

Вопросы ихтиологии, 2020, T. 60, № 4, стр. 383-391
Новые данные о короткожаберном гимнелопсе Gymnelopsis brevifenestrata (Zoarcidae) из Охотского моря
Н. В. Чернова 1, *, М. В. Назаркин 1
1 Зоологический институт РАН – ЗИН РАН
Санкт-Петербург, Россия
* E-mail: nchernova@mail.ru
Поступила в редакцию 15.07.2019
После доработки 16.09.2019
Принята к публикации 18.09.2019
Аннотация
Короткожаберный гимнелопс Gymnelopsis brevifenestrata (Zoarcidae, Gymnelinae) − эндемик Охотского моря, до недавнего времени был известен по 12 типовым особям из музейных коллекций. На основании изучения нескольких новых экземпляров и паратипов охарактеризована изменчивость вида и уточнено его распространение. Впервые описана прижизненная окраска самцов, дающая новые признаки для полевого определения вида. G. brevifenestrata обитает в западных и северных районах Охотского моря на глубинах 76−200 м. Учитывая приуроченность G. brevifenestrata к переохлаждённым водам арктического характера, его следует относить к группе гляциально-охотоморских видов.
Гимнелопсы рода Gymnelopsis Soldatov, 1922 – бельдюговые рыбы (Zoarcidae) небольших размеров, обитающие в Охотском и Японском морях (Солдатов, Линдберг, 1930; Линдберг, Красюкова, 1975; Anderson, 1982, 1994; Назаркин, Чернова, 2003). Населяют шельф и верхнюю часть континентального склона на глубинах от 70 до 783 м, ведут донный образ жизни (Anderson, 1982, 1994). Как и другие представители подсемейства Gymnelinae, гимнелопсы характеризуются удлинённым низким телом, отсутствием брюшного плавника, большим числом позвонков (86−115). От представителей наиболее близкого рода Gymnelus Reinhardt, 1834 они отличаются наличием чешуи, меньшим числом лучей хвостового плавника (5−8 против 9−12), наличием корональной поры. Последняя у видов Gymnelus отсутствует (кроме G. popovi, который, возможно, относится к роду Commandorella Taranetz et Andriashev, 1935, в котором был первоначально описан).
Состав рода Gymnelopsis вызывает дискуссии. В последней ревизии (Anderson, 1982) в род включены четыре вида: G. ocellata Soldatov, 1922, G. brashnikovi Soldatov, 1922, G. brevifenestrata Anderson, 1982 и G. ochotensis (Popov, 1931); пятый вид, япономорский гимнелопс G. japonica Katayama, 1943, сведён в синонимию вида G. ochotensis, хотя японскими ихтиологами (Katayama, 1943; Toyoshima, 1981) были показаны значительные различия этих видов, географически разделённых большим расстоянием. Считая их аргументы убедительными, ряд авторов продолжают рассматривать G. japonica в качестве самостоятельного вида (Назаркин, Чернова, 2003; Shinohara et al., 2011, 2014). Нет единого мнения и о родовой принадлежности G. ochotensis. Одни исследователи (Toyoshima in Masuda et al., 1984; Шейко, Федоров, 2000; Федоров и др., 2003; Balushkin et al., 2011) продолжают включать его в род Derjuginia Popov, 1931, в котором он был описан. Другие, следуя Андерсону (Anderson 1982, 1994; Anderson, Fedorov, 2004), считают его гимнелопсом (Shinohara et al., 2011; Парин и др., 2014). В данной работе мы принимаем в составе Gymnelopsis шесть видов, учитывая, кроме выше перечисленных, G. humilis Nazarkin et Chernova, 2003 (Назаркин, Чернова, 2003).
Короткожаберный гимнелопс G. brevifenestrata был описан из Охотского моря по 12 экз., собранным экспедициями 1912−1949 гг. и хранившимся в ЗИН (Anderson, 1982). Вид относили к категории редких (Федоров и др., 2003), так как его не находили до последнего времени, и лишь недавно он был указан в материалах траловой съёмки (Савельев и др., 2019).
Поводом для написания данной работы послужили 2 экз. G. brevifenestrata из сборов научно-исследовательского судна (НИС) “Зодиак” в Охотском море (2001 г.), обнаруженные при разборе хранящихся в ЗИН коллекций. Прижизненная окраска рыб этого вида не была известна (Anderson, 1982), поэтому интерес представляют фотографии экземпляров, выполненные при их поимке. Для подтверждения правильности определения G. brevifenestrata, на тот момент известного лишь по первоописанию, были исследованы его паратипы. Розыски дополнительных материалов выявили в коллекциях ЗИН ещё несколько особей вида среди Gymnelopsis sp. и Derjuginia sp. В результате оказалось возможным исследовать в общей сложности 23 экз. G. brevifenestrata (бóльшая часть – молодь). Мы не ставили своей задачей выполнить полное переописание вида: в этом нет необходимости, поскольку его описание было выполнено на высоком уровне (Anderson, 1982).
Цель настоящей работы − дополнить имеющиеся сведения новыми данными и обобщить известные материалы. В задачи работы входило охарактеризовать морфологическую изменчивость G. brevifenestrata, а также провести критический анализ опубликованных данных, исправить номенклатурные и иные неточности, уточнить распространение вида, условия его обитания и зоогеографическую характеристику.
МАТЕРИАЛ И МЕТОДИКА
Использованы методы, применявшиеся при обработке рыб подсемейства Gymnelinae, включая методику изучения пор сейсмосенсорной системы (рис. 1) (Чернова, 1998; Назаркин, Чернова, 2003). Число костных элементов подсчитано по рентгенограммам, на которых различимы все позвонки до конца хвостовой части тела (n = 12). Андерсон (Anderson, 1982) указывает 1-й луч спинного плавника (D) как колючий, но на рентгенограммах передний луч D не отличается от последующих, поэтому мы его не выделяем. Протяжённость очешуения измеряли на боках тела от наиболее передних до наиболее задних чешуек (последние обычно расположены вблизи основания хвостового плавника). Число зубов на челюсти подсчитывали во внешнем (наиболее длинном) ряду, во внутреннем (более коротком) и в среднем ряду (если таковой имеется). Измерения (выполнены по экземплярам хорошей сохранности) приведены в процентах стандартной длины (SL) и длины головы (c). Формула для пересчёта пропорций относительно абсолютной длины тела (TL): SL = 0.98TL (n = 15).
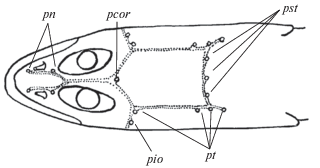
Рис. 1.
Сейсмосенсорные поры (⚪) и каналы (· · ·) Gymnelopsis: pn – носовые (=anterior supraorbital – по: Anderson, 1982), pcor – корональная (= interorbital), pst – супратемпоральные (=occipital). pio – инфраорбитальные (=suborbital) (показана последняя пора в инфраорбитальном канале), pt – темпоральные (=postorbital, 1 + 3). Поры преоперкуломандибулярного канала, расположенные на нижней стороне головы, не изображены.
Прижизненная окраска приводится по полевым записям и фото двух самцов (ЗИН № 56454, 56455).
В работе приняты следующие обозначения морфометрических признаков: c – длина головы (до кожного края operculum); hc, wc – высота и ширина головы, ао – длина рыла, о – горизонтальный диаметр глаза, io − межглазничное расстояние (костное); aD, aA – антедорсальное и антеанальное расстояния; H1, H2 – высота тела над грудным плавником и над началом анального плавника; lP, lС – длина грудного и хвостового плавников; lmx − длина верхней челюсти; D, A, P, С – число лучей в спинном, анальном, грудном и хвостовом плавниках; Dabd – число лучей D в предхвостовом (=туловищном) отделе; vert., vert. abd., vert.c. – число позвонков общее, туловищных и хвостовых.
В квадратные скобки помещены комментарии, уточняющие данные из опубликованных источников.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Gymnelopsis brevifenestrata Anderson, 1982 – короткожаберный гимнелопс (рис. 2, 3)
Рис. 2.
Рентгенограмма Gymnelopsis brevifenestrata − самка TL 104.5 мм, паратип ЗИН № 33751; in1 – первая interneurale, D1 – первый (укороченный) луч спинного плавника.

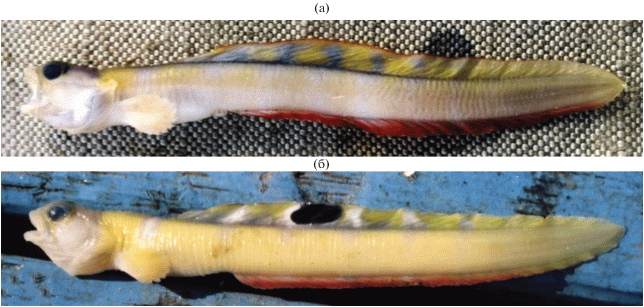
Рис. 3.
Прижизненная окраска самцов короткожаберного гимнелопса Gymnelopsis brevifenestrata: а – TL 111.5 мм (ЗИН № 56454), б − TL 99 мм (ЗИН № 56455); указана длина при поимке.
Gymnelopsis brevifenestratus Anderson, 1982. Р. 52. Fig. 33 (Охотское море, 100−200 м, SL ≤ 122 мм; грамматический род Gymnelopsis ошибочно принят как мужской).
Gymnelopsis brevifenestrata: Anderson, 1994. Р. 32, 112 (частью: Охотское море, но не Япония; SL 89 мм относится к голотипу). Федоров и др., 2003. С. 110 (высокобореальный приазиатский вид, эндемик северо-западной части Охотского моря; глубина 76−200 м; рисунок из: Anderson 1982. Fig. 33 ). Anderson, Fedorov, 2004. Р. 4 (частью: Охотское море, но не Япония).
Derjuginia sp. (non Popov, 1931): Balushkin et al., 2011. Р. 960 (в каталоге коллекций ЗИН, частью: № 44705). Балушкин и др., 2012. С. 28 (то же).
Gymnelopsis brashnikovi (non Soldatov, 1922): Balushkin et al., 2011. Р. 960 (частью: ЗИН № 44729). Балушкин и др., 2012. С. 29 (то же).
Gymnelopsis brevifenestratus: Balushkin et al., 2011. Р. 960−961 (частью: кроме ЗИН № 46785). Балушкин и др., 2012. С. 29 (то же).
Gymnelopsis sp.: Balushkin et al., 2011. Р. 961 (частью: ЗИН № 32119 [2 из 3 экз.], № 33273). Балушкин и др., 2012. С. 30 (то же).
Gymnelopsis brevifenestrata: Парин и др., 2014. С. 389 (северо-западная часть Охотского моря, редкий; глубина 76−200 м). Савельев и др., 2019. С. 405 (Притауйский район, в составе уловов траловой съёмки, глубина 97−150 м).
Типовая серия: 12 экз. (голотип и 11 паратипов) с пяти станций. [В описании G. brevifenestrata в составе типовой серии перечислены 12 экз. (Anderson 1982. Р. 53), но в тексте и таблице промеров приводятся данные 13 экз. Причину разночтений установить не удалось.]
Голотип: (был передан А.М. Поповым в Национальный музей США (United States National Museum − USNM), ныне − Национальный музей естественной истории, Смитсониановский институт, Вашингтон, США (National Museum of Natural History, Smithsonian Institution, Washington, USA) из ЗИН в 1932 г.) USNM № 92587 − молодая самка SL 89 мм, Охотское море, между м. Аян и р. Иней, 57°50′ с.ш., 141°47′ в.д., глубина 80 саженей [146 м], грунт жёлтый песок и камни, 30.06.1912 г., гидрографическая экспедиция Восточного океана (ГЭВО), траулер “Охотск”, ст. 11, коллектор Ф.А. Дербек.
Изученный материал – 23 экз. (11 паратипов и 12 нетиповых экземпляров), все из Охотского моря.
Паратипы. ЗИН № 23944 − TL 73 мм, SL 71 мм, пойман вместе с голотипом; ЗИН № 23952 – TL 99.0 мм, SL 96.5 мм, пойман вместе с голотипом; ЗИН № 23953 − TL 122 мм, SL 119 мм, 54°14′ с.ш., 143°45′ в.д., ГЭВО, ст. I 17, 12.07.1918 г. [3 экз., указанные в этом номере (Balushkin et al., 2011; Балушкин и др., 2012), − опечатка.]; ЗИН № 33334 – 6 экз. TL 74−95 мм, 58°50′ с.ш., 146°48′ в.д., ст. III 18, 20.07.1916 г., ГЭВО; ЗИН № 33751 − самка TL 104.5 мм, SL 102.0 мм, НИС “Витязь”, ст. 60, 56°59′ с.ш., 149°12′ в.д., глубина 175 м, 29.08.1949 г., бим-трал, сборы ИО АН СССР; ЗИН № 34842 − самка TL 120.0 мм, SL 117.5 мм, 58°41.5′ с.ш., 149°47.50′ в.д., 08.08.1915 г., ГЭВО.
Нетиповой материал. ЗИН № 32119 − 2 самца TL 117 и 113 мм, SL 115.0 и 111.5 мм, между островами Коровий и Ионы, 58°41′ с.ш., 147°45′ в.д., глубина 66 саженей [121 м], ст. 20, 22.08.1912 г., коллектор Ф.А. Дербек; ЗИН № 33273 − juv. TL 88.0 мм, SL 86.5 мм, у входа в Тауйскую губу, 58°28.5′ с.ш. [но не 53°28.5′ (Balushkin et al., 2011; Балушкин и др., 2012) − опечатка], 148°41′ в.д., 75 саж. [137 м], ст. IХ № 5, 27.08.1914 г., сборы ГЭВО; ЗИН № 44705 − juv. TL 74 мм, SL 73 мм, глубина 103 м, большой морозильный рыболовный траулер (БМРТ) “Посейдон”, ст. 144, 30.07.1978 г. [в районе м. Энкан], грунт ил, песок, камни, коллектор В.Н. Кобликов; ЗИН № 44727 − 4 экз. TL 57−96 мм, 55°00′ с.ш., 141°09′ в.д., 28.08.1978 г., БМРТ “Посейдон”, рейс 21, ст. 270, глубина 140 м, песчанистый ил с галькой, придонная температура 1.73°С, шлюпочный трал Сигсби, коллектор В.A. Павлючков; ЗИН № 44729 − TL 82 мм, Охотское море, у [северной оконечности] Сахалина, БМРТ “Посейдон”, рейс 22, ст. 269, глубина 102 м, 28.08.1978 г., заиленный песок, шлюпочный трал, коллектор В.A. Павлючков; ЗИН № 48107 − самец TL 116 мм, 48°00′ с.ш., 144°01′ в.д., глубина 80−76 м, БМРТ “Мыс Бабушкина”, трал 130, 10.08.1986 г., коллектор Л.А. Борец; ЗИН № 56454 − самец TL 107 (при поимке 111.5) мм, SL 105 мм, Охотское море, 58°55′ с.ш., 148°00′ в.д., глубина 113 м, НИС “Зодиак”, трал 18, 19.08.2001 г., коллектор М.В. Назаркин. ЗИН № 56455 − самец TL 96.0 (при поимке 99.0) мм, SL 94.5 мм, Охотское море, 58°30′ с.ш. 149°44′ в.д., глубина 122 м, НИС “Зодиак”, трал 20, 19.08.2001 г., коллектор М.В. Назаркин.
Уточнённый диагноз. Вид отличается от прочих Gymnelopsis сочетанием следующих признаков: vert. 89−96, vert. abd. 16−18, D 80−89, А 73−80. Спинной плавник обычно начинается над концом грудного плавника (над 6−9-м позвонками), aD 22−29% SL. Чешуя имеется только на хвостовой части тела (на протяжении, равном 1−3 c; у молоди развита в меньшей степени, чем у взрослых). Жаберное отверстие расположено выше основания грудного плавника и достигает его верхнего луча. Подглазничных пор обычно 6 (реже 7−8).
Приводим признаки, дополняющие или уточняющие первоописание (по совокупности изученного материала). Тело удлинённое, его высота содержится 8.6−13.5 раза в SL. Голова содержится 5.1−6.5 раза в SL; её высота составляет около половины c (46−56%). Ширина головы у молоди примерно равна высоте, с возрастом несколько увеличивается (58−64% c). Лицевая часть головы более уплощённая, чем сжатая с боков. Глаз продольно-овальный и довольно большой, 25−32% c; его диаметр несколько больше рыла; у молодых немного навыкате. Межглазничное пространство в три−пять раз меньше диаметра глаза. Длина трубчатой ноздри меньше диаметра зрачка. Рот конечный; у взрослых самцов верхняя челюсть достигает вертикали заднего края глаза, у прочих особей заканчивается под задним краем зрачка. Длина верхней челюсти у наиболее крупных самцов (TL 116−122 мм) составляет 52−55% c, у самки сходной длины (120 мм) – 44% с. Жаберное отверстие меньше глаза и расположено полностью выше основания грудного плавника: не доходит до его верхнего луча или достигает его. Оперкулярная лопасть округлая, с кожистым краем, под которым на теле имеется хорошо выраженная кожная складка, плотно замыкающая жаберное отверстие (сифон). Жаберных лучей у всех особей 5 (4 + 1).
Зубы на верхней челюсти обычно трёхрядные, разноразмерные. Во внешнем ряду их 8−14, во внутреннем – 5−14, в среднем – 4−8; всего зубов 17−27. Внешние зубы крупные, клыковидные, на протяжении всей длины челюсти сидят с широкими промежутками. Внутренний ряд (развит на передней половине челюсти) короткий и состоит из мелких ровных зубов; зубы среднего ряда меньше прочих. На нижней челюсти зубы однорядные на большей части её длины, лишь спереди добавляются один (реже два) коротких ряда. Во внешнем ряду 12−18 зубов, они крупные, редко сидящие; во внутреннем ряду их 2−5, в среднем – 0−4; всего зубов 16−22. Зубов на сошнике 3−7 [в первоописании 7−10]; нёбных 4−9, в один ряд. С возрастом число зубов увеличивается. Нёбная дыхательная перепонка занимает примерно 2/3 длины челюсти, по ширине она ỳже полосы челюстных зубов. Нижнечелюстная перепонка развита на передней трети челюсти. Нёбо усажено папиллами, особенно многочисленными в области дыхательной перепонки.
Сейсмосенсорных пор в супратемпоральной комиссуре всегда три (pst); корональная пора (pcor) одна; носовых (pn) две, темпоральных (pt) – 1 + 3 (рис. 1). Андерсон (Anderson, 1982) указывает стабильное число пор также в инфраорбитальном (suborbital) и преоперкуломандибулярном каналах (соответственно 6 и 6), однако по нашим данным число этих пор изменчиво: pio 6−8 и ppm 6−8. При обычном числе пор pio 6 их может быть 6 справа и 7 слева (ЗИН № 44727), 7 (у паратипов ЗИН № 23944 и 23952 и у экземпляра ЗИН № 44727), 8 и 7 (ЗИН № 56454). При обычном числе пор ppm 6 их может быть 7 (ЗИН № 56455) или 8 (ЗИН № 56454).
Боковая линия медиолатеральная, состоит из свободно сидящих невромастов (заметна на передней части тела).
Спинной плавник начинается обычно над концом грудного плавника (реже немного смещён вперёд или назад) (рис. 2). У взрослых самцов D высокий (при расправленных лучах равен примерно половине высоты тела); анальный плавник ниже спинного. Грудной плавник удлинённый, закруглён на конце, его основание составляет 32−48 (в среднем 40.0)% длины Р [по первоописанию 28−44, в среднем 36%]; мембрана плавника у кончиков лучей отчётливо вырезана. Лучей Р 10−11 (редко 9).
Анус открывается перед началом А. Чешуя мелкая, редко сидящая, имеется обычно только на задней половине хвостовой части тела. На расстоянии, равном диаметру глаза, насчитывается пять−шесть чешуй. У молоди (TL 96−107 мм) чешуя занимает пространство, примерно равное длине головы (17−19% TL). У взрослых особей (TL 120−121.5 мм) она развита лучше: по протяжённости занимает пространство ~3 c (54−58% TL), а на боках немного заходит вперёд за середину длины тела.
Рентгенограммы (n = 12): vert. 89−96 (91.8), vert. abd. 16−18 (16.3), vert.c. 73−80 (75.4), D 80−89 (85.5), А 73−81 (76.0) (таблица). Наличие колючих лучей D (указанных в первоописании: D I 79−86), равно как и сегментация лучей, по рентгенограммам не выявлены. Свободных от лучей D птеригиофоров (interneuralia) 0−5 (чаще два), расположены между невральными отростками 3−8-го позвонков. Птеригиофор 1-го луча D находится между невральными отростками 5−9-го позвонков. Dabd 7−15 (10.8), передние один−четыре луча короче последующих. В анальном плавнике свободные птеригиофоры отсутствуют; переднему птеригиофору соответствуют один−два луча А. Начало А ассоциировано с 16−18-м позвонками.
Распределение исследованных экземпляров Gymnelopsis brevifenestratа по значениям счётных признаков
| Признак | Значение признака | Число экземпляров | Пределы варьирования признака | ||
|---|---|---|---|---|---|
| Наши данные | Anderson, 1982 | Всего | |||
| Число позвонков | 89–95 | 89–96 | 89–96 | ||
| 89 | 2 | ||||
| 90 | 2 | ||||
| 91 | 1 | ||||
| 92 | 3 | ||||
| 93 | 2 | ||||
| 94 | – | ||||
| 95 | 2 | ||||
| Число лучей D | 82–89 | 80–88 | 80–89 | ||
| 82 | 2 | ||||
| 83 | 2 | ||||
| 84 | – | ||||
| 85 | 1 | ||||
| 86 | 1 | ||||
| 87 | – | ||||
| 88 | 2 | ||||
| 89 | 2 | ||||
| Число лучей A | 73–80 | 72–80 | 72–80 | ||
| 73 | 2 | ||||
| 74 | 2 | ||||
| 75 | – | ||||
| 76 | 2 | ||||
| 77 | 2 | ||||
| 78 | – | ||||
| 79 | – | ||||
| 80 | 2 | ||||
| Число свободных interneuralia | 0–5 | 3–5 | 0–5 | ||
| 0 | 2 | ||||
| 1 | 1 | ||||
| 2 | 7 | ||||
| 3 | – | ||||
| 4 | 1 | ||||
| 5 | 1 | ||||
Измерения нетиповых G. brevifenestratа TL 82−117 мм (n = 8) [в квадратных скобках, если отличаются, приведены по первоописанию данные типовых экземпляров (n = 10)]. В % SL: aD 22.4−26.7 [28.8] (в среднем 24.7 [26.0]), aA 34.2−38.3 [38.8] ([35.9] 36.3), c [14.3] 15.1−19.1 ([15.7] 17.9), hc 8.2−10.0 (8.9), wc [6.2] 7.8−11.4 ([7.8] 9.5), H1 7.3−11.0 ([9.2] 9.4), ширина тела в области Р 5.0−8.0 (6.2), H2 7.9−10.6 (9.0), lР 8.5−10.6 (9.5), о 4.5−5.8 (5.1), протяжённость очешуения 16.9−65.9 (34.4). В % c: aD 125−156 (139), hc 46−56 (50), wc 43.5−60.0 (53.0), H1 44.0−59.5 (52.0), H2 44.0−59.0 (50.6), lС 7.5−15.0 (10.0), lР 47.0−59.0 (52.0 [56.6]), о 25.0−32.1 [37.7] (28.4 [32.6]), ао [15.8] 19.2−25.2 ([17.3] 23.0), io 5.5−12.5 ([7.2] 8.8), lmx 24.4−54.8 ([45.2] 45.3), длина жаберного отверстия 16.2−25.3 ([18.3] 21.5), протяжённость очешуения 94−365 (189).
Прижизненная окраска самцов (при поимке TL 111.5 и 99.0 мм) яркая, зеленовато-жёлтая на сером или более светлом основном фоне (рис. 3). На боках неотчётливо выделяются до восьми широких поперечных полос, перемежающихся размытыми палевыми пятнами, которые при переходе на D становятся белыми или сероватыми и приобретают более чёткие очертания. Первая светлая полоса проходит на уровне середины длины грудного плавника. Щёки, низ головы и грудной плавник светлые. Жаберно-ротовая полость и перитонеум не пигментированы. На спинном плавнике у трёх из 19 экз. (16%) имеется чёрное глазчатое (окаймлённое белым) пятно (или stigma). Этот признак характерен для всех видов подсемейства Gymnelinae (но выражен не у всех экземпляров). Анальный плавник у обоих самцов с отчётливой красной полосой по краю. По аналогии с видами близкого рода Gymnelus, самцы которых всегда имеют чёрную полосу по краю А (в то время как А самок всегда светлый), можно предполагать наличие полового диморфизма по этому признаку и у G. brevifenestratа. Отметим, что у Gymnelus чёрный край А выражен и у мелких ювенильных особей, т.е. признак связан с полом, а не с половым созреванием. При хранении экземпляров в спирте их окраска выцветает и становится однотонной. Исчезает и красно-бурая полоса по краю анального плавника у самцов (в отличие от самцов Gymnelus, сохраняющих чёрную кайму из меланофоров по краю А и после фиксации).
G. brevifenestratа достигает TL 122 мм (ЗИН № 23953). [Это именно TL, но не SL, как было указано в первоописании (Anderson, 1982. Р. 53)]. SL 89 мм в качестве максимальной указана ошибочно (Froese, Pauly, 2019), на самом деле это длина голотипа (Anderson, 1982, 1994).
Половая зрелость, вероятно, наступает по достижении длины не менее 90−100 мм. Молодая самка SL 89 мм, выловленная в июне, имела 25−30 икринок диаметром 1.8−1.9 мм (голотип). Самец прижизненной длиной TL 99 мм имел в августе незрелые гонады (II стадии зрелости).
Сравнительные замечания. Сравнение дополнительных экземпляров с паратипами и первоописанием (Anderson, 1982) показывает хорошее соответствие большей части признаков, что, во-первых, подтверждает определение и, во-вторых, свидетельствует о высокой достоверности описания. Измерения близки данным типовой серии (aD, aA, H1, H2, wc, lР, io, lmx и др.). Некоторые расхождения имеются по измерениям длины головы (примерно на 2% SL), а также рыла и глаза, что может быть обусловлено изменением пропорций при хранении. Выявленная изменчивость в числе сейсмосенсорных пор обусловлена в значительной мере увеличением количества изученных экземпляров.
Следует отметить, что видовой диагноз G. brevifenestratа в работе Андерсона включает ряд признаков, общих (или широко перекрывающихся) с признаками других видов Gymnelopsis: это число лучей грудного плавника и ширина его основания, наличие зубов на сошнике и нёбных, пять жаберных лучей, число стабильных пор (1 interorbital, 4 postorbital, 3 occipital). Поскольку такие признаки не имеют значения для идентификации этого вида, они исключены из нашего уточнённого диагноза.
От остальных видов рода G. brevifenestrata отличается небольшим числом позвонков (89−96 против > 100) и лучей А (73−81 против 89−97), а также коротким жаберным отверстием, расположенным полностью над основанием Р (у прочих видов оно заходит несколько ниже, достигая 2−6-го луча Р).
В Охотском море помимо короткожаберного гимнелопса указаны G. ocellata, G. brashnikovi, G. humilis и G. ochotensis (Федоров и др., 2003). G. brevifenestrata отличается от G. ocellata помимо меньшего числа позвонков сдвинутым назад началом D (у последнего он начинается над началом Р) и полным развитием подглазничных пор (отсутствуют у G. ocellata). G. brevifenestrata отличается от G. brashnikovi и от G. humilis числом позвонков и положением D, начало которого находится над передней третью Р у первого и заметно позади конца Р – у второго. G. brevifenestrata отличается от G. ochotensis s. str. (голотип ЗИН № 30359) и G. japonica небольшим числом позвонков (89–96) и положением начала D (плавник начинается заметно позади конца Р у двух упомянутых видов).
Прежние исследователи относили часть экземпляров G. brevifenestrata к виду G. ocellatus Soldatov 1922 (=G. ocellata): это следует из авторских этикеток А.М. Попова (1931 г., ЗИН № 23953) и П.Ю. Шмидта (1941 г., ЗИН № 23944, 23952). Можно полагать, таким образом, что в публикации Шмидта (1950) сведения по G. ocellata основаны на смешанных с G. brevifenestrata материалах.
Вид рассматривался как новый в 1940-е гг. А.М. Поповым (этикетки “Gymnelopsis evermanni Popov” имеются у экземпляров ЗИН № 23944, 23952) и в 1953 г. А.П. Андрияшевым (этикетка “Gymnelopsis suvorovi sp. n., голотип” у экземпляра ЗИН № 33751), но эти материалы не были опубликованы.
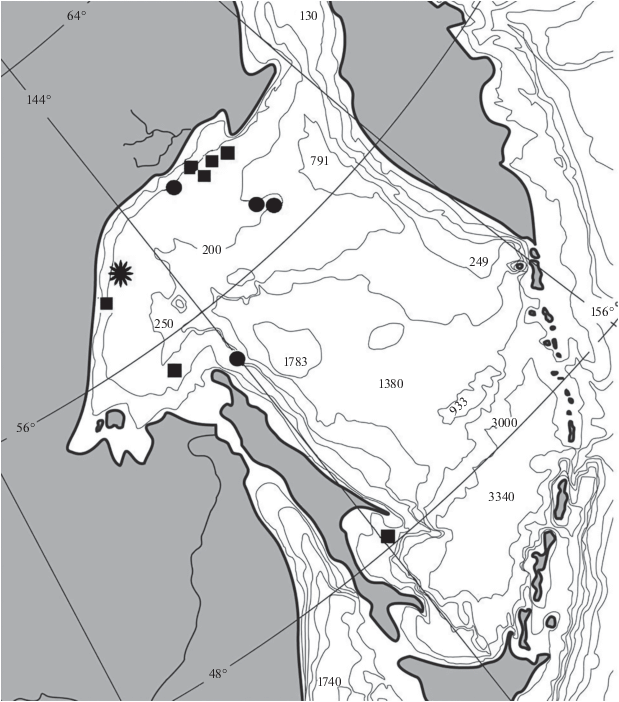
Распространение и данные по экологии. Типовые экземпляры G. brevifenestrata пойманы в Охотском море, в его северо-западной части и у северо-восточной оконечности Сахалина (рис. 4), на глубинах 100−200 м. Позднее, в каталоге материалов ЗИН, опубликованы сведения о трёх новых находках в Охотском море (Balushkin et al., 2011; Балушкин и др., 2012). Две из них подтверждаются настоящим исследованием (ЗИН № 44727, 48107), включая экземпляр из мелководного района восточнее южной части Сахалина (48°00′ с.ш., 144°01′ в.д., 76−80 м). Третий экземпляр (ЗИН № 46785: vert. 108, D 97, A 88) не относится к G. brevifenestrata.
Рис. 4.
Местонахождения Gymnelopsis brevifenestrata: (*) – голотип и пойманные вместе с ним паратипы ЗИН № 23944 и 23952, (⚫) – прочие паратипы, (◼) – нетиповые экземпляры; (——) − изобаты.
G. brevifenestrata считают эндемиком Охотского моря (Федоров и др., 2003). Однако он был отмечен и за его пределами, в водах префектуры Аомори, занимающей северную оконечность о‑ва Хонсю, Япония (Anderson, 1994). Это нахождение, нарушающее представление об охотоморском эндемизме вида, основано на экземпляре из сборов НИС “Витязь” (ст. 6668, без указания инвентарного номера коллекций ЗИН). Мы разыскали этот экземпляр (ЗИН № 48390). Как оказалось, он не относится к G. brevifenestrata (vert. 102, чешуя доходит вперёд до Р). Таким образом, эндемизм G. brevifenestrata для Охотского моря подтверждается. Все особи G. brevifenestrata пойманы на шельфе западной и северной части Охотского моря.
Глубины новых нахождений вписываются в известный ранее диапазон 76−200 м (Федоров и др., 2003): 97−150 м (Савельев и др., 2019) и 102−137 м (наши данные). Указание на поимки G. brevifenestrata на глубинах до 783 м (Froese, Pauly, 2019) ошибочно, так как приводится со ссылкой на работу, в которой батиметрический диапазон 70−783 м относится ко всем видам рода Gymnelopsis (Anderson, 1994), причём глубина 712–782 м указана для G. japonica и основана на его находке в Японском море у берегов Хоккайдо (USNM № 117956) (Anderson, 1994; Shinohara et al., 2011). Отсутствие G. brevifenestrata на больших глубинах подтверждается и тем, что в списке глубоководных рыб Японского моря он не значится (Shinohara et al., 2014).
Грунты в районах поимок (определены в четырёх случаях) включали ил, песок и камни в разных вариациях: жёлтый песок и камни, песчанистый ил с галькой, заиленный песок.
Придонная температура в районе поимки экземпляров к северо-западу от Сахалина (ЗИН № 44727) составляла 1.73°С. На севере Охотского моря G. brevifenestrata наиболее обычен в пределах вод с отрицательными значениями температуры: −1.0…−1.8°С (Савельев и др., 2019).
Зоогеографическая характеристика. G. brevifenestrata относили к группе приазиатских высокобореальных видов (Федоров и др., 2003; Парин и др., 2014). Однако характеристика “высокобореальный” требует уточнения. На самом деле это гляциально-охотоморский вид, входящий в состав охотоморской холодноводной фауны, приуроченной к переохлаждённым водам арктического характера (Савельев и др., 2019).
Список литературы
Балушкин А.В., Шейко Б.А., Природина В.П. 2012. Каталог фондовой коллекции Зоологического института РАН. Класс костистые рыбы (Osteichthyes). Отряд окунеобразные (Perciformis). Подотряд Zoarcoidei. Семейства Stichaeidae, Pholidae, Anarhichadidae // Исследования фауны морей. Т. 72. СПб.: Изд-во ЗИН РАН, 136 с.
Линдберг Г.У., Красюкова З.В. 1975. Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Ч. 4. Л.: Наука, 463 с.
Назаркин М.В., Чернова Н.В. 2003. Новый вид бельдюговых рыб, Gymnelopsis humilis sp. nov. (Zoarcidae) из северной части Охотского моря // Вопр. ихтиологии. Т. 43. № 5. С. 602–606.
Парин Н.В., Евсеенко С.А., Васильева Е.Д. 2014. Рыбы морей России: аннотированный каталог. М.: Т-во науч. изд. КМК, 733 с.
Савельев П.А., Метелёв Е.А., Сергеев А.С., Данилов В.С. 2019. Видовой состав и распределение донных рыб в элиторали северо-западной части Охотского моря в летний период // Вопр. ихтиологии. Т. 59. № 4. С. 405–415. https://doi.org/10.1134/S0042875219040179
Солдатов В.К., Линдберг Г.У. 1930. Обзор рыб дальневосточных морей // Изв. ТИНРО. Т. 5. 576 с.
Федоров В.В., Черешнев И.А., Назаркин М.В. и др. 2003. Каталог морских и пресноводных рыб северной части Охотского моря. Владивосток: Дальнаука, 202 с.
Чернова Н.В. 1998. Восстановление валидности вида Gymnelus bilabrus Andriashev, 1937 с уточнением видовой характеристики G. viridis (Fabricius, 1780) (Zoarcidae) // Вопр. ихтиологии. Т. 38. № 2. С. 182–188.
Шейко Б.А., Федоров В.В. 2000. Класс Cephalaspidomorphi – Миноги. Класс Chondrichthyes – Хрящевые рыбы. Класс Holocephali – Цельноголовые. Класс Osteichthyes – Костные рыбы // Каталог позвоночных Камчатки и сопредельных морских акваторий. Петропавловск-Камчатский: Камчат. печат. двор. С. 7–69.
Шмидт П.Ю. 1950. Рыбы Охотского моря. М.; Л.: Изд-во АН СССР, 370 с.
Anderson M.E. 1982. Revision of the fish genera Gymnelus Reinhardt and Gymnelopsis Soldatov (Zoarcidae), with two new species and comparative osteology of Gymnelus viridis // Nat. Mus. Can. Natur. Sci. Publ. Zool. № 17. 76 p.
Anderson M.E. 1994. Systematics and osteology of the Zoarcidae (Teleostei: Perciformes) // Ichthyol. Bull. J.L.B. Smith Inst. Ichthyol. №. 60. 120 p.
Anderson M.E., Fedorov V.V. 2004. Family Zoarcidae Swainson 1839 – eelpouts // Calif. Acad. Sci. Annot. Checklists Fish. № 34. P. 1–58.
Balushkin A.V., Sheiko B.A., Fedorov V.V. 2011. Catalog of the archival collection of the Zoological Institute, Russian Academy of Sciences: class Osteichthyes (bony fishes), order Perciformes, family Zoarcidae // J. Ichthyol. V. 51. № 10. P. 950–1034. https://doi.org/10.1134/S0032945211100031
Froese R., Pauly D. (eds.). 2019. FishBase. World Wide Web electronic publication (http://www.fishbase.org. Version 02/2019).
Katayama M. 1943. On two new ophidioid fishes from the Japan Sea // Ann. Zool. Jpn. V. 22. № 2. P. 101–104.
Masuda H., Amaoka K., Araga C. et al. 1984. The fishes of the Japanese Archipelago. Tokyo: Tokai Univ. Press, 437 p.+ + pls. 370.
Shinohara G., Shirai A.M., Nazarkin M.V., Yabe M. 2011. Preliminary list of the deep-sea fishes of the Sea of Japan // Bull. Nat. Mus. Natur. Sci. Ser. A. V. 37. № 1. P. 35–62.
Shinohara G., Nakae M., Ueda Y. et al. 2014. Annotated checklist of deep-sea fishes of the Sea of Japan // Nat. Mus. Natur. Sci. Monographs. № 44. P. 225–291.
Toyoshima M. 1981. Revision of the eelpout genus Derjuginia // Jpn. J. Ichthyol. V. 28. № 3. P. 254–258.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии