

Вопросы ихтиологии, 2020, T. 60, № 4, стр. 417-425
Распределение и размерно-половой состав ската Таранца Bathyraja taranetzi в тихоокеанских водах северных Курильских островов и у берегов Юго-Восточной Камчатки в летний период
В. В. Панченко 1, *, А. А. Баланов 1, А. Б. Савин 2
1 Национальный научный центр морской биологии Дальневосточного отделения РАН − ННЦМБ ДВО РАН
Владивосток, Россия
2 Тихоокеанский филиал Всероссийского научно-исследовательского института рыбного хозяйства
и океанографии – ТИНРО
Владивосток, Россия
* E-mail: vlad-panch@yandex.ru
Поступила в редакцию 19.06.2019
После доработки 11.07.2019
Принята к публикации 19.09.2019
Аннотация
Скат Таранца Bathyraja taranetzi в летний период в тихоокеанских водах северных Курильских о-вов и Юго-Восточной Камчатки обитает на глубинах от ~100 до 757 м; концентрируется у северных Курильских о-вов в диапазоне 250−360 м. В уловах отмечены особи длиной (TL) 11.3−67.5 см. На глубине <200 м обитают только крупные рыбы, стабильно присутствующие в уловах до 450-метровой изобаты. На начальных этапах развития скат Таранца в основном концентрируется в диапазоне 301−450 м. До 300 м прослеживается тенденция уменьшения средней длины рыб, далее размерный состав близок, глубже 450 м, по данным единичных поимок, длина рыб увеличивается. Взрослые особи летом обитают на внешнем шельфе и верхнем склоне при температуре 1.8−3.9°С, рыбы TL < 37 см предпочитают температуру 3.7°С и выше. Среди неполовозрелых особей соотношение полов равное. В размерной группе TL 52−58 см преобладают самцы, что обусловлено их более ранним по сравнению с самками половым созреванием. Среди рыб TL > 60 см начинают доминировать самки, что связано с достижением ими бóльших, чем самцы, размеров. В целом соотношение полов в популяции ската Таранца близко 1 : 1.
Скаты подотряда Rajoidei широко распространены в северной части Тихого океана и играют заметную роль в донных сообществах. Эти хищные рыбы могут являться как пищевыми конкурентами промысловых видов, так и их потребителями (Mito, 1974; Brodeur, Livingston, 1988; Livingston, deReynier, 1996; Orlov, 1998, 2003; Чучукало, Напазаков, 2002; Антоненко и др., 2007; Панченко и др., 2017). Во многих зарубежных странах скаты успешно осваиваются рыбопромысловым флотом (Orlov, 2005; Haas et al., 2016). Постепенно развивается и их отечественный промысел. По данным официальной статистики, за последние 5 лет (2014−2018 гг.) российский среднегодовой вылов скатов в Дальневосточном бассейне составил 4.86 тыс. т, тогда как в предыдущие годы этот показатель был гораздо меньше: в 2009−2013 гг. − 3.51 тыс. т, а в 2004−2008 гг. – лишь 1.65 тыс. т.
Скат Таранца Bathyraja taranetzi – высокобореальный тихоокеанский вид (Долганов, 1999; Долганов, Тупоногов, 1999; Шейко, Федоров, 2000; Mecklenburg et al., 2002; Парин и др., 2014). На внешнем крае шельфа и в верхней части тихоокеанского склона северных Курильских о-вов и Юго-Восточной Камчатки в настоящее время он является субдоминантным видом скатов, уступая по биомассе и численности лишь фиолетовому скату Bathyraja violacea (Savin et al., 2019). Однако распределение этого вида и его биология изучены слабо; имеющиеся в литературе сведения отрывочны и фрагментарны. Целенаправленные исследования экологии ската Таранца предпринимались лишь в прикурильских и прикамчатских водах Тихого океана (Орлов и др., 2006; Orlov et al., 2006; Orlov, Tokranov, 2010).
В настоящей работе на основе новых данных подробно рассматривается горизонтальное и вертикальное распределение, а также размерный и половой состав ската Таранца в летний период в тихоокеанских водах северных Курильских о-вов и Юго-Восточной Камчатки.
МАТЕРИАЛ И МЕТОДИКА
В основу работы положены данные траловой съёмки внешнего шельфа и верхнего отдела склона, прилегающих к океанской стороне северной части Курильской гряды и юга Камчатки (рис. 1), в которой авторы принимали непосредственное участие. Исследования проводили на научно-исследовательском судне ТИНРО-центра “Профессор Леванидов” 23.08−10.09.2018 г. и включали 86 тралений донным тралом ДТ/ТМ-27.1/24.4 м на глубинах 120−952 м. Методика работ и расчётов подробно изложена в нашей предыдущей публикации (Savin et al., 2019). При вычислениях биомассы и численности ската Таранца коэффициент уловистости принят равным 0.5. Карты распределения построены с помощью программы Surfer.
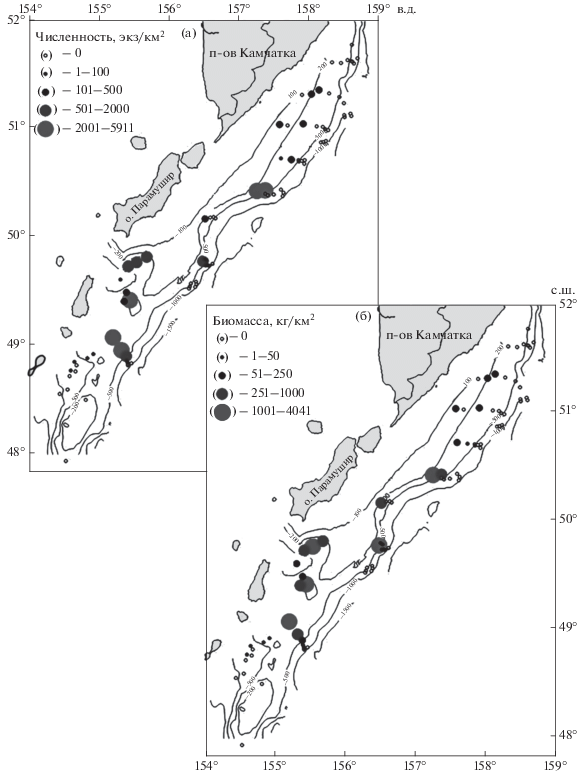
Рис. 1.
Пространственное распределение ската Таранца Bathyraja taranetzi в тихоокеанских водах северных Курильских о-вов и у Юго-Восточной Камчатки в летний период: а – численность, б – биомасса; (—) – изобаты.
Измерили полную длину (TL) 587 экз. ската Таранца, из них у 273 экз. определили массу, у 584 экз. – пол. При анализе горизонтального распределения разбивку по батиметрическим диапазонам проводили через каждые 100 м, помимо глубин >600 м, которые объединены. На глубинах до 200 м проведено пять тралений, 201−300 м – 9, 301−400 м – 12, 401−500 м – 14, 501−600 м – 11, >600 м – 35 тралений. Анализ размерного состава представлен более дробно – через 50 м. На глубинах до 150 м промерено 3 экз. ската Таранца, 151−200 м – 3 экз., 201−250 м – 108 экз., 251−300 м – 103 экз., 301−350 м – 167 экз., 351−400 м – 139 экз., 401−450 м – 57 экз., 451−500 м – 2 экз., 501−550 м – 1 экз., 551−600 м – 3 экз., >600 м – 1 экз.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Как показали наши исследования, распределение ската Таранца на акватории северных Курильских о-вов и в прилегающих водах Камчатки отличается неоднородностью (рис. 1). На самом северном разрезе он отсутствовал в уловах. Далее, с продвижением на юг, поимки начали отмечаться, однако вдоль всего обследованного восточного побережья Камчатки скоплений этот вид не образовывал. Концентрировался скат Таранца у Северных Курил, за исключением юга исследованного района, где значительных концентраций отмечено не было. Здесь хотелось бы отметить, что скат Таранца относится к мезобентальным видам, предпочитающим диапазон глубин 200−500 м (Федоров, 2000; Шейко, Федоров, 2000; Орлов и др., 2006). Между тем по ряду причин (сложный для проведения тралений рельеф дна, напряжённый график работ) в 2018 г. на юго-западе (южнее 48°55′ с.ш. и западнее 155° в.д.) в диапазоне 200−500 м выполнено лишь три траления. Возможно, именно с этим связано отсутствие здесь в зоне облова его скоплений. Орлов с соавторами (Orlov et al., 2006; Орлов, 2010; Orlov, Tokranov, 2010) при описании горизонтального распределения ската Таранца также указывают на предпочтение им центральной области рассматриваемого района, однако и на юге отмечают периодически его значительные уловы.
Известная минимальная длина при вылуплении дальневосточных скатов составляет 12.3 см (Долганов, 1998а). В пределах ареала наиболее крупный экземпляр ската Таранца (TL 77 см) зарегистрирован в восточной части Берингова моря (Ebert, 2005). В западной части Берингова моря этот показатель ниже – 72 см (Orlov, Tokranov, 2010), а в районе наших исследований – в тихоокеанских северокурильских и прилегающих прикамчатских водах – ещё меньше – лишь 70 см (Орлов и др., 2006; Orlov et al., 2006). В наших уловах скат Таранца был представлен широким размерным составом, начиная от недавно вылупившихся рыб TL 11.3 см, часть из которых ещё имела остатки желточного мешка, до 67.5 см.
В целом районы концентраций ската Таранца в численном и весовом выражениях были идентичны, однако степень доминирования каждого из уловов по биомассе и численности порой существенно различалась, что обусловлено изменением с ростом у этого вида батиметрических предпочтений. Наибольшая плотность численности (5911 экз/км2) и биомассы (4041 кг/км2) зарегистрирована на траверзе о-ва Парамушир (рис. 1), однако в первом случае глубина составила 350 м, а во втором – 251 м. Высокая удельная биомасса здесь ската Таранца на глубине 251 м была обусловлена значительным уловом особей среднего и крупного размера. Тогда как в этом районе мористее, на глубине 350 м, улов был представлен исключительно молодью TL 14−31 см с преобладанием рыб TL 15−16 см. Ни в одном из остальных тралений не было отмечено столь массового присутствия ската Таранца размером, близким к минимальному. Концентрации его отмечались и южнее на сходных глубинах в относительно равномерно охваченной тралениями зоне.
По данным разных авторов, скат Таранца в Северной Пацифике встречается на глубинах 130−1000 м (Долганов, 1999; Федоров, 2000), 105−1000 м (Долганов, Тупоногов, 1999), 81−1000 м (Шейко, Федоров, 2000; Mecklenburg et al., 2002; Парин и др., 2014), 58−1054 м (Ormseth et al., 2008). В тихоокеанских водах Камчатки и северных Курильских о-ов ранее этот вид отмечен в батиметрическом интервале 100−700 м (Орлов и др., 2006; Orlov et al., 2006), летом – 104−690 м (Orlov, Tokranov, 2010). Минимальная глубина обнаружения нами ската Таранца в 2018 г., на которой он был представлен лишь единичными экземплярами, составила 125 м. Так как на меньших глубинах было проведено лишь одно траление у сходной изобаты (120 м), уточнить верхнюю границу его обитания в летний период мы не можем. Однако, судя по мизерным уловам ската Таранца на близких к минимальным обследованным глубинам, верхняя граница его обитания близка 100-метровой изобате. Нижняя же зафиксированная нами граница обитания рассматриваемого вида составила 757 м, т.е. несколько ниже отмеченной ранее для данного района. На указанной глубине был пойман самец TL 62.2 см.
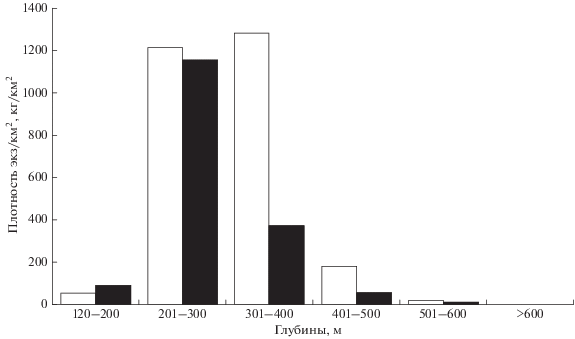
Ранее Орлов и Токранов (Orlov, Tokranov, 2010) указывали, что в рассматриваемом районе в летний период скат Таранца предпочитает глубины 340−390 м. В период наших исследований концентрации рыб наблюдались несколько мельче: наибольшая плотность численности была отмечена на глубинах от 250 до 360 м; при небольшом преобладании показателя в диапазоне 301−400 м по сравнению с 201−300 м (рис. 2). Максимальная удельная биомасса зарегистрирована в диапазоне 201−300 м, за которым с большим отрывом следовали глубины 301−400 м. Таким образом, наибольшие концентрации по биомассе располагались на несколько меньших, чем по численности, глубинах, что уже отмечалось выше при рассмотрении горизонтального распределения и связывалось с проходящими в процессе роста изменениями батиметрических предпочтений.
Рис. 2.
Средняя удельная плотность численности (◻) и биомассы (◼) ската Таранца Bathyraja taranetzi в разных диапазонах глубин в тихоокеанских водах северных Курильских о-вов и у Юго-Восточной Камчатки в летний период.
В наших сборах на относительно небольших глубинах (<200 м) в единичных поимках присутствовали только крупные особи TL 61.0−67.5 см (рис. 3). Все они оказались самками с развивающимися яйцами (диаметром > 10−20 мм) в яичниках, из чего можно заключить, что на минимальных глубинах распространения вида летом обитают только взрослые особи. Половая зрелость самцов ската Таранца наступает при TL 53.1−63.2 см, самок – 61.4−66.3 см (Долганов, 1998б; Ebert, 2005; Орлов, 2006). По нашим данным, созревание некоторых особей в прикурильских водах происходит и при меньшей длине. Отдельные самцы с полностью кальцинированными птеригоподиями имели TL 51.0 см, а наименьшая длина самки с развивающимися половыми продуктами составила 59.1 см.
Рис. 3.
Размерный состав (TL) ската Таранца Bathyraja taranetzi в уловах донного трала в разных диапазонах глубин в тихоокеанских водах северных Курильских о-вов и у Юго-Восточной Камчатки в летний период: (-) – среднее значение, (I) – пределы варьирования показателя.
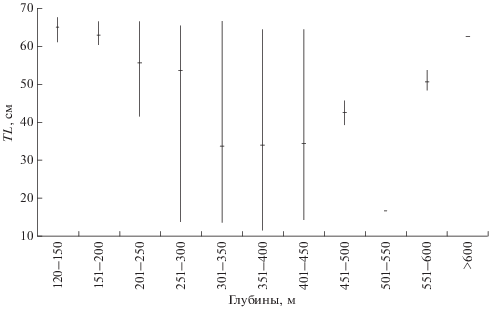
Крупные особи стабильно присутствовали в уловах до глубин ~ 450 м (рис. 3). В диапазоне 201−250 м помимо взрослых рыб стали отмечаться и неполовозрелые TL ≥ 42 см. Единичные поимки мелкой молоди появились в диапазоне 251−300 м, в массе – 301−450 м. Следовательно, в летний период на этих глубинах обитает молодь начальных этапов развития и, вероятно, проходит нерест ската Таранца. Далее, на глубинах 451−550 м, рассматриваемый вид был представлен лишь тремя особями TL 16.1−45.1 см; ниже – двумя неполовозрелыми рыбами (TL 48.7 и 49.2 см) и двумя половозрелыми самцами (TL 54.0 и 62.2 см). Таким образом, до 300-метровой изобаты прослеживается тенденция уменьшения среднего размера рыб, а глубже 450 м (по данным единичных поимок) общая направленность меняется на увели-чение размеров.
Ранее уже отмечалась тенденция уменьшения среднего размера ската Таранца с глубиной вследствие предпочтения крупными особями по сравнению с молодью меньших глубин как в целом в течение года (Орлов и др., 2006; Orlov et al., 2006), так и в отдельные сезоны, включая летний (Orlov, Tokranov, 2010). Однако представленные этими авторами данные по глубоководным диапазонам (>550 м) не соответствуют выявленной нами закономерности.
По данным литературы (Орлов и др., 2006; Orlov et al., 2006; Orlov, Tokranov, 2010), для ската Таранца, пойманного в исследуемом районе в апреле−декабре, аллометрическая зависимость массы (W, г) от длины (TL, см) имеет вид: W = 0.0029 × × TL3.1614 (R2 = 0.9777). Выведенная нами аналогичная формула оказалась близка: W = 0.0031 × × TL3.1262 (R2 = 0.9901) (рис. 4), однако масса ската Таранца оказалась несколько меньшей. Возможно, различия параметров размерно-весовой зависимости обусловлены разными размерами учтённых рыб. В наших уловах отмечены особи TL 11.3−67.5 см, а в выборке Орлова с соавторами (2006) минимальный размер ската Таранца был выше на 5 см (17 см), а максимальный – на 2 см (70 см). Отличалось и соотношение размерных групп. В нашей выборке существенную долю составляет молодь TL 15−31 см и рыбы TL 50−63 см, среди которых доминируют размерные группы 56 и 58 см (рис. 5). Средняя длина рыб в наших измерениях составила 41.8 см. В сравниваемой работе при небольшой доле молоди и рыб среднего размера относительно высокая численность приходилась на рыб TL 52−66 см (т.е. близких к созреванию и взрослых особей) с доминированием размерных групп 56−59 и 64 см. Соответственно, средний размер оказался на 10 см выше – 51.7 см (Орлов и др., 2006).
Рис. 4.
Зависимость между длиной (TL) и массой ската Таранца Bathyraja taranetzi в тихоокеанских водах северных Курильских о-вов и у Юго-Восточной Камчатки в летний период.
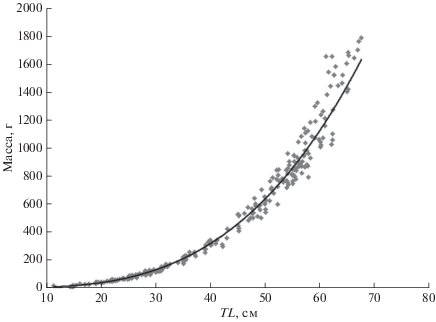
Рис. 5.
Размерный состав (TL) ската Таранца Bathyraja taranetzi в уловах донного трала (%) в тихоокеанских водах северных Курильских о-вов и у Юго-Восточной Камчатки в летний период: (◻) – самцы, (◼) – самки, (—) – оба пола.
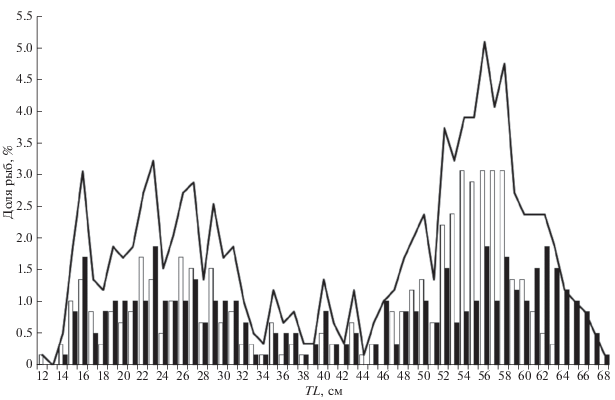
По данным Долганова (1998б), соотношение полов у эмбрионов дальневосточных скатов близко 1 : 1 и сохраняется далее. Лишь в старших размерно-возрастных группах начинают преобладать самки. Это же для большинства исследованных им видов скатов, в том числе для ската Таранца, позднее констатировал в восточноберинговоморских водах Иберт (Ebert, 2005). Равное соотношение полов характерно для ската Таранца и в западной части Берингова моря (Orlov, Tokranov, 2010). Однако в прикурильских водах для этого вида отмечено существенное преобладание самок над самцами (в 1.8 раза), причём доминирование самок проявляется не только среди крупных рыб TL 61−70 см, но и среди особей средней размерной группы 41−50 см (Орлов и др., 2006; Orlov et al., 2006). Между тем результаты наших измерений оказались близки классической схеме: лишь среди рыб TL > 60 см прослеживается увеличение доли самок, тогда как у особей мелкого и среднего размера существенные различия в соотношении полов не наблюдаются, а среди рыб TL 52−58 см (т.е. при размерах полового созревания самцов) в каждом из сантиметровых классов самцы существенно преобладают над самками (рис. 5). В целом же соотношение полов оказалось близко 1 : 1 с несущественным преобладанием самцов, доля которых составила 51.7%.
Средняя длина самцов несколько больше самок (41.9 против 41.6 см), что может объясняться неполным охватом нашей выборкой особей максимального размера. По данным Орлова с соавторами (Орлов и др., 2006; Orlov et al., 2006), самки ската Таранца в прикурильских водах достигают TL 70 см, самцы – 66 см. В нашей выборке максимальные размеры рыб обоих полов были ниже − соответственно 67.5 и 62.2 см.
Долганов (1998в) указывает, что скат Таранца в течение года встречается при температуре −0.2…+3.4°С. Позднее было выявлено, что в летний период в прикурильских водах его температурный диапазон сдвинут в сторону более высоких значений: –0.1…+5.5°С, а предпочитаемый термический фон в течение года варьирует в узких пределах 2.5−3.5°С (Orlov, Tokranov, 2010). В период проведения летней съёмки в 2018 г. на глубинах 120−952 м температура составляла 1.8−3.9°С (таблица), при этом скат Таранца встречался в пределах всего этого температурного градиента. При анализе его распределения относительно придонной температуры использованы данные с диапазона глубин 120−600 м, так как, хотя на расположенных ниже изобатах температура воды (2.7−3.7°С) находилась в пределах диапазона обитания вида, скат Таранца был представлен единственной выбивающейся из общего ряда поимкой. Несомненно, что избегание им глубин >600 м обусловлено не термическим фоном, а какими-то иными факторами.
Средняя плотность, частота встречаемости и длина ската Таранца Bathyraja taranetzi при разных значениях придонной температуры
| Показатель | Температура, °С | ||||
|---|---|---|---|---|---|
| 1.80−2.00 | 2.01−2.50 | 2.51−3.00 | 3.01−3.50 | 3.51−3.90 | |
| Плотность, экз/км2 | 44.3 | 964.3 | 1163.2 | 63.3 | 622.3 |
| Частота встречаемости, % | 33.3 | 100.0 | 100.0 | 50.0 | 35.5 |
| Длина (TL), см | 64.8 | 52.9 | 55.1 | 56.5 | 34.4 |
| Число рыб, экз. | 3 | 133 | 89 | 9 | 352 |
| Число тралений | 3 | 6 | 3 | 8 | 31 |
В период проведений исследований температура воды ниже 2°С отмечалась лишь в части тралений на глубинах <170 м. При этом термическом фоне в одном из трёх проведённых ловов было поймано лишь 3 экз. ската Таранца. Его плотность и частота встречаемости оказались здесь по сравнению с остальными температурными диапазонами минимальными (таблица). При фоновых значениях в пределах 2−3°С все проанализированные траления были для вида результативными. Высокой здесь (в особенности в пределах 2.51−3.00°С) оказалась и средняя численность ската Таранца на единицу площади. При возрастании температуры до 3.01−3.50°С этот показатель резко понизился, а частота встречаемости упала до 50%. Однако при температуре >3.5°С плотность ската Таранца по сравнению со смежным диапазоном возросла на порядок, хотя встречаемость вида продолжила падать, достигнув 35.5%. Повышение здесь плотности рыб обусловлено предпочтением молодью условий с относительно высокими значениями температуры. Мелкие особи TL < 37 см в массе отмечались лишь при температуре 3.7°С и более, в результате чего средний размер ската Таранца в верхнем диапазоне температуры (3.51−3.90°С) оказался наименьшим – 34.4 см. В отличие от молоди крупные особи встречались во всём характерном для вида температурном диапазоне. Следует лишь отметить, что самый крупный экземпляр был пойман при минимальной в период работ температуре (1.8°С), причём высоки здесь были и размеры остальных особей ската Таранца, чем и объясняется наибольшая его средняя длина при температурном фоне ниже 2°С.
ЗАКЛЮЧЕНИЕ
Как показали результаты донной траловой съёмки, проведенной у океанской стороны северной части Курильской гряды и юга Камчатки на глубинах 120−952 м, скат Таранца в летний период предпочитает акваторию северных Курильских о-вов на глубинах 250−360 м. Наименьшая глубина его обитания в это время близка 100-метровой изобате, наибольшая соответствует 757 м.
Минимальный размер вылупившихся особей составляет 11.3 см. Районы концентраций мелких и крупных рыб совпадают, однако батиметрическое распределение различается. На относительно небольших глубинах (<200 м) обитают только крупные особи, стабильно присутствующие в уловах до 450-метровой изобаты. С возрастанием глубины минимальные размеры ската Таранца снижаются. На начальных этапах развития он обитает в основном в диапазоне 301−450 м; здесь же, вероятно, располагаются и нерестилища. В связи с возрастанием доли молоди до 300-метровой изобаты прослеживается тенденция уменьшения средней длины ската Таранца. На глубинах 301−450 м размеры рыб близки, глубже (по данным единичных поимок) размеры рыб повышаются.
Наиболее эвритермными являются взрослые особи ската Таранца, обитающие летом на внешнем шельфе и верхнем склоне при температуре 1.8−3.9°С. Мелкие рыбы (TL < 37 см), являясь более теплолюбивыми, предпочитают температуру 3.7°С и выше.
Среди неполовозрелых особей соотношение полов равное. В размерной группе TL 52−58 см преобладают самцы, что обусловлено их более ранним по сравнению с самками половым созреванием. Среди рыб TL > 60 см начинают доминировать самки, что связано с достижением ими больших, чем самцы, размеров. В целом соотношение полов в популяции ската Таранца близко 1 : 1.
Список литературы
Антоненко Д.В., Пущина О.И., Соломатов С.Ф. 2007. Распределение и некоторые черты биологии щитоносного ската Bathyraja parmifera (Rajidae, Rajiformes) в северо-западной части Японского моря // Вопр. ихтиологии. Т. 47. № 3. С. 311−319.
Долганов В.Н. 1998а. Питание скатов семейства Rajidae и их роль в экосистемах дальневосточных морей // Изв. ТИНРО. Т. 124. С. 417–424.
Долганов В.Н. 1998б. Размножение скатов семейства Rajidae дальневосточных морей России // Там же. Т. 124. С. 425–428.
Долганов В.Н. 1998в. Абиотические условия среды обитания скатов семейства Rajidae дальневосточных морей России // Там же. Т. 124. С. 429–432.
Долганов В.Н. 1999. Географическое и батиметрическое распространение скатов семейства Rajidae в дальневосточных морях России и сопредельных водах // Вопр. ихтиологии. Т. 39. № 3. С. 428–430.
Долганов В.Н., Тупоногов В.Н. 1999. Определительные таблицы скатов родов Bathyraja и Rhinoraja (сем. Rajidae) дальневосточных морей России // Изв. ТИНРО. Т. 126. С. 657−664.
Орлов А.М. 2006. К обоснованию промысловой меры дальневосточных скатов (сем. Rajidae) на примере массовых западноберинговоморских видов // Тр. ВНИРО. Т. 146. С. 252−264.
Орлов А.М. 2010. Количественное распределение демерсального нектона тихоокеанских вод северных Курильских островов и юго-восточной Камчатки. М.: Изд-во ВНИРО, 336 с.
Орлов А.М., Токранов А.М., Фатыхов Р.Н. 2006. Условия обитания, относительная численность и некоторые особенности биологии массовых видов скатов прикурильских и прикамчатских вод Тихого океана // Исследования водных биологических ресурсов Камчатки и северо-западной части Тихого океана. Вып. 8. С. 38–53.
Панченко В.В., Пущина О.И., Бойко М.И., Калчугин П.В. 2017. Распределение и некоторые черты биологии ската Берга Bathyraja bergi в северо-западной части Японского моря // Вопр. ихтиологии. Т. 57. № 4. С. 415−423. https://doi.org/10.7868/S0042875217040117
Парин Н.В., Евсеенко С.А., Васильева Е.Д. 2014. Рыбы морей России: аннотированный каталог. М.: Т-во науч. изд. КМК, 733 с.
Федоров В.В. 2000. Видовой состав, распределение и глубины обитания видов рыбообразных и рыб северных Курильских островов // Промыслово-биологические исследования рыб в тихоокеанских водах Курильских о-вов и прилежащих районах Охотского и Берингова морей в 1992−1998 гг. М.: Изд-во ВНИРО. С. 7−41.
Шейко Б.А., Федоров В.В. 2000. Глава 1. Рыбообразные и рыбы. Класс Cephalaspidomorphi – Миноги. Класс Chondrichthyes – Хрящевые рыбы. Класс Holocephali – Цельноголовые. Класс Osteichthyes – Костные рыбы // Каталог позвоночных Камчатки и сопредельных морских акваторий. Петропавловск-Камчатский: Камчат. печат. двор. С. 7−69.
Чучукало В.И., Напазаков В.В. 2002. Питание и трофологический статус массовых видов скатов (Rajidae) западной части Берингова моря // Изв. ТИНРО. Т. 130. С. 422–428.
Brodeur R.D., Livingston P.A. 1988. Food habits and diet overlap of various Eastern Bering Sea fishes // US Dept. Commer. NOAA Tech. Memo. NMFS F/ NWC–127. 76 p.
Ebert D.A. 2005. Reproductive biology of skates, Bathyraja (Ishiyama), along the eastern Bering Sea continental slope // J. Fish Biol. V. 66. P. 618–649. https://doi.org/10.1111/j.1095-8649.2005.00628.x
Haas D.L., Ebert D.A., Cailliet G.M. 2016. Comparative age and growth of the Aleutian skate, Bathyraja aleutica, from the eastern Bering Sea and Gulf of Alaska // Environ. Biol. Fish. V. 99. № 11. P. 813−828. https://doi.org/10.1007/s10641-016-0518-5
Livingston P.A., deReynier Y. 1996. Groundfish food habits and predation on commercially important prey species in the Eastern Bering Sea from 1990 to 1992 // AFSC Proc. Rept. 96–04. 214 p.
Mecklenburg C.W., Mecklenburg T.A., Thorsteinson L.K. 2002. Fishes of Alaska. Bethesda, Maryland: Amer. Fish. Soc., 1037 p.
Mito K. 1974. Food relationships among benthic fish populations in the Bering Sea on the Theragra chalcogramma fishing grounds in October and November of 1972. M.S. Thesis. Hakodate: Hokkaido Univ. Grad. School, 135 p.
Orlov A.M. 1998. The diets and feeding habits of some deep-water benthic skates (Rajidae) in the Pacific waters off the northern Kuril Islands and southeastern Kamchatka // Alaska Fish. Res. Bull. V. 5. № 1. P. 1–17.
Orlov A.M. 2003. Diets, feeding habits, and trophic relations of six deep-benthic skates (Rajidae) in the western Bering Sea // Aqua. J. Ichthyol. Aquat. Biol. V. 7. № 2. P. 45–60.
Orlov A.M. 2005. Groundfish resources of the northern North Pacific continental slope: from science to sustainable fishery // Proc. 7-th North Pacific Rim Fish. Conf. Anchorage: Alaska Pacif. Univ. P. 139–150.
Orlov A.M., Tokranov A.M. 2010. Reanalysis of long-term surveys on the ecology and biology of mud skate (Rhinoraja taranetzi Dolganov, 1985) in the northwestern Pacific (1993–2002) // J. Appl. Ichthyol. V. 26. № 6. P. 861–871. https://doi.org/10.1111/j.1439-0426.2010.01512.x
Orlov A.M., Tokranov A.M., Fatykhov R.N. 2006. Common deep-benthic skates (Rajidae) of the northwestern Pacific: basic ecological and biological features // Cybium. V. 30. № 4. Suppl. P. 49−65.
Ormseth O., Matta B., Hoff J. 2008. Bering Sea and Aleutian Island skates [online]. http://www.afsc.noaa.gov/REFM/ docs/2008/BSAIskate.pdf. Version 09/2008.
Savin A.B., Balanov A.A., Panchenko V.V. 2019. The current structure of the ichthyocene on the continental shelf’s outer edge and upper slope of the Northern Kuril Islands and Southeastern Kamchatka // J. Ichthyol. V. 59. № 4. P. 499−515. https://doi.org/10.1134/ S0032945219040143
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии