − lig. integumentum posterior, iop – interoperculum, laX − r. lateralis (X), lav − m. levator arcus palatini, lfp − lig. frontale-palatinum, lim − lig. interoperculomandibulare, lmd − lig. mandibulare, lml − lig. maxillolacrimale, lmx − pr. lacrimalis maxillare, lo − m. levator operculi, lq − рr. lateralis quadratum, mVII − r. mandibularis (VII), mA1a − pars medialis ap. portio A1a, mh2 − membrana hyomandibularis 2, mxV − r. maxillaris (V), mxc – cr. lateralis maxillare, o – operculum, oh − pr. opercularis hyomandibulare, pdr – pr. dentalis praemaxillare, pll − lig. palatolacrimale, pop − praeoperculum, ptt − posttemporale, qu – quadratum, sA1a – перегородка (septa) ap. portio A1a, sA2 – перегородка (septa) ap. portio A2, sd − ковш (suffusorium) dentale, scl − supracleithrum, so – suboperculum, srA1б − subdivisio rostrale portio A1б, sua − m. supracarinalis anterior, vA1a − pars ventralis ap. portio A1a; ост. обозначения см. на рис. 1, 2.
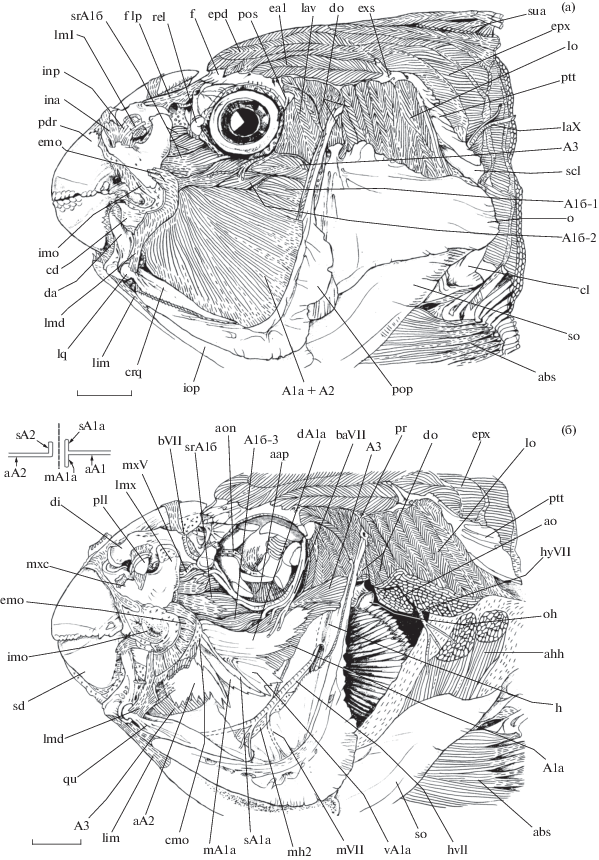
Рис. 4.
Некоторые мускулы, соединительнотканные структуры и нервы висцерального аппарата рубиново-пепельной рыбы-попугая Scarus rubroviolaceus, вид сбоку: а – мускульные волокна порций А1а, А2 и каудальных подразделений порции А1б удалены; б – manubrium maxillare, pr. alveolaris premaxillare сколоты, pr. coronoidalis dentale удалён частично; I – n. olfactorius (I), II – n. opticus (II), A3a − portio A3a, Awb – portio Awb, aA1б − ap. subdivisio rostrale portio A1б, aA2 − ap. portio A2, apx − pr. ascendens praemaxillare, arl − lig. articulodentale lateralis, cen − cr. entopterygoideum, com − сухожильные спайки (comissura), ec − ectopterygoideum, ep1 − lig. ectoethmoideum-palatinum 1, fA3 − fasciculus portio A3, imx − integumentum apparatus maxillaris, lpa − lig. praeoperculum-articulare, lpp − lig. palatine-praemaxillare, mA3V – r. mandibularis portio A3, mad − membrana adductores, mdV − r. mandibularis (V), mh1 − membrana hyomandibularis 1, mo − глазодвигательные мышцы (m. bulbus oculi), mx − maxillare, mxd − lig. maxillo-dentale, orp − textus organum palatinum, sy − symplecticum, t1A1a − t. portio A1a № 1, tA1a − t. portio A1a, tA2 − t. portio A2, tA3 − t. portio A3, vde − pr. ventromedialis descendens; ост. обозначения см. на рис. 1, 3.
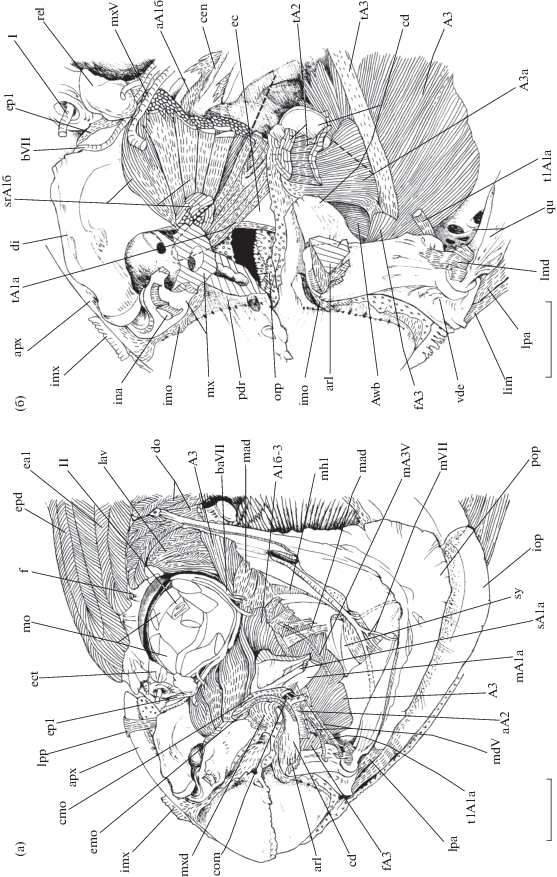
Рис. 5.
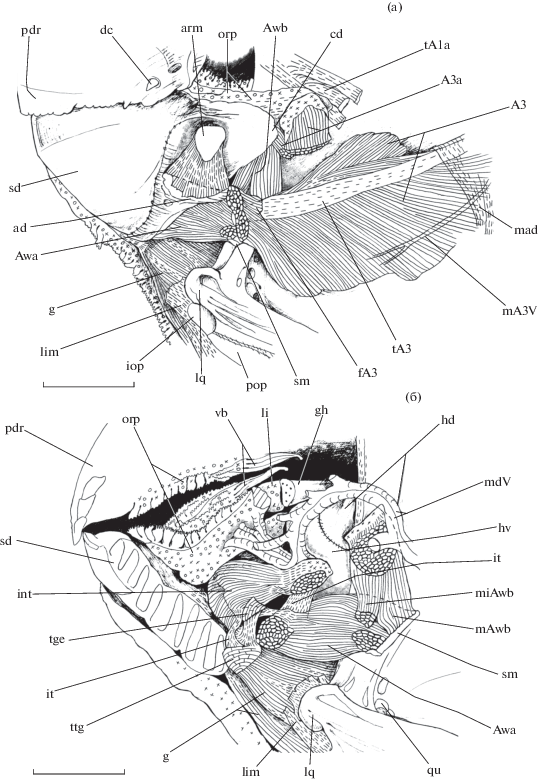
Порции m. adductor mandibulae рубиново-пепельной рыбы-попугая Scarus rubroviolaceus в области нижней челюсти, вид сбоку (аnguloarticulare, за исключением spina medialis, удалено): а – pr. coronoidalis dentale частично сколот; б − левое dentale полностью удалено, виден симфизный шов правого dentale; ad − pr. angulare dentale, arm − lig. articulodentale medialis, Awa – portio Awa, dc − клыковидный зуб (dentis caninus), g − m. geniohyoideus, gh − glossohyale, hd − hypohyale dorsale, hv − hypohyale ventrale, int – m. intermandibularis, it − сухожильный промежуток (intervallum tendineus), li − lingua, mAwb − caput magnus Awb, miAwb − caput minimus Awb, sm − spina medialis, tA1a − t. portio A1a, tge − t. geniohyoideus, ttg − locus tendinosus terminalis m. geniohyoideus, vb − ротовые клапаны (valvula buccalis); ост. обозначения см. на рис. 3, 4.
Более крупный рr. lateralis является продолжением мощного cr. quadratum. Рr. lateralis формирует протяжённую выпуклую сочленовную поверхность, располагающуюся на его верхней, передней и нижней сторонах. При взгляде на сустав спереди заметно, что эта поверхность состоит из двух частей – верхней (planum lateralis portio dorsalis quadratum) и нижней (planum lateralis portio ventralis quadratum), разделённых желобком. Вентральный край верхней части поверхности нависает козырьком (tectum) над желобком и её нижней частью. Меньший по размеру pr. medialis имеет выпуклую сочленовную поверхность (planum medialis quadratum) на своих передней и верхней сторонах; нижняя сторона отростка несёт вогнутый участок для сочленения с костным ключом.
В области перехода между сочленовными поверхностями pr. lateralis et medialis находится глубокое вдавление приблизительно треугольной формы – полость замка (cavum claustrum). Своим остиём эта полость ориентирована в сторону желобка сочленовной поверхности pr. lateralis. В каудальном направлении эта полость сливается с более обширной вентральной (cavum ventralis), которая уходит внутрь тела quadratum. Там её стенки становятся многократно перфорированными.
Латеральная сторона вертикальной пластины quadratum в области контакта pr. lateralis et medialis несёт маленькую костную площадку c плоской суставной поверхностью (planum articularis) круглой формы.
Внутренняя сторона anguloarticulare (рис. 2б, 2в, 3а, 4б) в месте перехода лопатообразного pr. dorsolateralis ascendens в pr. ventromedialis descendens сильно перфорирована. В основании pr. dorsolateralis ascendens, непосредственно позади spina medialis, находится плоская сочленовная поверхность (planum quadratus) для образования соотвествующего сустава с quadratum. Эта поверхность ориентирована вниз и медиально. Spina medialis хорошо заметным швом отделён от тела anguloarticulare. Немного выше точки прикрепления spina medialis pr. dorsolateralis ascendens несёт маленький колпачок os coronomeckeli (рис. 2б, 2в). На вентральной стороне pr. ventromedialis descendens расположено вытянутое retroarticulare значительного размера.
Каудальная сторона anguloarticulare несёт двураздельную вогнутую суставную поверхность для причленения двух соотвествующих мыщелков quadratum. Эта суставная поверхность ориентирована под острым углом относительно оси тела anguloarticulare и состоит из двух долей – латеральной с большей площадью (planum lateralis anguloarticulare) и меньшей медиальной доли (planum medialis anguloarticulare). Так же, как и в случае quadratum, planum lateralis anguloarticulare включает верхнюю (planum lateralis portio dorsalis аnguloarticulare) и нижнюю (planum lateralis portio ventralis аnguloarticulare) части и разделяющий их желобок. Дорсальный край нижней части planum lateralis anguloarticulare выдвинут каудально относительно полости желобка и верхней части planum lateralis anguloarticulare.
В области перехода между своими суставными поверхностями кость формирует выдающийся назад pr. clavis − костный ключ треугольной формы. При взгляде на anguloarticulare каудально ключ имеет короткую медиальную сторону с выпуклой поверхностью, длинную латеральную сторону с плоской поверхностью и центральное остриё. По-видимому, pr. clavis является преобразованным ретроартикулярным отростком (pr. postarticularis).
Передние челюсти: dentale и praemaxillare
Dentale (рис. 3−5) контрлатеральных сторон головы рыбы прочно связаны между собой зубчатым швом так, что левая и правая кость совместно формируют глубокий ковш (suffusorium) нижней челюсти. Задненижняя доля suffusorium образует треугольный pr. angulare, по латеральной стороне которого расходятся небольшие гребни. Задняя часть dentale представлена широким pr. coronoidalis. При взгляде на нижнюю челюсть спереди заметно, что тело pr. coronoidalis ориентировано дорсолатерально, а его вогнутая наружная поверхность покрыта мелкими рёбрышками. Между pr. coronoidalis и pr. angulare находится глубокая вырезка внутринижнечелюстного сустава (intramandibular joint − IMJ) (по: Konow et al., 2008) dentale c pr. dorsolateralis ascendens anguloarticulare.
Передняя доля рraemaxillare (рис. 3а, 4, 5) формирует pr. dentalis c широкой гладкой поверхностью. Кости левой и правой сторон головы соединены между собой при помощи симфиза. Вентральная часть рraemaxillare образует pr. alveolaris lateralis et medialis, которые обхватывают manubrium maxillare соответственно снаружи и изнутри, фиксируя относительно praemaxillare. Длина pr. ascendens превышает таковую pr. dentalis. Латеральная поверхность pr. ascendens покрыта рёбрышками, ориентированными в основании отростка спереди−назад, а на его конце – задневентрально.
Зубы, находящиеся на dentale и praemaxillare, сливаются между собой и с несущими их костями, образуя монолитные зубные пластинки.
Особенности строения некоторых висцеральных и соматических мышц аппарата питания
Сложно устроенный m. adductor mandibulae (рис. 3–6) является самым крупным мускулом висцерального аппарата и включает семь порций, которые оканчиваются как на верхней, так и на нижней челюсти. После снятия кожи с головы рыбы открывается внешне единый поверхностный слой порций А1а + А2 (рис. 3а); граница раздела между порциями А1а и А2 снаружи не прослеживается. Волокна этого поверхностно единого слоя берут начало с узких участков костей по периметру fossa adductores суспензориума − вентральной ветви hyomandibulare вокруг отверстия для выхода r. hyomandibularis VII (рис. 3б), узкого участка praeoperculum, заднего угла quadratum и небольшой его полоски на медиальной стороне cr. quadratum. Местом окончания служит задний край marsupium oris. Сверху мускульные волокна описываемого мускульного слоя контактируют с волокнами подразделений порции А1б. По мере удаления волокон поверхностного слоя m. adductor mandibulae в его глубине появляются две крупные пластины конечных апоневрозов, обозначающие наличие в мускуле двух отдельных порций – дорсальной А1а и вентральной А2.
Рис. 6.
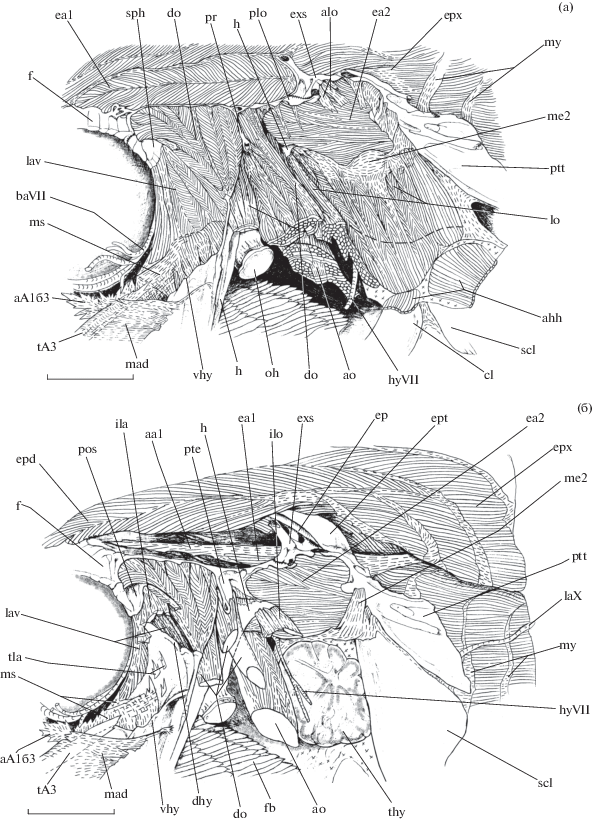
Mускулы в области причленения суспензориума к нейрокраниуму и membrana suspensorii рубиново-пепельной рыбы-попугая Scarus rubroviolaceus, вид сбоку: а – поверхностный слой ((− −) − граница прикрепления m. levator operculi к operculum); б – более глубокий слой: aA1б3 − ap. subdivisio № 3 portio А1б, aa1 − ap. epaxialis anterior 1, alo − ap. m. levator operculi, dhy − cr. dorsalis hyomandibulare, ea2 − m. epaxialis anterior 2, ept − pr. epioticus posttemporale, fb − filum branchiale, ila − ap. initialis m. levator arcus palatini, ilo − ap. initialis m. levator operculi, me2 − membrana m. epaxialis anterior 2, ms − membrana suspensorii, my − myosepta, tla − ap. terminalis m. levator arcus palatini, thy − thymus, vhy − cr. ventralis hyomandibulare; ост. обозначения см. на рис. 1, 3, 4 .
Веерообразная порция А1а (рис. 3б, 4, 5а) берёт начало с каудального угла quadratum, symplecticum, задней части metapterygoideum, передней области praeoperculum, вентральной ветви hyomandibulare, а также c поверхности membrana hyomandibularis 2. Кроме того, областью начала порции служит нижняя часть наружной поверхности membrana adductores. Мускульные волокна сходятся на крупном конечном ap. portio A1a с его наружной и внутренней стороны. В пластине ap. portio A1a можно выделить несколько составляющих: дорсальную (pars dorsalis), вентральную (pars ventralis) и медиальную (pars medialis) части, а также перегородку (septa ap. portio A1a).
Основная доля апоневроза представлена крупными pars dorsalis (имеющей парасагиттальную ориентацию) и pars ventralis, нижняя часть которой наклонена в медиальном направлении, т.е. в сторону поверхности суспензориума. Pars dorsalis и pars ventralis разделены между собой рядом соединительнотканных рёбрышек, идущих по пластине апоневроза. Верхний край pars dorsalis утолщён в виде валика. Таким образом, эти две части пластины представляют собой две плоскости, расположенные под углом друг к другу и являющиеся местом окончания большинства волокон порции А1а. Вдоль передневентрального края pars dorsalis и pars ventralis тянется septa ap. portio A1a, выдающаяся латерально. Продолжением septa ap. portio A1a и вышеупомянутой основной доли пластины апоневроза в медиальном направлении служит его pars medialis. Она ориентирована под углом 90° к плоскости суспензориума. От нижней поверхности рострального участка pars medialis отходит длинное и блестящее t. portio A1a № 1. Оно оканчивается, постепенно сужаясь, на внутренней стороне anguloarticulare ниже os coronomeckeli. Это сухожилие ориентировано передневентрально и проходит медиальнее волокон порции А2, но латеральнее волокон порции А3.
Впереди ap. portio A1a утолщается и сужается, осуществляя присоединение к задней стенке marsupium oris, а также тянущимися сухожильными спайками (comissura) – к медиальной поверхности конечного участка пластины ap. portio A2. После этого ростральный конец ap. portio A1a распадается на многочисленные блестящие сухожилия, ориентированные вверх и вперёд. Они образуют крупного размера толстый пучок, прикрепляющийся к внутренней стороне manubrium maxillare впереди и выше области окончания собрания сухожилий ростральных подразделений порции А1б. Таким образом, скопление сухожилий порции А1а и скопление сухожилий подразделений порции А1б оканчиваются на maxillare под разными углами.
Порция А1б (рис. 3, 4, 6) представлена серией мускульных подразделений (subdivisio) с умеренно выраженными границами. Число подразделений и степень их дифференцировки возрастают с размером особи: у исследованных экземпляров их было от пяти до восьми. Подразделения выстроены чередой друг за другом вдоль верхнего отдела суспензориума от hyomandibulare до entopterygoideum. Передняя часть позади лежащего подразделения накладывается на заднюю часть впереди расположенного. Кроме того, каждое впереди находящееся подразделение сверху несколько нависает над позади лежащим, а медиально они взаимно переходят друг в друга. Часто в щель на границе двух соседних подразделений проникают rr. mandibularii V, а также венозные и артериальные сосуды, обсуживающие остальные части m. adductor mandibulae. Опишем особь рубиново-пепельной рыбы-попугая с шестью подразделениями порции А1б.
Волокна подразделения № 1 А1б (рис. 3а) берут начало с фрагмента вентральной ветви hyomandibulare сразу же ниже области прикрепления порции А3. Оканчиваются на участке дорсокаудальной поверхности ap. portio A1a pars dorsalis, пластина которого в небольшой степени закручивается вокруг подразделения № 1 А1б, формируя сухожильную раковину и тем самым частично обособляя его от находящегося медиально и впереди подразделения № 2. Эта раковина охватывает волокна подразделения № 1 А1б сверху, спереди, латерально и изнутри. Снизу волокна подразделений № 1−3 переходят в состав порции А1а.
Подразделение № 2 А1б (рис. 3а) начинается с участка вентральной ветви hyomandibulare непосредственно каудальнее задней границы прикрепления membrana adductores, а также с блестящей наружной стороны membrana adductores. Волокна подразделения оканчиваются на верхнем утолщённом крае и верхней области медиальной поверхности ap. portio A1a pars dorsalis.
Основная масса волокон подразделения № 3 А1б (рис. 3б, 4а) начинается с крупной пластины начального ap. portio А1б № 3, прикрепляющегося к костному гребню. Последний формируется верхними краями костей суспензориума − metapterygoideum и entopterygoideum. Местом отхождения ap. portio А1б № 3 служит область этого гребня, где расположен зубчатый шов между двумя вышеупомянутыми костями; апоневроз ориентирован рострокаудально. Помимо ap. portio А1б № 3 подразделение № 3 А1б начинается с передней зоны membrana adductores и с соседних участков metapterygoideum и entopterygoideum. Волокна оканчиваются на верхней области медиальной поверхности ap. portio A1a pars dorsalis, а также на коротких маленьких пластинках конечных апоневрозов, которые формируют верхний край ap. portio A1a на уровне его прикрепления к marsupium oris.
Ростральные подразделения (у нашей особи это подразделения № 4, 5 и 6 А1б) (рис. 3, 4б) ориентированы переднекаудально и берут начало с сильновогнутой поверхности entopterygoideum, покрытой для этого несколькими хорошо дифференцированными гребнями, расходящимися веерообразно. Каждый из этих гребней несёт по пластине начального апоневроза для отхождения волокон соответствующего подразделения. Областью начала служат также небольшой участок передней поверхности metapterygoideum и верхняя часть quadratum, над которыми латерально нависает порция А3. Степень слияния медиальных областей подразделений нарастает в ростральном направлении. Латеральные волокна подразделения № 4 не имеют чёткой границы с волокнами порции А1а, прикрепляющимися к медиальной стороне ap. portio A1a. Волокна ростральных подразделений оканчиваются на поверхностных и глубинных узких пластинах конечных апоневрозов, имеющихся по несколько штук для каждого подразделения. Впереди эти апоневрозы переходят в состав многочисленных толстых и блестящих сухожилий. Эти сухожилия собираются в крупный пучок и прикрепляются к внутренней поверхности manubrium maxillare в основании pr. lacrimalis (рис. 3б). Вентрально скопление сухожилий подразделений А1б не имеет чёткой границы с группой конечных сухожилий порции А1а.
Волокна порции А2 (рис. 3б, 4) начинаются с медиальной стороны выраженного cr. quadratum и с вогнутой поверхности нижнего отдела quadratum, вентральнее и каудальнее области прикрепления на этой кости порции А3. Областью крепления также служит небольшой участок symplecticum и фрагмент membrana hyomandibularis 2. Волокна порции следуют вверх и вперёд и оканчиваются на пластине ap. portio A2. На уровне прохождения septa ap. portio A1a по пластине апоневроза порции А1 верхний край ap. portio A2 загибается латерально, таким образом морфологически формируя подобную же перегородку. Рострально ap. portio A2 имеет прикрепление к задней стенке marsupium oris, а затем делится на многочисленные блестящие сухожилия, оканчивающиеся плотным пучком на медиальной поверхности задней половины pr. coronoidalis dentale (рис. 3а, 4, 5а). Недалеко от области прикрепления описываемых сухожилий порции А2, между последними и вентральными соединительнотканными волокнами сухожилий порции А1а, имеется щель, в которой расположено крупное сплетение венозных сосудов.
Таким образом, зона взаимного перехода мускульных волокон между порцией А1а и порцией А2 находится между верхним краем ap. portio A2, представленным его перегородкой, и нижней поверхностью ap. portio A1a, образованной pars medialis и septa ap. portio A1a. Точной границы между этими двумя порциями провести нельзя.
Уплощённая порция А3 (рис. 3, 4, 5а, 6) протягивается наклонно вдоль центральной зоны fossa adductores суспензориума, нисходя от его дорсокаудальной области вентрорострально. Таким образом, порция А3 расположена под углом по отношению к порциям А1а и А2. Волокна порции А3 начинаются с наружной стороны membrana suspensorii, маленького фрагмента поверхности hyomandibulare между линиями крепления membrana suspensorii и membrana adductores, верхнего участка membrana hyomandibularis, а также срединных областей metapterygoideum и quadratum. У крупных особей волокна сходятся на небольшом апоневрозе, который быстро переходит в длинное уплощённое t. portio A3, тянущееся вдоль всей длины порции; у особей меньшего размера имеется только t. portio A3 без апоневроза. Порция А3 оканчивается при помощи t. portio A3 на os coronomeckeli, а также мускульно, прикрепляясь к заднему краю pr. dorsolateralis ascendens ниже os coronomeckeli.
Стоит отметить, что в области окончания t. portio A3 из порции выделяется маленький пучок мускульных волокон (fasciculus portio A3) (рис. 4, 5а), для которых участком отхождения служит латеральная поверхность t. portio A3, а местом окончания − задний край pr. dorsolateralis ascendens выше os coronomeckeli.
Треугольной формы уплощённая порция А3а (рис. 4б, 5а) широко начинается с дорсальной поверхности конечного участка t. portio A3, а также с небольшой области веерообразно ориентированных мускульных волокон порции А3, которые сходятся на t. portio A3, прикрепляясь к его верхнему краю. Помимо этого передненижний угол порции А3а соединён сухожильным промежутком с fasciculus portio A3. Волокна порции А3а оканчиваются в плотной соединительнотканной выстилке медиальной стороны pr. coronoidalis dentale, напрямую не контактируя с костью. Это место прикрепления находится медиальнее и немного впереди области окончания порции А2. Таким образом, зоны крепления порций А3а и А2 накладываются одна на другую.
Порция Аwа (рис. 5) ориентирована горизонтально и представляет собой пучок волокон, соединяющий находящиеся каудально spina medialis и маленький фрагмент внутренней поверхности pr. dorsolateralis ascendens вентроростральнее os coronomeckeli с расположенным впереди участком медиальной стороны нижней области suffusorium dentale (рис. 3б, 5). Место окончания порции на dentale находится немного выше и латеральнее locus tendinosus terminalis m. geniohyoideus, но ниже участка крепления t. geniohyoideus (рис. 5) (Громова, Махотин, 2019). Верхнепередняя область порции Awa связана одним сухожильным промежутком с t. geniohyoideus, а другим – с m. intermandibularis в районе прикрепления последнего к dentale.
Порция Аwb (рис. 4б, 5) расположена впереди и медиальнее по отношению к порции А3а. Волокна порции Аwb начинаются двумя головками с внутренней стороны anguloarticulare. Меньшая по размеру головка отходит от маленького фрагмента медиальной поверхности pr. dorsolateralis ascendens дорсоростральнее os coronomeckeli. Более крупная уплощённая головка берёт начало с верхней половины spina medialis. После объединения двух головок порция Awb широко оканчивается на медиальной стороне pr. coronoidalis в области перехода его в suffusorium dentale. Это место крепления, помимо кости, также включает в себя передний фрагмент медиальной поверхности конечного участка порции А3а, а также соединительнотканную выстилку медиальной стороны pr. coronoidalis.
Небольшой m. intermandibularis (рис. 5б) ориентирован горизонтально таким образом, что левая и правая части мускула, направляясь навстречу друг к другом, в поперечной плоскости граничат между собой посредством центральной миосепты. Мускульные волокна соединяют между собой левую и правую доли suffusorium dentale, прикрепляясь к срединным участкам внутренней поверхности половинок нижней челюсти.
Абдукторный комплекс суспензориума включает в себя три слабо дифференцированных друг от друга мускула – m. levator arcus palatini, m. dilatator operculi и m. levator operculi.
Конусовидный m. levator arcus palatini (рис. 3а, 4а, 6) образует задний край глазницы, берёт своё начало с рr. postorbitalis и небольшой лежащей ниже него области fossa dilatator operculi непосредственно над сочленовной впадиной для hyomandibulare. При взгляде сбоку хорошо видно перистое строение этого мускула, выраженное в правильном чередовании сухожильных промежутков и упорядоченно уложенных между ними мускульных волокон. М. levator arcus palatini включает в себя чрезвычайно большое количество узких пластин начальных апоневрозов, веерообразно расходящихся и насыщающих весь объём мускула. Местом окончания мускульных волокон служит латеральная поверхность ориентированного каудовентрально cr. dorsalis hyomandibulare (рис. 6б) и наружная сторона срединной области кости ниже этого отростка, но выше cr. ventralis hyomandibulare, а также вентральная половина pr. cap. articularis anterior hyomandibulare и внутренняя поверхность membrana suspensorii. У крупных особей на поверхности hyomandibulare можно обнаружить небольшие конечные апоневрозы значительно меньшего размера по сравнению с начальными. Приблизительной границей раздела между m. levator arcus palatini и волокнами позади лежащего m. dilatator operculi служит cr. dorsalis hyomandibulare.
На нейрокраниуме m. dilatator operculi (рис. 3, 4а, 6) берёт своё начало с участка frontale, формирующего переднюю область cr. dilatator operculi, таким образом слегка нависая над местом отхождения волокон m. levator arcus palatini, а также с поверхности fossa dilatator operculi, с передней и задней сторон pr. praeopercularis, и c небольшого фрагмента pteroticum. Последний располагается непосредственно перед ростральным краем m. epaxialis anterior 2. Кроме того, мускул начинается с поверхности верхнего отдела hyomandibulare дорсальнее и медиальнее его cr. dorsalis, с верхней половины pr. cap. articularis anterior и pr. opercularis (рис. 3б, 6а), а также с pr. cap. articularis posterior. М. dilatator operculi насыщен большим количеством сухожильных образований, с преобладанием конечных из них над начальными. В области fossa dilatator operculi волокна m. dilatator operculi прикрепляются к небольшим начальным апоневрозам, отходящим от костных рёбрышек составляющих рельеф её поверхности. Малая доля поверхностных волокон мускула, берущих начало с cr. dilatator operculi, собирается на вершине cr. dorsalis hyomandibulare. Основная масса волокон устремляется вентрокаудально, проходя медиальнее дорсального конца praeoperculum, и сходится на больших, толстых и вытянутых пластинах конечных апоневрозов, которые прикрепляются ко внутренней стороне верхней области operculum; также присутствует и мускульное окончание. Размер и толщина пластин увеличиваются в направлении вперёд: наиболее крупные из них крепятся к pr. dorsalis operculum. Чёткая граница между m. dilatator operculi и находящимися позади волокнами m. levator operculi не прослеживается: приблизительно мы провели её на уровне переднего края m. epaxialis anterior 2.
Широкий, сильно уплощённый в парасагиттальной плоскости m. levator operculi (рис. 3, 6) начинается с латеральной поверхности membrana m. epaxialis anterior 2, а также с узких участков костей по её периметру: с маленького фрагмента наружной стороны дорсокаудального угла hyomandibulare, верхнего края рr. m. levator operculi pteroticum, нижнего края extrascapulare и переднего края posttemporale. Толщина мускула постепенно увеличивается в каудоростральном направлении. М. levator operculi содержит большое количество узких начальных апоневрозов, которые крепятся к костным гребням рr. m. levator operculi pteroticum, кромке extrascapulare и переднему краю posttemporale. Толстые и блестящие их пластины, отходящие от hyomandibulare, частично переходят в состав ростровентральной доли membrana m. epaxialis anterior 2. М. levator operculi оканчивается на медиальной поверхности верхнего отдела operculum. При этом вентральный край области прикрепления контактирует c дорсальным краем участка окончания на данной кости mm. adductores hyohyoidei (рис. 3б, 6а).
После удаления operculum становится виден компактный, толстый в поперечном сечении m. adductor operculi. (рис. 3б, 6). Мускул нисходит вентролатерально от участка нижней области pr. intercalare pteroticum и граничащего с ним швом фрагмента exoccipitale. Местом окончания m. adductor operculi служит маленькая зона медиальной поверхности operculum, расположенная сразу же каудальнее сустава данной кости с pr. opercularis hyomandibulare. Эта область прикрепления мускула находится на уровне выделенной нами приблизительной границы m. dilatator operculi и m. levator operculi. М. adductor operculi включает в себя значительную долю сухожильного компнента.
M. adductor arcus palatini формирует дно глазницы, в области которой его наружная сторона выстлана слоем прочной блестящей соединительной ткани. Это покрытие распространяется от верхнего края parasphenoideum по поверхности мускула; направление волокон соединительной ткани и мускула совпадает. Каудально область начала m. adductor arcus palatini включает в себя узкую полоску переднего края prooticum сразу же ниже его pr. aculeus. Ниже участок отхождения волокон перемещается на поверхность parasphenoideum в районе перехода его cr. ventralis в pr. anterior. Ростральнее мускул начинается с латеральной стороны pr. anterior и остальной впереди расположенной поверхности данной кости, немного вогнутой нижней стороны ectoethmoideum, а также со всей обширной вентральной площади vomer, включая его передний отдел. Центральный зубчатый гребень vomer служит границей раздела между m. adductor arcus palatini контрлатеральных сторон головы рыбы. В составе мускула количество и размер пластин начальных апоневрозов возрастают в направлении вперёд. Наименьшая толщина m. adductor arcus palatini наблюдается в его задней области, там, где участок его отхождения от нейрокраниума весьма узок. Чрезвычайно тонкий слой мускульных волокон присутствует в составе соединительнотканной мембраны, формирующей задненижнюю поверхность глазницы и прикрепляющейся к переднему краю hyomandibulare. Мы полагаем, что эта группа волокон может относиться к m. adductor arcus palatini. В ростральном направлении площадь поперечного сечения мускула возрастает, достигая максимума в срединном отделе m. adductor arcus palatini. Область окончания на внутренней стороне суспензориума включает в себя membrana hyomandibularis 1, верхнюю половину metapterygoideum, entopterygoideum и малый фрагмент каудальной области ectopterygoideum. Стоит отметить, что медиальная поверхность entopterygoideum в районе прикрепления m. adductor arcus palatini снабжена многочисленными хорошо дифференцированными гребнями. Таким образом, основная доля волокон мускула оканчивается на суспензориуме, однако небольшая часть медиальных из них завершается в слое ткани нёбного органа, находящегося в крыше ротовой полости рыбы (Громова, Махотин, 2020).
M. adductor hyomandibularis не выражен.
М. epaxialis (рис. 3, 6) в области нейрокраниума служит источником нескольких подразделений, отличающихся по областям своего прикрепления на черепе: m. epaxialis dorsalis, m. epaxialis anterior 1 и 2, а также m. levator pectoralis.
M. epaxialis dorsalis (рис. 3а, 4а, 6б) представлен двумя контрлатеральными половинами, разделёнными cr. sagittalis, и формируют мощный массив соматической мускулатуры, располагающейся на дорсальной поверхности нейрокраниума. M. epaxialis dorsalis прикрепляется к верхней стороне frontale, начиная с переднего края этой кости на границе с mesethmoideum. Мускул занимает всю площадь frontale, за исключением латеральной области, которая формирует крышу глазницы и fossa posttemporale. В каудальном направлении зона отхождения волокон сбоку ограничена cr. posttemporalis и включает дорсальные поверхности parietale, supraoccipitale, epioticum и pr. epioticus posttemporale, а также верхний край extrascapulare. M. epaxialis dorsalis содержит в себе миосепты, позади непрерывен с m. epaxialis и на нейрокраниуме располагается медиально по отношению к m. epaxialis anterior 1.
M. epaxialis anterior 1 (рис. 3а, 4а, 6) имеет вид рострокаудально ориентированного длинного мускульного пера, которое берёт своё начало с поверхности fossa posttemporale при помощи вытянутого в горизонтальной плоскости ap. epaxialis anterior 1. В районе завершения fossa posttemporale и на уровне нахождения extrascapulare латеральные волокна задней части мускула непрерывны с волокнами m. epaxialis anterior 2. Каудально пластина ap. epaxialis anterior 1 включается в состав одной из миосепт позади расположенной массы m. epaxialis, в которую без заметной границы переходит m. epaxialis anterior 1.
M. epaxialis anterior 2 (рис. 6) начинается с нижнего края extrascapulare, латеральной поверхности рr. m. levator operculi, заднемедиальной стороны pr. levator pectoralis, а также с задней кромки и внутренней поверхности дорсокаудального угла hyomandibulare. Область отхождения мускула от hyomandibulare сильно сухожильна. Глубинные волокна m. epaxialis anterior 1 и 2 берут начало с миосепты, которая крепится к заднему краю pteroticum и задненижней кромке epioticum. Эта миосепта прободается pr. intercalare posttemporale и отделяет описываемые мускулы от m. levator posterior. Поверхностый слой волокон m. epaxialis anterior 2 оканчивается на переднем крае posttemporale и supracleithrum. Глубинные волокна мускула переходят в состав m. epaxialis. Нижний край m. epaxialis anterior 2 слегка нависает над верхним краем m. levator pectoralis. Чёткой границы между этими двумя мускулами выделить нельзя − они взаимно переходят друг в друга.
Конусовидный m. levator pectoralis располагается непосредственно медиальнее тимуса (рис. 6б) так, что нижний край мускула совпадает с нижним краем железы. М. levator pectoralis берёт начало при помощи блестящего толстого пучка соединительнотканных волокон, отходящего от кончика pr. levator pectoralis. Область отхождения мускула расположена непосредственно медиальнее места прикрепления вентральных волокон m. epaxialis anterior 2 к hyomandibulare. Каудально m. levator pectoralis расходится веерообразно и оканчивается на передней поверхности supracleithrum и cleithrum, выше участка крепления t. corpus muscularis (Громова, Махотин, 2019). Часть глубинных волокон мускула непрерывна с волокнами m. epaxialis.
Соединительнотканные элементы аппарата питания
Угол рта рыбы снабжён широким соединительнотканным карманом (marsupium oris) (рис. 3, 4а); он становится виден после снятия кожи c головы животного в области pr. coronoidalis dentale, pr. alveolaris lateralis praemaxillare и ручки (manubrium) maxillare. Карман представляет собой полое образование, которое имеет наружную, внутреннюю и заднюю стенки; его входное отверстие расположено спереди. Сверху пространство кармана ограничено lacrimale, каудально – задним краем pr. coronoidalis и manubrium maxillare, а снизу оканчивается в области контакта участка pr. dorsolateralis ascendens anguloarticulare и нижнего края pr. coronoidalis.
Наружная стенка marsupium oris (рис. 3, 4a) составлена толстым слоем блестящих соединительнотканных волокон, которые веерообразно расходятся в каудальном направлении, а рострально сходятся в конус и образуют наружную губу верхней челюсти рыбы. Нижний край этой губы тянется приблизительно параллельно озубленному краю praemaxillare. Каудальная линия крепления наружной стенки marsupium oris включает задние края manubrium maxillare и pr. coronoidalis dentale. Волокна нижней области стенки участвуют в формировании lig. mandibulare. Наружная стенка кармана в области своей каудальной границы переходит в состав его маленькой по размеру задней стенки. Задняя стенка представлена плотной соединительнотканной выстилкой. Она связывает участки заднего края и медиальной поверхности pr. coronoidalis dentale с внутренней стороной manubrium maxillare.
Внутренняя стенка marsupium oris (рис. 3, 4б) представлена пигментированным соединительнотканным слоем, выстилающим наружные поверхности pr. coronoidalis dentale, pr. alveolaris lateralis praemaxillare и ручки (manubrium) maxillare. Этот слой частично проникает в щели между этими костями, сверху становясь наружной выстилкой pr. dentalis praemaxillare (рис. 3а, 5) в месте его перехода в pr. ascendens, а снизу включается в состав lig. mandibulare. Вентральный край pr. coronoidalis является участком встречи и срастания наружной и внутренней стенок marsupium oris. Часть волокон внутренней стенки у крупных экземпляров может проникать в область сустава anguloarticulare и dentale и участвовать в формировании lig. articulodentale lateralis и medialis.
Подвешивающая перепонка суспензориума (membrana suspensorii) (рис. 6) развита довольно слабо. Она служит для крепления начальной области волокон порции А3 m. adductor mandibulae и конечной части волокон m. levator arcus palatini. После снятия кожи головы membrana suspensorii имеет вид узкого сухожильного промежутка между двумя этими мускула. Целиком она открывается лишь после удаления их волокон. Membrana suspensorii представляет собой плёнку, состоящую из двух слоёв (наружного и внутретреннего) и прикрепляющуюся к cr. ventralis hyomandibulare, который ориентирован передневентрально. Волокна наружного слоя направлены назад и вверх, а внутреннего – вверх и вперёд, таким образом повторяя ориентацию волокон соответствующего мускула, прилегающего к перепонке изнутри либо снаружи. В области передневентрального угла m. levator arcus palatini толщина membrana suspensorii максимальна. На этом участке она охватывает данный мускул изнутри и снаружи, а ниже переходит в состав плёнки (membrana hyomandibularis 1) (рис. 4а), затягивающей небольшое отверстие треугольной формы между задним краем metapterygoideum и передним краем hyomandibulare. Передний участок membrana hyomandibularis 1 и ростральные волокна membrana adductores участвуют в формировании пластины начального апоневроза одного из срединных подразделений порции А1б.
Около ½ наружной поверхности порции А3, которая скрывается под массой волокон порций А1а, А2 и подразделений порции А1б, латерально затянута membrana adductores (рис. 4а, 6). Эта плёнка начинает формироваться вентрально посредством сливающихся друг с другом веерообразно расположенных пучков соединительнотканных волокон. Ориентация пучков в направлении снизу вверх постепенно меняется с дорсоростральной на переднекаудальную; при этом толщина плёнки нарастает снизу вверх. По достижении треугольника membrana hyomandibularis 1 membrana adductores включается в её состав. Линия прикрепления membrana adductores каудально тянется по поверхности hyomandibulare параллельно и ниже cr. ventralis, а далее − по задней 1/3 наружной стороны metapterygoideum. Впереди (относительно порции А3) полоска крепления плёнки проходит вдоль передней 1/3 metapterygoideum. Передняя и задняя линии прикрепления membrana adductores приблизительно параллельны друг другу. Волокна порции А3 не крепятся к внутренней поверхности membrana adductores.
Membrana hyomandibularis 2 (рис. 3б) затягивает отверстие в плоскости суспензориума между нижним краем symplecticum, quadratum, передним краем praeoperculum и концом вентральной ветви hyomandibulare.
Membrana hyomandibularis 3 закрывает отверстие, имеющееся между pr. cap. articularis anterior и posterior hyomandibulare и нижним краем fossa dilatator operculi, который сформирован sphenoticum и pteroticum.
Прозрачная membrana m. epaxialis anterior 2 (рис. 6) снаружи покрывает одноимённый мускул, отделяя его волокна от находящегося латерально m. levator operculi. Толщина плёнки нарастает в направлении сверху вниз. Впереди область её отхождения включает малый участок pteroticum позади места начала m. dilatator operculi и дорсокаудальный угол hyomandibulare. Cверху плёнка прикрепляется к полоске дорсальной поверхности рr. m. levator operculi pteroticum и нижнему краю extrascapulare, сзади – к ростральной стороне posttemporale и переднему краю supracleitrum. Нижняя граница membrana m. epaxialis anterior 2 приблизительно совпадает с вентральной кромкой m. epaxialis anterior 2. В дальнейшем плёнка делится на два толстых и непрозрачных соединительнотканных листа. Один из них выстилает медиальную поверхность жаберной крышки, в том числе внутреннюю сторону m. levator operculi и m. adductor opercula, другой покрывает медиальную поверхность оперкулярной полости и включает в себя тонкий на срезе тимус и ткань оперкулярной железы рыбы.
Ниже при описании связок, если не упомянуто обратное, подразумевается, что все они имеют блеск.
Lig. frontale-palatinum (рис. 3а) – прочная, нетянущаяся и неблестящая связка, которая при взгляде на череп сбоку следует вниз и вперёд. Зона начала этой связки включает передний край frontale в области границы с mesethmoideum и ectopterygoideum, затем её волокна спускаются вдоль ростральной поверхности ectoethmoideum, проходя медиальнее f. r. ophthalmicus superficialis (VII) и f. olfactorius, и оканчиваются на верхнем крае и дорсальном участке медиальной стороны задней половины пальцевидного отростка (pr. digitatus) palatinum (рис. 3б, 4б).
Толстое lig. ectoethmoideum-palatinum 1 (рис. 4) начинается на передненижней стороне pr. preorbitalis ectoethmoideum, отделённой от f. olfactorius костным гребешком, и оканчивается на малом участке латеральной стороны и на заднем крае palatinum в основании pr. digitatus. Дорсомедиальное направление данной связки совпадает с ориентацией описываемой поверхности pr. preorbitalis. Часть волокон непрерывны с таковыми lig. frontale-palatinum.
Вентральный фрагмент поверхности pr. preorbitalis ectoethmoideum формирует ямку для отхождения короткой lig. ectoethmoideum-palatinum 2. При взгляде на нейрокраниум спереди эта связка ориентирована вниз и медиально, а при взгляде сбоку – вниз и вперёд. На поперечном срезе она имеет значительную толщину и оканчивается на заднем крае palatinum сразу же ниже области прикрепления lig. ectoethmoideum-palatinum 1.
Lig. ethmopalatinum 1 уплощено в парасагиттальной плоскости. Связка направлена вверх и вперёд, оканчиваясь на медиальной поверхности pr. digitatus, ниже его продольного гребня. Размер и толщина крепкого lig. ethmopalatinum 2 сходны с таковыми lig. ethmopalatinum 1. Lig. ethmopalatinum 2 направлено вентрокаудально и также уплощено в парасагиттальной плоскости. Участок его окончания расположен на медиальной поверхности суспензориума, на границе palatinum и entopterygoideum.
Очень короткое lig. articulodentale lateralis (рис. 4) имеет значительную площадь поперечного сечения. Волокна связки соединяют латеральную поверхность верхней части pr. dorsolateralis ascendens, начиная с его переднего края, с медиальной стороной переднего отдела pr. coronoidalis dentale, который накладывается на anguloarticulare сбоку. Небольшого размера lig. articulodentale medialis (рис. 5а), имеющее большую толщину, ориентировано дорсовентрально; начинается с малого участка на внутренней стороне верхней части pr. dorsolateralis ascendens и оканчивается на ребристой поверхности pr. angulare dentale.
Lig. mandibulare (рис. 3, 4б) ориентировано дорсовентрально, снизу прикрепляется к заднему краю pr. dorsolateralis ascendens, передней головке и шейке quadratum, а дорсально расходится веерообразно и постепенно исчезает в составе соедительнотканного слоя кожи головы. Глубинные волокна связывают только anguloarticulare и quadratum.
Lig. maxillo-dentale (рис. 4а) − широкий тяж, прочно соединяющий наружную поверхность вентральной области pr. alveolaris lateralis praemaxillare и manubrium maxillare c задним краем pr. coronoidalis dentale. Данная связка расположена медиальнее внутренней стенки marsupium oris. Стоит отметить, что нижние отделы рr. alveolaris lateralis praemaxillare и manubrium maxillare крепко связаны между собой короткими соединительнотканными волокнами, а латеральная поверхность pr. coronoidalis dentale обита прочной, у крупных особей охрящевевающей, соединительной тканью. Эти соединительнотканные структуры также расположены медиально относительно внутренней стенки marsupium oris.
Нетянущееся, без блеска lig. interoperculomandibulare (рис. 3б, 4б, 5) несильно развито. Соединяет передний конец interoperculum с обширной вентральной поверхностью retroarticulare.
Lig. hyointeroperculare – небольшая группа волокон, отходящих от гребня на латеральной поверхности epihyale непосредственно ниже сустава этой кости с interhyale и веерообразно оканчивающихся на костном выступе (pr. articularis) в центре медиальной стороны interoperculum.
Сr. lateralis maxillare (рис. 3б) служит началом трёх скоплений блестящих соединительнотканных волокон, которые, однако, имеют разные места своего окончания, включаясь в состав других соединительнотканных образований или же скрепляя maxillare с другими костями черепа: lig. integumentum anterior et posterior, а также lig. maxillolacrimale. Границы этих связок лучше дифференцированы у более крупных экземпляров.
Lig. integumentum anterior (рис. 3а, 4б) – ростральная группа волокон, которые тянутся вверх и медиально и сливаются с пучком волокон связки контрлатеральной стороны для образования переднего края соединительнотканного чехла максиллярного аппарата (integumentum apparatus maxillaris).
Сзади lig. integumentum anterior взаимно переходит в веерообразное lig. integumentum posterior (рис. 3а). Последняя связка формирует латерально ту область integumentum apparatus maxillaris, которая покрывает сверху подушку из желеобразной ткани над pr. digitatus palatinum каждой стороны головы рыбы. Заднедорсальные волокна lig. integumentum posterior присоединяются к переднему отделу nasale, а также частично непрерывны с lig. maxillolacrimale.
Самое каудальное lig. maxillolacrimale (рис. 3а) скрепляет maxillare с ростродорсальным краем lacrimale. Часть волокон этой связки устремляется на медиальную сторону lacrimale и сверху нависает над lig. palatolacrimale, частично включаясь в его состав.
Очень прочное, толстое, неблестящее lig. palatolacrimale (рис. 3б) соединяет латеральную поверхность срединной области pr. digitatus palatinum и участок медиальной стороны lacrimale. При взгляде на нейрокраниум спереди связка направлена латерально и немного вниз. После удаления lacrimale видно, что участок крепления lig. palatolacrimale на lacrimale находится непосредственно в районе выемки maxillare при переходе тела кости в pr. lacrimalis.
ОБСУЖДЕНИЕ
Процесс питания рубиново-пепельной рыбы-попугая включает многократное выполнение силовых укусов кормового субстрата, что привело к уникальному изменению конструкции и особенностей работы висцерального аппарата животного, в частности систем его передних челюстей и суспензориума. Строение данных отделов аппарата питания в значительной степени взаимосвязано с морфологией и функционированием самого крупного и сложно устроенного мускула спланхнокраниума − m. adductor mandibulae. Высокий уровень перистости этого мускула и большой объём пространства, занимаемый им на суспензориуме, сближают рубиново-пепельную рыбу-попугая с хищными видами Teleostei, использующими в качестве главного способа кормодобывания как укус (ромбовидная пиранья Serrasalmus rhombeus (Grubich et al., 2012) и морской угорь Conger conger (Eagderi, Adriaens, 2010)), так и всасывание (клариевый сом Clariallabes longicauda (Wassenbergh et al., 2004)). Однако это сходство m. adductor mandibulae у упомянутых видов и объекта нашего исследования имеет под собой различную трофическую основу. Полученные нами данные позволяют сделать вывод о том, что мускул рубиново-пепельной рыбы-попугая совмещает в себе способность развивать большую силу сокращения наряду с возможностью осуществлять тонкую регулировку степени приведения нижней челюсти и ретракции maxillare, которая имеет место после его выдвижения в ходе работы максиллярного аппарата. Реализации этого функционального потенциала m. adductor mandibulae способствует наличие внутринижнечелюстного сустава (IMJ) между anguloarticulare и dentale. Это сочленение относится к категории расширяющих амплитуду раскрытия передних челюстей у коралловых рыб, использующих укус в качестве основного способа кормодобывания, в отличие от внутринижнечелюстного сустава рыб-ангелов (Pomacanthidae), ограничивающего размер ротового отверстия (Konow et al., 2008). В результате dentale рубиново-пепельной рыбы-попугая получило способность к самостоятельным движениям относительно anguloarticulare. Взаимная подвижность этих костей сделала невозможным присутствие единой порции Аw m. adductor mandibulae, которая у Teleostei обычно заполняет внутреннее пространство единой нижней челюсти (Winterbottom, 1974). По мнению Иорданского (2008), основной функцией порции Аw является распределение нагрузки, исходящей со стороны верхнечелюстных порций m. adductor mandibulae, по медиальной поверхности нижней челюсти, что способствует повышению устойчивости системы висцерального аппарата в целом. Развитие у рубиново-пепельной рыбы-попугая специализированной двучленной структуры нижней челюсти привело к биомеханически выгодному смещению областей нагрузки, прикладываемых к ней верхнечелюстными порциями m. adductor mandibulae, относительно расположения этих областей на dentale и anguloarticulare у генерализованных представителей Teleostei (Winterbottom, 1974) без ущерба для прочности конструкции челюсти. В рамках вышеописываемой системы dentale играет роль рычага первого рода, а верхний конец pr. dorsolateralis ascendens anguloarticulare становится точкой опоры, вокруг которой происходят повороты рычага. Для осуществления контроля движений плеч этого рычага нижний отдел m. adductor mandibulae претерпевает значительную дифференцировку. Он формирует несколько порций, сокращение которых оказывает противоположный эффект, закрывая или открывая dentale. Наличие такого рычажного механизма позволяет уменьшить нагрузку, оказываемую на anguloarticulare и, следовательно, на нижнечелюстной сустав во время питания, в сравнении с генерализованными представителями Teleostei, такими как сёмга Salmo salar (Громова, Махотин, 2016). Кроме того, по-видимому, наличие внутринижнечелюстного сустава облегчает поддержание челюстей в приоткрытом состоянии за счёт опускания dentale относительно anguloarticulare во время периодов быстрого передвижения животного с распахнутым ртом в поисках источников пищи (Clements, Bellwood, 1988; Alfaro et al., 2001). Такое поведение позволяет рыбе начать укус субстрата с более выгодного положения уже частично отведённой нижней челюсти. Выполнению этого действия также способствует поднятие нейрокраниума путём сокращения гипертрофированного m. epaxialis dorsalis. Судя по положению относительно найденной нами os coronomeckeli, spina medialis, по-видимому, является окостеневшим c. meckeli, который вследствие разделения челюсти на две части вторично потерял связь с dentale (рис. 2). Благодаря изменению типичной для большинства лучепёрых рыб (Actinopterygii) каудоростральной ориентации (Diogo, Abdala, 2010) на вентродорсальную, а также с приобретением жёсткости своей структуры spina medialis стал выгодным местом для крепления антагонистических порций Awa и Awb в сравнении с исходной единой порцией Аw. Для уточнения полученных данных требуются дополнительные исследования развития нижней челюсти в раннем онтогенезе у объекта нашей работы.
Стоит отметить, что внутринижнечелюстной сустав рубиново-пепельной рыбы-попугая не является типичным (Нейматов, Сабинин, 2012) вследствие наличия специфических особенностей конструкции. Медиальная и латеральная поверхности pr. dorsolateralis ascendens, которые являются основными областями контакта с dentale, связаны с последним только lig. articulodentale lateralis et medialis и не формируют гладких, покрытых слоем хряща сочленовных поверхностей (рис. 4б, 5а). Описываемое место взаимодействия anguloarticulare и dentale не имеет суставной сумки или менисков. Lig. articulodentale lateralis et medialis обеспечивают упругую мягкость поворотов dentale относительно anguloarticulare и поддерживают целостность этого сустава. Также они задействуются в ходе открывания или закрывания рта при перемещениях anguloarticulare: во время отведения этой кости натягивается lig. articulodentale lateralis, а в ходе её приведения − lig. articulodentale medialis. В обоих случаях это приводит к передвижению dentale в соответствующем направлении. По-видимому, подобный контроль смещений dentale с помощью связок обеспечивает расширение спектра возможностей тонкого воздействия на изменение местоположения этой кости относительно anguloarticulare в сравнении с вариантом сустава, который обладает фиксированным числом осей поворотов, предполагающим бóльшую степень стандартизированности движений (Hollister et al., 1992).
Вследствие расширения спектра объектов питания с увеличением размера и возраста животного, рубиново-пепельной рыбе-попугаю необходимо регулировать силу закрывания нижней челюсти. Для этого у неё развивается уникальная система тонкого контроля степени приведения dentale, состоящая из трёх специализированных для этой цели порций m. adductor mandibulae. Эти порции формируют ряд последовательно расположенных мускульных образований, которые постепенно уменьшают размер и длину волокон по мере приближения к внутринижнечелюстному суставу. В ростральном направлении конусовидная форма этих порций с расширенным на суспензориуме основанием понемногу сменяется на пучкообразную. Самую значительную силу сокращения из них может развивать порция А2. За счёт механически выгодного крепления к самой каудальной точке pr. coronoidalis dentale (т.е. к самой отдалённой от оси рычага точке плеча dentale) эта порция является наиболее эффективным аддуктором. Далее в порядке уменьшения силы и амплитуды сокращения следуют порции А3а и Аwb. По мере приближения к внутринижнечелюстному суставу эффективность работы этих порций в качестве аддукторов уменьшается (рис. 4б, 5а). Таким образом, наличие данного механизма регулировки движений dentale является морфологическим свидетельством, иллюстрирующим способность рубиново-пепельной рыбы-попугая модулировать характеристики укуса в процессе питания.
Система порций m. adductor mandibulae попугаевых копателей характеризуется большей степенью дифференцировки и их большей мускульной массой в сравнении с таковыми у видов-соскребателей (Bellwood, 1985). В ходе работы мы обнаружили ранее не отмеченные особенности строения описываемого мускула у рубиново-пепельной рыбы-попугая, которые могут послужить структурной основой, позволяющей данному представителю питаться по способу видов из группы копателей. Сложно устроенный апоневротический каркас, образующий внутреннюю структуру порций А1а, А1б и А2 m. adductor mandibulae рубиново-пепельной рыбы-попугая, способствует повышению разнообразия функциональных результатов, возникающих вследствие сокращения различных областей в рамках массы волокон внушительного по размеру сложноперистого мускула (Herring et al., 1979). В то же время наличие областей взаимного перехода между этими порциями позволяет рыбе в случае необходимости включать в процесс сокращения более обширную долю мускульных волокон в сравнении с принадлежащими лишь одной из порций, тем самым многократно увеличивая силу сокращения. Неглубокий единый поверхностный слой порций А1а + А2 смещает друг к другу каудальные концы manubrium maxillare и pr. coronoidalis dentale после их отведения в процессе открывания рта, тем самым возвращая челюсти в состояние покоя и стабилизируя относительно друг друга (рис. 3а). Наличие двух отдельных пластин апоневрозов у порции А1а и А2 позволяет дифференцировать их в качестве отдельных образований, в отличие от мнения Белвуда (Bellwood, 1985), который объединял эти порции в одну. Детальное анатомирование дало нам возможность уточнить особенности соединений данных апоневрозов между собой и marsupium oris. Наличие таких морфологических связей говорит в пользу необходимости поддержания структурного единства этих трёх основных соединительнотканных элементов в области важного биомеханического узла составляющих висцерального аппарата в углу рта животного. Обращает на себя внимание многочленность конечных сухожильных пучков апоневрозов порций А1а и А2, оканчивающихся соответственно на maxillare и dentale. Такое устройство свидетельствует о приспособлении этих сложноперистых мускульных образований к созданию большой силы сокращения, а также о способности осуществлять динамический контроль степени сокращения путём задействования разного числа конечных сухожилий в рамках их многочленного пучка.
Порция А1б рубиново-пепельной рыбы-попугая имеет схожее строение с таковой у попугаевых копателей, т.е. состоит из нескольких последовательно расположенных подразделений, что не было отмечено предшествующими авторами (Bellwood, 1985; Clements, Bellwood, 1988). Гребни, проходящие по поверхности entopterygoideum, наличие пластин начальных апоневрозов в составе ростральных подразделений, а также многочленность пучка их конечных сухожилий по типу строения такового у порций А1а и А2 свидетельствуют о подобии функциональных возможностей порции А1б с соответствующими им у вышеупомянутых порций. Обращает на себя внимание тот факт, что объём мускульных волокон порций А1а и А1б, управляющих движениями maxillare (с оговоркой касательно порции А1б), приблизительно можно соотнести с суммарным объёмом порций А2, А3, A3a, Аwa, Awb, обсуживающих нижнюю челюсть. В отличие от этого у других видов Teleostei, использующих укус во время питания, имеется иная ситуация (Eagderi, Adriaens, 2010). У последней группы рыб практически вся масса волокон m. adductor mandibulae вовлечена в процесс закрывания нижней челюсти, тогда как maxillare остаётся неподвижным и не принимает активного участия в захвате добычи (Grubich et al., 2012). Небольшая степень подвижности максиллярного аппарата у рубиново-пепельной рыбы-попугая способствует работе расширяющего размер ротового отверстия механизма внутринижнечелюстного сустава. Возможность значительного выдвижения praemaxillare привела бы к снижению устойчивости конструкции аппарата питания в условиях повышенных нагрузок. Когда в процессе укуса режущий край praemaxillare проникает в кормовой субстрат, pr. ascendens praemaxillare смещается вверх и назад по поверхности этмоидального отдела нейрокраниума, а manubrium maxillare вместе с pr. alveolaris praemaxillare поворачиваются вперёд и вверх. В связи с этим рубиново-пепельной рыбе-попугаю необходимо удерживать траекторию поворота manubrium maxillare в строго определённом положении в зависимости от требуемой степени открывания рта, затрачивая при этом значительную силу, чтобы не дать этой кости вылететь из суставов с нейрокраниумом, palatinum и praemaxillare под воздействием большой нагрузки, оказываемой на неё со стороны субстрата (maxillare управляет движением praemaxillare (Alexander, 1967)). Таким образом, уникальная черта аппарата питания объекта нашего исследования состоит в том, что maxillare у него принимает активное динамически регулируемое участие в выполнении силовых укусов субстрата в сравнении с его пассивной ролью у других представителей Teleostei, использующих укус в качестве основного способа кормодобывания (Eagderi et al., 2016; Meyer et al., 2018). Особенно важным становится удержание maxillare путём сокращения порций А1а и А1б в ходе «заякоривания» premaxillare в грунте, когда укус осуществляет одна нижняя челюсть; подобный метод питания попугаевых копателей может использовать и рубиново-пепельная рыба-попугай (Bellwood, Choat, 1990). Стоит отметить, что значительная степень дифференцировки и обширная масса m. adductor mandibulae данного вида конвергентно схожи с таковыми у представителей другой специализированной группы рыб − иглобрюхообразных (Tetraodontiformes) (Turingan, Wainwright, 1993). Наличию этого морфологического подобия способствует дурофагия, характерная для рыб этой группы, однако спектр объектов их питания шире, чем у попугаевых (Bruggemann et al., 1996; Ong, Holland, 2010), и включает в основном различных беспозвоночных (Moyer, Sano, 1987; Krumme et al., 2007; Hammoud, Salama, 2016). Обслуживание maxillare порциями А1а и A1b (помимо многочисленных порций, управляющих нижней челюстью) у иглобрюхообразных связано с необходимостью сочетания большой силы схватывания добычи со способностью их ротовых челюстей манипулировать жертвой и осуществлять её обработку (Turingan, Wainwright, 1993; Friel, Wainwright, 1999). У рубиново-пепельной рыбы-попугая, напротив, пища после захвата сразу же проглатывается и направляется в аппарат глоточных челюстей, осуществляющий её перемалывание (Андрияшев, 1941, 1944, 1946, 1948б; Громова, Махотин, 2020).
Дифференцировка каудальных подразделений порции А1б связана с развитием у данного вида уникального механизма приведения anguloarticulare с использованием пластины апоневроза порции А1а (рис. 4а). Сокращение этих подразделений порции А1б вызывает подтягивание ap. portio A1a в дорсокаудальном направлении, что служит причиной натяжения t. portio A1a № 1: следствием этого процесса является аддукция anguloarticulare. Описываемое конечное сухожилие прикрепляется очень близко к челюстному суставу, поэтому суммарный миовектор каудальных подразделений проходит почти через ось его вращения, делая их сокращение малоэффективным (Иорданский, 1990). Приемлемым решением этой проблемы стало развитие порции А3 с подобной же точкой окончания t. portio A3 на anguloarticulare, но с экстремально увеличенным плечом своего миовектора, которое подходит к оси anguloarticulare практически под прямым углом, делая всю силу сокращения этой порции вращательной; следовательно, последняя работает только на аддукцию рычага данной кости (Громова и др., 2017). Кроме того, мы включаем в систему тонкой регулировки степени закрывания anguloarticulare и fasciculus portio A3, маленький размер которого свидетельствует в пользу его способности приводить эту кость с наименьшей в сравнении с другими порциями m. adductor mandibulae силой. Наличие связи этого пучка при помощи сухожильного промежутка с порцией А3а (рис. 4б) говорит о возможности сокращения этих образований в качестве частей единого двубрюшного мускула в целях повышения скорости закрывания рта рыбы (Иорданский, 1990). Таким образом, anguloarticulare имеет индивидуальную систему приведения различными компонентами m. adductor mandibulae, отличную от таковой для dentale; в процессе питания эти две системы аддукции элементов нижней челюсти вступают между собой во взаимодействие.
Нижнечелюстной сустав рубиново-пепельной рыбы-попугая во время совершения укуса подвергается значительной нагрузке, вследствие чего его целостность может быть нарушена. Эта ситуация, возникающая в процессе кормодобывания данного вида, конвергентно схожа с той, что имеет место в ходе работы челюстных суставов таких хищных представителей Teleostei как, например, сёмга (Громова, Махотин, 2016) и судак Stizostedion lucioperca (Elshoud-Oldenhave, 1979). Энергичные толчки вырывающейся добычи способны заставить нижнюю челюсть этих видов рыб сместиться вперёд и вниз и таким образом разобщить сочленовные поверхности, образующие челюстной сустав. Во избежание этого задняя доля anguloarticulare формирует хорошо развитый ретроартикулярный отросток (pr. postarticularis), предохраняющий челюстной сустав сзади. Также такое приспособление позволяет ограничить степень открывания нижней челюсти лишь до определённого предела: пока ретроартикулярный отросток не встретит упор quadratum. В отличие от этого в процессе питания нижняя челюсть рубиново-пепельной рыбы-попугая широко не отводится, поэтому этот вид лишён подобного механизма предотвращения высокоамплитудного открывания рта: ретроартикулярный отросток редуцирован и не обхватывает челюстной сустав сзади. Рr. postarticularis внедряется в челюстной сустав и приобретает специализированную функцию ограничения слишком сильного приведения anguloarticulare, которое может произойти вследствие работы мощного комплекса m. adductor mandibulae. В конце траектории поворота данной кости в процессе аддукции ретроартикулярный отросток, словно ключ, вставляется в специальную полость quadratum (cavum claustrum), обеспечивая замыкание “замочного механизма”. Также замыкание происходит в рамках латерального мыщелка нижнечелюстного сустава: дорсальный край planum lateralis portio ventralis аnguloarticulare встречает упор козырька (tectum) вентрального края planum lateralis portio dorsalis quadratum (рис. 2). (Хасселт (Hasselt, 1979) отмечает наличие у некоторых представителей лабриновых (Labrinae) клинообразного отростка quadratum, однако, упоминая его в тексте работы, не показывает его на изображениях данной кости; рисунки anguloarticulare также отсутствуют.) Для уточнения валидности соответствия pr. clavis и рr. postarticularis других Teleostei необходимо исследовать развитие данного костного образования у нашего вида попугаевых в ходе раннего онтогенеза.
М. levator operculi является основным мускулом, который опускает нижнюю челюсть рубиново-пепельной рыбы-попугая, поскольку для данного вида приемлема небольшая степень открывания рта (Громова, Махотин, 2020). Мера отведения нижней челюсти контролируется lig. mandibulare. Кроме того, в осуществление её абдукции при помощи m. levator operculi могут вносить вклад m. sternohyoideus и m. geniohyoideus (рис. 5), сокращение которых будет приводить dentale к anguloarticulare; подробности этого процесса описаны нами ранее (Громова, Махотин, 2019). Ввиду того, что контралатеральные половинки suffusorium dentale скреплены прочным швом, исключающим подвижность между ними, значение первоначальной роли m. intermandibularis – прижимать dentale обеих сторон головы друг к другу – снижается. Однако этот мускул сохраняется, поскольку приобретает новую функцию – его сокращение формирует упругий передний предел, ограничивающий то небольшое смещение гиоида вперёд, которое возникает в ходе работы m. geniohyoideus. О взаимосвязанности сокращений m. intermandibularis и m. geniohyoideus говорит наличие между ними сухожильного промежутка (рис. 5б). Помимо m. sternohyoideus и m. geniohyoideus функционирование порции Аwа прижимает suffusorium dentale к anguloarticulare. В результате эта порция способствует открыванию рта рыбы, в отличие от остальных шести порций m. adductor mandibulae, закрывающих нижнюю челюсть. Небольшой размер порции Аwa можно связать с тем, что dentale под её воздействием в процессе перемещения не преодолевает какой-либо нагрузки, кроме сопротивления воды. Кроме того, уменьшению массы мускульных волокон способствует выгодность точек крепления порции на костях, увеличивающая эффективность её работы в качестве абдуктора.
М. levator arcus palatini, m. dilatator operculi и m. levator operculi вторично теряют границы между собой, формируя единый комплекс, отводящий суспензориум и жаберную крышку (рис. 6а). Данное устройство конвергентно напоминает слияние отдельных мускулов, имеющееся у более примитивных рыб, например у сёмги, которое у неё реализуют m. adductor arcus palatini, m. adductor hyomandibularis и m. adductor operculi (Громова, Махотин, 2016). Причиной такого объединения служит необходимость повышения силы сокращения, достигаемая за счёт увеличения массы мускульных волокон, входящих в состав такого единого сложноперистого мускула. Слабое развитие membrana suspensorii у рубиново-пепельной рыбы-попугая в сравнении с сёмгой (Громова, Махотин, 2016) или серебряной араваной Osteoglossum bicirrhosum (Громова и др., 2017) свидетельствует об отсутствии значимого взаимодействия между m. levator arcus palatini и m. adductor mandibulae (посредством его порции А3). Многочисленные апоневротические рёбра, которыми насыщен абдукторный комплекс рубиново-пепельной рыбы-попугая, способствуют статической функции изометрического сокращения данного сверхмускула (Иорданский, 1990). Стоит отметить, что результатом работы волокон m. dilatator operculi, начинающихся с hyomandibulare, будут повороты operculum относительно последней кости; по-видимому, это связано с необходимостью работы жаберных крышек рыбы в латеромедиальном направлении при проточном способе дыхания. Компактный размер описываемого абдукторного комплекса рубиново-пепельной рыбы-попугая свидетельствует в пользу увеличения роли m. geniohyoideus и m. sternohyoideus в процессе расширения ротовой полости, который происходит в основном в её вентральном отделе (Громова, Махотин, 2019).
Обращает на себя внимание значительное развитие подразделений эпаксиальной мускулатуры, осуществляющей соединение верхнего отдела плечевого пояса с нейрокраниумом − m. epaxialis anterior 2 и m. levator pectoralis. Увеличение массы их волокон требует расширения областей прикрепления этих мускулов на черепе, которое в случае m. epaxialis anterior 2 включает даже hyomandibulare. Это означает, что латеромедиальные отклонения данной кости в процессе питания рыбы не столь велики, чтобы помешать функционированию m. epaxialis anterior 2. Гипертрофия мускулатуры, связывающей плечевой пояс и нейрокраниум у рубиново-пепельной рыбы-попугая, конвергентно схожа с той, что присутствует у донных видов Teleostei, у которых мощно развитые грудные плавники используются для передвижения по дну, например, у белоносого свинорыла Perryena leucometopon (Honma et al., 2013) и кораллового ползуна-пигмея Caracanthus unipinna (Shinohara, Imamura, 2005). Однако это подобие является следствием разных причин. Значительное развитие m. epaxialis anterior 2 и m. levator pectoralis у рубиново-пепельной рыбы-попугая обусловлено необходимостью создания плечевым поясом прочного упора для сочленяющейся с ним нижней глоточной челюсти (Bellwood, 1985). Поддержание целостности сустава плечевого пояса с нижней глоточной челюстью имеет важное значение для обработки корма в аппарате глоточных челюстей у данного вида (Vandewalle et al., 2000). Кроме того, мы полагаем, что укрепление сочленения плечевого пояса с нейрокраниумом способствует стабилизации конструкции черепа у рыб с лабриформным типом локомоции, совершающих при плавании взмахивающие движения плавников, при которых плечевой пояс подвергается более значительным нагрузкам, чем при гребущих движениях (Thorsen, Westneat, 2005).
Устройство висцерального аппарата Teleostei предполагает присутствие в нём ряда входных механизмов, действие которых запускает цепь кинематических событий, в итоге приводящих в движение различные компоненты его конструкции (выходные механизмы) (Konow, Sanford, 2008). Так, в максиллярном аппарате изменения положения praemaxillare обусловлены перемещениями maxillare, в свою очередь находящегося под влиянием движений суспензориума. На основе исследованного нами морфологического материала мы можем заключить, что функционирование суспензориумов у рубиново-пепельной рыбы-попугая отличается от такового, присущего более генерализованным окунеобразным, например судаку (Elshoud-Oldenhave, 1979) или окуню Perca fluviatilis (Osse, 1969). Обращает на себя внимание значительное различие в способе крепления задней и передней областей суспензориума к нейрокраниуму у объекта нашей работы. Сap. articularis posterior hyomandibulare причленяется к pteroticum малоподвижно, поскольку амплитуда поворотов, предоставляемая сочленовной впадиной на pteroticum, невелика. Задний угол hyomandibulare прочно прикрепляется к pr. levator pectoralis pteroticum. В отличие от этого обширная внутренняя поверхность передней сочленовной впадины для hyomandibulare, формируемая sphenoticum и pteroticum, свидетельствует о возможности в ней большего объёма движений (передняя впадина ориентирована перпендикулярно по отношению к задней). Ростральный отдел суспензориума в процессе питания подвергается значительным нагрузкам, однако он не образует сколь бы то ни было прочного и устойчивого сустава c нейрокраниумом, который имеется у других видов Teleostei, использующих укус в качестве основного способа кормодобывания (Elshoud-Oldenhave, 1979). Palatinum рубиново-пепельной рыбы-попугая соприкасается лишь с маленьким участком ectoethmoideum, формируя там плоское скользящее сочленение, допускающее неограниченные возможности перемещений в разных направлениях. Эффективное функционирование нескольких суставов с разной мерой мобильности в рамках единого комплекса суспензориума свидетельствует о присутствии некоторой степени самостоятельной подвижности составляющих его костных элементов, благодаря чему повышается вариабельность результатов работы связанных с ним систем аппарата питания рыбы, таких как максиллярный механизм. При манипуляциях с синдесмологическиими препаратами голов рубиново-пепельной рыбы-попугая было обнаружено, что передний отдел суспензориума, включающий palatinum, quadratum, ecto-, ento- и metapterygoideum, может отгибаться относительно hyomandibulare, поскольку две последних кости не формируют неподвижного шва и связаны лишь при помощи соединительной ткани; hyomandibulare тоже способно к небольшим поворотам вокруг своей продольной оси. Присутствие подобных смещений отдельных составляющих суспензориума по отношению к другим было найдено у ряда видов: карповых (Cyprinidae) – карпа Cyprinus carpio (Ballintijn et al., 1972) и белого толстолобика Hypophthalmichthys molitrix (Gromova, Makhotin, 2018), кифозовых (Kyphosidae) − гиреллы Girella laevifrons (Vial, Ojeda, 1990), губановых (Labridae) − радужного губана Labrus berggylta (Hasselt, 1979) и в значительной степени у большеротого губана Epibulus insidiator (Westneat, Wainwright, 1989). У рубиново-пепельной рыбы-попугая возможность этих сдвигов обеспечивается тем, что передний и задний (hyomandibulare) отделы суспензориума обслуживаются целиком разными мускулами, которые являются антагонистами друг друга – гипертрофированным m. adductor arcus palatini и небольшим m. levator arcus palatini. Стоит подчеркнуть, что области окончания описываемых мышц на костях у рубиново-пепельной рыбы-попугая не пересекаются, тогда как в висцеральном аппарате других Teleostei кости имеют двойной контроль абдуктором и аддуктором во избежание возникновения областей механической нестабильности в суспензориуме (Kirchhoff, 1958; Elshoud-Oldenhave, 1979; Громова и др., 2017). Помимо этого при взгляде на голову рубиново-пепельной рыбы-попугая спереди заметно, что palatinun, ectopterygoideum и передняя доля quadratum изогнуты латерально относительно плоскости суспензориума, тогда как hyomandibulare ориентировано, напротив, медиально. Направления небольших перемещений, которые может осуществлять подвижный передний отдел суспензориума у объекта нашего исследования, ясно определяются ориентацией связок, окружающих плоский сустав palatinum и ectoethmoideum (Anker, 1974). Это – сдвиги в парасагиттальной плоскости вверх и вперёд, чья амплитуда ограничивается lig. ethmopalatinum 1, и противоположные им передвижения вниз и назад, ограничиваемые lig. ethmopalatinum 2. Смещения palatinum вверх и медиально, возникающие в процессе сокращения мощного m. adductor arcus palatini, находятся под контролем lig. ectoethmoideum-palatinum 1. Сдвиги вперёд, вниз и медиально ограничены lig. ectoethmoideum-palatinum 2, а lig. frontale-palatinum препятствует излишнему перемещению данной кости ростровентрально. Подобные движения palatinum небольшой амплитуды, происходящие в ходе абдукции и аддукции суспензориумов, были также описаны для атлантической трески Gadus morhua (Petersen, 1914) и трёхиглой колюшки Gasterosteus aculeatus (Kampf, 1961; Anker, 1974). Возможно, в случае рубиново-пепельной рыбы-попугая мы имеем дело с кинематической моделью, подобной той, что была предложена Хасселтом (Hasselt, 1979) для описания работы висцерального аппарата губановых, в рамках которой конструкция с небольшим входным движением (суспензориум) обусловливает функционирование выходного механизма (передние челюсти) с более высокоамплитудными перемещениями. Однако строение спланхнокраниума объекта нашего исследования характеризуется рядом морфологических особенностей, отличающих его от видов губановых (Hasselt, 1979). Так, lacrimale рубиново-пепельной рыбы-попугая, прочно сочленяющееся с pr. praeorbitalis сзади и жёстко скреплённое мощным lig. palatolacrimale с pr. digitatus palatinum спереди, формирует небольшую суставную поверхность с медиальной стороной pr. lacrimalis maxillare и будет ограничивать его повороты, а значит, и всей этой кости вокруг её продольной оси, способствуя осуществлению лишь качательных движений (Alexander, 1967). Выдвижение верхней челюсти и значение в этом процессе движений суспензориума у данного вида попугаевых будут более подробно описаны в нашей следующей публикации, посвящённой работе максиллярного аппарата.
ЗАКЛЮЧЕНИЕ
Висцеральный аппарат рубиново-пепельной рыбы-попугая демонстрирует глубокую степень адаптации своей конструкции к многократному совершению укусов, выполняемых животным в процессе питания. Спланхнокраниум данного вида характеризуется значительным уровнем дифференцировки мускулатуры, сложное устройство которой представляет собой фундамент для ранее выявленной значительной степени индивидуальной изменчивости процесса кормодобывания у представителей Scaridae. Однако некоторые мускулы вторично утрачивают границы между собой в пользу достижения возможности развивать увеличенную мощность своего сокращения (абдукторный комплекс суспензориума). Подавляющее большинство мускулов спланхнокраниума имеют перистое строение и сложный апоневротический каркас. Гипертрофированный m. adductor mandibulae демонстрирует структурные адаптации, которые свидетельствуют о его способности сочетать большую силу своего действия с тонким контролем движения элементов скелета, что конвергентно напоминает морфофункциональные приспособления того же мускула в висцеральном механизме иглобрюхообразных (Tetraodontiformes). Подобное устройство m. adductor mandibulae позволяет рубиново-пепельной рыбе-попугаю пластично подстраивать работу своего аппарата питания под различные рельефы субстратов на коралловых рифах для эффективного соскабливания с них корма. В строении этого мускула обнаружены признаки, характерные для m. adductor mandibulae видов попугаевых копателей, что предоставляет морфологическую основу для ранее полученных данных о фактах использования такого способа кормодобывания рубиново-пепельной рыбой-попугаем, которая в литературе обычно рассматривается как соскребатель. Обнаруженная нами некоторая степень подвижности костей, составляющих суспензориум, относительно друг друга свидетельствует о том, что функциональные последствия работы такой механической конструкции будут отличаться от тех, что имеют место в результате абдукции и аддукции суспензориумов неспециализированных Acanthopterygii. В работе представлены оригинальные изображения anguloarticulare и quadratum, сочленение которых формирует специфический замочный механизм, ограничивающий степень приведения нижней челюсти.
Список литературы
Андрияшев А.П. 1941. Функционально-морфологическая характеристика глоточных зубов // Рефераты работ учреждений Отд. биол. наук АН СССР за 1940 г. М.; Л.: Изд-во АН СССР. С. 211–212.
Андрияшев А.П. 1944. О методике функционально-морфологического исследования глоточного аппарата костистых рыб // Зоол. журн. Т. 23. Вып. 6. С. 319−329.
Андрияшев А.П. 1945а. Функционально-морфологические упрощения в глоточном аппарате Blennius sanguinolentus Pallas // Рефераты работ учреждений Отд. биол. наук АН СССР за 1941–1943 гг. М.; Л.: Изд-во АН СССР. С. 166–167.
Андрияшев А.П. 1945б. Функциональные особенности глоточного аппарата саргана // Рефераты НИР за 1944 г. Отд. биол. наук АН СССР. М.; Л.: Изд-во АН СССР. С. 114–115.
Андрияшев А.П. 1946. Об упрощениях в строении и функции глоточного аппарата некоторых растительноядных рыб // Зоол. журн. Т. 25. Вып. 4. С. 339–346.
Андрияшев А.П. 1948а. Роль глоточного аппарата в питании кефали // Сб. памяти акад. С.А. Зернова. Л.: Изд-во ЗИН АН СССР. С. 108–112.
Андрияшев А.П. 1948б. Функционально-морфологическая характеристика глоточного аппарата горбыля // Тр. Севастоп. биол. ст. Т. 6. С. 319–323.
Громова Е.С., Махотин В.В. 2016. Функциональная морфология висцерального аппарата сёмги Salmo salar (Salmonidae) // Вопр. ихтиологии. Т. 56. № 4. С. 1−17. https://doi.org/10.7868/S0042875216040068
Громова Е.С., Махотин В.В. 2019. Детали строения и функционирования аппарата глоточных челюстей рубиново-пепельной рыбы-попугая Scarus rubroviolaceus (Scaridae) // Там же. Т. 59. № 6. С. 693−714.
Громова Е.С., Дзержинский Ф.Я., Махотин В.В. 2017. Морфофункциональные особенности висцерального аппарата серебряной араваны Osteoglossum bicirrhosum (Osteoglossidae) // Там же. Т. 57. № 4. С. 379–392. https://doi.org/10.7868/S0042875217040038
Иорданский Н.Н. 1990. Эволюция комплексных адаптаций: челюстной аппарат амфибий и рептилий. М.: Наука, 308 с.
Иорданский Н.Н. 2008. Внутринижнечелюстные мышцы и некоторые проблемы эволюции челюстного аппарата позвоночных // Зоол. журн. Т. 87. № 1. С. 49−61.
Нейматов Э.М., Сабинин С.Л. 2012. Основы биомеханики движения тела. Настольная книга остеопата. М.: Мед. информ. агентство, 480 с.
Ромейс Б. 1953. Микроскопическая техника. М.: Изд- во иностр. лит-ры, 718 с.
Alexander R.McN. 1967. The functions and mechanisms of the protrusible upper jaws of some acanthopterygian fish // J. Zool. V. 151. P. 43−64. https://doi.org/10.1111/j.1469-7998.1967.tb02865.x
Alfaro M., Westneat M.W. 1999. Motor patterns of herbivorous feeding: electromyographic analysis of biting in the parrotfishes Cetoscarus bicolor and Scarus iseri // Brain Behav. Evol. V. 54. P. 205−222. https://doi.org/10.1159/000006624
Alfaro M., Janovetz J., Westneat M.W. 2001. Motor control across trophic strategies: muscle activity of biting and suction feeding fishes // Amer. Zool. V. 41. P. 1266−1279. https://doi.org/10.1093/icb/41.6.1266
Alwany M.A., Thaler E., Stachowitsch M. 2009. Parrotfish bioerosion on Egyptian Red Sea reefs // J. Exp. Mar. Biol. Ecol. V. 371. P. 170 –176. https://doi.org/10.1016/j.jembe.2009.01.019
Anker G.Ch. 1974. Morphology and kinetics of the head of the stickleback, Gasterosteus aculeatus // Trans. Zool. Soc. London. V. 32. P. 311–416. https://doi.org/10.1111/j.1096-3642.1974.tb00030.x
Anker G.Ch. 1978. The morphology of the head-muscles of a generalized Haplochromis species: H. elegans Trewavas 1933 (Pisces, Cichlidae) // Netherl. J. Zool. V. 28. № 2. P. 234–271. https://doi.org/10.1163/002829678X00071
Ballintijn C.M., Burg A., Egberink B.P. 1972. An electromyographic study of the adductor mandibulae complex of a free-swimming carp (Cyprinus carpio L.) during feeding // J. Exp. Biol. V. 57. P. 261−283.
Barel C.D., Witte F., Van Oijen M.J. 1976. The shape of the skeletal elements in the head of a generalized Haplochromis species: H. elegans Trewavas 1933 (Pisces, Cichlidae) // Netherl. J. Zool. V. 26. № 2. P. 163–265.
Bellwood D.R. 1985. The functional morphology, systematics and behavioural ecology of parrotfihes (Scaridae): Ph.D. thesis. N. Queensland, Townsville: James Cook Univ., 489 p.
Bellwood D.R. 1994. A phylogenetic study of the parrotfishes family Scaridae (Pisces: Labroidei), with a revision of genera. Sydney: Austral. Mus., 86 p.
Bellwood D.R. 1996. Production and reworking of sediment by parrotfishes (family Scaridae) on the Great Barrier Reef, Australia // Mar. Biol. V. 125. № 4. P. 795–800. https://doi.org/10.1007/BF00349262
Bellwood D.R., Choat H. 1990. A functional analysis of grazing in parrotfishes (family Scaridae): the ecological implications // Environ. Biol. Fish. V. 28. P. 189–214. https://doi.org/10.1007/978-94-009-2065-1_11
Bonaldo R.M., Bellwood D.R. 2009. Dynamics of parrotfish grazing scars // Mar. Biol. V. 156. № 4. P. 771–777. https://doi.org/10.1007/s00227-009-1129-x
Bonaldo R.M., Krajewski J.P., Bellwood D.R. 2011. Relative impact of parrotfish grazing scars on massive Porites corals at Lizard island, Great Barrier Reef // Mar. Ecol. Progr. Ser. V. 423. P. 223–233. https://doi.org/10.3354/meps08946
Bonaldo R.M., Hoey A.S., Bellwood D.R. 2014. The ecosystem roles of parrotfishes on tropical reefs // Ocean. Mar. Biol. Ann. Rev. V. 52. P. 81–132.
Bruggemann J.H., Van Kessel A.M., Van Rooij J.M., Breeman A.M. 1996. Bioerosion and sediment ingestion by the Caribbean parrotfish Scarus vetula and Sparisoma viride: implications of fish size, feeding mode and habitat use // Mar. Ecol. Progr. Ser. V. 134. P. 59–71. https://doi.org/10.3354/meps134059
Clements K.D., Bellwood D.R. 1988. A comparison of the feeding mechanisms of two herbivorous labroid fishes, the temperate Odax pullus and the tropical Scarus rubroviolaceus // Austal. J. Mar. Freshwat. Res. V. 39. № 1. P. 87–107. https://doi.org/10.1071/MF9880087
Diogo R., Abdala V. 2010. Muscles of vertebrates. Comparative anatomy, evolution, homologies and development. Boca Raton: CRC Press, 500 p.
Eagderi S., Adriaens D. 2010. Head morphology of the duckbill eel, Hoplunnis punctata (Regan, 1915; Nettastomatidae: Anguilliformes) in relation to jaw elongation // Zool. V. 113. № 3. P. 148–157. https://doi.org/10.1016/j.zool.2009.09.004
Eagderi S., Christiaens J., Boone M. et al. 2016. Functional morphology of the feeding apparatus in Simenchelys parasitica (Simenchelyinae: Synaphobranchidae), an alleged parasitic eel // Copeia. V. 104. № 2. P. 421–439. https://doi.org/10.1643/CI-15-329
Elshoud-Oldenhave M.J.W. 1979. Prey capture in the pike-perch, Stizostedion lucioperca (Teleostei, Percidae): a structural and functional analysis // Zoomorphology. V. 93. № 1. P. 1−32. https://doi.org/10.1007/BF02568672
Francini-Filho R.B., Moura R.L., Ferreira C.M., Coni E.O.C. 2008. Live coral predation by parrotfishes (Perciformes: Scaridae) in the Abrolhos Bank, eastern Brazil, with comments on the classification of species into functional groups // Neotrop. Ichthyol. V. 6. № 2. P. 191–200. https://doi.org/10.1590/S1679-62252008000200006
Friel J.P., Wainwright P.C. 1999. Elolution of complexity in motor patterns and jaw musculature of Tetraodontiform fishes // J. Exp. Biol. V. 202. P. 867–880.
Gierse V.A. 1904. Untersuchungen uber das Gehirn und die Kopfnerven von Cyclothone acclinidens // J. Morph. V. 32. P. 602–686.
Goedel V.W. 1974. A contribution to the comparative and functional anatomy of the head of Tilapia (Chichlidae, Teleostei) // Zool. Jb. Anat. V. 92. P. 220−274.
Gromova E.S., Makhotin V.V. 2018. Maxillary apparatus in the feeding of the silver carp Hypophthalmichthys molitrix (Cyprinidae) // J. Ichthyol. V. 58. № 6. P. 857–877. https://doi.org/10.1134/S0032945218060036
Grubich J.R., Huskey S., Crofts St. et al. 2012. Mega-bites: extreme jaw forces of living and extinct piranhas (Serrasalmidae) // Sci. Rept. V. 2. № 1009. P. 1–9. https://doi.org/10.1038/srep01009
Hammoud V.M., Salama L. 2016. Food and feeding habits of the invasive puffer fish Lagocephalus sceleratus (Tetraodontidae) in the Syrian marine waters // J. King Abdul. Univ. Sci. V. 28. № 2. P. 21–30.
Harrison G. 1981. The cranial nerves of the teleost Trichiurus lepturus // J. Morphol. V. 167. № 1. P. 119–134. https://doi.org/10.1002/jmor.1051670111
Hasselt M.J.F.M. 1978. A kinematic model for the jaw movements in some Labrinae (Pisces, Perciformes) // Netherl. J. Zool. V. 28. № 3−4. P. 545–558. https://doi.org/10.1163/002829678X00143
Hasselt M.J.F.M. 1979. Morphology and movements of the jaw apparatus in some Labrinae (Pisces, Perciformes) // Ibid. V. 29. № 1. P. 52–108. https://doi.org/10.1163/002829679X00115
Herring S.W., Grimm A.F., Grimm B.R. 1979. Functional heterogeneity in a multipinnate muscle // Amer. J. Anat. V. 154. № 4. P. 563−576. https://doi.org/10.1002/aja.1001540410
Hollister A., Buford W.L., Myers L.M., Giurintano D.J., Novick A. 1992. The axes of rotation of the thumb carpometacarpal joint // J. Orthop. Res. V. 10. P. 454–460. https://doi.org/10.1002/jor.1100100319
Honma Y., Imamura H., Kawai T. 2013. Anatomical description of the genus Perryena, and proposal to erect a new family for it based on its phylogenetic relationships with related taxa (Scorpaeniformes) // Ichthyol. Res. V. 60. № 2. P. 122–141. https://doi.org/10.1007/s10228-012-0321-z
Howard K.G., Claisse J.T., Clark T.B. et al. 2013. Home range and movement patterns of the Redlip Parrotfish (Scarus rubroviolaceus) in Hawaii // Mar. Biol. V. 160. № 7. P. 1583–1595. https://doi.org/10.1007/s00227-013-2211-y
Kampf W.D. 1961. Vergleichende funktionsanalytische Untersuchungen an den Viscerocranien einiger rauberisch lebender Knochenfische // Zool. Beitr. Neue Folge. V. 6. P. 391–496.
Kirchhoff H. 1958. Funktionell-anatomische Untersuchung des Visceralapparates von Clupea harengus L. // Zool. Jb. Anat. V. 76. P. 462–540.
Konow N., Sanford C.P.J., 2008. Biomechanics of a convergently derived prey-processing mechanism in fishes: evidence from comparative tongue bite apparatus morphology and raking kinematics // J. Exp. Biol. V. 211. P. 3378−3391. https://doi.org/10.1242/jeb.023564
Konow N., Bellwood D.R., Wainwright P.C., Kerr A.M. 2008. Evolution of novel jaw joints promote trophic diversity in coral reef fishes // Biol. J. Linn. Soc. V. 93. № 3. P. 545−555. https://doi.org/10.1111/j.1095-8312.2007.00893.x
Krumme U., Keuthen H., Saint-Paul U., Villwock W. 2007. Contribution to the feeding ecology of the banded puffer fish Colomesus psittacus (Tetraodontidae) in north Brazilian mangrove creeks // Braz. J. Biol. V. 67. № 3. P. 383–392. https://doi.org/10.1590/S1519-69842007000300002
Liem K.F. 1979. Modulatory multiplicity in the feeding mechanism in cichlid fishes, as exemplified by the invertebrate pickers of lake Tanganyika // J. Zool. V. 189. № 1. P. 93–125. https://doi.org/10.1111/j.1469-7998.1979.tb03954.x
Liem F.L., Osse L.W.M. 1975. Biological versatility, evolution, and food resource exploitation in african cichlid fishes // Amer. Zool. V. 15. № 2. P. 427–454. https://doi.org/10.1093/icb/15.2.427
Maheshwari S.C. 1965. The cranial nerves of Mastacembelus armatus (Lacepede) // Jpn. J. Ichthyol. V. 12. № 3−6. P. 89–98. https://doi.org/10.11369/jji1950.12.89
Manigk W. 1933. Der trigemino-facialiskomplex und die innervation der kopfseitenorgane der elritze (Phoxinus laevis) // Zoomorphology. V. 28. № 1. P. 64–106. https://doi.org/10.1007/BF00408348
Melgarejo-Damian M.P., Gonzalez-Acosta A.F., Cruz-Escalona V.H., Moncayo-Estrada R. 2018. A comparison of feeding biomechanics between two parrotfish species from the Gulf of California // Ibid. V. 137. № 1. P. 165–176. https://doi.org/10.1007/s00435-017-0383-6
Meyer J., Goethals T., Wassenbergh S. et al. 2018. Dimorphism throughout the European eel’s life cycle: are ontogenetic changes in head shape related to dietary differences? // J. Anat. V. 233. № 3. P. 289−301. https://doi.org/10.1111/joa.12836
Mikami Y. 2013. Phylogenic relationship of Labridae species deduced from comparative dissection // Anat. Rec. V. 296. № 5. P. 788–797. https://doi.org/10.1002/ar.22687
Moyer J.T., Sano M. 1987. Feeding habits of two sympatric ostraciid fishes at Miyake-jima, Japan // Jpn. J. Ichthyol. V. 34. № 1. P. 108–112. https://doi.org/10.11369/jji1950.34.108
Nanami A. 2016. Parrotfish grazing ability: interspecific differences in relation to jaw-lever mechanics and relative weight of adductor mandibulae on an Okinawan coral reef // PeerJ. V. 4. P. e2425. https://doi.org/10.7717/peerj.2425
Nakae M., Sasaki K. 2006. Peripheral nervous system of the ocean sunfish Mola mola (Tetraodontiformes: Molidae) // Ichthyol. Res. V. 53. № 3. P. 233–246. https://doi.org/10.1007/s10228-006-0339-1
Nakae M., Sasaki K. 2007. Review of spino-occipital and spinal nerves in Tetraodontiformes, with special reference to pectoral and pelvic fin muscle innervation // Ibid. V. 54. № 4. P. 333–349. https://doi.org/10.1007/s10228-007-0409-z
Nelson J.S. 2006. Fishes of the world. Hoboken, New Jersey: John Wiley and Sons Inc., 624 p.
Ong L., Holland K.N. 2010. Bioerosion of coral reefs by two Hawaiian parrotfishes: species, size differences and fishery implications // Mar. Biol. V. 157. № 6. P. 1313–1323. https://doi.org/10.1007/s00227-010-1411-y
Osse J.W.M. 1969. Functional morphology of the head of the perch (Perca fluviatilis L.): an electromyographic study // Netherl. J. Zool. V. 19. № 3. P. 289–392. https://doi.org/10.1163/002829669X00134
Petersen H. 1914. Studien zur vergleichenden und allgemeinen Mechanik des Tierk6rpers. I. Das Kiefergelenk des Kabeljaus, Gadus morhua // W. Roux’ Arch f. Entw.-Mech. Org. V. 39. № 1. P. 54–111. https://doi.org/10.1007/BF02286973
Randall J.E., Nelson G. 1979. Scarus japanensis, S. quoyi and S. iserti − valid names for parrotfishes presently known as S. capistratoides, S. blochii and S. croicensis // Copeia. № 2. P. 206–212. https://doi.org/10.2307/1443405
Rognes K. 1971. Head skeleton and jaw mechanism in Labrinae (Teleostei: Labridae) from norwegian waters // Acta Univ. Berg. Ser. Math. Rer. Nat. № 4. P. 1–151.
Saxena P.K. 1969. The cranial nerves in Hilsa ilisha (Ham.) and Cirrhina mrigala (Ham.) // Acta Anat. V. 74. P. 197–213.
Shinohara G., Imamura H. 2005. Anatomical description and phylogenetic classification of the orbicular velvetfishes (Scorpaenoidea: Caracanthus) // Ichthyol. Res. V. 52. № 1. P. 64–76. https://doi.org/10.1007/s10228-004-0256-0
Springer V.G., Freihofer W.C. 1976. Study of the monotypic fish family Pholidichthyidae (Perciformes) // Smith. Contr. Zool. № 216. P. 1–41.
Stiassny M.L.J. 1981a. The phyletic status of the family Cichlidae (Pisces, Perciformes): a comparative anatomical investigation // Netherl. J. Zool. V. 31. № 2. P. 275–314.
Stiassny M.L.J. 1981b. Phylogenetic versus convergent relationship between piscivorous cichlid fishes from lakes Malawi and Tanganyika // Bull. Brit. Mus. Nat. Hist. Zool. V. 40. № 3. P. 67–101.
Stiassny M.L.J. 1990. Tylochromis, relationships and the phylogenetic status of the african Cichlidae // Amer. Mus. Nov. № 2993. P. 1–14.
Tedman R.A. 1980a. Comparative study of the cranial morphology of the labrids Choerodon venustus and Labroides dimidiatus and the scarid Scarus fasciatus (Pisces: Perciformes) I. Head skeleton // Austal. J. Mar. Freshwat. Res. V. 31. № 3. P. 337–349. https://doi.org/10.1071/MF9800337
Tedman R.A. 1980b. Comparative study of the cranial morphology of the labrids Choerodon venustus and Labroides dimidiatus and the scarid Scarus fasciatus (Pisces: Perciformes). II. Cranial myology and feeding mechanisms // Ibid. V. 31. № 3. P. 351–372. https://doi.org/10.1071/MF9800351
Thorsen D.H., Westneat M.W. 2005. Diversity of pectoral fin structure and function in fishes with labriform propulsion // J. Morphol. V. 263. № 2. P. 133–150. https://doi.org/10.1002/jmor.10173
Turingan R.G., Wainwright P.C. 1993. Morphological and functional bases of durophagy in the queen triggerfish, Balistes vetula (Pisces, Tetraodontiformes) // Ibid. V. 215. № 2. P. 101–118. https://doi.org/10.1002/jmor.1052150202
Vandewalle P., Parmentier E., Chardon M. 2000. The branchial basket in teleost feeding // Cybium. V. 24. № 4. P. 319–342.
Vial Cl.I., Ojeda F.P. 1990. Cephalic anatomy of the herbivorous fish Girella laevifrons (Osteichthyes: Kyphosidae): mechanical considerations of its trophic function // Rev. Chil. Hist. Natur. V. 63. № 3. P. 247–260.
Wassenbergh S.V., Herrel A., Adriaens D., Aerts P. 2004. Effects of jaw adductor hypertrophy on buccal expansions during feeding of air breathing catfishes (Teleostei, Clariidae) // Zoomorph. V. 123. P. 81–93. https://doi.org/10.1007/s00435-003-0090-3
Westneat M.W., Wainwright P.C. 1989. Feeding mechanism of Epibulus insidiator (Labridae; Teleostei): evolution of a novel functional system // J. Morphol. V. 202. № 2. P. 129–150. https://doi.org/10.1002/jmor.1052020202
Winterbottom R. 1974. A descriptive synonymy of the striated muscles of the Teleostei // Proc. Acad. Nat. Sci. Phil. V. 125. P. 225–317.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии