

Вопросы ихтиологии, 2020, T. 60, № 3, стр. 283-294
Крупные африканские усачи: векторы диверсификации особей генерализованной формы как основа морфологического разнообразия комплекса Barbus intermedius в озере Тана, Эфиопия
1 Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
2 Институт биологии внутренних вод РАН – ИБВВ РАН
Ярославская область, Борок, Россия
* E-mail: adissa@mail.ru
Поступила в редакцию 24.04.2019
После доработки 13.08.2019
Принята к публикации 13.08.2019
Аннотация
Анализировали морфологическую диверсификацию особей генерализованной формы комплекса Barbus intermedius в сравнении с диверсификацией специализированных морфотипов и аллопатрических видов крупных африканских усачей. В совокупностях генерализованных особей выявлено четыре главных вектора диверсификации, у морфотипов и видов – по два. Сравнение пар одноранговых векторов диверсификации мономорфных популяций генерализованных особей из озёр Лангано и Аваса обнаруживает их сходство между собой и существенные отличия от соответствующих векторов диверсификации генерализованных особей из оз. Тана. Векторы диверсификации танских морфотипов весьма сходны как с одноранговыми векторами аллопатрических видов, так и с соответствующими векторами генерализованной формы из оз. Тана. Высокая степень этого сходства даёт основания полагать, что диверсификация особей генерализованной формы из оз. Тана, диверсификация танских морфотипов и диверсификация аллопатрических видов – это последовательные этапы одного процесса, и диверсификация морфотипов и видов идёт в направлениях, заданных диверсификацией особей генерализованной формы. Результаты исследования можно рассматривать как весомое свидетельство в пользу гипотезы о близости (или тождестве) генерализованной формы и формы предковой, давшей начало разнообразию комплекса B. intermedius. Исследование основано на оценке корреляций между признаками особей изучаемых совокупностей.
Крупные африканские усачи комплекса Barbus intermedius (sensu Banister, 1973) привлекают внимание исследователей прежде всего тем, что в ряде водоёмов Эфиопии они образуют симпатрические формы (морфотипы), специализированные в отношении питания. Наибольшее разнообразие форм усачей наблюдается в оз. Тана, где различия между некоторыми морфотипами сопоставимы с различиями родов карповых рыб (Cyprinidae) Евразии (Nagelkerke et al., 1994). Генерализованными в системе фенетического разнообразия крупных африканских усачей комплекса B. intermedius (sensu Banister, 1973) именуются особи без явных признаков специализации питания. Будучи всеядными, в ареале они вездесущи. Такие особи слагают мономорфные популяции усачей во многих реках и озёрах Эфиопии. В полиморфных популяциях комплекса, в которых вместе со всеядной присутствуют формы (морфотипы) в пищевом отношении в той или иной степени специализированные, генерализованные особи доминируют по численности. Считается, что генерализованная форма близка к предковой, в процессе своей диверсификации давшей начало всему многообразию комплекса B. intermedius (Nagelkerke et al., 1994; Mina et al., 1996a; Лёвин, 2003; Голубцов, 2010; Levin et al., 2019). Гипотеза о близости генерализованной формы к предковой заставляет задуматься о закономерностях диверсификация генерализованных особей, живущих в настоящее время. Одинаковы ли эти закономерности в разных (мономорфных и полиморфных) популяциях? Сходны ли они с закономерностями диверсификации особей в процессе дивергенции морфотипов и видов? Такое сходство, имей оно место, можно было бы рассматривать как очевидное свидетельство в пользу гипотезы о близости (или тождестве) генерализованной и предковой форм.
Цель исследования − оценить сходство векторов диверсификации особей генерализованной формы усачей в разных популяциях с векторами дивергенции морфотипов и видов.
МАТЕРИАЛ И МЕТОДИКА
Материал для исследования собран в трёх озёрах Эфиопии: Тана в 1992–2010 гг., Аваса в 1997−2016 гг. и Лангано в 2007−2011 гг. (рис. 1). Рыб отлавливали жаберными и накидными сетями, а также брали из уловов местных рыбаков. В оз. Тана, расположенном на Эфиопском нагорье, обитает полиморфная популяция крупных африканских усачей; генерализированная форма (далее GF11) этой популяции описана как морфотип intermedius22 (Nagelkerke et al., 1994; Мина и др., 2016). Выборки из озёр Аваса и Лангано, расположенных в Рифтовой долине, рассматриваются как практически мономорфные. Фенетические проявления микроэволюционных процессов в популяциях двух этих озёр едва уловимы (Мина и др., 2016). В оз. Аваса положение рта некоторых особей можно рассматривать как полунижнее, тогда как у подавляющего большинства генерализованных усачей в этом озере рот конечный. Среди усачей из оз. Лангано можно выделить особей относительно высокотелых, изредка встречаются особи с положением рта, близким к нижнему, в 2007 г. была поймана особь, напоминающая морфотип white hunch из оз. Тана, а в 2011 г. – две особи с губами, сходными с губами морфотипа Lip. Результаты анализа остеологических признаков GF обоих озёр провизорно можно трактовать лишь как отражающие едва наметившуюся дивергенцию на две формы: эврифагов и ихтиофагов (Мина и др., 2016). В расчётах использовали особей одного размерного ряда: стандартная длина (SL) от 17 до 29 см; в оз. Тана таких оказалось 119, в оз. Аваса – 45, в оз. Лангано – 50.
Наряду с генерализованными в исследовании рассматриваются усачи специализированных морфотипов из оз. Тана: acute (далее обозначается как ac) – 75 экз., barbel (ba) – 17, bighead (bh) – 11, bigmouth big eye (be) – 56, bigmouth mini eye (me) – 10, bigmouth small eye (se) – 51, carplike (ca) – 12, dark (da) – 35, troutlike (tr) – 41, white hunch (wh) – 48, zurki (zu) – 54, shorthead (sh) – 13, Lip (Li) – 27 экз. Средние оценки признаков определяли с учётом полного объёма выборки каждого морфотипа. Для оценки корреляций между признаками усачей разных морфотипов использованы 14 референсных групп, ранее послуживших основой серии исследований (Мина и др., 2011, 2013; Mina et al., 2012). Каждая такая группа состоит из пяти особей, принадлежность которых к данному морфотипу по диагностическим признакам определялась однозначно. Таким образом, в расчётах корреляций каждый морфотип представлен равным числом особей; размерный ряд модельной совокупности из 70 особей морфотипов равномерно (без выраженных хиатусов) варьирует в пределах SL 11−37 см, т.е. полностью включает в себя размерный ряд выборок GF.
Кроме выборок симпатрических морфотипов из оз. Тана в исследовании использованы выборки четырёх аллопатрических видов крупных африканских усачей: B. ethiopicus Zolezzi, 1939, B. binny (Forsskål, 1775), B. gananensis Vinciguerra, 1895 и B. zaphiri Boulenger, 1906. Как и в случае танских морфотипов, в расчёте корреляций каждый вид представлен пятью особями, размеры которых равномерно (без выраженных хиатусов) варьируют от 12 до 28 см.
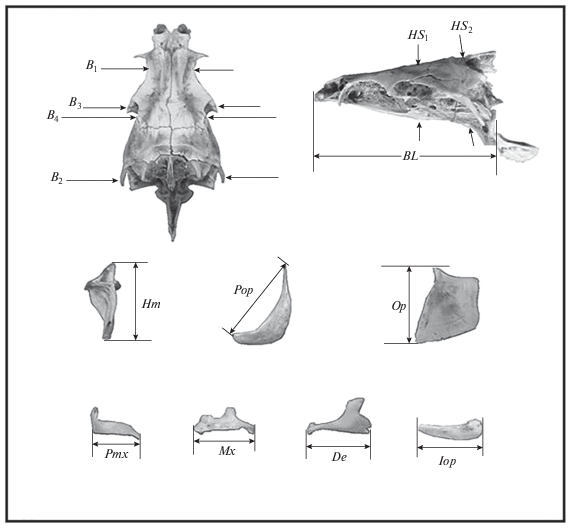
У каждой особи сделаны 14 промеров черепа (рис. 2). Как показали исследования изменчивости крупных африканских усачей, данный набор признаков, обладая высокой разрешающей способностью и хорошей воспроизводимостью результатов измерений как одним, так и разными операторами, позволяет достаточно уверенно оценивать фенетические отношения большинства морфотипов (Mina et al., 1993, 1996а, 1996b, 1998; Мина и др., 2005; Мироновский, 2006).
Рис. 2.
Схема промеров черепа рыб комплекса Barbus intermedius: BL − базальная длина черепа; B1, B2, B3 – расстояние между внешними краями соответственно frontalia, pterotica и sphenotica; B4 – ширина черепа на уровне соединения frontale и pteroticum; HS1, HS2 – высота черепа на уровне соответственно изгиба parasphenoideum и заднего края parasphenoideum; Hm – высота hyomandibulare, Pop − длина praeoperculum, Op – высота передней части operculum; Pmx, Mx, De, Iop – длина соответственно praemaxillare, maxillare, dentale и interoperculum.
Статистическая обработка данных выполнена средствами пакетов NTSYS 2.02k (Rohlf, 1998) и Статистика 6. В расчётах использованы десятичные логарифмы индексов, вычисляемых как отношения абсолютных значений промеров к базальной длине черепа (BL). В анализе главных компонент (АГК) собственные векторы рассчитывали по корреляционной матрице; длину вектора принимали равной 1.
Для анализа корреляционных связей признаков использованы подходы, основанные на идеях разных авторов (Терентьев, 1959; Андреев, Решетников, 1978; Андреев, 1980; Животовский, 1984; Изюмов, 1984; Дгебуадзе и др., 2008), а также на собственном опыте учёта корреляций при изучении структуры фенетического разнообразия Cyprinidae (Мироновский, 1985, 2017). Корреляции между признаками (индексами промеров черепа) вычисляли при помощи коэффициента Пирсона (r). Статистическую значимость коэффициентов корреляции и уравнений регрессии оценивали для уровня α = 0.05, соответственно алгоритмам, указанным в монографиях Плохинского (1980), а также Ивантера и Коросова (2011). Группы скоррелированных признаков выявляли, кластеризуя корреляционные матрицы методом полной связи. Результаты кластерного анализа представлены в виде дендрограмм. Следуя рекомендации Животовского (1984), для признаков, вошедших в разные кластеры дендрограмм, главные компоненты вычисляли раздельно; при построении двумерных распределений по оси абсцисс откладывали значения первой главной компоненты (ГК1) признаков одного кластера, по оси ординат − значения ГК1 признаков другого кластера (Мироновский, 2017).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В процессе морфологической диверсификации признаки особей изменяются не порознь (каждый признак сам по себе), а согласованно, образуя плеяды (Терентьев, 1959) или ансамбли (Андреев, Решетников, 1978) параметров организма, в той или иной степени коррелирующих между собой в процессе формирования внутри- и межпопуляционных различий. Анализ таких корреляций позволил получить ряд важных результатов при изучении изменчивости особей как природных популяций, так и в процессе селекционной работы с сельскохозяйственными растениями и животными (Ростова, 2002). Это даёт основания полагать, что, анализируя структуру корреляционных матриц признаков разных совокупностей, мы сможем сличить особенности процессов диверсификации особей сравниваемых совокупностей и таким образом оценить степень сходства и различий этих процессов. В нашем случае сравниваемые совокупности особей – это выборки GF из озёр Тана, Аваса и Лангано, а также искусственные агломерации особей симпатрических морфотипов из оз. Тана и особей аллопатрических видов крупных африканских усачей. На первом этапе исследования для оценки особенностей диверсификации особей означенных совокупностей проведён кластерный анализ пяти соответствующих матриц корреляций, результаты которого иллюстрируют дендрограммы на рис. 3.
Рис. 3.
Дендрограммы корреляций (r) между признаками особей изучаемых совокупностей африканских усачей комплекса Barbus intermedius: а − модельной агломерации 14 танских морфотипов, достоверны r > 0.240; б − модельной агломерации четырёх аллопатрических видов, достоверны r > 0.444; в − генерализированной формы (GF) оз. Тана, достоверны r > 0.200; г – GF оз. Аваса, достоверны r > 0.280; д – GF оз. Лангано, достоверны r > 0.270. Обозначения признаков см. на рис. 2.
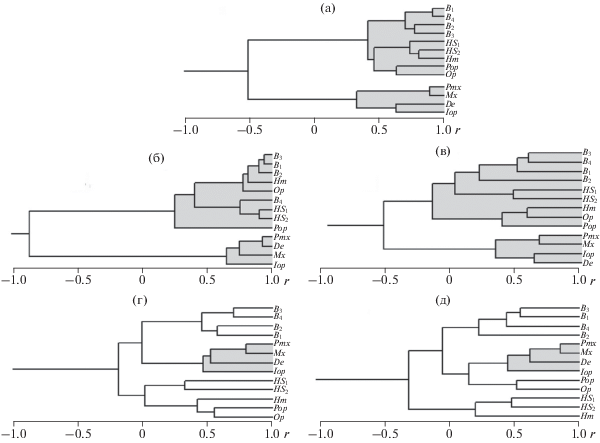
Корреляционные связи между признаками агломерации из 70 усачей 14 танских морфотипов (включая GF) отражает рис. 3а. Чётко выраженные кластеры дендрограммы делят признаки на две группы. В одну входят шесть признаков осевого черепа – расстояние между внешними краями frontalia (B1), pterotica (B2) и sphenotica (B3), ширина черепа на уровне соединения frontale и pteroticum (B4), высота черепа на уровне изгиба parasphenoideum (HS1) и заднего края parasphenoideum (HS2) – и три признака висцерального черепа: высота hyomandibulare (Hm), длина praeoperculum (Pop), высота передней части operculum (Op). Во вторую – четыре признака висцерального черепа, три из которых – длина praemaxillare (Pmx), maxillare (Mx) и dentale (De) – отражают морфологию челюстей, что даёт основания рассматривать их изменчивость как прямо связанную с особенностями питания (Мироновский, 2017). Четвёртый признак второй группы – длина interoperculum (Iop) – характеризует длину межкрышечной кости. В черепе усачей эта кость располагается за зубной, что, видимо, и определяет тесную корреляцию между нею и De. Высокие корреляции внутри каждого кластера дают основание рассматривать выделяемые группы признаков как корреляционные плеяды в понимании Терентьева (1959). Объединение в одной плеяде признаков, прямо связанных с питанием, показывает, что в процессе диверсификации морфотипов они изменяются согласованно, как единое целое. Отрицательная связь между плеядами согласуется со знакомой каждому ихтиологу зависимостью между образом жизни рыб и их обликом (Никольский, 1963). В частности, как правило, чем длиннее челюсти (и больше рот), тем ниже и у́же голова.
Результаты АГК средневыборочных оценок признаков GF и 13 специализированных морфотипов из оз. Тана приведены на рис. 4, собственные векторы и значения, а также доли связанной дисперсии − в табл. 1.
Рис. 4.
Распределение морфотипов африканских усачей комплекса Barbus intermedius оз. Тана (▼) на плоскости главных компонент (ГК): по оси абсцисс – ГК1 признаков плеяды челюстной дуги и interoperculum, по оси ординат – ГК1 признаков плеяды осевого черепа, подъязычной дуги и жаберной крышки; пояснения см. в тексте.
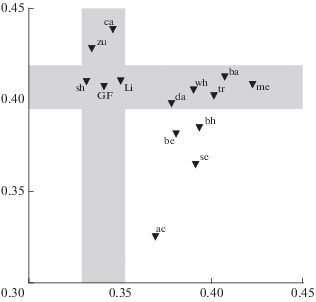
Таблица 1.
Собственные значения и весовые вклады признаков в первые собственные векторы главных компонент корреляционных плеяд признаков морфотипов африканских усачей комплекса Barbus intermedius оз. Тана
| Признак | Собственные векторы | |
|---|---|---|
| СВ1, ось x | СВ1, ось y | |
| B3 | 0.376 | |
| B2 | 0.364 | |
| B1 | 0.355 | |
| B4 | 0.368 | |
| HS1 | 0.309 | |
| HS2 | 0.224 | |
| Hm | 0.374 | |
| Pop | 0.227 | |
| Op | 0.357 | |
| iOp | 0.452 | |
| Pmx | 0.554 | |
| Mx | 0.468 | |
| De | 0.518 | |
| Собственные значения | 3.03 | 6.49 |
| Объяснённая дисперсия, % | 75.9 | 72.2 |
Примечание. Здесь и в табл. 2: B1, B2, B3 – расстояния между внешними краями соответственно frontalia, pterotica и sphenotica; B4 – ширина черепа на уровне соединения frontale и pteroticum; HS1, HS2 – высота черепа на уровне изгиба parasphenoideum и заднего края parasphenoideum, Hm – высота hyomandibulare, Pop − длина praeoperculum, Op – высота передней части operculum; Pmx, Mx, De, Iop – длина соответственно praemaxillare, maxillare, dentale и interoperculum.
Как видим, ось х отражает без малого 76% изменчивости признаков челюстной дуги и Iop, ось y – >72% изменчивости признаков осевого черепа, жаберной крышки и подъязычной дуги. При этом в каждом векторе вклады признаков одинаковы по знаку и близки по абсолютной величине. Из чего следует, что двумерное распределение на рис. 4 с большой полнотой и минимальными искажениями отображает фенетические отношения морфотипов в многомерном пространстве исходных параметров. Затенением на рис. 4 выделены два канала, идущие параллельно осям координат; длина и ширина каждого соотносятся так, что находящиеся в каналах точки-выборки различаются между собой в основном по значениям ГК1 признаков одной плеяды, мало различаясь по значениям ГК1 второй плеяды. В центре пересечения каналов находится выборка GF.
Проанализируем распределение точек на рис. 4, исходя из гипотезы, что генерализованная форма близка к предковой, давшей начало многообразию форм крупных усачей, в рассматриваемом случае – совокупности танских морфотипов. Как видим, лишь zu и ca отличаются от GF главным образом по оси y, тогда как sh, Li, da, wh, tr, ba и me, напротив, от GF отличаются, прежде всего, по оси x. Символы морфотипов bh, be, se и ac лежат вне затенённых каналов, вклады координатных осей в их различия с GF сопоставимы по величине, и только у морфотипа ac вклад оси y существенно выше вклада оси x. Таким образом, раздельный АГК признаков сравниваемых плеяд с использованием ГК1 каждой плеяды в качестве координатной оси наглядно показывает, что в диверсификацию морфотипов крупных усачей оз. Тана основной вклад вносят параметры, прямо связанные с процессом захвата и удержания пищевых объектов. Это хорошо согласуется с мнением об определяющей роли разделения пищевых ресурсов в формировании пучка форм оз. Тана (Sibbing et al., 1998; Sibbing, Nagelkerke, 2001).
Дендрограмма, отражающая структуру корреляций между признаками 20 особей агломерации четырёх видов крупных африканских усачей, приведена на рис. 3б; её топография аналогична таковой дендрограммы на рис 3а. И там, и там признаки образуют две группы, состав групп совпадает, корреляции внутри групп – положительны и достаточно высоки, чтобы рассматривать группы как корреляционные плеяды. Между плеядами корреляция отрицательна. Сходство топографии дендрограмм свидетельствует о сходстве закономерностей диверсификации особей симпатрических морфотипов оз. Тана и аллопатрических видов.
Два кластера отчётливо обособлены и на дендрограмме, отражающей корреляционные связи признаков особей GF из оз. Тана (рис. 3в), их состав полностью совпадает с таковым на рис. 3а и 3б. Есть, однако, и разница: если на рис. 3а и 3б корреляции между признаками B1,,B2, B3, B4, HS1, HS2, Hm, Pop и Op достаточно высоки, чтобы рассматривать их в качестве корреляционной плеяды, то на рис. 3в уровень корреляций между ними много ниже, и в некоторых ветвлениях древа от нуля отличается не значимо. Таким образом, здесь говорить можно лишь о группе девяти признаков, составивших кластер, отрицательно коррелирующий с кластером челюстной дуги и Iop. У усачей из озёр Аваса и Лангано (рис. 3г, 3д) означенные девять признаков общий кластер не формируют: часть их вначале кластеризуется с плеядой челюстной дуги и уже вместе с ней объединяется с остальными.
Таким образом, если плеяда признаков челюстной дуги и Iop формируется во всех пяти анализируемых совокупностях, то прочие девять признаков в общий противостоящий ей кластер объединяются не всегда. Именно это – противостояние общего кластера девяти прямо не связанных с питанием признаков и кластера признаков челюстной дуги и Iop − отличает диверсификацию GF в оз. Тана от диверсификации GF в мономорфных популяциях, одновременно сближая её с диверсификацией танских морфотипов и аллопатрических видов усачей. На рис. 3а–3в плеяда челюстной дуги противостоит кластеру девяти прочих признаков, на рис. 3г и 3д девять прочих признаков разделены на два кластера, среди которых морфология челюстей, образно говоря, теряется.
Продолжим анализ, развивая и адаптируя к задачам настоящего исследования подход, использованный при изучении изменчивости популяций Coregonidae (Андреев, Решетников, 1978; Андреев, 1980). В табл. 2 приведены наиболее весомые собственные векторы (СВ) и собственные значения корреляционных матриц, по которым строились дендрограммы на рис. 3. В качестве наиболее весомых рассматриваются векторы с собственными значениями >1. В выборках GF таких оказалось по четыре, в агломерациях особей морфотипов и видов − по два. Чтобы сравнить диверсификацию особей разных совокупностей, на осях системы прямоугольных координат отложим значения признаковых коэффициентов одноранговых векторов (СВ1) той и другой совокупности. Если направления изменчивости в сравниваемых совокупностях одинаковы (т.е. диверсификационные процессы совпадают), точки, соответствующие значениям коэффициентов СВ, лягут на прямую, расположенную под углом 45° к осям координат, и корреляция между коэффициентами сравниваемых векторов будет равна 1. Картина, весьма близкая к описанной, имеет место при сравнении СВ1 матриц корреляций признаков танских морфотипов и аллопатрических видов: точки-признаки вытянулись вдоль линии регрессии, её угол наклона ~47°, r = 0.989 (рис. 5а, табл. 3). При сравнении СВ1 агломерации танских морфотипов и СВ1 совокупности особей GF из Таны (рис. 5б, табл. 3) угол наклона на 1° больше, коэффициент корреляции на 0.027 меньше, т.е. в целом распределение аналогично предыдущему. Примечательно, что сравнение корреляционной матрицы GF из Таны с матрицей аллопатричных видов (рис. 5в, табл. 3) выявляет даже бóльшую близость первых собственных векторов, чем предыдущее её сравнение с симпатричными морфотипами. При сравнении GF из Таны, танских морфотипов и аллопатрических видов с выборкой GF из оз. Лангано коэффициенты корреляции не столь высоки, но достоверны и отрицательны (рис. 5г–5е; табл. 3). Это свидетельствует о том, что в популяции GF из Лангано процессы диверсификации по первому вектору изменчивости во многом противоположны соответствующим процессам в трёх сопоставляемых с нею совокупностях. О сходстве процессов диверсификации GF оз. Тана, танских морфотипов и аллопатических видов с процессами диверсификации GF оз. Аваса уверенно судить сложно: на рис. 5ж–5и, где сравниваются первые векторы соответствующих матриц, линии регрессии почти параллельны оси х, коэффициенты корреляций от нуля отличаются недостоверно (табл. 3). И, наконец, результаты анализа рис. 5к и табл. 3 не оставляют сомнений в сходстве двух первых собственных векторов корреляционных матриц признаков особей GF из озёр Аваса и Лангано.
Используя данные табл. 3 как матрицу корреляций между вкладами признаков в сравниваемые СВ1, проведём кластерный анализ и построим дендрограмму (рис. 6а), резюмирующую результаты анализа рис. 5. Как видим, совокупность выборок делится на два кластера: в один входят выборки GF и морфотипов из Таны и выборка аллопатрических видов, во второй − выборки GF из двух мономорфных популяций. Корреляции внутри первого кластера заметно выше, чем внутри второго; между кластерами корреляция отрицательна.
Таблица 2.
Собственные векторы матриц корреляций между признаками пяти совокупностей африканских усачей комплекса Barbus intermedius
| Признак | Корреляционные матрицы между признаками особей | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| морфотипов | видов | генерализованной формы | ||||||||||||||
| оз. Taна | оз. Аваса | оз. Лангано | ||||||||||||||
| СВ1 | СВ2 | СВ1 | СВ2 | СВ1 | СВ2 | СВ3 | СВ4 | СВ1 | СВ2 | СВ3 | СВ4 | СВ1 | СВ2 | СВ3 | СВ4 | |
| B3 | 0.330 | 0.065 | 0.315 | 0.144 | 0.300 | 0.113 | 0.354 | 0.347 | 0.268 | 0.389 | 0.121 | 0.180 | 0.317 | 0.223 | 0.284 | 0.391 |
| B2 | 0.331 | 0.139 | 0.288 | 0.281 | 0.203 | 0.292 | –0.161 | 0.209 | 0.270 | 0.291 | 0.081 | 0.128 | 0.270 | 0.286 | 0.052 | 0.204 |
| B1 | 0.317 | 0.121 | 0.297 | 0.250 | 0.320 | 0.213 | 0.126 | 0.403 | 0.266 | 0.309 | 0.339 | –0.149 | 0.324 | 0.308 | 0.156 | –0.081 |
| B4 | 0.347 | 0.017 | 0.314 | 0.029 | 0.309 | 0.144 | 0.401 | 0.125 | 0.368 | 0.258 | 0.119 | –0.008 | 0.267 | 0.344 | 0.261 | –0.045 |
| HS1 | 0.309 | –0.115 | 0.292 | –0.194 | 0.332 | –0.063 | 0.307 | –0.466 | –0.026 | 0.060 | –0.531 | –0.550 | 0.001 | 0.219 | –0.349 | –0.575 |
| HS2 | 0.341 | 0.046 | 0.328 | 0.0004 | 0.340 | 0.107 | 0.009 | –0.133 | 0.247 | 0.188 | –0.171 | –0.585 | 0.155 | 0.401 | –0.018 | –0.496 |
| Hm | 0.341 | 0.123 | 0.327 | 0.072 | 0.283 | 0.306 | –0.323 | –0.203 | 0.217 | 0.257 | –0.440 | 0.226 | 0.014 | 0.283 | –0.407 | 0.253 |
| Pmx | –0.170 | 0.465 | –0.274 | 0.340 | –0.369 | 0.317 | 0.032 | 0.174 | 0.326 | –0.309 | 0.179 | –0.095 | 0.353 | –0.336 | –0.009 | –0.191 |
| Pop | 0.129 | 0.369 | 0.149 | 0.434 | 0.011 | 0.422 | 0 | –0.521 | 0.229 | –0.144 | –0.490 | 0.408 | 0.253 | 0.096 | –0.466 | 0.199 |
| Op | 0.269 | 0.322 | 0.261 | 0.346 | 0.086 | 0.429 | –0.348 | –0.019 | 0.385 | –0.087 | –0.172 | 0.103 | 0.183 | –0.028 | –0.530 | 0.235 |
| Iop | –0.213 | 0.352 | –0.234 | 0.325 | –0.246 | 0.264 | 0.448 | –0.150 | 0.253 | –0.373 | –0.038 | 0.085 | 0.388 | –0.283 | 0.144 | –0.039 |
| Mx | –0.061 | 0.494 | –0.191 | 0.422 | –0.271 | 0.385 | –0.055 | 0.210 | 0.308 | –0.384 | 0.031 | –0.130 | 0.424 | –0.263 | –0.047 | –0.128 |
| De | –0.256 | 0.326 | –0.274 | 0.296 | –0.296 | 0.213 | 0.385 | –0.117 | 0.274 | –0.294 | 0.206 | –0.155 | 0.273 | –0.310 | –0.124 | –0.087 |
| Собственные значения | 7.05 | 3.08 | 8.38 | 2.73 | 3.70 | 2.58 | 1.78 | 1.19 | 4.59 | 2.17 | 1.58 | 1.14 | 3.70 | 2.42 | 1.51 | 1.30 |
| Объяснённая дисперсия, % | 54.3 | 23.7 | 64.5 | 21.0 | 28.5 | 19.9 | 13.7 | 9.2 | 35.3 | 16.7 | 12.2 | 8.8 | 28.4 | 18.7 | 11.6 | 10.0 |
Рис. 5.
Анализ сходства первых собственных векторов (СВ1) матриц корреляций между признаками особей рассматриваемых совокупностей африканских усачей комплекса Barbus intermedius: а − танские морфотипы и аллопатрические виды, б − танские морфотипы и генерализированная форма (GF) оз. Тана, в − аллопатрические виды и GF оз. Тана, г − танские морфотипы и GF оз. Лангано, д − аллопатрические виды и GF оз. Лангано, е − GF оз. Тана и GF оз. Лангано, ж − танские морфотипы и GF оз. Аваса, з − аллопатрические виды и GF оз. Аваса, и − GF оз. Тана и GF оз. Аваса, к − GF оз. Аваса и GF оз. Лангано.
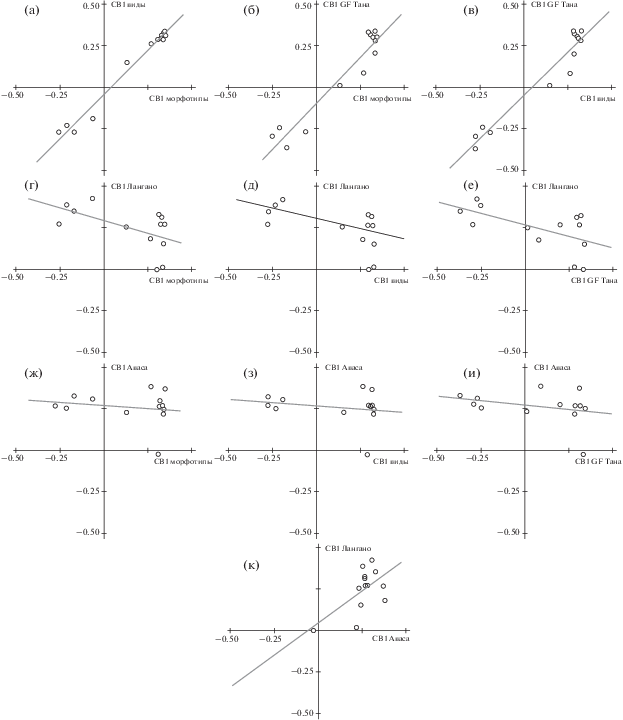
Таблица 3.
Параметры линий регрессии при сравнении одноранговых собственных векторов матриц корреляций между признаками особей пяти рассматриваемых совокупностей африканских усачей комплекса Barbus intermedius
| Выборка | СВ1 | СВ2 | CВ3 | СВ4 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Морфотип | Вид | GF Тана | GF Аваса | Морфотип | Вид | GF Тана | GF Аваса | GF Тана | GF. Аваса | GF Тана | GF Аваса | |
| Вид | $\frac{{{\mathbf{0}}.{\mathbf{989}}}}{{{\mathbf{1}}.{\mathbf{070}}}}$ | $\frac{{{\mathbf{0}}.{\mathbf{905}}}}{{{\mathbf{0}}.{\mathbf{896}}}}$ | ||||||||||
| GF Тана | $\frac{{{\mathbf{0}}.{\mathbf{962}}}}{{{\mathbf{1}}.{\mathbf{114}}}}$ | $\frac{{{\mathbf{0}}.{\mathbf{973}}}}{{{\mathbf{1}}.{\mathbf{043}}}}$ | $\frac{{{\mathbf{0}}.{\mathbf{816}}}}{{{\mathbf{0}}.{\mathbf{610}}}}$ | $\frac{{{\mathbf{0}}.{\mathbf{872}}}}{{~{\mathbf{0}}.{\mathbf{659}}}}$ | ||||||||
| GF Аваса | $\frac{{ - 0.173}}{{ - 0.072}}$ | $\frac{{ - 0.203}}{{ - 0.078}}$ | $\frac{{ - 0.306}}{{ - 0.110}}$ | $\frac{{ - {\mathbf{0}}.{\mathbf{817}}}}{{ - {\mathbf{1}}.{\mathbf{247}}}}$ | $\frac{{ - {\mathbf{0}}.{\mathbf{575}}}}{{ - {\mathbf{0}}.{\mathbf{886}}}}$ | $\frac{{ - 0.444}}{{ - 0.906}}$ | $\frac{{0.316}}{{0.327}}$ | $\frac{{0.059}}{{0.058}}$ | ||||
| GF Лангано | $\frac{{ - {\mathbf{0}}.{\mathbf{558}}}}{{ - {\mathbf{0}}.{\mathbf{303}}}}$ | $\frac{{ - {\mathbf{0}}.{\mathbf{596}}}}{{ - {\mathbf{0}}.{\mathbf{299}}}}$ | $\frac{{ - {\mathbf{0}}.{\mathbf{597}}}}{{ - {\mathbf{0}}.{\mathbf{280}}}}$ | $\frac{{{\mathbf{0}}.{\mathbf{591}}}}{{{\mathbf{0}}.{\mathbf{771}}}}$ | $\frac{{ - {\mathbf{0}}.{\mathbf{841}}}}{{ - {\mathbf{1}}.{\mathbf{240}}}}$ | $\frac{{ - {\mathbf{0}}.{\mathbf{635}}}}{{~ - {\mathbf{0}}.{\mathbf{945}}}}$ | $\frac{{ - 0.439}}{{ - 0.865}}$ | $\frac{{{\mathbf{0}}.{\mathbf{926}}}}{{{\mathbf{0}}.{\mathbf{895}}}}$ | $\frac{{{\mathbf{0}}.{\mathbf{597}}}}{{{\mathbf{0}}.{\mathbf{602}}}}$ | $\frac{{{\mathbf{0}}.{\mathbf{763}}}}{{{\mathbf{0}}.{\mathbf{747}}}}$ | $\frac{{0.268}}{{0.267}}$ | $\frac{{{\mathbf{0}}.{\mathbf{922}}}}{{{\mathbf{0}}.{\mathbf{928}}}}$ |
Представляется, что в обсуждаемом аспекте о сходстве собственных векторов корреляционных матриц достаточно уверенно можно судить на основании данных табл. 3, и двумерные распределения на рис. 5 приведены главным образом для графической иллюстрации сути используемого подхода. Поэтому, сопоставляя диверсификацию рассматриваемых совокупностей особей по вторым, третьим и четвёртым собственным векторам, ограничимся анализом данных табл. 3.
Рис. 6.
Сходство процессов диверсификации пяти сравниваемых совокупностей африканских усачей комплекса Barbus intermedius: а − по первому собственному вектору (СВ1), достоверны r > 0.553; б − по второму собственному вектору (СВ2), достоверны r > > 0.553.
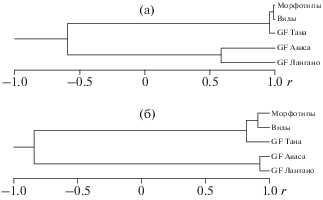
Как видим, сходство диверсификации пяти рассматриваемых совокупностей особей по СВ2 в целом соответствует их сходству по СВ1 (табл. 3). Весьма сходна и топология дендрограмм на рис. 6а и 6б, разница в том, что по СВ2 выборки из мономорфных популяций между собой коррелируют теснее, чем коррелируют между собой обе выборки из оз. Тана и аллопатрических видов – по СВ1 было наоборот. Это позволяет сказать, что если по первому направлению диверсификации между мономорфными популяциями усачей Авасы и Лангано некоторые различия наметились, то по второму направлению различия, видимо, минимальны. По СВ3 и СВ4, данные по которым в табл. 3 приведены только для трёх совокупностей особей GF, отрицательных корреляций между сравниваемыми выборками нет. Между выборками из Лангано и Авасы корреляции по обоим векторам высоки и достоверны, корреляция между выборками из Таны и Авасы недостоверна, но достоверна корреляция по СВ3 между выборками из Таны и Лангано.
Как следует из табл. 2, два первых собственных вектора диверсификации особей модельной совокупности танских морфотипов объясняют без малого 78% общей изменчивости рассматриваемых признаков. В модельной совокупности аллопатрических видов доля общей дисперсии, вкупе объясняемой СВ1 и СВ2, превышает 85%. Сопоставление пар одноранговых векторов показывает, что в модельных совокупностях они почти совпадают по направлению, и корреляция между ними близка к 1 (рис. 5а, табл. 3). Это даёт веские основания полагать, что в данных совокупностях два первых вектора дисперсии отражают одни и те же направления диверсификации. При этом в обеих модельных совокупностях собственные значения лишь СВ1 и СВ2 > 1, т.е., согласно правилу Кайзера (Kaiser, 1960), упорядоченная, направленная изменчивость (сигнал) сосредоточена именно в них. В оставшихся 11 векторах (чьи собственные значения <1) преобладает изменчивость стохастическая, случайная (шум). Из чего следует, что два первых вектора практически полностью определяют направленную конструктивную изменчивость, обусловливающую дивергенцию рассматриваемых морфотипов и видов.
Сопоставление СВ1 и СВ2 изменчивости морфотипов и видов с соответствующими векторами матрицы корреляций особей GF из оз. Тана (рис. 5б, 5в; табл. 3) выявляет уровень совпадения сравниваемых параметров почти столь же высокий, как в аналогичном сопоставлении векторов морфотипов и видов между собой. Из чего следует, что каждое из двух выявленных направлений дивергенции морфотипов и видов следует вдоль направлений, заданных двумя направлениями диверсификации генерализованных особей из Таны. Это, очевидно, можно рассматривать как весомое свидетельство в пользу гипотезы о близости GF из оз. Тана к форме предковой, давшей начало многообразию морфотипов и видов крупных африканских усачей, причём “близость” − это в отношении аллопатричных ей видов. В отношении симпатричных морфотипов GF из Таны, видимо, тождественна предковой форме, т.е. не исключено, что она ею и является. Различия структуры корреляций в совокупностях особей GF из Таны, танских морфотипов и аллопатических видов, с одной стороны, и особей GF из озёр Аваса и Лангано − с другой, показывает, что главные направления диверсификации в мономорфных совокупностях существенно отличаются от таковых в совокупностях полиморфных. Именно эти различия, видимо, составляют разницу между популяциями GF, дающими начало разнообразию форм, и популяциями, где такого разнообразия нет.
Следует отметить, что в случае GF из Таны, Авасы и Лангано единицу превышают собственные значения не только СВ1 и СВ2, но также СВ3 и СВ4 (табл. 2). В Тане они отражают без малого 23% общей изменчивости, в Авасе и в Лонгано примерно по 21%, причём отрицательные корреляции между одноранговыми парами СВ3 и СВ4 в трёх озёрах не выявлены. Для оценки роли этих двух направлений в микроэволюции усачей необходимо специальное исследование, в частности, анализ вкладов в собственные векторы матриц корреляций не только 13 рассматриваемых признаков вкупе, но и парциальных вкладов групп признаков, образующих кластеры на дендрограммах (рис. 3). Пока можно лишь предположить, что третье и четвёртое направления изменчивости второстепенны и либо вовсе не ведут к образования новых форм, либо формы, дивергировавшие в этих направлениях, крайне малочисленны и пока в поле зрения исследователей не попали.
В рамках представления о генерализованных особях как о форме, близкой к предковой, давшей начало всему многообразию комплекса B. intermedius, полученные результаты позволяют рассматривать диверсификацию GF в оз. Тана как основу фенетического разнообразия обитающих здесь форм крупных африканских усачей. Неясным остаётся вопрос о размахе и пределах диверсификации GF в настоящее время: ставши некогда основой фенетического разнообразия усачей оз. Тана, остаётся ли она его источником сейчас? Есть ли в потомстве генерализованных усачей особи с внешними признаками специализированных морфотипов или же, дивергировав некогда от GF, ныне каждый морфотип воспроизводит себя сам? Ответ на эти вопросы может дать продолжение исследований, в частности, оценка корреляционных связей признаков в процессе диверсификации не только GF, но и особей каждого специализированного морфотипа усачей Таны в отдельности.
ВЫВОДЫ
1. В процессе морфологической диверсификации признаки особей генерализованной формы, морфотипов и видов крупных африканских усачей изменяются не порознь (каждый признак сам по себе), а согласованно, образуя кластеры параметров, в той или иной степени между собой коррелирующих в процессе формирования внутри- и межгрупповых различий. Некоторые из таких кластеров могут рассматриваться как корреляционные плеяды признаков в понимании Терентьева (1959).
2. Состав выявленных плеяд и уровень их различий между собой не одинаков в рассмотренных совокупностях особей. При сравнении диверсификации усачей GF из оз. Тана, морфотипов и видов уровень различий и состав плеяд весьма сходны, но существенно отличаются от состава и уровня различий плеяд, возникающих при диверсификации GF мономорфных популяций озёр Аваса и Лангано.
3. В каждой из трёх рассмотренных совокупностей генерализованных усачей выявлено четыре главных вектора диверсификации, у морфотипов и видов главных векторов диверсификации − по два.
4. При сравнении мономорфных популяций озёр Лангано и Аваса первый и второй главные векторы диверсификации генерализованных особей весьма сходны и существенно отличаются от соответствующих векторов диверсификации генерализованных особей в полиморфной популяции оз. Тана. По третьим и четвёртым главным векторам существенных различий между диверсификацией особей в полиморфной и в мономорфных популяциях не выявлено.
5. Главные векторы диверсификации танских морфотипов и аллопатрических видов сходны как между собой, так и с двумя соответствующими векторами диверсификации генерализованных особей из оз. Тана. Высокая степень этого сходства даёт основания полагать, что диверсификация генерализованных особей в оз. Тана, диверсификация особей танских морфотипов и аллопатрических видов – это последовательные этапы одного процесса, и дивергенция морфотипов, а затем видов идёт в направлениях, заданных диверсификацией особей генерализованной формы.
6. Результаты исследования можно рассматривать как веское свидетельство в пользу гипотезы о близости (или тождестве) генерализованной формы усачей и формы предковой, давшей начало всему разнообразию комплекса B. intermedius (sensu Banister, 1973).
Список литературы
Андреев В.Л. 1980. Классификационные построения в экологии и систематике. М.: Наука, 142 с.
Андреев В.Л., Решетников Ю.С. 1978. Анализ фенотипической изменчивости географически удалённых популяций одного вида // Математические методы в экологии и географии. Владивосток: ДВНЦ АН СССР. С. 98−110.
Голубцов А.С. 2010. “Пучки видов” рыб в реках и озерах: симпатрическая дивергенция в фаунистически обеднённых рыбных сообществах как особый модус эволюции // Актуальные проблемы современной ихтиологии. К 100-летию Г.В. Никольского. М.: Т-во науч. изд. КМК. С. 96–123.
Дгебуадзе Ю.Ю., Мина М.В., Мироновский А.Н. 2008. К оценке фенетических отношений алтайских османов (Oreoleuciscus, Cyprinidae) из трех озер Монголии по признакам черепа // Вопр. ихтиологии. Т. 48. № 3. С. 315−323.
Животовский Л.А. 1984. Интеграция полигенных систем в популяциях: проблемы анализа комплекса признаков. М.: Наука, 183 с.
Ивантер Э.В., Коросов А.В. 2011. Введение в количественную биологию. Петрозаводск: Изд-во ПетрГУ, 302 с.
Изюмов Ю.Г. 1984. Популяционная структура леща Abramis brama (L.) волжских водохранилищ // Биологические ресурсы водохранилищ. М.: Наука. С. 227−242.
Лёвин Б.А. 2003. Структура локальных популяционных систем у рыб рода Barbus // Матер. ІІ міжнар. наук. конф. “Біорізноманіття та роль зооценозу в природних і антропогенних екосистемах”. Днепропетровск: Изд-во ДНУ. С. 53−55.
Мина М.В., Лёвин Б.А., Мироновский А.Н. 2005. О возможностях использования в морфометрических исследованиях рыб оценок признаков, полученных разными операторами // Вопр. ихтиологии. Т. 45. № 3. С. 331−341.
Мина М.В., Мироновский А.Н., Капитанова Д.В. 2011. Фенетические отношения и вероятные пути морфологической диверсификации африканских усачей комплекса Barbus intermedius из озера Тана (Эфиопия) // Там же. Т. 51. № 2. С. 149−163.
Мина М.В., Дзержинский К.Ф, Мироновский А.Н., Капитанова Д.В. 2013. Фенетическое разнообразие крупных усачей Barbus intermedius complex sensu Banister из прибрежной зоны Бахардарского залива (озеро Тана, Эфиопия) // Там же. Т. 53. № 6. С. 627−644. https://doi.org/10.7868/S0042875213050081
Мина М.В., Мироновский А.Н., Дгебуадзе Ю.Ю. 2016. Полиморфизм по пропорциям черепа у крупных африканских усачей Barbus intermedius sensu Banister, 1973 (Cyprinidae) из озер Аваса и Лангано (Рифтовая долина, Эфиопия) // Там же. Т. 56. № 4. С. 403−409. https://doi.org/10.7868/S0042875216040081
Мироновский А.Н. 1985. К использованию скоррелированных признаков в популяционных исследованиях // Биология внутрен. вод. № 68. С. 64−68.
Мироновский А.Н. 2006. Факторы, обуславливающие сопоставимость данных, полученных путем оценки пластических признаков рыб // Вопр. ихтиологии. Т. 46. № 2. С. 240−251.
Мироновский А.Н. 2017. Место генерализованной формы в структуре фенетического разнообразия крупных африканских усачей комплекса Barbus intermedius. Метод корреляционных плеяд // Там же. Т. 57. № 4. С. 393−404. https://doi.org/10.7868/S0042875217040105
Никольский Г.В. 1963. Экология рыб. М.: Высш. шк. С. 368.
Плохинский Н.П. 1980. Алгоритмы биометрии. М.: Изд-во МГУ, 150 с.
Ростова Н.С. 2002. Корреляции: структура и изменчивость. СПб.: Изд-во СПбГУ, 308 с.
Терентьев П.В. 1959. Метод корреляционных плеяд // Вестн. ЛГУ. № 9. С. 137–141.
Banister K.E. 1973. A revision of the large Barbus (Pisces, Cyprinidae) of East and Central Africa. Pt. II // Bull. Brit. Mus. Nat. Hist. Zool. V. 26. P. 1–148.
Kaiser H.F. 1960. The application of electronic computers to factor analysis // Educ. Psychol. Measmt. V. 20. № 1. P. 141−151.
Levin B.A., Casal-López M., Simonov E. et al. 2019. Adaptive radiation of barbs of the genus Labeobarbus (Cyprinidae) in the East African river // Freshwater Biol. V. 64. Iss. 10. P. 1721–1736. https://doi.org/10.1111/fwb.13364
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. 1993. Alometria et divergence entre les barbeaux du lac Tana (Ethiopie) // Cahiers d’Ethologie. V. 13. P. 219–222.
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. 1996a. Morphometry of barbel of Lake Tana, Ethiopia: multivariate ontogenetic channels // Folia Zool. V. 45. Suppl. 1. P. 109–116.
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. 1996b. Lake Tana large barbs: phenetics, growth and diversification // J. Fish Biol. V. 48. P. 383–404.
Mina M.V., Mironovsky A.N., Golubtsov A.S., Dgebuadze Yu.Yu. 1998. ‘Barbus’ intermedius species flock in Lake Tana (Ethiopia). II. Morphological diversity of “large barbs” from Lake Tana and neighbouring areas: homoplasies or synapomorphies? // Ital. J. Zool. V. 65. Suppl. P. 9–14.
Mina M.V., Shkil F.N., Dzerzhinskii K.F. et al. 2012. Morphological diversity and age dependent transformations in progeny of the large barbs (Barbus intermedius complex sensu Banister) of several morphotypes from Lake Tana (Ethiopia). Results of a longtime experiment // J. Ichthyol. V. 52. № 11. P. 821–837. https://doi.org/10.1134/S0032945212110070
Nagelkerke L.A.J., Sibbing F.A., van den Boogaart J.G.M. et al. 1994. The barbs (Barbus spp.) of Lake Tana: a forgotten species flock? // Environ. Biol. Fish. V. 39. P. 1–21.
Rohlf F.J. 1998. NTSYS-pc: numerical taxonomy and multivariate analysis system. Version 2.0. N.Y.: Exeter software, 31 p.
Sibbing F.A., Nagelkerke L.A.J. 2001. Resource partitioning by Lake Tana barbs predicted from fish morphometrics and prey characteristics // Rev. Fish Biol. Fish. V. 10. № 4. P. 393–437.
Sibbing F.A., Nagelkerke L.A.J., Stet R.J.M., Osse J.W.M. 1998. Speciation of endemic Lake Tana barbs (Cyprinidae, Ethiopia) driven by trophic resource partitioning; a molecular and ecomorphological approach // Aquat. Ecol. V. 32. P. 217–227.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии