

Вопросы ихтиологии, 2020, T. 60, № 4, стр. 426-441
Динамика популяции леща Abramis brama Псковско-Чудского озера
М. Б. Данилов 1, *, Е. А. Криксунов 1, А. Е. Бобырев 2, 3, А. Д. Шереметьев 3, М. М. Мельник 4, С. О. Северин 4, П. В. Васильев 1, С. В. Чистов 1
1 Московский государственный университет
Москва, Россия
2 Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
3 Всероссийский научно-исследовательский институт рыбного хозяйства и океанографии – ВНИРО
Москва, Россия
4 Псковский филиал Всероссийского научно-исследовательского института рыбного хозяйства
и океанографии – ПсковНИРО
Псков, Россия
* E-mail: danilov84@rambler.ru
Поступила в редакцию 25.07.2019
После доработки 06.08.2019
Принята к публикации 07.08.2019
Аннотация
С помощью когортных моделей проанализирована динамика популяции леща Abramis brama в Псковском и Чудском озёрах в 1980−2009 гг. Стабильное возрастание запаса леща в Псковском озере наблюдалось до 1999 г., в Чудском – до 2004 г., затем наметилась тенденция к его снижению. В 2000-х гг. зарегистрировано снижение среднего возраста в популяции, обусловленное как ростом интенсивности промысла, так и высоким пополнением в конце 1990-х гг. Высокочисленные генерации леща появляются в обоих озёрах синхронно. Во второй половине 2000-х гг. в обоих озёрах отмечены низкие значения пополнения. Зависимость в системе запас–пополнение наилучшим образом отображает кривая Рикера. Выявлена связь величины пополнения леща с прозрачностью и температурой воды, числом дней в году с температурой воздуха выше 20°С и среднегодовым уровнем воды. Существующий уровень промысловой нагрузки на запас леща близок к оптимальному. Дальнейшее увеличение интенсивности промысла в обоих озёрах нецелесообразно. Текущий уровень равновесного улова может быть достигнут при гораздо меньшей интенсивности промысла.
Вопросы организации и регулирования промысла рыб в Псковско-Чудском озере, в том числе леща Abramis brama как одного из основных объектов добычи, привлекают большое внимание специалистов (Kangur, 1996; Pihu, 1996; Pihu, Kangur, 2001; Мельник, Концевая, 2005; Kangur et al., 2007; Концевая, 2009; Saat et al., 2010; Бобырев и др., 2013). В рамках исследования динамики популяций основных промысловых видов рыб Псковско-Чудского озера в одной из наших недавних работ (Данилов и др., 2018) сделана попытка проанализировать состояние запасов, промысла и воспроизводства судака Sander lucioperca в 1980−2009 гг. на основе методов ретроспективного анализа, а также некоторых традиционных моделей взаимодействий в системе запас–пополнение. Используя данный методический подход, мы продолжили начатые исследования и в настоящей работе уделили внимание популяции леща, доля которого в общем улове рыб Псковско-Чудского озера может достигать в отдельные годы 30%, в среднем составляя около 9 и 15% годового улова для Чудского и Псковского озер соответственно.
Хотя сами процессы популяционных изменений всё ещё остаются слишком сложными для их надёжной предсказуемости, ретроспективный анализ статистики промысла позволяет провести обобщение накопленных данных, увязать изменения основных популяционных показателей с результатами промысловой деятельности на водоёме за период, охватывающий события новейшей истории. Проведению такого анализа посвящена настоящая работа, рассматривающая ход популяционных процессов в период с 1980 по 2009 гг.
МАТЕРИАЛ И МЕТОДИКА
Исследование базируется на результатах наблюдений в 1980−2009 гг., представленных в публикациях и фондовых материалах Псковского отделения ГосНИОРХ (с 2018 г. – Псковский филиал ВНИРО), включающих выполненные для разных сезонов каждого года оценки соотношения возрастных групп в уловах различных орудий (ставные сети, механизированные мутники, заколы), данные по линейному и весовому росту, плодовитости, темпу полового созревания, общему годовому улову, температурному, уровенному режимам водоёма и т.д. В анализ вовлечены лишь данные официальной статистики; объёмы нелегального вылова не учитывали. Статистическая обработка данных выполнена с помощью пакета прикладных программ Statistica 6.0.
Предварительно для каждого календарного года выполнен расчёт возрастного состава улова леща, в качестве которого рассматривалась средневзвешенная по сезонам и орудиям лова оценка соотношения разных возрастных групп в суммарном годовом объёме добычи. Возрастной ряд ограничен возрастом 17+, поскольку поимки рыб более старшего возраста единичны и не образуют представительную выборку для анализа. Непрерывные ряды уловов, подразделённых на возрастные категории, использованы как входные переменные для когортного анализа, параметры которого заданы оценками возрастной динамики относительной улавливаемости рыб, их естественной смертности, темпа полового созревания, терминальных значений промысловой смертности. Ввиду относительной обособленности Псковского и Чудского озёр и независимой деятельности рыбодобывающих предприятий, ведущих промысел в отдельных частях водоёма, когортный анализ выполнен раздельно для двух субъединиц запаса леща, локализованных в Чудском и Псковском озёрах. Для Чудского озера используются объединённые данные для эстонской и российской частей водоёма.
В качестве основного средства ретроспективного анализа состояния запасов и промысла леща использована аналитическая когортная модель ADAPT-VPA (версия 2.8) (Gavaris, 1988, 1993; Mohn, Cook, 1993; Patterson, Kirkwood, 1995), реализованная в пакете прикладных программ Fisheries Toolbox (National Oceanic and Atmospheric Administration − NOAA) / National Marine Fisheries Services − NMFS, США). В качестве настроечных индексов использовали различные показатели улова на единицу промыслового усилия: тонны промыслового улова на одного рыбака, численность улова сеголеток за 1 ч траления, численность и биомасса уловов молоди (длиной < 30 см) и взрослых рыб (длиной > 30 см) за 1 ч траления экспериментальным тралом.
Описание линейного и весового роста выполнено с использованием функции Берталанфи. Оценки возрастных значений мгновенных коэффициентов естественной смертности получены методом Чена−Ватанабе (Chen, Watanabe, 1989), связывающим темп убыли с параметрами созревания и линейного роста рыб; оценки коэффициентов относительной улавливаемости – методом Южаковой−Снеткова (Южакова, Снетков, 1983; Криксунов, Концевая, 1987).
Для оценки современного состояния промысла леща использована модель динамического запаса (Бивертон, Холт, 1969), реализованная с помощью прикладной программы YPR (Yield per Recruit Model) версия 3.1 (NOAA/NMFS, США). Проанализированы изменения численности рекрутов (2+) и относительной биомассы нерестового стада леща SSB/R (Gabriel et al., 1989). Методика анализа описана ранее (Данилов и др., 2018). В ходе проведённого анализа получены также описания связей между величиной родительского стада леща и численностью продуцируемого потомства моделями пополнения Бивертона−Холта, Рикера и Шепарда. Последняя модель (Shepherd, 1982) представляет собой формальное обобщение двух первых и обеспечивает (за счёт введения дополнительного параметра формы) бóльшую гибкость в возможности описания эмпирических данных по сравнению с классическими функциями.
Изучение факторов, влияющих на изменчивость пополнения леща в Чудском озере, проведено на основе использования обобщённых линейных моделей (Generalized Linear Models – GLM), преимуществом которых перед традиционным регрессионным анализом является отсутствие ограничений на тип распределения данных. В качестве зависимой переменной рассматривалась численность пополнения, в качестве независимых – биомасса нерестового стада, среднегодовые уровень, температура и прозрачность воды, число дней в году с температурой воздуха выше 20°С. Независимые переменные использованы в анализе по отдельности и в различных сочетаниях. При выборе наилучшего статистического описания основными показателями служили величина остаточной дисперсии и информационный критерий Акайке (Akaike Information Criterium – AIC), оценивающий качество аппроксимации с учетом числа параметров (степеней свободы) модели. Анализ осуществлен в программной среде R (версия 3.0).
РЕЗУЛЬТАТЫ
Рост и смертность. Различия между значениями параметров уравнения линейного роста леща Псковского и Чудского озера очень малы, значительно меньше величин стандартных ошибок. Поэтому для описания линейного роста используется обобщённое уравнение вида: Lt = 72 × (1 – e0.09(t– 0.8)), где Lt – средняя длина тела рыб в возрасте t, см.
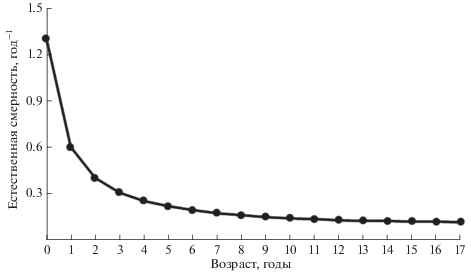
Столь же большое сходство демонстрируют значения возрастных коэффициентов естественной смертности. Они варьируют в пределах 1.30−0.12 год–1, монотонно снижаясь с увеличением возраста (рис. 1). Средневзвешенные оценки мгновенных коэффициентов естественной смертности составили 0.18 год–1 для Чудского озера и 0.19 год–1 – для Псковского.
Рис. 1.
Динамика изменения мгновенного коэффициента естественной смертности леща Abramis brama в зависимости от возраста, усреднённые данные для Псковского и Чудского озёр.
Динамика обилия и возрастной состав популяций. За исследуемый период уловы леща в Чудском озере выросли почти в пять раз: минимальный улов (190.3 т) зарегистрирован в 1980 г., максимальный (1027.0 т) – в 2007 г., средний вылов составил 504.9 т. (рис. 2а). В Псковском озере уловы леща также значительно возросли (примерно в 6 раз) и варьировали от 52.8 (1980 г.) до 322.0 т (2002–2003 гг.), в среднем составив 163.2 т (рис. 2а). Коэффициент вариации уловов (CV) был достаточно высоким и имел сходные значения в обоих озёрах – соответственно 50 и 43%. Наиболее существенный рост добычи пришёлся на вторую половину 1990-х гг. В Чудском озере возрастание уловов продолжалось вплоть до конца 2000-х гг., в Псковском озере уловы стали снижаться с середины 2000-х гг. Статистическая связь уловов в обоих озёрах довольно велика – коэффициент корреляции (r) равен 0.75 (p < 0.001).
Рис. 2.
Динамика уловов (а) и биомассы (б) леща Abramis brama в Чудском (——) и Псковском (- - -) озёрах.
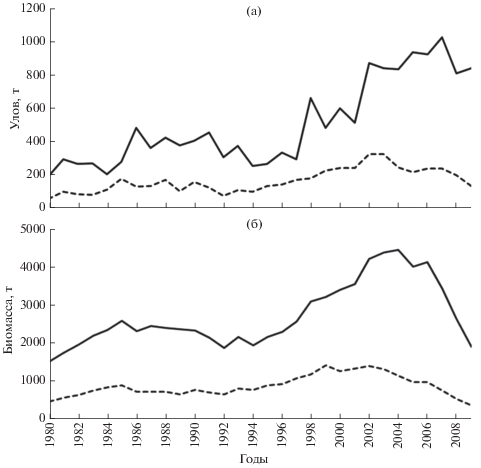
Сходно с уловами изменялась общая биомасса запаса леща. В Чудском озере её оценки варьировали от 1510 (1980 г.) до 4445 (2004 г.) т, составляя в среднем 2713 т; в Псковском − от 345 (2009 г.) до 1408 т (1999 г.), в среднем 859 т (рис. 2б). Осреднённая оценка общей численности популяции (возрастные группы 1+–17+) составила в Чудском озере 10.1 млн экз., в Псковском − 3.3 млн экз.
В изменениях запаса леща в Псковском и Чудском озёрах прослеживается определённая сопряжённость, о чём свидетельствует достаточно высокий уровень связи между изменениями его биомассы в этих озёрах (r = 0.77, p < 0.001) и численности (r = 0.76, p < 0.001). И в том, и в другом озере до начала 2000-х гг. прослеживалась общая тенденция к возрастанию запасов, которая затем сменилась на противоположную, обусловив тем самым возврат популяции к уровню, характерному для начала рассматриваемого периода (начало 1980-х гг.).
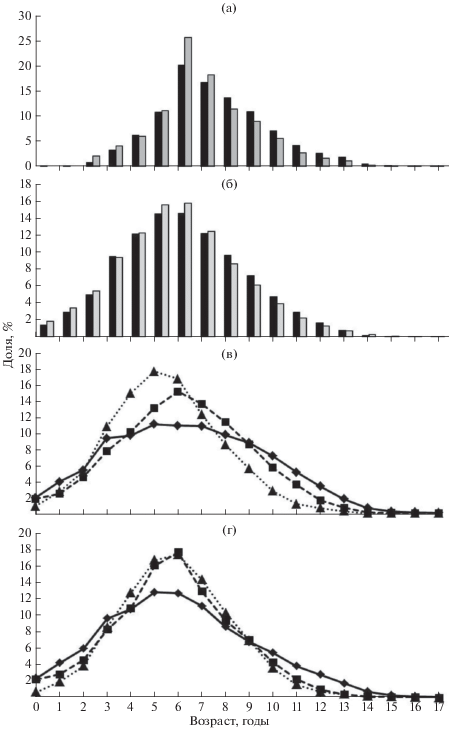
И в Псковском, и в Чудском озёрах основу биомассы популяции составляют рыбы возраста 4–7 лет (53–56%), эти же группы формируют большую часть улова (рис. 3а, 3б). Рассматривая изменения возрастного состава леща по десятилетиям, можно отметить некоторое увеличение среднего возраста рыб в 1990-х гг., происходящее на фоне общего роста запаса. В 2000-х гг. средний возраст рыб снизился, возможно, из-за роста интенсивности промысла или вступления в промысел высокочисленных генераций конца 1990-х гг. (рис. 3в, 3г).
Рис. 3.
Возрастной состав улова (а) и запаса (б) леща Abramis brama в Чудском (◼) и Псковском ( ) озёрах (усреднённые данные за 1980−2009 гг.), а также возрастной состав запаса в
Чудском (в) и Псковском (г) озёрах в 1980-х (- ◼ -), 1990-х (−◆−) и 2000-х (···▲···) гг.
) озёрах (усреднённые данные за 1980−2009 гг.), а также возрастной состав запаса в
Чудском (в) и Псковском (г) озёрах в 1980-х (- ◼ -), 1990-х (−◆−) и 2000-х (···▲···) гг.
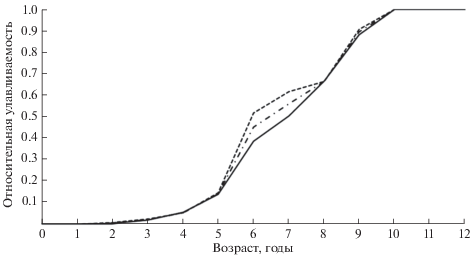
Оценка возрастных коэффициентов относительной улавливаемости показала, что уже в 2-летнем возрасте часть молодых особей леща становятся доступными для используемых орудий лова. С возрастом доля рекрутов плавно возрастает, достигая 100% к 10-у году жизни (рис. 4). Темп вступления в состав промыслового запаса особей разного возраста в наибольшей степени связан с переходом к образу жизни взрослых рыб, поэтому представленную на рис. 4 обобщённую кривую можно рассматривать как огиву созревания.
Рис. 4.
Коэффициенты относительной улавливаемости леща Abramis brama разного возраста в Чудском и Псковском озёрах; (−•−) − обобщённая кривая, обозначения озёр см. на рис. 2.
Интенсивность эксплуатации. В Чудском озере динамика коэффициентов промысловой смертности, взвешенных по численности и биомассе запаса, демонстрирует слабый отрицательный тренд, обусловленный увеличением запаса и улова (рис. 5а). В Псковском озере взвешенные коэффициенты промысловой смертности имели высокие значения в начале 1980-х и в конце 2000-х гг. (рис. 5б), что связано с низкими значениями запаса и уловов в эти периоды. В Чудском озере в динамике промысловой смертности леща отчётливо проявляется цикличность с периодом ~ 10 лет.
Рис. 5.
Интенсивность эксплуатации леща Abramis brama в Чудском и Псковском озёрах в 1980−2009 гг.: а − взвешенные по численности (×106) и по биомассе (×103) коэффициенты промысловой смертности в Чудском озере, б – то же в Псковском озере; в − средний коэффициент промысловой смертности разных возрастных групп в 1980-х, 1990-х и 2000-х гг. в Чудском озере; г – то же в Псковском озере; д − коэффициент эксплуатации в Чудском и Псковском озёрах; взвешенные коэффициенты: (···) – по численности, (−•−) – по биомассе; ост. обозначения см. на рис. 2 и 3.
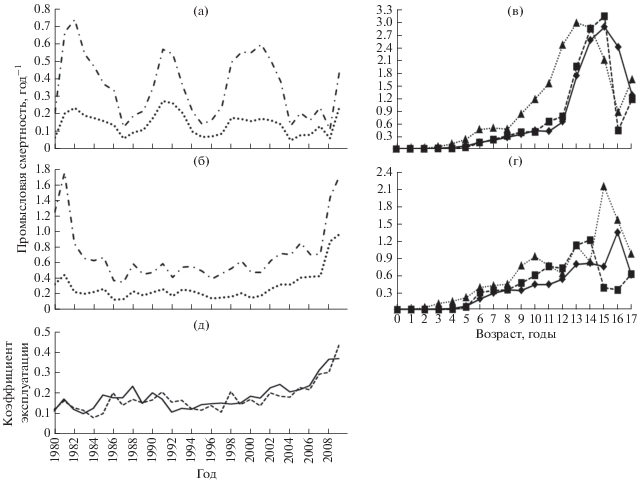
Максимальные значения промысловой смертности леща приходятся на старшие возрастные группы – 10–15 лет (рис. 5в, 5г). Начиная с 2000-х гг. просматривается тенденция к повышению эксплуатации рыб средних возрастных групп.
Коэффициенты эксплуатации (отношение массы улова к общей биомассе запаса) в Чудском и Псковском озёрах изменялись в пределах соответственно 0.11−0.38 (0.19) и 0.09−0.44 (0.18) (рис. 5д). В обоих озёрах они имеют сходную динамику, возрастая в конце 2000-х гг.
В пользу возрастающей промысловой нагрузки говорит увеличение числа промысловых рыбаков в Чудском озере с начала 1990-х гг. почти в три раза; в Псковском озере число рыбаков оставалось относительно стабильным, с локальным повышением в начале 2000-х гг. Однако в динамике количества орудий лова в целом по водоёму прослеживается обратная тенденция: число крупноячейных ставных сетей и заколов снизилось к середине 1990-х гг. соответственно на 60 и 47%. Разнонаправленность трендов в динамике факторов, обусловливающих изменения интенсивности промысловой эксплуатации, может быть вызвана недоучётом продолжительности использования указанных орудий лова.
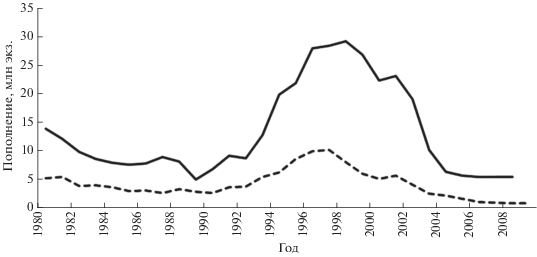
Урожайность молоди. Численность пополнения (0+) леща варьировала в широком диапазоне (CV = 59%): в Чудском озере − 5.0−29.1 (13.2) млн экз., в Псковском – 0.9−10.1 (4.2) млн экз. В обоих озёрах появление высокочисленных возрастных классов приходится на одни и те же годы − 1995–2003 (рис. 6).
Рис. 6.
Динамика численности пополнения (0+) леща Abramis brama в Чудском и Псковском озёрах; обозначения озёр см. на рис. 2.
На рис. 7 представлены результаты аппроксимации связи в системе родители–потомки, полученные с помощью различных моделей пополнения. Заметим, что лишь одна из них, а именно модель Рикера, даёт статистически значимое описание данных, относящихся к Чудскому озеру. В качестве основного средства оценки надёжности аппроксимации использованы t-критерий Стьюдента и величина стандартной ошибки для параметров аппроксимирующих функций (Фёрстер, Рёнц, 1981). Представленные данные указывают на то, что определённая роль в формировании пополнения леща принадлежит внутренним механизмам популяционной регуляции, сдерживающим эффективность воспроизводства при высокой численности родительского запаса. Это отчётливо видно на рис. 7а, интерпретирующем данные воспроизводства леща в Чудском озере. После определённых значений дальнейшее увеличение родительского запаса приводит к снижению численности потомства. Отсюда относительно хорошее соответствие эмпирических данных теоретической модели Рикера для Чудского озера. В то же время из приведённых графиков следует, что значительное влияние на урожайность молоди леща оказывают не учитываемые моделью факторы, порождающие наблюдаемые отклонения оценок обилия от модельного предсказания. Поскольку такие факторы предположительно связаны с межгодовыми вариациями гидроклиматического режима водоёма, представляет интерес выявление возможных связей между динамикой урожайности молоди в разные годы анализируемого периода и условиями среды. Результаты статистического анализа сопряжённости динамики пополнения (0+) леща с условиями внешней среды и величиной родительского запаса, выполненного с использованием моделей GLM, представлены в табл. 1. Поскольку распределение оценок пополнения леща близко к логнормальному, зависимая переменная преобразована к виду x← ln(x).
Рис. 7.
Аппроксимация связи биомасса−пополнение леща Abramis brama Чудского (а, в) и Псковского (б, г) озёр функциями Рикера (а, б), Шепарда (в) и Бивертона−Холта (г).
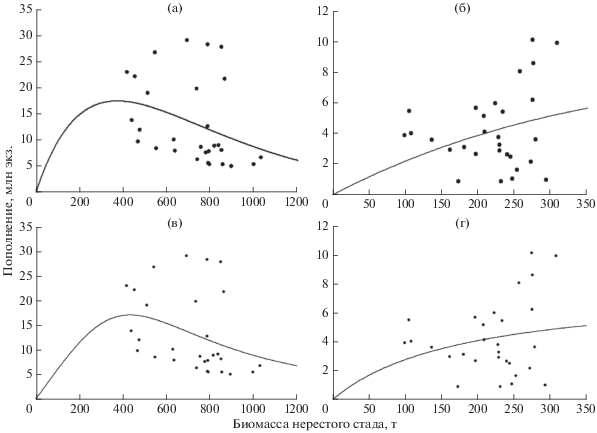
Таблица 1.
Регрессионные модели, тестированные методом обобщённых линейных моделей (GLM)
| № | Модель | Остаточная дисперсия |
AIC |
|---|---|---|---|
| 1 | lnR ~ 17.291000 – 0.001485 SSB | 7.707 | 49.87 |
| 2 | lnR ~16.227552 – 0.004663 H | 8.993 | 54.34 |
| 3 | lnR ~ 16.2276 + 0.2223 T | 8.589 | 53.01 |
| 4 | lnR ~ 16.091225 + 0.001359 D | 9.353 | 55.48 |
| 5 | lnR ~ 10.50 + 3.26 Sd | 7.873 | 50.49 |
| 6 | lnR ~17.214130 – 0.001378 SSB – 0.002881 H | 7.522 | 51.16 |
| 7 | lnR ~ 17.173811 – 0.001321SSB + 0.164486 T | 7.228 | 50.01 |
| 8 | lnR ~ 17.157580 – 0.001478 SSB + 0.001282 D | 7.573 | 51.36 |
| 9 | lnR ~10.317979 – 0.001843SSB + 4.111582 Sd | 5.218 | 40.56 |
| 10 | lnR ~16.227552 – 0.004691 H + 0.223103 T | 8.072 | 53.21 |
| 11 | lnR ~16.095669 – 0.004619 H + 0.001314 D | 8.852 | 55.89 |
| 12 | lnR ~ 9.506010 – 0.006747 H + 3.822803 Sd | 6.853 | 48.46 |
| 13 | lnR ~ 16.209890 + 0.218139 T + 0.000176 D | 8.586 | 55.00 |
| 14 | lnR ~ 10.7262 + 0.2058 T + 3.1288 Sd | 7.092 | 49.46 |
| 15 | lnR ~ 0.1057 + (4.617 × 10–4) D + 3.191 Sd | 7.856 | 52.42 |
| 16 | lnR ~ 17.086008 – 0.001199 SSB – 0.003134 H + 0.170359 T | 7.010 | 51.12 |
| 17 | lnR ~ 17.083922 – 0.001372 SSB – 0.002846 H + 0.001260 D | 7.392 | 52.66 |
| 18 | lnR ~ 9.613544 – 0.001690 SSB – 0.004904 H + 4.449793 Sd | 4.698 | 39.51 |
| 19 | lnR ~ 17.13407 – 0.00133 SSB + 0.15309 T + 0.00046 D | 7.214 | 51.95 |
| 20 | lnR ~ 10.473432 – 0.001704 SSB + 0.126756 T + 3.966671 Sd | 4.937 | 40.95 |
| 21 | lnR ~ 10.336558 – 0.001841 SSB + 0.000113 D + 4.093697 Sd | 5.217 | 42.55 |
| 22 | lnR ~ 16.2155423 – 0.0046867 H + 0.2202399 T + 0.0001197 D | 8.071 | 55.21 |
| 23 | lnR ~ 9.7415 – 0.0067 H + 0.2039 T + 3.6889 Sd | 6.086 | 47.02 |
| 24 | lnR ~ 10.6221846 + 0.2234750 T – 0.0007612 D + 3.2314299 Sd | 7.052 | 51.29 |
| 25 | lnR ~ 17.0524814 – 0.0012075 SSB – 0.0031084 H + 0.1604913 T + 0.0003964 D | 6.999 | 53.07 |
| 26 | lnR ~ 9.757577 – 0.001540 SSB – 0.005037 H + 0.132957 T + 4.306927 Sd | 4.389 | 39.54 |
| 27 | lnR ~ 9.609 – (1.690 × 10–3) SSB – (4.907 × 10–3) H – (2.768 × 10–5) D + 4.454 Sd | 4.698 | 41.51 |
| 28 | lnR ~ 10.3871987 – 0.0016999 SSB + 0.1417114 T – 0.0006358 D + 4.0502434 Sd | 4.909 | 42.79 |
| 29 | lnR ~ 9.585165 – 0.006817 H + 0.227529 T – 0.001018 D + 3.835904 Sd | 6.016 | 48.68 |
| 30 | lnR ~ 9.6281436 ‒ 0.0015308 SSB – 0.0051436 H + 0.1529036 T – 0.0008424 D + + 4.4248672 Sd | 4.341 | 41.22 |
Примечание. AIC − информационный критерий Акайке, R – величина пополнения, экз.; SSB – биомасса нерестового стада, т; H – среднегодовой уровень воды, по данным станции наблюдения Раскопель, см; Т – среднегодовая температура воды, по данным станции наблюдения Раскопель, °С; D – число дней в году с температурой воздуха выше 20°С, дни; Sd – прозрачность по диску Секки, м (по: Kangur et al., 2013).
Данные табл. 1 указывают на наличие отрицательной связи величины пополнения леща с биомассой нерестового стада и среднегодовым уровнем воды (уравнения 1–2) и положительной связи с прозрачностью, температурой воды, числом дней в году с температурой воздуха выше 20°С (уравнения 3–5). Принимая во внимание экологические особенности воспроизводства леща, отрицательная связь с уровнем воды представляется сомнительной. Действительно, из результатов анализа входных временны́х рядов следует, что отрицательная связь с уровенным режимом и биомассой нерестового стада определяется наличием разнонаправленных трендов: положительным для величины пополнения и отрицательным для среднегодового уровня воды, особенно с середины 1990-х до середины 2000-х гг., а также положительным для биомассы нерестового стада и отрицательным для пополнения, особенно в начале 1980-х гг. При удалении трендов знак зависимости меняется с отрицательного на положительный. Таким образом, можно утверждать, что, несмотря на общую тенденцию к понижению уровня воды в Псковско-Чудском водоёме, его отклонения от основного тренда в положительную сторону способствуют появлению более мощных возрастных классов леща. Так, в 1999 г. фиксировался уровень воды, близкий к абсолютному максимуму, а в 2003 и 2006–2007 гг. – близкий к абсолютному минимуму (Mäemets et al., 2018). Одновременно в указанные годы наблюдались соответствующие максимумы и минимумы в динамике пополнения леща.
С формальной точки зрения наилучшим вариантом описания динамики пополнения леща является уравнение 26 (табл. 1), связывающее величину пополнения с биомассой нерестового стада, прозрачностью, температурой и среднегодовым уровнем воды. В то же время отдельные факторы, привлекаемые к анализу, обладают примерно равной предсказательной способностью. Возможно, это связано со сложной организацией процессов формирования урожайности поколений, при которой помимо биомассы нерестового стада важен одновременный вклад нескольких абиотических факторов – уровня, прозрачности и температуры воды.
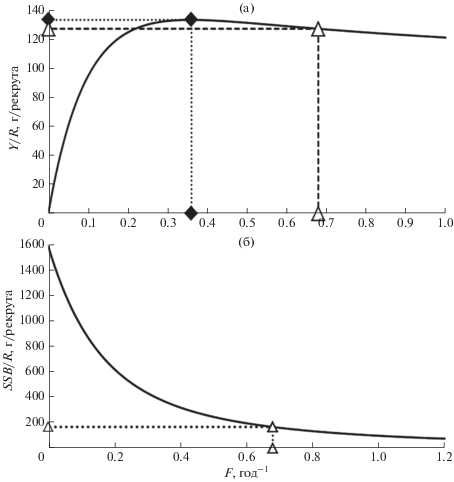
Оценка равновесного вылова. Анализ модели динамического запаса показывает, что в Чудском озере при интенсивности промысла на уровне второй половины 2000-х гг. (F2005–2009 = 0.68 год–1) обеспечивается равновесный улов леща, составляющий 127.7 г в пересчёте на одного рекрута (двугодовика) (рис. 8а). Данная оценка на 5% ниже максимально возможной – 133.9 г/рекрута, которая может быть достигнута при значительно меньшей промысловой смертности Fmax = 0.36 год–1. Полученной оценке величины относительного вылова соответствует равновесная биомасса нерестового запаса 164.8 г/рекрута. Это около 10% максимальной биомассы нерестового запаса на единицу пополнения (1578 г/рекрута), которая могла бы быть достигнута при отсутствии промысла (рис. 8б). Для леща Псковского озера данные параметры имели сходные значения: Fmax = 0.35 год–1, Y/R(Fmax) = 131.6 г/рекрута; F2005–2009 = 0.66 год–1, Y/R (F2005–2009) = 125.6 г/рекрута.
Рис. 8.
Зависимость равновесного улова (Y/R) (а) и биомассы нерестового стада (SSB/R) (б) от промысловой смертности (F) леща Abramis brama в Чудском озере: (◆) − Fmax, (△) – F 2005−2009 гг..
При увеличении возраста вступления в промысел с 2 до 8 лет максимальный равновесный улов незначительно возрастает с 133.9 до 140.1 г/рекрута, а при увеличении возраста до 10 лет снижается до 137.1 г/рекрута (табл. 2). При этом значения коэффициента промысловой смертности возрастают с 0.36 до 0.42 год–1, а равновесная биомасса нерестового стада – с 356.4 до 450.4 г/рекрута.
Таблица 2.
Зависимость максимального равновесного улова (Y/R) и биомассы нерестового стада (SSB/R) от промысловой смертности (Fmax) при разном возрасте вступления рекрутов леща Abramis brama в промысел в Чудском озере
| Возраст, годы | Fmax, год–1 | ${{{Y \mathord{\left/ {\vphantom {Y R}} \right. \kern-0em} R}}_{{{{F}_{{\max }}}}}}$, г/рекрута | ${{{{SSB} \mathord{\left/ {\vphantom {{SSB} R}} \right. \kern-0em} R}}_{{{{F}_{{\max }}}}}}$, г/рекрута |
|---|---|---|---|
| 2 | 0.36 | 133.9 | 356.4 |
| 3 | 0.36 | 134.0 | 356.9 |
| 4 | 0.36 | 134.6 | 359.5 |
| 5 | 0.37 | 135.6 | 355.6 |
| 6 | 0.39 | 136.9 | 352.0 |
| 7 | 0.41 | 139.2 | 380.6 |
| 8 | 0.43 | 140.1 | 413.8 |
| 9 | 0.42 | 138.7 | 441.1 |
| 10 | 0.42 | 137.1 | 450.4 |
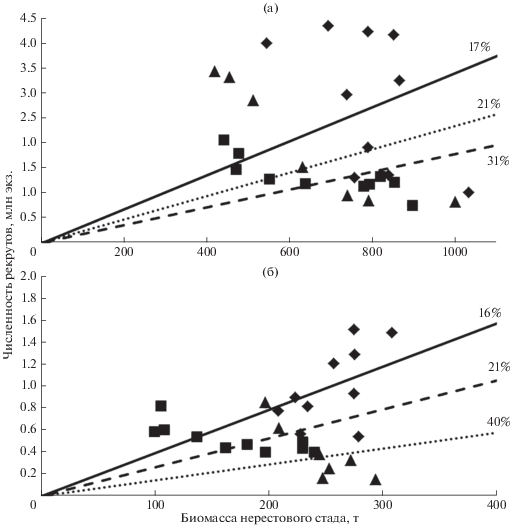
Замещающий уровень пополнения. В Чудском озере отношение биомассы нерестового стада (SSB) к численности рекрутов (R) (возрастная группа 2+) определено для разных десятилетий. Для 1980-х гг. наклон прямой, выраженный в процентах максимального уровня, составил 31%, в 1990-е – 17%, в 2000-е – 21%, в среднем за весь период − 22% (рис. 9а). Несколько иное соотношение характерно для Псковского озера − соответственно 21, 16 и 40%, в среднем за период также 22% (рис. 9б). В 1990-е гг. условия среды были благоприятны для формирования высокочисленных годовых классов. Точки, относящиеся к отдельным годам, располагаются на диаграмме рассеивания выше линии замещения, что говорит о достаточности пополнения для поддержания запаса на стабильном уровне при сложившейся интенсивности промысла (Чудское озеро – F = 0.84 год–1, Псковское озеро – F = 0.43 год–1). Во второй половине 2000-х гг. урожайность пополнения снизилась, в отдельные годы его уровень оказался недостаточным для стабильного поддержания запаса при текущем режиме промысла (Чудское озеро – F = 1.14 год–1, Псковское озеро – F = 0.71 год–1).
Рис. 9.
Соотношение между биомассой нерестового стада и численностью рекрутов (2+) леща Abramis brama в Чудском (а) и Псковском (б) озёрах в 1980-х, 1990-х и 2000-х гг.; обозначения см. на рис. 3.
ОБСУЖДЕНИЕ
Количественные оценки, характеризующие состояние запасов и промысла леща, охватывают относительно непродолжительный период. С начала 1980-х до конца 1990-х гг. запасы леща в Псковском Чудском озёрах увеличивались. Особенно благоприятные условия для воспроизводства запаса сложились во второй половине 1990-х гг., когда популяция пополнилась рядом высокоурожайных генераций, что обеспечило общий рост запасов леща, происходивший синхронно в обоих озёрах. К концу 2000-х гг. запасы начали снижаться. Общая картина динамики запасов леща в Псковском и Чудском озёрах характеризуется высоким сходством, несмотря на значительные различия между этими частями единой водной системы и использование их биоресурсов добывающими предприятиями разных стран (России и Эстонии), хотя и координирующими промысловую деятельность, но различающимися в подходах к управлению биоресурсами (Vetemaa et al., 2008). Такое сходство, видимо, отражает общность популяции Псковско-Чудского озера, год от года разделяющейся между его различными частями в более или менее фиксированных долях.
При анализе динамики промысловых видов рыб в качестве основных факторов обычно рассматривается промысловая деятельность (Saat et al., 2010), изменение экологического режима водоёма (например, его эвтрофирование или загрязнение (Pihu, Kangur, 2001)) или общие гидроклиматические изменения в бассейне (Kangur et al., 2007). Первостепенная роль обычно отводится промыслу как постоянно действующему фактору, оказывающему не только прямое, но и опосредованное (через изменения состава сообщества и трофических отношений между видами) воздействие за запасы рыб. Среди недавних публикаций можно отметить работу Кангур с соавторами (Kangur et al., 2007), в которой на основе использования самообучающейся модели нейронной сети анализируется динамика состава уловов рыб в Чудском озере за 70-летний период (1931−2002) и делается вывод о том, что рыболовство является наиболее важным фактором данного процесса. Сходные утверждения имеются во многих других публикациях, посвящённых анализу динамики промысла рыб Псковско-Чудского озера, в том числе в работе Концевой (2009), анализирующей изменения уловов леща в послевоенный и современный периоды. По мнению Кангура (Kangur, 1996), для увеличения популяции леща необходимы защитные меры в период его нереста.
В 2000-е гг. некоторые участки нерестилищ в дельте р. Великая в Псковском озере утратили своё значение в результате заиливания, засорения и зарастания водной растительностью. Например, в 2000 г. нерест леща был неактивным и растянутым, заполняемость береговых нерестилищ была низкой, значительная часть производителей отнерестилась на глубине (Лебедева, 2006). Мечение леща показало существование его обширных нерестилищ в р. Эмайыги (Emajõgi) в 95 км выше Чудского озера и его миграцию на нагул и зимовку в Чудское озеро. Уловы леща в этой реке в 1990-е гг. увеличились в десять раз по сравнению с предыдущим десятилетием. На фоне сокращения площади нерестилищ в Чудском озере в 2000-е гг. стал расти вклад запаса речного леща в его общий улов в озере (Tambets et al., 2002).
Численность леща может быть выше, поскольку продукция его основного пищевого ресурса – зообентоса – недоиспользуется в трофической сети водоёма (Kangur, 1996). Данное утверждение подтверждается и результатами исследования балансовой трофической модели водоёма (Бобырев и др., 2013). В годы с аномально высоким обилием кормовых ресурсов наблюдается появление высокочисленных генераций леща. Так, особенно высокие биомассы бентоса в обоих озёрах зарегистрированы в 1999 и 2003 гг., что было связано с благоприятными гидрометеорологическими условиями (высокая теплообеспеченность и низкая ветровая активность) в открытый период в эти годы (Мельник, Концевая, 2005). Именно в этот период отмечена высокая численность пополнения леща в обоих озёрах. Возможно, это связано с разными спектрами питания молоди и взрослых особей.
Важная роль, приписываемая промыслу, во многом объясняется тем, что рыболовство является не только одним из немногих хорошо измеряемых видов воздействия, но и практически единственным доступным способом управления запасами. Практика такого управления не всегда приводит к ожидаемым результатам, но некоторые эффекты промысла удаётся обнаружить при анализе изменений численности рыб на интервалах, охватывающих значительные исторические периоды. Результаты проведённого анализа не дают оснований для тревожных ожиданий и свидетельствуют не столько об угрозе запасам со стороны промысла, сколько о способности последнего достигать оптимального баланса между продукционными характеристиками запаса и величиной усилия, отвечающей сложившимся приёмам и способам лова. Отчасти эта способность обеспечивается современной системой управления рыболовством, но в большей мере – стихийными процессами, присущими промысловой деятельности, ориентированной на добычу водных биоресурсов. Можно заключить, что дальнейшее увеличение интенсивности промысла (промысловой смертности) в обоих озёрах нецелесообразно, поскольку не приведёт в росту улова, а текущий уровень равновесного улова может быть достигнут при гораздо меньшей интенсивности промысла.
Из анализа модели улова на единицу пополнения следует, что современный уровень промысловой смертности леща обеспечивает добычу, близкую к максимальной. Сама по себе динамика промысловой смертности и коэффициента эксплуатации леща (рис. 5а), не повторяя в точности ход колебаний биомассы запаса, демонстрирует общую тенденцию к возрастанию. Её уровень, достигнутый к концу 2000 гг., тем не менее нельзя считать чрезвычайно высоким, несмотря на некоторое снижение среднегодовой численности популяции (в том числе её репродуктивной части) и сокращение доли старших возрастных групп (Концевая, 2009). Подобные изменения состояния промысловой популяции обратимы и являются обычной реакцией на усиление эксплуатации, в том числе вызванное появлением обильных генераций. Их нельзя считать признаком перелова. Примечательно, что, несмотря на довольно сильные флуктуации численности пополнения, его величина в большинстве случаев превышает так называемый замещающий уровень (рис. 9). Иными словами, популяция леща обладает достаточным резервом воспроизводства, позволяющим компенсировать его «провалы» в отдельные годы.
Простое сопоставление динамики общего запаса леща (рис. 2б) и межгодовых изменений величины пополнения (рис. 6) совершенно очевидно указывает на то, что факторы, определяющие урожайность пополнения, играют решающую роль в популяционной динамике этого вида. Аналогичная связь показана для леща Вислинского залива Балтийского моря (Шибаев и др., 2012). Природа этих факторов различна; они включают в себя не только внешние воздействия, но и внутренние механизмы популяционной регуляции, способные оказывать значительное влияние на темп формирования пополнения. Наличие таких механизмов подтверждается выраженной связью между величиной родительского стада леща и численностью потомства (рис. 7, 9). Рассматривая промысел через призму отношений в системе родители–потомки, нетрудно заключить, что любое усиление эксплуатации должно приводить к снижению величины родительского запаса (и, как следствие, величины нарождающихся генераций) и подавлению компенсаторных реакций, проявляющихся в форме плотностного контроля выживания и роста рыб, действующего в период формирования урожайности отдельных возрастных классов. Таким образом, давление промысла должно приводить к более отчётливому проявлению эффектов внешних воздействий. Чем сильнее это давление, тем заметнее роль внешних факторов в динамике популяции рыб.
Синхронность в появлении урожайных генераций в обоих озёрах, вероятно, обусловлена нерестовой миграцией части производителей из Чудского озера в Псковское. По всей видимости, мигрирующий чудской лещ вносит основной вклад в количество отложенной икры в Псковском озере и определяет общую динамику пополнения. Для леща характерны нерестовые миграции в пределах водоёма – нагул и зимовка в открытой части Чудского озера, а нерест в южной части Чудского озера и в Псковском (Pihu, 1996). В Псковском озере одно из основных нерестилищ леща располагается в дельте р. Великая (Лебедева, 2006). Обычно нерестовая миграция из Чудского озера в Псковское начинается в I декаде мая. Осенний ход леща из Псковского озера через Тёплое озеро в Чудское начинается в конце июля и длится до середины сентября. Однако эти сроки чаще всего определяются погодными условиями. Набор показателей, включённых нами в статистический анализ динамики пополнения леща, не даёт возможности прийти к однозначным выводам относительно главных из них. Показано наличие связи величины пополнения леща с прозрачностью, температурой воды, числом дней в году с температурой воздуха выше 20°С и среднегодовым уровнем воды. Последний фактор, по всей видимости, играет ведущую роль. Можно лишь в самых общих чертах утверждать, что положительные отклонения уровня воды в озере от основного тренда способствуют появлению более мощных возрастных классов леща. Совокупность таких факторов, как прозрачность, температура воды и число дней в году с температурой воздуха выше 20°С, в комплексе также благоприятно влияют на величину пополнения. Полученные результаты хорошо соотносятся с данными для других популяций леща, например, из Волго-Каспийского и Азово-Черноморского бассейнов (Куцын, 2016; Белоголова, Солохина, 2017; Васильченко, Пономарев, 2017). Для более детального анализа необходимы продолжительные статистические ряды и данные синхронизированных наблюдений за ходом биологических процессов, которые определяют величину пополнения, и за влияющими на эти процессы показателями озёрной среды.
По понятным причинам мы не может рассчитывать на безусловную точность полученных оценок состояния промысла леща. Результаты анализа равновесной модели и полученные на её основе оценки следует рассматривать как ориентировочные, дающие общие представления о режиме промыслового использования.
Ретроспективная модель виртуального популяционного анализа (ВПА) базируется исключительно на данных официальной статистики. Она не учитывает других видов промысла (любительского, браконьерского), масштабы которых могут быть достаточно велики. Поэтому большой интерес представляет сопоставление полученных ВПА-оценок с таковыми, основанными на иных принципах и подходах. Для такого сопоставления можно воспользоваться результатами выполненного нами ранее картографического анализа, основанного на данных траловых учётов с конца прошлого столетия, проводимых несколько раз в сезон в акватории Псковского и Чудского озёр (Васильев и др., 2018). Эти результаты интересны тем, что позволяют восстановить общую картину внутрисезонного распределения рыб в акватории водоёма.
На рис. 10 представлены карты пространственного распределения леща в Псковском озере в весенне-летние (май–июнь) и летне-осенние (август–октябрь) сезоны 2003, 2006 и 2008 гг11. Особенно характерны различия в пространственном распределении леща в весенний (май) и осенний (октябрь) периоды. В начале сезона его основные скопления регистрируются в южной и центральной части озера, осенью – на севере, ближе к проливу, соединяющему Псковское и Теплое озёра. Такая картина в целом соответствует представлениям о сроках и характере нерестовой миграции леща. Надо заметить, что картографическое отображение даёт относительную картину пространственного распределения рыб. Каждая из представленных на рис. 10 карт отображает распределение именно той численности, которая зарегистрирована на момент тралового учёта. Иными словами, штриховая кодировка участков акватории не является совместимой − для каждой даты она своя.
Рис. 10.
Распределение леща Abramis brama в Псковском озере в весенне-летний (а, б, в) и летне-осенний (г, д, е) периоды в
2003 (а, г), 2006 (б, д) и 2008 (в, е) гг. Зона плотности: ( ) – высокая, (
) – высокая, (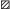 ) – средняя, без штриховки – низкая; (→) – траектория траления, (⚫) – средняя точка
траектории траления (исходная точка для интерполяции), (—) – граница области интерполяции. Масштаб: 6 км.
) – средняя, без штриховки – низкая; (→) – траектория траления, (⚫) – средняя точка
траектории траления (исходная точка для интерполяции), (—) – граница области интерполяции. Масштаб: 6 км.
Математико-картографическое моделирование позволяет провести общее суммирование показателей плотности рыб по акватории для каждой траловой съёмки и тем самым получить независимую оценку общей численности леща. Как видно из табл. 3, картографические оценки весенней и осенней численности леща могут существенно различаться. Эти различия особенно заметны при сравнении оценок, относящихся к маю и октябрю (материалы 2003 и 2006 гг.), что, всей видимости, связано с появлением в составе уловов, полученных в ходе октябрьских съёмок, возрастной группы 0+. В отличие от ВПА картографические оценки на основе траловых съёмок могут не учитывать распределение рыб на мелководных участках, что также влияет на конечные оценки численности. Можно полагать, что разница в оценках численности между траловыми съёмками и расчётами ВПА обусловлена несколькими причинами, поскольку оценки могут относиться к разным сезонам года (необходимо учитывать появление и смертность сеголеток в течение года), охватывают разные районы водоёма (могут не учитывать мелководные участки) и базируются на разных источниках информации (данные официальной статистики и учётные съёмки).
Таблица 3.
Сопоставление оценок обилия леща Abramis brama (млн экз.) Псковского озера, полученных на основе картографического метода и виртуально-популяционного анализа (ВПА)
| Год | Дата, сезон (месяц) | Метод оценки | |
|---|---|---|---|
| ВПА | картографический | ||
| 2003 | 1 января | 6.5 | |
| Весна (май) | 5.1 | ||
| Осень (октябрь) | 9.0 | ||
| 2006 | 1 января | 3.1 | |
| Весна (май) | 2.4 | ||
| Лето (август) | 3.2 | ||
| 2008 | 1 января | 1.9 | |
| Лето (июнь) | 4.7 | ||
| Осень (октябрь) | 4.4 | ||
Имеющиеся расхождения в оценках тем не менее никак не влияют на общий вывод, касающийся состояния промысла леща, поскольку настройка процедуры ADAPT–VPA проводится на основе различных индексов, некоторые из которых определяются независимо от данных промысловой статистики.
Список литературы
Белоголова Л.А., Солохина Т.А. 2017. Численность сеголеток воблы, леща и судака в западной части Северного Каспия в 2012–2016 гг. // Вестн. АГТУ. Сер. Рыб. хоз-во. № 3. С. 9–16. https://doi.org/10.24143/2073-5529-2017-3-9-16
Бивертон Р., Холт С. 1969. Динамика численности промысловых рыб. М.: Пищ. пром-сть, 248 с.
Бобырев А.Е., Криксунов Е.А., Мурзов Н.Н. и др. 2013. Состояние запасов и современные тенденции в динамике популяций промысловых рыб Псковско-Чудского водоёма // Вопр. ихтиологии. Т. 53. № 1. С. 44–56. https://doi.org/10.7868/S0042875213010037
Васильев П.В., Чистов С.В., Криксунов Е.А., Бобырев А.Е. 2018. Картографическое моделирование пространственно-временной динамики рыб Псковского озера // Матер. Междунар. конф. “ИнтерКарто/ИнтерГИС”. Т. 24. Ч. 2. С. 292–305. https://doi.org/10.24057/2414-9179-2018-2-24-292-305
Васильченко О.М., Пономарев С.В. 2017. Об особенностях естественного воспроизводства леща (Abramis brama orientalis. 1758) на нерестилищах низовий реки Волги в многоводном 2013 и маловодном 2015 годах // Вестн. АГТУ. Сер. Рыб. хоз-во. № 3. С. 17–28. https://doi.org/10.24143/2073-5529-2017-3-17-24
Данилов М.Б., Бобырев А.Е., Криксунов Е.А. и др. 2018. Динамика популяции судака Sander lucioperca Псковско-Чудского озера // Вопр. ихтиологии. Т. 58. № 4. С. 450–463. https://doi.org/10.1134/S0042875218040021
Концевая Н.Я. 2009. Состояние запасов и промысла леща в Чудском озере // Вестн. ПГУ. Сер. Естеств. и физ.-мат. науки. № 8. С. 34–38.
Криксунов Е.А., Концевая Н.Я. 1987. Анализ состояния запаса и промысла чудского сига // Вестн. МГУ. Сер. 16. Биология. № 4. С. 43–49.
Куцын Д.Н. 2016. Структура популяций и рост леща Аbramis brama (L. 1758) и плотвы Rutilus rutilus (L. 1758) Азовского моря в условиях антропогенного преобразования гидрологического режима: Автореф. дис. … канд. биол. наук. Мурманск: ММБИ КНЦ РАН, 26 с.
Лебедева О.А. 2006. Экосистема дельты реки Великой и ее влияние на Псковско-Чудское озеро // Псков. регион. журн. № 1. С. 107–121.
Мельник М.М., Концевая Н.Я. 2005. Состояние кормовой базы и рост леща в Псковско-Чудском озере // Матер. IV (XXVII) Междунар. конф. “Биологические ресурсы Белого моря и внутренних водоемов Европейского Севера” Ч. 2. Вологда: Изд-во ВГПИ. С. 5–7.
Фёрстер Э., Рёнц Б. 1981. Методы корреляционного и регрессионного анализа. М.: Финансы и статистика, 302 с.
Шибаев С.В., Голубкова Т.А., Рябчун В.А. 2012. Когортный анализ динамики численности леща (Abramis brama L.) Вислинского (Калининградского) залива Балтийского моря // Изв. КГТУ. № 24. С. 95–102.
Южакова Г.Г., Снетков М.А. 1983. Анализ промысла сига в озере Севан методами теории динамического запаса // Вопр. ихтиологии Т. 23. Вып. 5. С. 858–859.
Chen S., Watanabe S. 1989. Age dependence of natural mortality coefficient in fish population dynamics // Nippon Suisan Gakkaishi. V. 55. P. 205–208.
Gabriel W.L., Sissenwine M.P., Overholtz W.J. 1989. Analysis of spawning stock biomass per recruit: an example for Georges bank haddock // N. Amer. J. Fish. Management. V. 9. P. 383–391.
Gavaris S. 1988. An adaptive framework for the estimation of population size // CAFSAC Res. Doc. № 88/29. 12 p.
Gavaris S. 1993. Analytical estimates of reliability for the projected yield from commercial fisheries // Can. Spec. Publ. Fish. Aquat. Sci. V. 120. P. 185–191.
Kangur K., Park Y.–S., Kangur A. et al. 2007. Patterning long-term changes of fish community in large shallow Lake Peipsi // Ecol. Modell. V. 203. P. 34–44. https://doi.org/10.1016/j.ecolmodel.2006.03.039
Kangur K., Kangur A., Kangur P. et al. 2013. Long-term effects of extreme weather events and eutrophication on the fish community of Lake Peipsi (Estonia/Russia) // J. Limnol. V. 72. P. 376–387. https://doi.org/10.4081/jlimnol.2013.e30
Kangur P. 1996. On the biology of bream Abramis brama (L.) in Lake Peipsi in 1994 // Hydrobiologia. V. 338. P. 173–177.
Mäemets H., Laugaste R., Palmik K., Haldna M. 2018. Response of primary producers to water level fluctuations of Lake Peipsi // Proc. Eston. Acad. Sci. V. 67. № 3. P. 231–245. https://doi.org/10.3176/proc.2018.3.06
Mohn R.K., Cook R. 1993. Introduction to sequential population analysis // NAFO Sci. Council Stud. № 17. 110 p.
Patterson K.R., Kirkwood G.P. 1995. Comparative performance of ADAPT and Laurec–Shepherd methods for estimating fish population parameters and in stock management // ICES J. Mar. Sci. V. 52. P. 183–196.
Pihu E. 1996. Fishes, their biology and fisheries management in Lake Peipsi // Hydrobiologia. V. 338. P. 163–172.
Pihu E., Kangur A. 2001. Fishes and fisheries management // Lake Peipsi. Flora and fauna / Eds. Pihu E., Haberman J. Tartu: Sulemees Publ. P. 100–111.
Saat T., Vaino V., Afanasiev E., Koncevaya N. 2010. Effects of fisheries management on fish community of Lake Peipsi-Pihkva // Peipsi Vesikonna. Kalad Ja Kalandus / Ed. Saat T. Tartu: Keskkonnainvesteeringute Keskus. P. 58–69.
Shepherd J.G. 1982. A versatile new stock–recruitment relationship for fisheries, and the construction of sustainable yield curves // ICES J. Mar. Sci. V. 40. P. 67–75.
Tambets M., Jarvalt A., Tamberts J. 2002. Migration of bream Abramis brama L. in Lake Peipsi basin: new management approach need // EIFAC Symposium of Inland Fisheries and the Aquatic Environment. V. 22. P. 23.
Vetemaa M., Vaino V., Saat T., Kuldin S. 2008. Co-operative fisheries management of the cross border Lake Peipsi-Pihkva // Fish. Managmt. Ecol. V. 8. № 4−5. P. 283–462. https://doi.org/10.1111/j.1365-2400.2001.00259.x
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии