

Вопросы ихтиологии, 2020, T. 60, № 3, стр. 251-260
О некоторых малоизученных Melanostomiidae с описанием нового вида Eustomias
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН – ИПЭЭ РАН
Москва, Россия
2 Институт океанологии им. П.П. Ширшова РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 14.09.2019
После доработки 04.10.2019
Принята к публикации 06.11.2019
Аннотация
Описан новый вид Eustomias mikhailini sp. nova, близкий к E. macrura. Предложено восстановить подрод Urostomias из синонимии подрода Spilostomias. Приведена определительная таблица подродов рода Eustomias с большим числом лучей грудного плавника и видов подрода Urostomias. Описан экземпляр Eustomias gr. melanostigma, который в структуре подбородочного усика сочетает признаки разных видов данной группы, что вызывает сомнения в обоснованности выделения E. melanonema и E. melanostigmoides. Сообщено о новых нахождениях редких видов Leptostomias cf. bilobatus и Photonectes xenopogon, впервые обнаруженных в юго-восточной части Тихого океана; приведены подробные морфологические описания экземпляров; уточнены пределы изменчивости диагностически значимых признаков.
Стомиеобразные рыбы являются одной из доминирующих групп в океанской мезо- и отчасти батипелагиали. Наиболее продвинутых представителей этого отряда можно обособить в семейство Melanostomiidae, характеризующееся более или менее удлинённым телом с голой подвижной кожей, смещёнными назад супротивными вертикальными плавниками; как правило, развитым подбородочным усиком, концевая часть которого в большинстве случаев образует специфические структуры, играющие важную роль в дифференциации видов, и рядом анатомических признаков (Fink, 1985). В современной систематике (Nelson, 2006; Nelson et al., 2015; Fricke et al., 2018) меланостомовые обычно рассматриваются в ранге подсемейства широко понимаемого семейства Stomiidae sensu Fink (1985), по своему составу соответствующего подотрядам Lepidophotodermi и Gymnophotodermi Парра (Parr, 1927). Однако Melanostomiidae (включая ранее выделявшиеся семейства Idiacanthidae и Malacosteidae) являются монофилетической кроновой группой, морфологически хорошо отличимой от других “классических” семейств, также включаемых в состав Stomiidae sensu Fink (1985) (Astronesthidae, Chauliodontidae и Stomiidae s. str.), и оснований для выделения их в ранг семейства не меньше, чем для выделения самого семейства Stomiidae sensu Fink (1985), поскольку отношения последнего с предковым семейством Phosichthyidae также представляют собой граду, а не клады (Harold, Weitzman, 1996). Меланостомовые всё ещё продолжают оставаться недостаточно изученной в систематическом отношении группой, и видовой состав многих родов до сих пор окончательно не установлен. Ряд видов известны по единичным поимкам, и их ареалы пока тоже остаются невыясненными. В связи с этим представляет интерес публикация сведений о новых нахождениях за пределами ранее известных ареалов и о морфологической изменчивости трёх малоизученных видов меланостомовых рыб из родов Eustomias Vaillant, 1888, Leptostomias Gilbert, 1905 и Photonectes Günther, 1887, обнаруженных мною в коллекциях Института океанологии РАН (ИО РАН) и Парижского национального музея естественной истории (MNHN). Кроме того, в статье описывается своеобразный новый вид Eustomias, голотип и единственный известный экземпляр которого ранее уже упоминался в литературе (Парин и др., 1978) в открытой номенклатуре. Этот экземпляр в своё время отсылали ныне покойному Р. Гиббсу, США (R.H. Gibbs), подтвердившему его принадлежность к новому для науки виду.
МАТЕРИАЛ И МЕТОДИКА
Методика изучения и терминология соответствуют общепринятым (Morrow, Gibbs, 1964; Gibbs, 1968; Gibbs et al., 1983; Прокофьев, 2018, 2019). В случае отличий в счётных значениях одних и тех же признаков на разных сторонах тела одной рыбы показатели разделены знаком “/”. В работе использованы следующие сокращения: SL – стандартная длина; D, A, P, V, С – соответственно спинной, анальный, грудные, брюшные и хвостовой плавники; vert. – общее число позвонков; э/с – экспедиционное судно, ст. – океанографическая станция. Используется стандартная номенклатура фотофоров стомиеобразных рыб (Morrow, Gibbs, 1964) и структур подбородочного усика (Gibbs et al., 1983; Прокофьев, 2018).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
(рис. 1)
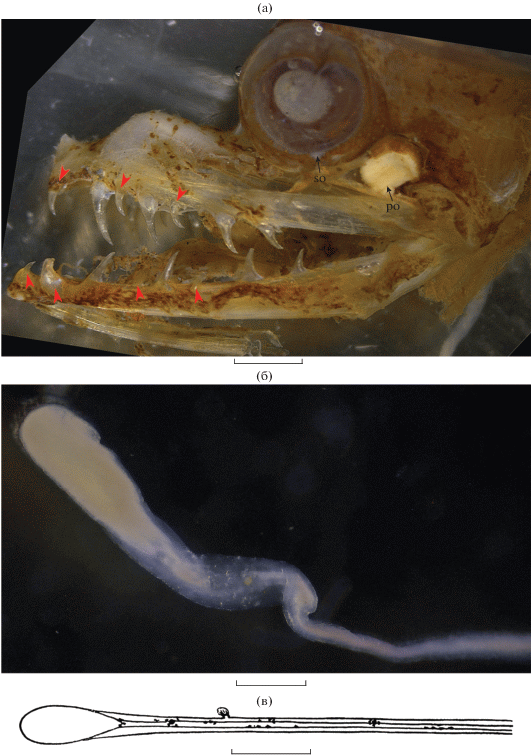
Рис. 1.
Eustomias mikhailini sp. nova, голотип SL 115 мм, экваториальная Восточная Атлантика: а – озубление челюстей и окологлазничные светящиеся органы; б, в (рисунок Г.Н. Похильской) – дистальный конец подбородочного усика. (▶) – фиксированные зубы; so, po – суборбитальный и посторбитальный органы. Масштаб: а – 1.5, б – 0.4, в – 1.0 мм.
Eustomias sp.: Парин и др., 1978. С. 172. Рис. 2.
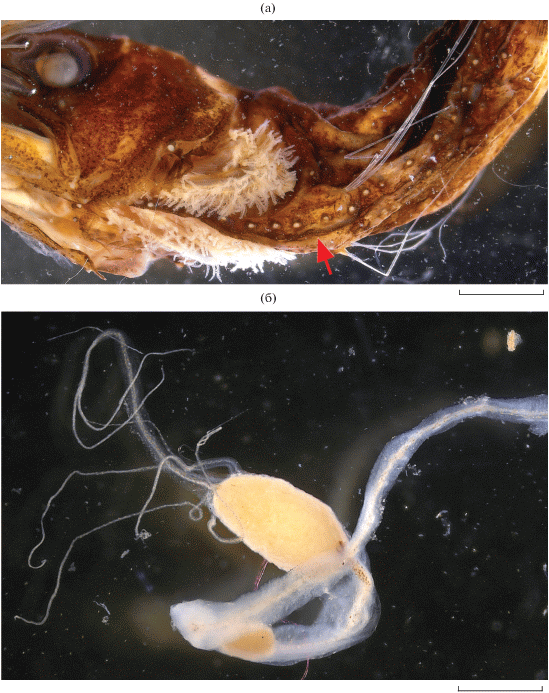
Рис. 2.
Eustomias gr. melanostigma SL 74 мм, экваториальная Западная Пацифика: а – рудимент брюшной бороздки (→), б – дистальный конец подбородочного усика. Масштаб: а – 2, б – 1 мм.
Материал. Голотип, ИО РАН № 03599, SL 115 мм, 00°15′ ю.ш., 06°36′ в.д., пелагический трал, горизонт траления 25 м, э/с “Фиолент”, рейс ФАО-1, 06.01.1976 г. Парин с соавт. (1978) приводят для данного экземпляра SL 118 мм. Отмечаемое мною изменение пропорций, очевидно, связано с дегидратацией ткани после более 40 лет хранения в спирте.
Диагноз. Вид рода Eustomias с семью лучами Р и V, с подбородочным усиком простого строения (с одной крупной овальной терминальной луковицей без придатков), со светящейся железой на верхних дополнительных лучах С.
Описание. D 26, A 41, P 7, V 7. Фотофоров в сериях: IP 7, PV 27, OV 27, VAV 13 (из них 5 над А); в сериях BR, VAL и AC органы сохранились лишь частично. Туловище сплошь покрыто многочисленными вторичными фотофорами, особенно густыми на его вентральной поверхности. Пятна светящейся ткани на брюхе отсутствуют. Хотя кожа на хвостовом стебле сорвана, на верхних дополнительных лучах С сохранились остатки чёрной ткани, подлежащей под светящейся железой (вероятно, была овальной формы, занимала область всех верхних дополнительных лучей). Посторбитальный орган (рис. 1а) округлой формы, относительно крупный для Eustomias (примерно вдвое короче диаметра глаза). Суборбитальный орган маленький. Ветви нижней челюсти густо покрыты вторичными фотофорами, на боках головы они более разреженные. Брюшная бороздка доходит до пятого фотофора IP. На praemaxillare семь зубов, из них первый, третий и пятый фиксированные, второй – наибольший. Зубов dentale восемь, из них первый, второй, четвертый и шестой фиксированы, самый длинный, вероятно, также второй, но его вершина обломана (рис. 1а). На maxillare имеются только очень маленькие бугорковидные выступы в задней части кости. Жаберные дуги голотипа сильно повреждены.
Подбородочный усик (рис. 1б, 1в) тонкий, вдвое длиннее головы, его ствол почти не пигментирован, с одной крупной (4.8% длины всего усика) овальной терминальной луковицей без придатков. Парин с соавт. (1978. С. 172) отметили наличие на стволе усика недалеко от начала терминальной луковицы “короткого выступа, несущего пять очень маленьких филаментов (при препаровке … был утрачен)” (согласно приводимому ими рисунку, расстояние от основания усика до выступа в 1.2 раза меньше полной длины усика).
Измерения, в % SL: длина головы 13.0, длина рыла 4.8, горизонтальный диаметр глаза 3.0, ширина костного межглазничного промежутка 1.7, длина верхней и нижней челюстей 9.6, длина посторбитального органа 1.3, длина подбородочного усика и его концевой луковицы соответственно 23.5 и 1.1, максимальная высота тела 4.3; предорсальное, преанальное, превентральное и вентроанальное расстояния соответственно 80.9, 70.4, 60.9 и 9.6.
Этимология. Вид назван в честь одного из коллекторов рейса ФАО-1 и соавторов сообщения, в котором впервые был упомянут экземпляр, послуживший голотипом нового вида, – отечественного ихтиолога С.В. Михайлина (1943–1981), специалиста по трихиуроидным рыбам, трагически погибшего при спасении людей из горящего поезда.
Сравнительные замечания. Только три из 11 номинальных подродов Eustomias включают виды с шестью или более лучами в P: Spilostomias Regan et Trewavas, 1930, Urostomias Regan et Trewavas, 1930 и Triclonostomias Regan et Trewavas, 1930 (Regan, Trewavas, 1930; Gibbs et al., 1983; Прокофьев, 2018). Новый вид хорошо отличается от представителей подрода Triclonostomias простым строением и слабой пигментацией подбородочного усика, терминальная луковица которого лишена филаментов, а ствол – группы из трёх длинных придатков, отходящих из одного места; семью (против восьми) лучами V и малым числом подвижных зубов в челюстях (четыре против 7–11). Гиббс с соавторами (Gibbs et al., 1983) объединили подроды Spilostomias и Urostomias, что было с оговоркой принято и мною (Прокофьев, 2018. Таблица. С. 4). Вместе с тем типовые виды указанных подродов существенно отличаются друг от друга по особенностям развития поверхностной светящейся ткани, пигментации ствола усика и числу лучей V. Последний признак отличается исключительной стабильностью в пределах подродов Eustomias (Прокофьев, 2018), поэтому наблюдаемое различие по числу лучей V между Urostomias и Spilostomias, на мой взгляд, свидетельствует о достаточно давнем расхождении этих групп. После обнаружения E. mikhailini, по всем перечисленным признакам соответствующего виду E. macrura Regan et Trewavas, 1930 – типовому для подрода Urostomias, мне представляется неоправданным объединение Urostomias и Spilostomias в один подрод. Морфологические различия между обсуждаемыми подродами и видами подрода Urostomias могут быть суммированы в следующей определительной таблице.
1(2) Лучей Р 6–16 ………………………….............. 3
2(1) Лучей Р 4–5 (подрод Rhynchostomias) или 0–3 (остальные подроды), если в Р более трёх лучей, то V 8. Подвижных зубов в челюстях намного больше, чем фиксированных …………….................. ............................ все остальные подроды Eustomias
3(4, 5) Лучей V 8. Ствол подбородочного усика полностью пигментирован, с тремя крупными ветвями, отходящими от одного места проксимальнее терминальной луковицы. Нет пятен светящейся ткани на вентральной поверхности туловища или железы в верхней части основания С. Брюшная бороздка достигает 15–24-го фотофора PV. Подвижных зубов в челюстях намного больше, чем фиксированных ………………...................... .............................................. подрод Triclonostomias
4(3, 5) Лучей V 8. Ствол подбородочного усика полностью пигментирован, с двумя филаментами в основании терминальной луковицы (иногда очень слабо развитыми). Пятна светящейся ткани на вентральной поверхности крупные, выражены лучше, чем вторичные фотофоры; нет светящейся железы на С. Брюшная бороздка достигает 5–8-го фотофора PV. Все или почти все зубы в челюстях фиксированные …………… подрод Spilostomias
(один вид: E. (S.) braueri Zugmayer, 1911 (синоним: E. spilopleura Regan et Trewavas, 1930))
5(4, 3) Лучей V 7. Ствол подбородочного усика не пигментирован, за исключением скоплений меланофоров вокруг фотофоров, без развитых придатков (могут иметься крошечные выросты проксимальнее терминальной луковицы). Пятна светящейся ткани на вентральной поверхности отсутствуют или едва различимы, если имеются, намного слабее вторичных фотофоров. Есть светящаяся железа у верхнего края С. Брюшная бороздка не заходит за 8-й фотофор PV. Фиксированных зубов в челюстях больше или столько же, сколько и подвижных (подрод Urostomias) ………. 6
6(7) Лучей Р 7. VAV 13. Брюшная бороздка не достигает вертикали основания Р. Подбородочный усик примерно вдвое длиннее головы, 23.5% SL. В челюстях по семь–восемь зубов, из них три–четыре фиксированы ….……............ E. (U.) mikhailini
7(6) Лучей Р 9. VAV 16–17. Брюшная бороздка заходит за вертикаль основания Р (до 5–8-го органа PV). Подбородочный усик примерно равен голове, 12.5% SL. В челюстях по шесть–десять зубов, из них пять–семь фиксированы ..……............. ....................................................... E. (U.) macrura11.
(рис. 2)
Материал. MNHN 1991–6420, 1 экз. SL 74 мм, 0°25′1.2″ с.ш., 139°04′58.8″ в.д., глубина 650 м, э/с “Coriolis”, рейс “Caride 1”, ст. 44b, 25.09.1968 г.
Описание. D 27, A 38, P 3, V 7. Фотофоров в сериях: BR 11, IP 7, PV 34, OV 32, VAV 16 (шесть над А), VAL 18 (восемь над А), AC 20 (19-й над последним лучом А). Посторбитальный орган маленький, овальный, вытянутый в длину и несколько более приострённый к дистальному концу, около трети диаметра зрачка. Имеется маленький круглый суборбитальный орган, чуть более трети длины посторбитального. Брюшная бороздка слабо развита, прослеживается как узкий желобок до 4-й пары фотофоров PV (рис. 2а). На лучах и межлучевых перепонках передней половины D скопления белой светящейся ткани. Зубов praemaxillare 11 (1, 2, 5, 8, 9-й подвижные) слева, 10 (1, 2, 4, 7, 8) справа; dentale 14 (3, 7, 10, 11) справа, 16 (3, 7, 8, 11, 12) слева; в задней части maxillare гребёнка из примерно восьми мелких зубов, не доходящих до дистального конца. На copula три пары острых, почти прямых зубов. Жаберные лепестки в верхних двух третях длины ceratobranchiale-1 сильно редуцированы.
Подбородочный усик и его внутренний ствол, луковицы и придатки не пигментированы, за исключением мелкого и редкого меланофорного крапа по стволу в промежутке между проксимальной и дистальной луковицами, сгущающегося к интенсивно пигментированному проксимальному полюсу дистальной луковицы, и по стволу основного придатка дистальной луковицы в его проксимальной половине. Две овальные луковицы, вытянутые в длину, разделены промежутком, в 1.5 раза меньшим, чем длина дистальной луковицы; проксимальная луковица втрое короче дистальной; длина дистальной луковицы вдвое больше её высоты. Основной придаток дистальной луковицы с широкой кожистой оторочкой, дистально разветвляется на два нитевидных придатка и несёт такой же придаток на вентральном крае непосредственно у своего основания. Длина придатка до развилки примерно в полтора раза больше длины дистальной луковицы. Ещё три длинных нитевидных придатка отходят обособленно от дистального конца луковицы (два выше и один ниже основного придатка). Все придатки лишены светящихся телец. Фотофоров и пигментных пятен на стволе усика нет (рис. 2б).
Измерения, в % SL: длина головы ca.13.5, горизонтальный диаметр глаза 3.0; предорсальное, преанальное и превентральное расстояния соответственно 81.1, 70.3 и 56.8; длина усика до проксимальной луковицы и всего усика соответственно 47.3 и 56.8; длина промежутка между луковицами 1.6, длина дистальной луковицы и её основного придатка с ветвями – соответственно 2.4 и 4.1. Сумма длин проксимальной и дистальной луковиц – 2.3 мм.
Сравнительные замечания. Изученный экземпляр относится к группе “melanostigma”, характеризующейся двумя разобщёнными терминальными луковицами, из которых дистальная крупнее и несёт несколько нитевидных придатков, лишённых светящихся телец. Группа включает три трудноразличимых вида: помимо E. melanostigma, известного из всех трёх океанов, также E. melanonema Regan et Trewavas, 1930 из тропической Восточной Атлантики и E. melanostigmoides Gibbs, Clarke et Gomon, 1983 из Гавайского района (Gibbs et al., 1983). По удлинённой дистальной луковице, высота которой вдвое меньше длины, описываемый экземпляр соответствует E. melanonema, тогда как у E. melanostigma и E. melanostigmoides высота дистальной луковицы содержится в длине не более полутора раз. Абсолютная длина дистальной луковицы у изученной рыбы близка к нижнему пределу значений, известному для E. melanonema, но суммарная абсолютная длина проксимальной и дистальной луковиц (2.3 мм) явственно меньше наблюдаемой у этого вида (2.7–4.3 мм при SL ≥ 75 мм), но хорошо соответствует таковой E. melanostigma и E. melanostigmoides (1.5–2.7 мм). По величине интербульбарного промежутка (1.6% SL) описываемый экземпляр хорошо соответствует E. melanostigmoides (1.0–2.4% SL). Однако Гиббс с соавторами (Gibbs et al., 1983. Fig. 24 ) приводят 1 экз. E. melanostigma SL 72 мм, у которого интербульбарный промежуток составляет лишь около 1.5% SL (вместо 2.1–4.1% у других экземпляров SL > 70 мм). В диагнозе вида они данного значения не указывают. По длине дистальной луковицы (2.4% SL) описываемый экземпляр гораздо ближе к E. melanostigma ((0.5)1.0–2.2% SL), чем к E. melanostigmoides (0.7–1.5% SL), и укладывается в известные пределы изменчивости у E. melanonema (1.7–2.8% SL).
Таким образом, изученный экземпляр сочетает признаки разных видов группы “melanostigma”, в наибольшей степени соответствуя E. melanonema. Однако он не может быть отождествлён с последним, так как для E. melanonema предполагается ограниченный ареал в Восточной тропической Атлантике (Gibbs et al., 1983). Является ли описанная здесь рыба ещё одним, ранее не известным, видом группы “melanostigma” или её признаки представляют собой крайний вариант изменчивости E. melanostigma (что, в свою очередь, поднимает вопрос о границах между этим видом и E. melanonema и о реальности последнего), будет возможно сказать лишь после новых нахождений рыб с похожим набором признаков.
Одним из диагностических признаков подрода Nominostomias является отсутствие брюшной бороздки, однако у изученного экземпляра на истмусе прослеживается небольшой желобок (рис. 2а), который я трактую как следы брюшной бороздки. Возможно, что наличие брюшной бороздки является исходной синапоморфией рода Eustomias, но она вторично утрачивается у некоторых продвинутых подродов.
(рис. 3)
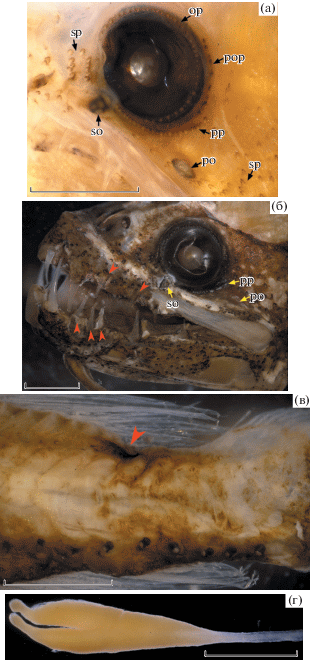
Рис. 3.
Leptostomias cf. bilobatus, Юго-Восточная Пацифика (а, в, г – экз. SL 67 мм; б – экз. SL 116.5 мм): а – светящиеся органы вокруг глаза; б – озубление челюстей, зубы наружного ряда (▶); в – пигментное пятно под концом основания D (▶), г – дистальный конец подбородочного усика. op – ряд фотофоров по краю глазного яблока, po – посторбитальный светящийся орган, pop – ряд фотофоров вдоль заднего края орбиты, pp – ряд фотофоров по краю посторбитальной кожной складки, so – суборбитальный светящийся орган, sp – вторичные фотофоры перед глазом и в нижней части щеки. Масштаб: а – 1.5, б и г – 2, в – 1 мм.
Материал. Всего 2 экз. (ИО РАН № 03612): SL 116.5 мм, 18°00′ ю.ш., 82°30′ з.д., э/с “Профессор Месяцев”, рейс 15, трал 34, 23.09.1984 г.; SL 67 мм, 20°00′ ю.ш., 79°00′ з.д., э/с “Профессор Месяцев”, рейс 15, трал 39, 28.09.1984 г.
Описание (первыми приведены показатели экз. SL 116.5 мм, за ними в скобках – отличающиеся значения экз. SL 67 мм). D 19, A 22, P 10 (11), V 7. Фотофоров в сериях: BR 18 (17), IP 10, PV 47 (44), OV 46 (44), VAV 20 (23) (четыре над А), VAL 21 (22) (пять над А), AC 12. Фотофоры серии IP расставлены на равные промежутки; расстояние от вершины истмуса до первой пары органов IP в 1.5 (2.0) раза превышает промежуток между IP-1 и IP-2. Фотофоры в латеральных (OV и VAL) и вентральных (PV и VAV) рядах расположены на одной прямой. Последний фотофор VAV находится на вертикали середины промежутка между последним и предпоследним органами VAL. Голова и тело несут многочисленные вторичные фотофоры. На голове они особенно густо покрывают рыло над верхней челюстью, ветви нижней челюсти, щёку непосредственно у конца верхней челюсти (но слабо развиты за глазом) и нижнюю часть жаберной крышки; разреженные в верхней части жаберной крышки и в задней части верха головы. На туловище вторичные фотофоры очень густо покрывают брюхо между вентральными рядами серийных фотофоров и непосредственно над ними, более разреженные на боках и спине. Мелкие фотофоры окаймляют глазное яблоко и образуют короткий ряд вдоль заднего края орбиты. Есть ряд хорошо развитых фотофоров по краю кожной складки у нижнезаднего края глаза (посторбитальная складка) (рис. 3а). Суборбитальный орган хорошо развит, его дорсальная часть образует выступ у вентрального края глазного яблока. Посторбитальный орган очень мал, соизмерим с суборбитальным, его передний конец отстоит от свободного края посторбитальной кожной складки (рис. 3а, 3б). Посторбитальный орган овальный у экз. SL 67 мм, суженный каудально у экз. SL 116.5 мм. Лучи D и А покрыты светящейся тканью. Истмус с глубокой медиальной бороздкой, оканчивающейся у IP-7. Под основанием последнего луча D имеется чёрное оконтуренное пигментное пятно (рис. 3в).
На praemaxillare 8/6 (9/7) разноразмерных зубов, из них второй зуб значительно длиннее остальных. У экз. SL 116.5 мм третий/третий–четвертый и пятый/шестой премаксиллярные зубы расположены кнаружи от остальных зубов (бисериальный тип озубления: Прокофьев, 2019). На dentale 8 (6/5) разноразмерных зубов, из которых второй – наибольший. У экз. SL 116.5 мм третий, четвертый и пятый зубы расположены кнаружи от остальных (рис. 3б). У экз. SL 67 мм все зубы на praemaxillare и dentale расположены в один ряд (унисериальное озубление). Нижнечелюстные зубы собраны в группы (первый и второй, третий–пятый (третий и четвертый), шестой и седьмой (пятый) и восьмой (шестой)), широко разобщённые друг от друга (рис. 3б). На maxillare у экз. SL 116.5 мм непосредственно перед рядом мелких прижатых назад зубов (занимающих около двух третей длины кости) с каждой стороны имеется один более крупный зуб, у экз. SL 67 мм присутствует только гребёнка из мелких зубов. На сошнике 1 + 1 (0 + 0) зубов (когда имеются, широко расставлены); нёбных зубов нет. На языке две отстоящие друг от друга пары слабых зубов. Зубчики на жаберных дугах длинные и острые, у меньшего экземпляра отстоящие друг от друга на равные промежутки, у большего – собранные в группы попарно. Жаберные лепестки ceratobranchiale-1 длинные.
Подбородочный усик длинный, достигает середины длины основания А у экз. SL 116.5 мм (немного не достигает начала А у экз. SL 67 мм). Пигментация усика прослеживается только в проксимальных 6–10% его длины. В основании и по ходу ствола усика нет филаментов. Концевая часть усика (рис. 3г) удлинённая, проксимально суженная, на вершине асимметрично раздвоённая, без филаментов.
Измерения, в % SL: длина головы 12.9 (13.4), длина рыла 3.4 (3.4), горизонтальный диаметр глаза 2.15 (2.2), ширина межглазничного промежутка 3.4 (3.7), длина верхней челюсти 7.7 (9.0); предорсальное, преанальное и превентральное расстояния соответственно 87.1 (86.6), 86.3 (84.3) и 65.7 (64.2); длина усика и его концевой части соответственно 90.1 (71.6) и ? (повреждена в проксимальной части) (6.0), длина посторбитального органа 0.4 (0.5).
Сравнительные замечания. Изученные экземпляры в целом согласуются с имеющимися описаниями этого вида (Koefoed, 1956; Morrow, Gibbs, 1964), за исключением несколько большего числа лучей в Р (10–11 против 9), но меньшего в D и A (соответственно 19 и 22 против 20–21 и 25–26), большего числа фотофоров PV и OV (соответственно 44–47 и 44–46 против 42–45 и 42) и отсутствия филаментов на концевой части усика и в его основании. Большинство из перечисленных отличий можно было бы связать с изменчивостью вида, но наличие или отсутствие филаментов в основании усика считается видоспецифичным признаком у Leptostomias (Morrow, Gibbs, 1964). Необходимо исследование дополнительных материалов для решения вопроса о таксономической значимости выявленных различий.
Морроу и Гиббс (Morrow, Gibbs, 1964. P. 439, 441) указывают в качестве отличительного признака L. bilobatus равнорасставленные зубчики жаберных дуг, однако из изученных мною рыб только меньший экземпляр характеризуется данным признаком, тогда как у большей рыбы они собраны в группы по двое. Поскольку последний экземпляр заметно крупнее рыб, описанных Куфудом и Морроу с Гиббсом (Koefoed, 1956; Morrow, Gibbs, 1964) (SL 116.5 мм против 67.8–82.0 мм), возможно, что равнорасставленные зубчики жаберных дуг являются ювенильной чертой и что взрослые экземпляры L. bilobatus по данному признаку неотличимы от других представителей рода.
Вид был описан по голотипу SL 82 мм из Северной Атлантики (29°08′ с.ш., 25°16′ з.д.), позднее утраченному (Koefoed, 1956); затем Морроу и Гиббс (Morrow, Gibbs, 1964) привели описание двух экземпляров SL 67.8 и 72.9 мм из Мексиканского залива (26°52′ с.ш., 89°44′ з.д.), меньший из которых был обозначен в качестве неотипа. Впоследствии L. bilobatus был указан из “широко распределённых местонахождений в Атлантическом океане и в юго-западной части Индийского океана” (Gibbs, Barnett, 1990. P. 325; Moore et al., 2003. P. 189) без приведения морфологических описаний или иллюстраций. В Каталоге рыб (Fricke et al., 2018) ареал вида охарактеризован как космополитичный, но без указания соответствующих литературных ссылок. Описанные экземпляры являются первым достоверным свидетельством присутствия данного вида в юго-восточной части Тихого океана. Учитывая некоторые различия между ними и ранее описанными рыбами из Северной Атлантики, необходимо детальное сравнение материалов по этому виду из разных районов Мирового ареала.
(рис. 4)
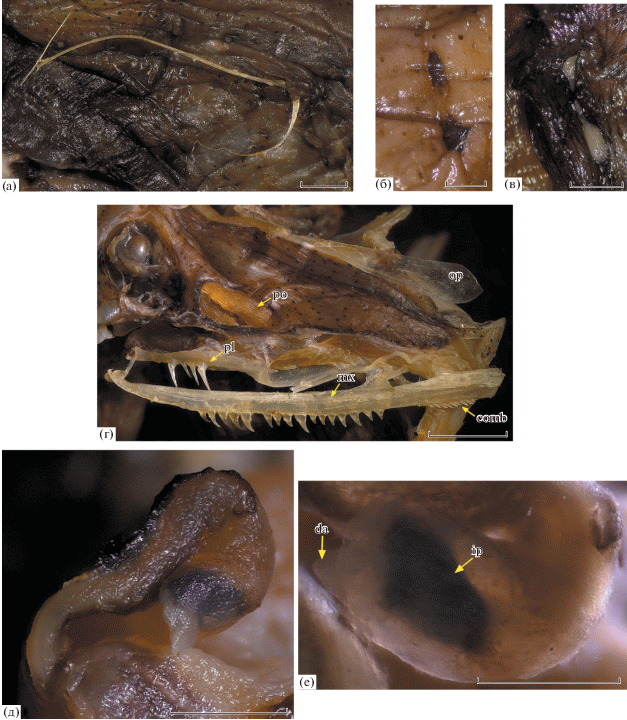
Рис. 4.
Photonectes xenopogon SL 120 мм (а, б, г–е) и Photonectes sp. SL 116 мм (в), Юго-Восточная Пацифика: а – лучи Р; б, в – пятна белой светящейся ткани позади верхнезаднего края жаберного отверстия; г – озубление maxillare, palatinum и заглазничная часть головы; д – подбородочный усик, е – луковица усика. comb – гребёнка мелких зубов в задней части maxillare, da – дистальный придаток луковицы усика, ip – внутренняя пигментация луковицы усика, mx – maxillare, op – operculum, pl – palatinum, po – посторбитальный светящийся орган. Масштаб: 1 мм.
Материал. ИО РАН № 03613 SL 120 мм в одной ёмкости с экз. SL 116 мм (см. дополнительный материал) и с двумя бирками, ранее прикреплёнными к рыбам, но на момент обнаружения пробы от них отлетевшими, которые содержат следующие этикеточные данные: 1) э/с “Ихтиандр”, рейс 5, трал 47, 15.10.1979 г.; 2) э/с “Ихтиандр”, рейс 6, трал 14, 16.08.1980 г. Район подводных хребтов Наска и Сала-и-Гомес. Экземпляр, по-видимому, является незрелым самцом (гонады слабо развиты). Голова сильно повреждена (раздавлена, часть костей утрачена).
Дополнительный материал. ИО РАН № 03614 SL 116 мм, подбородочный усик утрачен.
Описание. SL 120 мм. D 16, A 20, P 2 (передний луч крепкий и длинный, задний – короткий и слабый (рис. 4а)), V 7, vert. 60. Лучи D и A полностью покрыты толстой кожей. Длинный луч P состоит из двух плотно соединённых половин (разделённых до места его облома). Длина короткого луча Р около 4 мм (3.3% SL). Фотофоров в сериях: BR 10, IP 10, PV 31, OV 30, VAV ca.11 (восемь до ануса, девять до вертикали начала А), VAL ca.11, AC 10. С каждой стороны позади верхней части жаберного отверстия (рис. 4б) два горизонтально вытянутых чёрных “рубцевидных” пятна, представляющих собой подложку пятен белой светящейся ткани (рис. 4в) (сама ткань сорвана). Нет пятен светящейся ткани в дне рта и, по-видимому, в межглазничном промежутке; на вершине рыла кожа содрана. Голубая светящаяся ткань отсутствует. Посторбитальный орган крупный (соизмерим с диаметром орбиты), его передний край высокий, прямо срезанный, к каудальному концу суженный (рис. 4г).
Длина рыла примерно равна диаметру глаза. Praemaxillaria утрачены. На maxillare 16 слабо различающихся по величине зубов, за которыми через промежуток следует 12 гораздо более мелких зубов, направленных вершинами назад и образующих подобие гребёнки (рис. 4г). Величина промежутка между нормальными зубами и зубами гребёнки составляет примерно половину длины последней. Озубление dentale гомогенное унисериальное, лишь третий зуб немного длиннее соседних (сохранение ювенильной черты?), число зубов – 25. Зубы на palatinum не образуют группы, числом 4/4. Вершины всех зубов dentale и palatinum простые, но у отдельных зубов на maxillare с маленькими дополнительными зубчиками. Жаберные лепестки одинаково длинные на всем протяжении ceratobranchiale-1. Подбородочный усик (рис. 4д) короткий (1.2 раза в длине рыла), кожа на нем, по-видимому, сорвана, субдермальная пигментация слабая. Луковица небольшая, округлая, слабо утолщённая, с внутренней пигментацией, несёт короткий непигментированный лопастевидный дистальный придаток (рис. 4д, 4е).
Измерения, в % SL: длина рыла 2.5, ширина межглазничного промежутка 3.75, длина усика 2.9, длина костей maxillare и dentale соответственно 12.5 и 13.75, максимальная и минимальная высота тела соответственно 15.0 и 2.9, предорсальное, преанальное и превентральное расстояния соответственно 82.1, 79.6 и 66.7.
Экземпляр SL 116 мм. D 17, A 22, P 2, V 7, vert. 58. Фотофоры в сериях сохранились лишь частично. Позади верхней части жаберного отверстия с каждой стороны два чётко отграниченных пятна белой светящейся ткани (рис. 4в). Посторбитальный орган соизмерим с длиной рыла и диаметром глаза, к заднему концу сужен. Озубление praemaxillare и dentale гомогенное унисериальное, зубы на praemaxillare крупнее зубов на maxillare и dentale. На praemaxillare шесть зубов; на maxillare 12/14 зубов, за которыми следует гребёнка из более мелких, наклонённых назад зубов; на dentale более 20 зубов (часть утрачена); зубов на vomer 2 + 2. Зубы на palatinum разноразмерные, не образуют групп, числом 5/5. Жаберные лепестки ceratobranchiale-1 не укорочены.
Измерения, в % SL: длина головы 12.1, длина рыла, горизонтальный диаметр глаза и длина посторбитального органа 2.2, ширина межглазничного промежутка 3.45, длина верхней челюсти 12.9, максимальная и минимальная высота тела соответственно 12.9 и 2.0, предорсальное, преанальное и превентральное расстояния соответственно 81.0, 81.0 и 69.0.
Сравнительные замечания. Сохранившиеся у экз. SL 116 мм признаки хорошо согласуются с таковыми у экз. SL 120 мм, вследствие чего они могут быть конспецифичными, но, так как усик у экз. SL 116 мм утрачен и рыбы пойманы в разных экспедициях, в этом нельзя быть уверенным. Экз. SL 116 мм может принадлежать и к виду P. margarita (Goode et Bean, 1896) с широкотропическим циркумглобальным ареалом, включающим и воды восточной части Тихого океана (Прокофьев, 2015). Различия между P. margarita и P. xenopogon заключаются в строении подбородочного усика, не имеющего внутренней пигментации у первого вида, и его дистального придатка(ов) (одиночный, лопастевидный и непигментированный у P. xenopogon против множественных, по меньшей мере, частью нитевидных и пигментированных у P. margarita) (Прокофьев, 2019). Экз. SL 120 мм по строению усика хорошо соответствует голотипу P. xenopogon, за исключением того, что дистальный придаток луковицы у него более короткий и неподразделённый: ср. рис. 4д (настоящая работа) и рис. 6 г (Прокофьев, 2019). Это может быть связано как с индивидуальной изменчивостью, так и с повреждением усика у экз. SL 120 мм. Кроме того, у описываемого экземпляра два луча Р (задний – короткий и слабый), тогда как у типовых экземпляров P. xenopogon в Р имеется только один длинный и крепкий луч (состоящий, как и передний луч у экз. SL 120 мм, из двух плотно соединённых половинок). Однако, поскольку точно такая же изменчивость в строении Р отмечена для P. margarita (Прокофьев, 2019), это различие не может служить препятствием для отождествления описываемой рыбы с P. xenopogon.
Ранее P. xenopogon был известен только по двум экземплярам типовой серии, происходящим из моря Сулу. Нахождение в районе подводного хребта Наска свидетельствует о гораздо более широком распространении этого вида, по меньшей мере, в водах Тихого океана.
Список литературы
Парин Н.В., Похильская Г.Н. 1974. Обзор индо-тихоокеанских видов рода Eustomias Vaillant (Melanostomiatidae, Osteichthyes) // Тр. ИО АН СССР. Т. 96. С. 316–368.
Парин Н.В., Сазонов Ю.И., Михайлин С.В. 1978. Глубоководные пелагические рыбы в сборах НПС “Фиолент” в Гвинейском заливе и прилегающих районах // Там же. Т. 111. С. 169–184.
Прокофьев А.М. 2015. Материалы к ревизии подрода Trachinostomias (Melanostomiidae: Photonectes). Морфологическая изменчивость P. margarita с описанием двух новых видов // Вопр. ихтиологии. Т. 55. № 2. С. 131–138.
Прокофьев А.М. 2018. Три новых вида Eustomias (Melanostomiidae) из южносубтропических вод с описанием нового подрода // Там же. Т. 58. № 1. С. 2–9.
Прокофьев А.М. 2019. Материалы к ревизии рода Photonectes (Melanostomiidae) // Там же. Т. 59. № 4. С. 375–404.
Fink W.L. 1985. Phylogenetic interrelationships of the stomiid fishes (Teleostei: Stomiiformes). № 171. Michigan: Miscell. Publ. Mus. Zool. Univ., 127 p. + i–vii.
Fricke R., Eschmeyer W.N., van der Laan R. 2018. Catalog of fishes: genera, species, references.(http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Version 11/ 2018)
Gibbs R.H. 1968. Photonectes munificus, a new species of melanostomiatid fish from the south Pacific subtropical convergence, with remarks on the convergence fauna // Contrib. Sci. Los Angeles Co. Mus. № 149. P. 1–6.
Gibbs R.H., Barnett M.A. 1990. Melanostomiidae // Check-list of the fishes of the eastern tropical Atlantic (CLOFETA). V. 1 / Eds. Quero J.C. et al. Lisbon; Paris: JNICTSEI, UNESCO. P. 308–337.
Gibbs R.H., Clarke T.A., Gomon J.R. 1983. Taxonomy and distribution of the stomioid fish genus Eustomias (Melanostomiidae), I: subgenus Nominostomias // Smithson. Contrib. Zool. № 380. 139 p. + i–iv.
Harold A.S., Weitzman S.H. 1996. Interrelationships of stomiiform fishes // Interrelationships of Fishes / Eds. Stiassny M.L.J. et al. San Diego: Academic Press P. 333–353.
Koefoed E. 1956. Isospondyli. 1. Gymnophotodermi and Lepidophotodermi from the “Michael Sars” North Atlantic deep-sea expedition 1910 // Sci. Res. M. Sars N. Atlantic Deep-Sea Exped. 1910. V. 4. Pt. 2. № 5. P. 1–21.
Moore J.A., Hartel K.E., Craddock J.E., Galbraith J.K. 2003. An annotated list of deepwater fishes from off New England region, with new area records // Northeastern Naturalist. V. 10. № 2. P. 159–248.
Morrow J.E., Gibbs R.H. 1964. Melanostomiatidae // Fishes of the Western North Atlantic. V. 1. Pt. 4. New Haven: Sears Found. P. 351–522.
Nelson J.S. 2006. Fishes of the World. Hoboken, N.J.: John Wiley and Sons, Inc. 601 p.
Nelson J.S., Grande T.C., Wilson M.V.H. 2015. Fishes of the World. Hoboken, N.J.: John Wiley and Sons, Inc. 707 p.
Parr A.E. 1927. The Stomiatoid fishes of the suborder Gymnophotodermi (Astronesthidae, Melanostomiatidae, Idiacanthidae) with a complete review of the species // Bull. Bingham Oceanogr. Collect. Yale Univ. V. 3. Art. 2. 123 p.
Regan C.T., Trewavas E. 1930. The fishes of the families Stomiatidae and Malacosteidae // Dana Rept. V. 6. P. 1–143.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии