

Вопросы ихтиологии, 2020, T. 60, № 2, стр. 183-191
Динамика показателей линейного роста сельди Clupea pallasii залива Петра Великого (Японское море)
Л. А. Черноиванова *
Тихоокеанский филиал Всероссийского научно-исследовательского института рыбного хозяйства и океанографии − ТИНРО
Владивосток, Россия
* E-mail: chlas27@yandex.ru
Поступила в редакцию 29.05.2019
После доработки 06.06.2019
Принята к публикации 06.06.2019
Аннотация
Анализ размерно-возрастного состава тихоокеанской сельди Clupea pallasii зал. Петра Великого в 1941, 1944 и 1947−2017 гг. показал наличие статистически достоверно отличающихся линейными размерами категорий крупных, средних и мелких рыб. Их доли в поколениях подвержены долгопериодным и межгодовым изменениям. Выделенные группировки рыб различаются длительностью жизненного цикла, временем полового созревания и динамикой численности, но не являются полностью репродуктивно изолированными. В многолетней динамике соотношения размерных категорий в рассматриваемых поколениях в периоды подъёма запасов доминируют крупные и средние особи с положительными отклонениями линейного роста, а в годы низкой численности преобладают группировки мелких и средних рыб с отрицательными отклонениями индексов роста.
В дальневосточных морях известны популяции тихоокеанской сельди Clupea pallasii с быстрым, с медленным ростом и растущие в умеренном темпе. В каждой из них существует межгодовая изменчивость скорости роста, иногда достигающая различий межпопуляционного уровня и часто связанная с периодичностью изменений численности (Науменко, 2001). Считается, что сельдь зал. Петра Великого имеет быстрый темп линейного роста (Науменко, 2001, 2002). Судя по результатам анализа размерно-возрастного состава, с наиболее высокой скоростью рыбы этой популяции росли в периоды значительных запасов – в середине 1920-х и с середины 1970-х до конца 1980-х гг. (Амброз, 1931; Посадова, 1985, 1988). Показатели роста особей уменьшились в период депрессии популяции в 1990-х и 2000-х гг. (Черноиванова и др., 2009). На протяжении всего периода исследований с середины 1920-х гг. до настоящего времени, кроме долговременной и межгодовой вариабельности показателей роста, отмечено наличие быстро-, средне- и медленнорастущих рыб в пределах одновозрастных классов (Черноиванова, Ким, 2016). Исследования динамики численности в популяциях тихоокеанской сельди показали, что уровень запасов в значительной степени определяется скоростью роста поколений, его межгодовой и долгопериодной изменчивостью (Науменко, 2001).
В связи с этим цель работы – определить особенности линейного роста разных поколений сельди зал. Петра Великого.
МАТЕРИАЛ И МЕТОДИКА
Сбор материалов по размерно-возрастному составу сельди в зал. Петра Великого в нерестовые сезоны был начат с середины 1920-х гг. и проводится до настоящего времени. В работе использованы имеющиеся архивные данные ТИНРО-центра за 1941, 1944 и 1947−1995 гг., а также собственные материалы за 1996–2017 гг. Всего за рассматриваемый период (1941–2017 гг.) проанализировано >60 тыс. рыб в возрасте 2−13 полных лет. Данные, относящиеся к 1920-м гг., приведены по монографии Амброза (1931). Длину рыб измеряли по Смитту (FL); возраст определяли по чешуе.
При расчёте общих характеристик вариационного ряда длины полагали, что частотное распределение значений, ~95% которых находились в пределах двух стандартных отклонений от среднего, подобно нормальному. Границы нормы распределения длины в возрастных классах устанавливали в соответствии с правилом двух сигм (Гланц, 1999). Рыбы с длиной тела в пределах среднемноголетних значений отнесены к категории средних; с выходящей за пределы двух стандартных отклонений в сторону уменьшения или увеличения − к категории мелких или крупных (табл. 1).
Таблица 1.
Параметры распределений для категорий мелких, средних и крупных особей тихоокеанской сельди Clupea pallasii зал. Петра Великого (Черноиванова, Ким, 2016)
| Категория размеров | Показатель вариации | Возраст, годы (число рыб, экз.) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | ||
| (1031) | (7827) | (13229) | (12241) | (8061) | (7703) | (5508) | (2910) | (1081) | (331) | (108) | (15) | (2) | ||
| Мелкие | М | 12.80 | 19.60 | 22.20 | 26.30 | 28.10 | 30.90 | 31.90 | 32.20 | – | – | – | – | – |
| m | 0.06 | 0.04 | 0.02 | 0.03 | 0.04 | 0.03 | 0.07 | 0.09 | – | – | – | – | – | |
| σ | 0.96 | 1.28 | 1.21 | 1.43 | 1.54 | 1.09 | 1.00 | 0.57 | – | – | – | – | – | |
| min | 8.50 | 15.50 | 17.50 | 19.50 | 21.50 | 24.70 | 27.50 | 30.50 | – | – | – | – | – | |
| max | 13.00 | 22.50 | 25.00 | 28.50 | 31.50 | 33.50 | 34.00 | 34.00 | – | – | – | – | – | |
| Средние | М | 15.30 | 22.70 | 25.80 | 29.50 | 31.60 | 33.40 | 35.00 | 36.10 | 36.70 | 37.60 | 37.60 | 37.40 | 37.5 |
| m | 0.04 | 0.02 | 0.02 | 0.01 | 0.01 | 0.01 | 0.01 | 0.02 | 0.03 | 0.06 | 0.12 | 0.29 | – | |
| σ | 1.04 | 1.27 | 1.89 | 1.21 | 1.13 | 0.94 | 0.91 | 0.85 | 0.92 | 1.06 | 1.25 | 1.13 | – | |
| min | 13.20 | 20.20 | 22.10 | 27.10 | 29.50 | 31.40 | 33.20 | 34.10 | 34.50 | 35.10 | 35.10 | 35.10 | 37.0 | |
| max | 17.00 | 25.50 | 29.00 | 32.00 | 34.00 | 35.00 | 37.00 | 38.00 | 39.00 | 40.00 | 40.00 | 40.00 | 38.0 | |
| Крупные | М | 17.60 | 26.10 | 29.10 | 32.80 | 34.90 | 35.70 | 36.60 | – | – | – | – | – | – |
| m | 0.04 | 0.04 | 0.02 | 0.03 | 0.04 | 0.02 | 0.01 | – | – | – | – | – | – | |
| σ | 0.34 | 1.08 | 0.84 | 0.64 | 0.71 | 0.44 | 0.38 | – | – | – | – | – | – | |
| min | 16.90 | 24.50 | 28.30 | 32.00 | 33.00 | 34.00 | 35.90 | – | – | – | – | – | – | |
| max | 20.00 | 29.50 | 32.50 | 35.00 | 36.50 | 37.50 | 37.50 | – | – | – | – | – | – | |
Диапазоны варьирования длины мелких и крупных категорий сельдей определяли по соотношению между процентилями и числом стандартных отклонений от среднего (Гланц, 1999): М – 2σ = 2.5%, М – σ = 16.0%, М = 50.0%, М + σ = = 84.0%, М + 2σ = 97.5%. Во всех возрастных классах средняя длина рыб в выделенных категориях соответствовала доверительному интервалу среднего значения (порог доверительной вероятности – 95%).
В настоящей работе принято, что в поколениях сельди доминирует одна из категорий в тех случаях, когда в уловах частота её встречаемости в течение жизни генерации составляет 50–100% (массовое количество). При встречаемости данной категории в поколении 30–50% она количественно составляет значительную долю, 10–30% − заметную численность в поколении, <10% − её доля незначительная.
Изменение темпа линейного роста сельди в выделенных размерных категориях рассматривали на основании статистического описания динамического ряда (Ефимова и др., 2007). Индекс отклонения линейных размеров (T, %) в конкретном году вычисляли по формуле: Tni = xni/xnср × 100 – 100, где Tni – индекс отклонения линейных размеров сельди возрастного класса n данной размерной категории в i-том году, xni – средняя длина возрастного класса n данной размерной категории в i-том году, xnср – среднемноголетнее значение длины возрастного класса n данной размерной категории.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Жизненный цикл сельди зал. Петра Великого проходит в заливе и прилегающих водах юго-западной части Японского моря. По сравнению с другими дальневосточными популяциями морских сельдей уровень её запасов невысокий; для неё известны кратковременные периоды высокой численности и более продолжительные – низкой численности. Нерест этой популяции проходит в прибрежной части зал. Петра Великого и небольшой акватории прилегающих вод Северного Приморья, и только при увеличении запасов нерестилища распространяются до зал. Ольга (Гаврилов, Посадова, 1982; Вдовин, Черноиванова, 2006).
Анализ изменчивости линейного роста сельди зал. Петра Великого проводили по данным наблюдений в период нереста в 1926–1927, 1941, 1944 и 1947–2017 гг. В пределах одновозрастных классов были выделены три размерные категории рыб − средние, мелкие и крупные, имеющие при этом разную продолжительность жизни. Предельный возраст особей, представляющих ранг мелких, составил 8 полных лет, крупных – 7, средних − 13 (Черноиванова, Ким, 2016). Соотношение размерных категорий в рассматриваемых поколениях 1918−1925 и 1934−2013 гг. варьирует от полного преобладания одной из них до её отсутствия (табл. 2).
Таблица 2.
Соотношение размерных категорий тихоокеанской сельди Clupea pallasii зал. Петра Великого в поколениях 1918−1925 и 1934−2013 гг.
| Год рождения | Доля размерных категорий, % | ||
|---|---|---|---|
| мелкие | средние | крупные | |
| 1918 | 40.0 | 46.7 | 13.3 |
| 1919 | 42.4 | 51.5 | 6.1 |
| 1920 | 38.2 | 55.4 | 6.4 |
| 1921 | 5.4 | 84.0 | 10.6 |
| 1922 | 0 | 79.6 | 20.4 |
| 1923 | 0 | 62.2 | 37.8 |
| 1924 | 0 | 26.6 | 73.4 |
| 1925 | 1.4 | 49.5 | 49.1 |
| 1934 | 100 | 0 | 0 |
| 1935 | 78.1 | 21.9 | 0 |
| 1936 | 86.4 | 13.6 | 0 |
| 1937 | 86.6 | 13.4 | 0 |
| 1938 | 54.2 | 45.8 | 0 |
| 1939 | 38.9 | 61.1 | 0 |
| 1940 | 23.1 | 76.9 | 0 |
| 1941 | 1.8 | 96.5 | 1.7 |
| 1942 | 2.8 | 95.3 | 1.9 |
| 1943 | 80.0 | 20.0 | 0 |
| 1944 | 1.5 | 98.1 | 0.4 |
| 1945 | 1.0 | 98.4 | 0.6 |
| 1946 | 2.5 | 92.7 | 4.8 |
| 1947 | 0.3 | 91.3 | 8.4 |
| 1948 | 0.9 | 80.7 | 18.4 |
| 1949 | 0.5 | 75.7 | 23.8 |
| 1950 | 0 | 72.5 | 27.5 |
| 1951 | 0.8 | 92.7 | 6.5 |
| 1952 | 16.9 | 79.6 | 3.5 |
| 1953 | 1.3 | 93.3 | 5.4 |
| 1954 | 0 | 64.9 | 35.1 |
| 1955 | 0.1 | 75.7 | 24.2 |
| 1956 | 0.5 | 97.3 | 2.2 |
| 1957 | 0.4 | 92.6 | 7.0 |
| 1958 | 1.2 | 81.3 | 17.5 |
| 1959 | 19.6 | 68.6 | 11.8 |
| 1960 | 20.0 | 73.0 | 7.0 |
| 1961 | 24.7 | 69.8 | 5.5 |
| 1962 | 35.6 | 63.7 | 0.7 |
| 1963 | 3.2 | 65.4 | 31.4 |
| 1964 | 71.5 | 19.7 | 8.8 |
| 1965 | 9.6 | 89.8 | 0.6 |
| 1966 | 43.2 | 53.6 | 3.2 |
| 1967 | 14.2 | 85.0 | 0.8 |
| 1968 | 3.5 | 94.9 | 1.6 |
| 1969 | 26.0 | 72.2 | 1.8 |
| 1970 | 23.2 | 65.5 | 11.3 |
| 1971 | 1.1 | 91.5 | 7.4 |
| 1972 | 3.5 | 69.8 | 26.7 |
| 1973 | 12.6 | 79.3 | 8.1 |
| 1974 | 1.5 | 91.6 | 6.9 |
| 1975 | 1.5 | 89.3 | 9.2 |
| 1976 | 2.3 | 88.6 | 9.1 |
| 1977 | 1.4 | 93.6 | 5.0 |
| 1978 | 2.7 | 94.8 | 2.5 |
| 1979 | 10.3 | 84.9 | 4.8 |
| 1980 | 1.7 | 96.3 | 2.0 |
| 1981 | 0.5 | 89.9 | 9.6 |
| 1982 | 0.7 | 87.9 | 11.4 |
| 1983 | 0.5 | 91.5 | 8.0 |
| 1984 | 0.9 | 87.0 | 12.1 |
| 1985 | 25.9 | 68.7 | 5.4 |
| 1986 | 15.6 | 78.8 | 5.6 |
| 1987 | 8.2 | 81.5 | 10.3 |
| 1988 | 0.9 | 85.5 | 13.6 |
| 1989 | 8.6 | 79.1 | 12.3 |
| 1990 | 7.4 | 74.4 | 18.2 |
| 1991 | 29.3 | 57.4 | 13.3 |
| 1992 | 17.0 | 67.2 | 15.8 |
| 1993 | 13.8 | 64.2 | 22.0 |
| 1994 | 32.4 | 52.0 | 15.6 |
| 1995 | 55.9 | 41.4 | 2.7 |
| 1996 | 63.0 | 34.3 | 2.7 |
| 1997 | 45.1 | 54.3 | 0.6 |
| 1998 | 51.4 | 44.8 | 3.8 |
| 1999 | 67.2 | 31.7 | 1.1 |
| 2000 | 72.4 | 27.5 | 0.1 |
| 2001 | 23.8 | 74.0 | 2.2 |
| 2002 | 7.8 | 91.0 | 1.2 |
| 2003 | 16.7 | 81.9 | 1.4 |
| 2004 | 24.4 | 69.6 | 6.0 |
| 2005 | 48.2 | 48.5 | 3.3 |
| 2006 | 7.5 | 91.4 | 1.1 |
| 2007 | 32.0 | 59.7 | 8.3 |
| 2008 | 50.5 | 49.0 | 0.5 |
| 2009 | 38.5 | 61.2 | 0.3 |
| 2010 | 40.3 | 59.7 | 0 |
| 2011 | 40.5 | 58.9 | 0.6 |
| 2012 | 20.8 | 78.0 | 1.2 |
| 2013 | 23.6 | 75.5 | 0.9 |
Категория крупной сельди в массовом количестве (73.4%) была представлена только в поколении 1924 г. Крупные особи чаще составляли или значительную (30−50%), или заметную (10−30%) долю в поколениях, существовавших в периоды относительно высокой численности – в первой половине 1920-х, в 1948−1949, в 1950-х, в первой половине 1970-х и в серии поколений с 1982 по 1994 гг.
Мелкая сельдь доминировала или составляла значительную долю в поколениях 1934−1939, 1994−2000, 2005 и 2007−2011 гг. Заметная численность мелких рыб (10–30%) наблюдалась в отдельных поколениях 1950-х, 1960-х 1970-х, 1980-х и 2012−2013 гг. Появление генераций с мелкой сельдью в массовых или значительных количествах сопровождалось, как правило, снижением запасов. Например, 1930-е гг. и современный длительный период депрессии сельди в зал. Петра Великого характеризуются преобладанием мелкой категории рыб в поколениях и сокращением возрастного ряда. Категория средних рыб, присутствовавшая практически во всех рассматриваемых поколениях, за исключением одного – 1934 г. рождения, доминировала в подавляющем большинстве генераций. В отличие от категорий мелких и крупных рыб, в категории средних отмечены существенные изменения скорости роста особей в рассматриваемом ряду поколений, поэтому её динамику анализировали детальнее (рис. 1). В категории средних рыб отрицательные индексы отклонения линейных размеров от среднемноголетних значений практически для всех возрастных классов наблюдались в поколениях 1935–1942 и 1994−2015 гг. рождения. Исключения составили некоторые поколения 2000-х гг.: в возрасте 2 и 3 года их скорость роста превышала среднемноголетние показатели (рис. 1а, 1б). Отметим, что в категории средних рыб отрицательные отклонения от среднемноголетних значений длины чаще появлялись в поколениях со значительной долей или доминированием в них сельди мелкой категории. Положительные отклонения индекса линейного роста в категории средних рыб отмечались в поколениях сельди 1945−1959 гг. от 2-го до 5-го года жизни (рис. 1а–1г), начиная с 6-го года и старше в большинстве указанных поколений наблюдался замедленный темп роста (рис. 1д–1м). Положительными значениями показателя роста (индекс Т) для всех присутствующих возрастных классов характеризовались поколения 1973−1984 гг. (рис. 1а–1к).
Рис. 1.
Отклонения линейных размеров относительно среднемноголетних значений (T, %) тихоокеанской сельди Clupea pallasii разного возраста в поколениях 1935−2015 гг. для категории средних рыб в зал. Петра Великого: а – 2 года, б – 3 года, в – 4 года, г – 5 лет, д – 6 лет, е – 7 лет, ж – 8 лет, з – 9 лет, и – 10 лет, к – 11 лет, л – 12 лет, м – 13 лет; *данные по росту сельди за 1927 г.: Амброз, 1931, (- -) – уровень значимости 0.01.
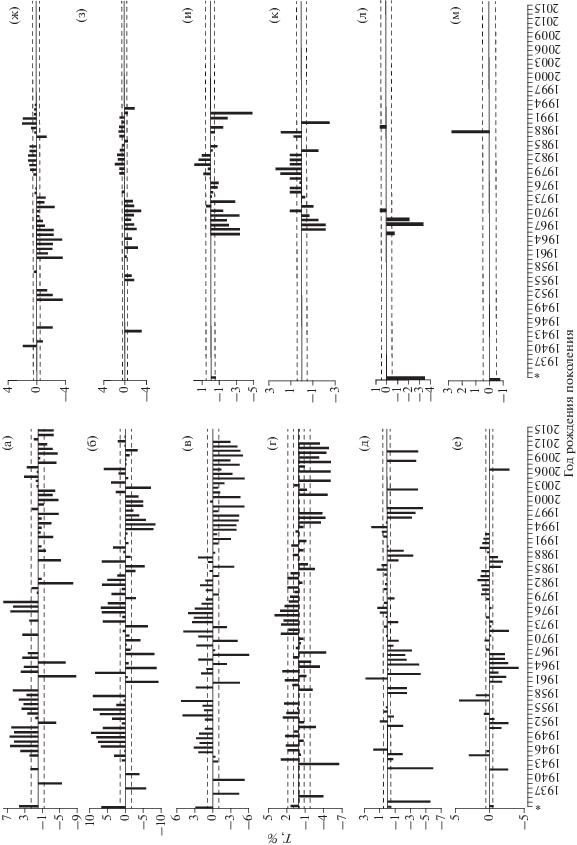
Межгодовые колебания замедления или ускорения темпа роста оказались наиболее выраженными среди поколений 1960−1972 гг., причём заметная межгодовая изменчивость отмечалась для рыб в возрасте 2−5 лет, в то время как у более старших особей этих генераций были только отрицательные значения индекса отклонения длины тела. В генерациях, родившихся в 1985−1994 гг., также наблюдалась межгодовая изменчивость роста в 2–5-годовалом возрасте, но группы старшего возраста в поколениях 1985−1987 гг. имели отрицательные отклонения от среднемноголетней длины, а темп роста особей того же возраста 1988−1992 гг. рождения превышал средние показатели. По материалам 1927 г. (Амброз, 1931), в поколениях 1917−1925 гг. линейные размеры в средней категории сельди либо превосходили, либо были близки к среднемноголетним значениям во всех возрастных классах (рис. 1).
Сопоставление динамики численности популяции сельди зал. Петра Великого и темпа роста рыб средней размерной категории показало наличие периодической изменчивости скорости роста поколений. Наиболее высокие показатели линейного роста наблюдались в поколениях 1917−1925, 1951−1957 и 1974−1982 гг., т.е. в фазах увеличения численности и её высокого уровня. В фазах снижения запасов и в переходные периоды между их высоким уровнем присутствовали поколения как с положительными, так и с отрицательными значениями индекса отклонения линейных размеров. После снижения численности в начале 1930-х гг. и в период затяжной депрессии (в поколениях сельди с 1994 г. до настоящего времени) показатели темпа роста в средней категории рыб имели заметные отрицательные отклонения.
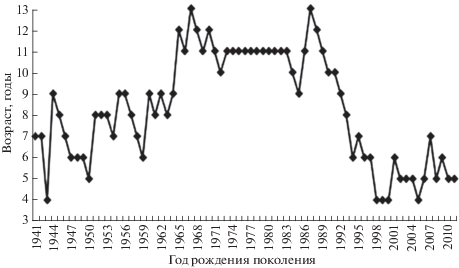
По нашим данным, в категории средних рыб вариабельность темпа роста сочеталась с изменениями максимального возраста. В поколениях с медленным ростом в годы низкой численности жизненный цикл сокращался. При высоком уровне запасов, в середине 1920-х и в конце 1970-х–начале 1980-х гг., в категории средних рыб не только доминировали особи с высоким темпом роста, но были отмечены высокая доля особей старшего возраста и максимальная продолжительность жизни поколений (рис. 2). В генерациях, появившихся в кратковременный период подъёма запасов в середине 1950-х гг., рыбы средней категории также имели высокий темп роста, но непродолжительный жизненный цикл (9 лет против максимума для данной категории 13 лет). В отмеченной выше межгодовой вариабельности темпа роста в категории средних рыб в 1960−1972 гг. в первой половине 1960-х гг. преобладали поколения с максимальным возрастом 8−9 лет, а с середины 1960-х гг. появились долгоживущие генерации, обеспечившие подъём численности 1970–1980-х гг. Межгодовые колебания темпа роста и продолжительности жизни поколений в средней категории рыб, наблюдающиеся в фазах снижения запасов и в годы их относительно низкой величины, отражают переход популяции сельди зал. Петра Великого к изменению таких параметров, сопряжённых с темпом роста, как длительность жизненного цикла, размеры впервые созревающих особей, возраст массового полового созревания, и к формированию другого уровня численности (Вдовин, Черноиванова, 2006; Черноиванова и др., 2009).
Рис. 2.
Динамика максимального возраста в поколениях средней размерной категории тихоокеанской сельди Clupea pallasii зал. Петра Великого, 1941−2015 гг.
Реализация потенциала популяции к наращиванию численности зависит от взаимодействия динамики основных биологических параметров её нерестовой части и измечивости среды. В популяции сельди зал. Петра Великого повторяемость урожайных поколений существенно возрастает в тёплые годы, если преобладают короткоцикловые генерации, и в периоды похолоданий, если доминируют долгоживущие. Причём уровень численности в разные климатические фазы различается. Долгоживущие быстрорастущие поколения в ответ на благоприятные условия способны по ряду биологических показателей (таких как более высокая плодовитость, длительность генеративного периода особей в течение жизни, более широкий нагульный и нерестовый ареалы, чем у короткоцикловых поколений) наращивать высокую численность (Черноиванова, 2013).
Таким образом, многолетняя динамика соотношения размерных категорий в рассматриваемых поколениях следует тенденции преобладания группировок с замедленным ростом в поколениях в годы низкой численности и доминирования сельди с высоким темпом роста в генерациях в периоды формирования значительных запасов.
Долгопериодные и межгодовые вариации ростовых параметров, замедляющиеся в годы низкой численности и ускоряющиеся в периоды роста запасов, которые наблюдаются в популяции зал. Петра Великого, известны также у восточноберинговоморской сельди, но в большинстве дальневосточных популяций в периоды высокой численности и в годы глубокой депрессии скорость линейного роста ниже, чем в периоды средней и низкой численности (Науменко, 2001).
Ранее было отмечено, что катастрофическое падение численности сельди в зал. Петра Великого после 1930 г. сопровождалось уменьшением числа возрастных групп (Гаврилов, Посадова, 1982). В современный период депрессии в популяции преобладают поколения с укороченным жизненным циклом, низким темпом роста, минимальной длиной впервые созревающих особей и массовым половым созреванием двухлеток (Черноиванова и др., 2009; Черноиванова, 2013).
Рассматриваемые особенности линейного роста различных размерных категорий сельди зал. Петра Великого позволили выделить несколько одновозрастных группировок с разным типом роста (рис. 3). Различия средней длины особей основных выделенных категорий (крупных, средних и мелких рыб), оцениваемые согласно критерию Стьюдента, оказались статистически достоверными при уровне значимости p < 0.001 (табл. 3). Наиболее различается темп роста группировок мелких и крупных рыб. Сельди этих категорий заметно различаются сроками нерестовых подходов: крупные особи подходят на нерест в феврале–марте, нерест мелких проходит в конце апреля и в мае. В период нагула мелкая сельдь располагается до 50-метровой изобаты, в осенне-зимний период держится в прибрежной части залива (в мелководной части залива или на местах летнего нагула вблизи нерестилищ) и заходит в эстуарии рек. Крупная сельдь в летний период распространяется до глубин 200–300 м, зимует в относительно тёплом промежуточном слое открытых вод зал. Петра Великого. Сельди категории мелких рыб лучше адаптированы к прибрежным условиями, категория крупных рыб соответствует типу морских сельдей по классификации Науменко (2001).
Рис. 3.
Линейный рост поколений тихоокеанской сельди Clupea pallasii зал. Петра Великого, относящихся к размерным категориям крупных (– –), средних (—) и мелких (- -) рыб, а также поколений категории средних рыб с положительными (▲) и отрицательными (⚫, ⚪) отклонениями от среднемноголетнего значения; (⚪) − поколения 1994–2015 гг. с укороченным жизненным циклом; пояснения см. в тексте.
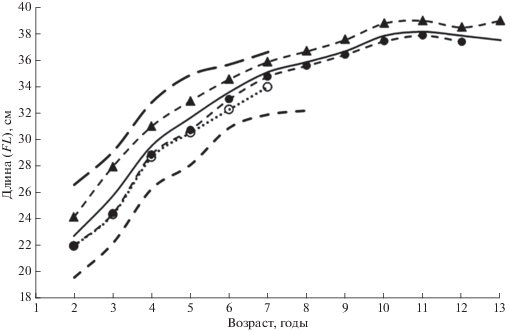
Таблица 3.
Критерий различий Стьюдента для средних значений длины выделенных размерных категорий тихоокеанской сельди Clupea pallasii зал. Петра Великого в зависимости от возраста
| Возраст, годы | Сравниваемые размерные категории | ||
|---|---|---|---|
| средние–мелкие | средние–крупные | мелкие–крупные | |
| 2 | 77.1 | 74.7 | 115.4 |
| 3 | 133.9 | 131.2 | 272.4 |
| 4 | 107.0 | 114.5 | 171.8 |
| 5 | 85.5 | 78.1 | 122.9 |
| 6 | 68.8 | 99.7 | 119.1 |
| 7 | 46.5 | 86.9 | 69.7 |
| 8 | 40.2 | ||
Категория средних рыб представляет собой основу численности популяции сельди зал. Петра Великого. В отличие от мелких и крупных она включает в себя поколения с периодически изменяющимся темпом роста особей, сочетающимся с изменениями длительности жизненного цикла. В этой категории в разные годы наблюдались (рис. 3): 1) долгоживущие поколения, колебания линейных размеров (положительные и отрицательные) которых во всех возрастных классах не превышали среднемноголетнее значение (p > 0.01); 2) долгоживущие поколения, линейные размеры всех возрастных классов которых превышали среднемноголетнее значение (p < 0.01); 3) долгоживущие поколения, линейные размеры всех возрастных классов которых меньше среднемноголетнего значения (p < 0.01); 4) поколения с укороченным жизненным циклом, линейные размеры всех возрастных классов которых меньше среднемноголетнего значения ( p < 0.01), поколения такого типа появлялись в 1994–2015 гг.
Для всех поколений в категории средних рыб характерны нерестовые подходы с середины марта до середины апреля, причём сроки массового нереста периодически смещаются в зависимости от размерного состава поколений: сельдь поколений с замедленным темпом роста и укороченным жизненным циклом нерестится преимущественно в апреле, а долгоживущих и лучше растущих поколений – в марте. Вероятно, вариации темпа роста в категории средних рыб обусловлены наибольшей возможностью их перемешивания с крайними категориями, особенно в нерестовый и посленерестовый периоды. В годы заметной численности крупных рыб в средней категории формировались типы долгоживущих поколений с темпом роста, близким или несколько превышающим среднемноголетний уровень. Напротив, влияние особей мелкой категории в годы их преобладания способствовало развитию поколений с укороченным жизненным циклом и невысоким темпом роста. Наибольшей численности популяция сельди в зал. Петра Великого достигала в периоды доминирования долгоживущих быстрорастущих поколений с заметной долей присутствия особей крупной категории.
Для тихоокеанской сельди характерно наличие локальных малочисленных популяций, периодически формирующих высокую численность. Они не являются репродуктивно изолированными и имеют возможность значительно перемешиваться. Например, известное в Японском море сахалино-хоккайдское стадо в период высоких запасов составляет единую суперпопуляцию, в годы со средней численностью имеет соподчинённые единицы раннего и позднего нереста, а в текущий период низкой численности существует в виде местных сельдей северной части Японского моря, юго-западного побережья Сахалин и западного побережья Хоккайдо. Эти группировки разобщенны по срокам и районам нереста, различаются по морфологическим и биологическим признакам и нагульным ареалам (Науменко, 2001; Ившина, 2008).
Выделенные в одновозрастных классах сельди зал. Петра Великого категории рыб, различающиеся по темпу роста, длительности жизненного цикла, минимальной длине и возрасту полового созревания, демонстрируют адаптивные возможности вида, даже в пределах популяции, к относительно быстрому во времени формированию группировок рыб: одни из них способны существовать в локальных условиях прибрежных районов, а другие совершают протяжённые миграции в периоды нагула и зимовки.
ЗАКЛЮЧЕНИЕ
В относительно небольшой по численности и занимаемой акватории популяции сельди зал. Петра Великого наблюдается неоднородность темпа линейного роста особей в одновозрастных классах. Категории крупных, средних и мелких рыб достоверно отличаются линейными размерами. Их численное соотношение в поколениях изменяется как в межгодовом, так и в долгопериодном аспектах. Выделенные группировки различаются длительностью жизненного цикла, размерами тела, возрастом полового созревания и динамикой численности, но в связи с ограниченным районом обитания не могут быть полностью репродуктивно изолированными. Наиболее обособленные категории мелких и крупных рыб, имея общий нерестовый ареал, разобщены во времени нерестовых подходов, а также в пространственном расположении районов нагула и зимовки. Сельди категории мелких рыб лучше адаптированы к прибрежным условиями, а категория крупных рыб соответствует экологической форме морских сельдей.
В категории средних рыб, составляющей основу численности сельди зал. Петра Великого, заметные периодические сдвиги в сторону замедления или ускорения темпа роста обусловлены большей возможностью перемешивания с категориями крупных и мелких рыб, особенно в нерестовый и посленерестовый периоды. Типы долгоживущих поколений, состоящих из рыб с положительными индексами линейного роста, или короткоцикловых – с отрицательными, формируются в соответствии с преобладающим влиянием категорий крупных или мелких рыб. Наибольшей численности популяция сельди в зал. Петра Великого достигает в периоды доминирования долгоживущих поколений среднего ранга с заметной долей особей крупной категории.
В многолетней динамике соотношения размерных категорий в рассматриваемых поколениях в периоды подъёма запасов доминируют крупные и средние сельди с положительными отклонениями линейного роста, а в годы низкой численности преобладают группировки рыб мелкого ранга и среднего с отрицательными индексами роста.
Список литературы
Амброз А.И. 1931. Сельдь (Clupea harengus pallasi C.V.) залива Петра Великого. Биологический очерк // Изв. ТИНРО. Т. 6. 313 с.
Вдовин А.Н., Черноиванова Л.А. 2006. Многолетняя динамика некоторых параметров популяции тихоокеанской сельди Clupea pallasii (Clupeidae) залива Петра Великого (Японское море) // Вопр. ихтиологии. Т. 46. № 1. С. 54−61.
Гаврилов Г.М., Посадова В.П. 1982. Динамика численности тихоокеанской сельди Clupea pallasi pallasi Valenciennes (Clupeidae) залива Петра Великого // Там же. Т. 22. Вып. 5. С. 760−772.
Гланц С. 1999. Медико-биологическая статистика. М.: Практика, 459 с.
Ефимова М.Р., Петрова Е.В., Румянцев В.Н. 2007. Общая теория статистики. М.: ИНФРА-М, 416 с.
Ившина Э.Р. 2008. Основные черты биологии и современное состояние запасов сельди (Clupea pallasii Valenciennes, 1847): Автореф. дис. … канд. биол. наук. Южно-Сахалинск: СахНИРО, 19 с.
Науменко Н.И. 2001. Биология и промысел морских сельдей Дальнего Востока. Петропавловск-Камчатский: Камчат. печат. двор, 330 с.
Науменко Н.И. 2002. О росте тихоокеанской сельди (Clupea pallasi) // Исследования водных биологических ресурсов Камчатки и северо-западной части Тихого океана. Вып. 6. Петропавловск-Камчатский: Изд-во КамчатНИРО. С. 67−74.
Посадова В.П. 1985. Межгодовая изменчивость нерестовых подходов сельди залива Петра Великого // Сельдевые северной части Тихого океана. Владивосток: Изд-во ТИНРО. С. 22−29.
Посадова В.П. 1988. Состояние запасов сельди залива Петра Великого // Изменчивость состава ихтиофауны, урожайности поколений и методы прогнозирования запасов рыб в северной части Тихого океана. Владивосток: Изд-во ТИНРО. С. 64−69.
Черноиванова Л.А. 2013. Эффективность воспроизводства тихоокеанской сельди Clupea pallasii Val залива Петра Великого (Японское море) в зависимости от биологического состояния производителей и изменчивости среды // Изв. ТИНРО. Т. 175. С. 42−55.
Черноиванова Л.А., Ким Л.Н. 2016. Дифференциация по размерам в одновозрастных классах сельди Clupea pallasii Val. залива Петра Великого (Японское море) // Сб. матер. Всерос. науч.-практ. конф. “Морские биологические исследования: достижения и перспективы”. Т. 3. Севастополь: ЭКОСИ−Гидрофизика. С. 475−478.
Черноиванова Л.А., Измятинский Д.В., Соломатов С.Ф. и др. 2009. Оценка биологического состояния репродуктивной части популяции сельди залива Петра Великого (Японское море) по результатам мониторинга 2007−2008 гг. // Матер. III Междунар. науч.-практ. конф. “Морские прибрежные экосистемы. Водоросли, беспозвоночные и продукты их переработки”. Владивосток: Изд-во ТИНРО-центр. С. 84−89.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии