

Вопросы ихтиологии, 2020, T. 60, № 4, стр. 478-487
Сравнение экспериментальных методов оценки мотивационной компоненты реореакции рыб (соотношения типов реореакции)
Д. С. Павлов 1, В. Ю. Паршина 1, В. В. Костин 1, *, Д. А. Прозоров 1
1 Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
* E-mail: povedenie@yandex.ru
Поступила в редакцию 23.01.2020
После доработки 03.02.2020
Принята к публикации 03.02.2020
Аннотация
На примере Danio rerio апробирована новая гидродинамическая установка “кольцевой лоток с лимнозоной” для определения одного из показателей мотивационной компоненты реореакции рыб – соотношения типов реореакции. В отличие от ранее использовавшихся в данной установке рыбам предоставлена возможность проявлять реореакцию четырёх, а не трёх типов. Проведён сравнительный анализ методов оценки реореакции с использованием трёх установок: “рыбоход”, “кольцевой лоток” и “кольцевой лоток с лимнозоной”. Показано, что предоставление возможности проявлять реореакцию факультативного типа не только количественно, но и качественно изменяет соотношение типов реореакции данио. Определена область применения рассмотренных методов.
Миграции рыб являются неотъемлемым звеном их жизненного цикла. Среди разнообразных механизмов миграций поведенческие играют особую роль. Именно они реализуют миграционное состояние рыб. Основным поведенческим механизмом начала, приостановки и завершения миграций является изменение мотивационной компоненты реореакции рыб (Павлов и др., 2010б; Pavlov et al., 2010; Звездин, 2016).
Реореакция (ранее реотаксис), или отношение рыб к течению, – это врождённая реакция, связанная с жизнью рыб в текучей воде. Она проявляется в выборе рыбами участка водоёма с оптимальными (для актуального поведения) скоростями течения и вектора собственного движения в нём. Проявление реореакции определяют три компоненты: ориентационная, связанная с восприятием стимула; мотивационная – выбор типа реореакции и локомоторная – реализация реореакции выбранного типа (Павлов, 1979; Pavlov еt al., 2010). К известным показателям мотивационной компоненты относятся: избираемые скорости течения (зоны скоростей течения) в реоградиенте (Павлов, 1979), реопреферендум рыб – выбор из двух каналов с течением и без него (Павлов и др., 2007), соотношение типов реореакции – соотношение всех экспериментальных оценок частот проявления реореакции разных типов (Pavlov et al., 2010).
Исследования избираемых скоростей в гидродинамическом лотке показали, что в реоградиенте наблюдаются две основные формы отношения рыб к течению (Павлов, 1979; Павлов, Скоробогатов, 2014). При первой форме рыбы избирают зону скоростей течения в диапазоне 0.30–0.95 критической скорости течения (Vk)11. Эти скорости существенно превышают пороговые для реореакции, поэтому её проявление носит обязательный характер. При второй форме отношения рыб к течению они избирают зону слабых течений в диапазоне 0–0.3 Vk. Эти скорости ниже пороговых для реореакции или близки к ним, поэтому реореакция носит факультативный характер: течения не заставляют особь постоянно проявлять реореакцию для компенсации сноса, её перемещения не обусловлены течением.
При первой форме отношения к течению у рыб может наблюдаться реореакция трёх типов: положительный тип реореакции (ПТР) – движение против течения (реотаксис), отрицательный (ОТР) – движение по течению, компенсаторный (КТР) – сохранение своего положения относительно неподвижных ориентиров (Павлов и др., 2010б; Pavlov et al., 2010; Johnston et al., 2017; Пономарева и др., 2017). Вторая форма отношения рыб к течению в данной работе рассматривается как 4-й тип реореакции – факультативный (ФТР). При данном типе реореакции вектор собственного движения особь выбирает независимо от направления течения. Таким образом, соотношение типов реореакции будет представлять собой совокупность четырёх величин − частот проявления ПТР, ОТР, КТР и ФТР.
Реореакцию этих типов мы наблюдаем (Павлов и др., 2010б; Pavlov et al., 2010; Звездин, 2016) и в естественных условиях, например, в реках: ПТР – при контранатантной миграции; ОТР – при денатантной (покатной) миграции; КТР – при сохранении места обитания (компенсации сноса) в речном потоке; ФТР – при нахождении рыб в зоне с пониженной скоростью течения или его отсутствия (в лимноусловиях: прибрежье; зона гидравлической тени за островами, косами, камнями, зарослями; зона слабых круговых течений; заливы и старицы).
Ранее показатель мотивационной компоненты реореакции оценивали по проявлению реореакции только трёх типов – ПТР, ОТР и КТР. Для получения этих оценок использовали две гидродинамические установки: “рыбоход” и “кольцевой лоток” (Pavlov et al., 2010; Пономарева и др., 2017). Поскольку в этих установках рыбы не могли проявлять ФТР, возникла необходимость модифицировать установку “кольцевой лоток”, создав в ней зону, моделирующую лимноусловия и позволяющую рыбам проявить ФТР. Эта установка названа нами “кольцевой лоток с лимнозоной”.
Цель работы – провести сравнительный анализ способов оценки соотношения типов реореакции рыб с использованием трёх экспериментальных гидродинамических установок и определить область их применения.
МАТЕРИАЛ И МЕТОДИКА
Исследования проводили в августе 2017 г. на традиционном модельном виде рыб для экспериментальных работ − данио Danio rerio стандартной длиной (SL) 20−36 мм. Рыб содержали в аэрируемых аквариумах при плотности посадки 1 экз/5 л воды. Температуру поддерживали с помощью терморегулятора на уровне 22°C. Освещённость с 10 до 19 ч составляла 450–500 лк; в остальное время суток искусственное освещение в аквариальном помещении выключали. Кормили рыб один раз в сутки после окончания опытов кормом для аквариумных рыб Tropical Fish Flakes (“Prodac”, Италия).
В экспериментах использовали две гидродинамические установки – “рыбоход” и “кольцевой лоток с лимнозоной”. Для сравнительного анализа методик привлечены результаты, полученные на данио в установке “кольцевой лоток” ранее (Пономарева и др., 2017).
Гидродинамические установки
Установка “рыбоход” представляет собой прямоточный лоток 110 × 20 см, разделённый поперечными перегородками на девять отсеков (рис. 1). Глубина воды в лотке составляла 10 см; ширина проходов между отсеками – 4 см; скорость течения в этих проходах − 22–24 см/с, или 0.53 от критической скорости течения для опытных рыб, которая составляла 43.6 см/с.
Рис. 1.
Схема гидродинамического лотка “рыбоход”: 1−9 − отсеки (5 – стартовый отсек), (– – –) – съёмные заградительные сетки, ( )– постоянные заградительные сетки, П – помпа, (→) − направление течения.
)– постоянные заградительные сетки, П – помпа, (→) − направление течения.
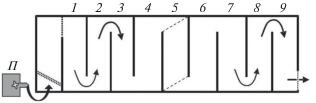
Каждую группу (8 экз.) или особь помещали в стартовый отсек (№ 5), расположенный в центре установки и закрытый с обеих сторон съёмными решётками. Время акклимации рыб к скорости потока и условиям установки составляло 20 мин, после чего открывали решётки и в течение 30 мин записывали поведение рыб видеокамерой JVC GC-XA1EU (“JVC”, Япония), установленной над “рыбоходом”. При просмотре видеозаписей фиксировали направление и время пересечения каждой особью (индивидуально даже в опытах с группой рыб) каждой границы отсека, а также распределение особей по отсекам “рыбохода” в конце опыта.
От установки “кольцевой лоток” (рис. 2а) (Пономарева и др., 2017) “кольцевой лоток с лимнозоной” (рис. 2б) отличается только наличием лимнозоны, все остальные параметры такие же. Установка изготовлена из оргстекла и представляет собой замкнутый канал шириной 9−10 см и внешним диаметром 80 см; высота стенок 15 см, внутренняя стенка на 8 см состоит из оргстекла, выше – из капронового сита, через которое стекает вода из рабочего канала к помпе. По визуальным оценкам, рыбы чаще всего двигались по центру канала, поэтому при расчётах принимали, что путь особи за один круг составляет 213 см. Лимнозона (рис. 2б) с шириной входа 30 см располагается с внутренней стороны канала. На стенки канала нанесены визуальные ориентиры в виде чёрных вертикальных полос шириной 1 см с промежутками в 8 см. Поток в рабочем коридоре создаётся с помощью помпы Atman AT-107 (Chuangxing Electrical Appliances Co., Ltd, Китай). Точка подачи воды располагается у внешней стенки канала в толще потока. Глубина воды составляла 10 см, общий объём циркулирующей воды в установке – 50 л, из них 24 л – в рабочем канале.
Рис. 2.
Схемы гидродинамических установок “кольцевой лоток”: а – без лимнозоны (по: Пономарева и др., 2017), б – с лимнозоной; Л – лимнозона, 1–8 – сектора установки (8 – стартовый сектор), (- - -) − границы секторов и лимнозоны; ост. обозначения см. на рис. 1.
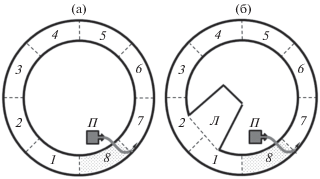
В рабочем коридоре установки наблюдается поперечный градиент скоростей течения (с максимальной скоростью у внешней стенки), зависящий от соотношения длин внешней и внутренней стенок рабочего коридора. Скорость течения у внешней стенки была 20 см/с, или 0.46 Vk для опытных рыб. Скорости течения в обеих установках измеряли при помощи измерительного комплекса, включающего в себя датчик скорости (микровертушка с диаметром лопасти 8 мм), усилитель, преобразователь сигнала и контролер на базе однокристальной ЭВМ (Павлов, Лупандин, 1994).
Перед началом опыта стартовый сектор изолировали сетками и помещали в него опытную особь. Время адаптации к условиям опыта составляло 20 мин, после чего сетки стартового сектора убирали. Далее на протяжении 30 мин фиксировали перемещение рыбы с помощью видеокамеры, расположенной над установкой. При просмотре видеозаписей фиксировали направление и время пересечения особью (всем телом) границ секторов. Перемещения особи в пределах сектора не фиксировали.
Схема экспериментов
Эксперименты каждый день начинали с группового теста (8 экз.) в “рыбоходе”, после него рыб помещали в индивидуальные сетчатые садки размером 10 × 10 см и глубиной воды 10 см, размещённые в аквариуме. Потом с каждой особью поочерёдно проводили индивидуальные тесты в “рыбоходе” и затем в “кольцевом лотке с лимнозоной”. В течение одного светового дня каждая особь проходила все три теста. Всего в “рыбоходе” проведено 36 опытов (из них четыре на группах рыб) на 32 особях, а в “кольцевом лотке с лимнозоной” – 31 опыт. Для экспериментов использовали воду из аквариумов, в которых содержали опытных рыб. Температура и освещённость в экспериментальных установках были такими же, как в аквариумах.
Методы расчёта
В нашей работе мы рассматриваем три метода расчёта частот проявления реореакции разных типов: 1) по конечному распределению рыб в “рыбоходе”; 2) по времени проявления реореакции того или иного типа в “рыбоходе”; 3) по времени проявления реореакции того или иного типа в кольцевых лотках. Метод расчёта № 2 ранее не использовали. Он необходим для того, чтобы оценить допустимость сравнения результатов, полученных при оценке соотношения типов реореакции по конечному положению рыб в “рыбоходе”, с результатами, полученными при оценке этого показателя по времени проявления реореакции разных типов в кольцевых лотках. Этот расчёт позволяет сравнивать оценки, полученные одним методом на разных установках, и определить, различаются ли результаты разных методов расчёта для одной установки.
Метод № 1. Частоту (Р) реореакции разных типов вычисляли относительно числа рыб в опыте по формулам:
Для методов расчёта № 2 и 3 потребовалось разработать объективный (не зависящий от оператора) способ оценки времени проявления КТР. Для этого введено понятие граничного времени. Граничное время (tg) – это расчётная оценка максимальной продолжительности прохода особью без остановок сектора кольцевых лотков (с лимнозоной и без неё) или отсека “рыбохода”. Величину tg определили по суммарной рабочей диаграмме частоты встречаемости времени, проведённого особями в отсеке “рыбохода” (во 2–8-м отсеках) или секторе кольцевых лотков. Граничное время определяли отдельно для движения по течению и против него. По диаграммам определяли момент времени, в котором указанная частота резко падала до принятой нами величины – в восемь раз и более. Если такого падения частоты не наблюдалось при исходной величине классового интервала (0.5–1.0 с), мы увеличивали его величину до появления искомого резкого различия. Если особь покидала отсек после истечения граничного времени, то мы считали, что она проявляла КТР в течение реального времени пребывания в отсеке (секторе) минус величина граничного времени.
Для “рыбохода” при проявлении как ПТР, так и ОТР tg = 10 с; для “кольцевого лотка” при ПТР tg = 10 с, а при ОТР tg = 2 с; для “кольцевого лотка с лимнозоной” при ПТР и ОТР − соответственно 9 и 2 с.
Метод № 2. В крайних отсеках “рыбохода” особи находились преимущественно у решёток, закрывающих выход из установки (табл. 1). Это указывает на то, что особи проявляли бы ПТР в 1-м отсеке и ОТР в 9-м отсеке, если бы решётки им не препятствовали. Поэтому мы считали, что если особь находилась в 1-м отсеке, то она проявляла ПТР, если в последнем (№ 9) – ОТР; в промежуточных отсеках (№ 2–8) рыба могла проявлять реореакцию всех трёх типов. То есть в отличие от расчёта № 1 учитывалось проявление КТР не только в центральном отсеке, но и в других, кроме крайних, отсеках. При этом считали, что особь проявляла ПТР в промежуточных отсеках, если до истечения граничного времени она покидала отсек против течения, а ОТР – по течению. Если особь покидала отсек после истечения граничного времени, то она проявляла КТР в течение реального времени пребывания в отсеке минус величина tg.
Таблица 1.
Относительное время нахождения данио Danio rerio в разных отсеках “рыбохода”, %
| Опыты | Номер отсека | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| Индивидуальные | 57.7 | 8.0 | 1.5 | 3.7 | 3.2 | 2.2 | 3.1 | 5.8 | 14.8 |
| Групповые | 77.6 | 7.1 | 5.2 | 2.7 | 4.6 | 0.9 | 0.7 | 0.4 | 0.8 |
Частоту (Р) типов реореакции вычисляли по доле времени проявления рыбами реореакции данного типа относительно длительности опыта:
Метод № 3 (кольцевые лотки). Частоту проявления реореакции разных типов, как и при расчёте методом № 2, вычисляли по доле времени её проявления относительно длительности опыта:
Обобщённые показатели реореакции. При расчёте методом № 1 индекс контранатантности (Ik) указывает, в каком направлении относительно течения и на сколько отсеков в среднем сместились рыбы в “рыбоходе”. Он изменяется от +1 (все рыбы поднялись в верхний отсек) до –1 (все рыбы опустились из стартового отсека в нижний) и вычисляется по формуле:
При расчётах с помощью методов № 2 и 3 в “рыбоходе” и кольцевых лотках индексу контранатантности соответствует перемещение рыб относительно стартового положения:
Таким образом, индексы контранатантности, полученные разными методами расчётов, отражают одно и то же – алгебраическую сумму путей, пройденных рыбами против течения и по нему. Индексы приведены к одной шкале, что и позволяет их сравнивать.
Помимо указанных выше показателей мотивационной компоненты реореакции благодаря непрерывному видеонаблюдению появилась возможность оценить дополнительные показатели поведения рыб в установке. Наиболее значимые показатели – это подвижность рыб и частота смены типов реореакции, характеризующая степень их стабильности/изменчивости.
Подвижность рыб в “рыбоходе” при методе расчёта № 1 оценивали по величине индекса подвижности (Im), который отражает, на сколько отсеков в среднем переместились особи относительно стартового независимо от направления движения, т.е. изменяется от 1 (все рыбы в крайних отсеках) до 0 (все рыбы остались в стартовом отсеке):
При расчётах методами № 2 и 3 подвижность оценивали по общему пройденному пути.
Частоту смены типов реореакции рассчитывали как частное от деления числа изменений типа реореакции на время опыта в минутах. Чем меньше этот показатель, тем в среднем стабильнее у рыбы проявление реореакции исследованных типов.
Затраты времени на получение результатов эксперимента − один из показателей для сравнения рассматриваемых в работе методик, при оценке которого учитывали только время получения исходных данных для вычисления указанных выше показателей, так как предполагается, что все последующие расчёты осуществляются средствами Excel и требуют равных затрат времени. Учитывали следующие слагаемые: продолжительность эксперимента и длительность обработки исходных данных (анализ видеозаписей и/или перенос результатов в компьютер).
В нашей работе принято, что сравниваемые соотношения типов реореакции достоверно различаются тогда, когда достоверно различаются частоты проявлений хотя бы одного из типов реореакции по критерию Стьюдента для долей (Лакин, 1973) .
РЕЗУЛЬТАТЫ
Соотношение частот проявления разных типов реореакции в трёх установках, рассчитанное разными методами, приведено в табл. 2. Сравнение способов расчёта для “рыбохода” по конечному положению особей (№ 1) и по времени проявления реореакции разных типов (№ 2) не выявило достоверных различий при групповых опытах, а при индивидуальных тестах достоверно отличается только КТР – тип реореакции с наименьшей частотой (табл. 3). Следовательно, оценки частот проявления ПТР, ОТР и КТР, полученные по конечному положению рыб в установке и по времени проявления типов реореакции, мало зависят от метода расчёта, и сравнение результатов, изложенных ниже, проведено корректно.
Таблица 2.
Частоты проявления данио Danio rerio разных типов реореакции в трёх установках, рассчитанные разными методами
| Установка | Опыт | Метод расчёта | Обозначение опыта | Частота | |||
|---|---|---|---|---|---|---|---|
| ПТР | КТР | ОТР | ФТР | ||||
| Рыбоход | Группа | № 1 | РГ1 | 0.97 | 0 | 0.03 | н.о. |
| № 2 | РГ2 | 0.86 | 0.09 | 0.05 | н.о. | ||
| Индивид | № 1 | РИ1 | 0.73 | 0 | 0.27 | н.о. | |
| № 2 | РИ2 | 0.64 | 0.17 | 0.19 | н.о. | ||
| Кольцевой лоток | № 3 | КИ3 | 0.67 | 0.23 | 0.10 | н.о. | |
| Кольцевой лоток с лимнозоной | № 3 | КлИ3 | 0.18 | 0.20 | 0.08 | 0.54 | |
Примечание. Обозначение типов реореакции здесь и в табл. 3−7: ПТР – положительный, КТР – компенсаторный, ОТР – отрицательный, ФТР – факультативный; н.о. – по данной методике невозможно определить показатель.
Таблица 3.
Типы реореакции данио Danio rerio, частоты проявления которых достоверно (р < 0.05, критерий Стьюдента для долей) различаются по результатам, полученным с использованием разных методов расчёта и установок
| Опыт | Опыт | ||||
|---|---|---|---|---|---|
| РГ1 | РГ2 | РИ1 | РИ2 | КИ3 | |
| РГ2 | – | ||||
| РИ1 | ПТР, ОТР | ОТР | |||
| РИ2 | ПТР, КТР, ОТР | ПТР | КТР | ||
| КИ3 | ПТР, КТР | – | КТР | – | |
| КлИ3 | ПТР, КТР | ПТР | ПТР, КТР, ОТР | ПТР | ПТР |
Примечание. “−“ – нет достоверных различий между одинаковыми типами реореакции, определёнными разными методиками; обозначение опытов см. в табл. 2.
При использовании “кольцевого лотка с лимнозоной”, позволяющего рыбам проявлять реореакцию всех типов, оценки их частоты достоверно отличаются от полученных в других установках (табл. 3). В “кольцевом лотке с лимнозоной” преобладал ФТР, а на втором месте был КТР; во всех других случаях преобладающим был ПТР, а на втором месте − ОТР (табл. 2). Таким образом, наличие лимнозоны принципиально изменяет частоты проявления реореакции разных типов – изменяется её преобладающий тип. Даже если не учитывать ФТР, то и в этом случае реореакция преобладающего типа различна: в “кольцевом лотке с лимнозоной” это КТР, а в других установках – ПТР.
Индекс контранатантности (табл. 4) по результатам индивидуальных тестов в “рыбоходе” оказался меньше, чем по результатам групповых, в 1.7 раза при расчёте по времени проявления реореакции разных типов и в 2.0 раза при расчёте по конечному положению особей в установке. Это указывает на то, что наличие других рыб меняет поведение особи и, как следствие, оценку показателя реореакции.
Таблица 4.
Обобщённые и дополнительные показатели реореакции данио Danio rerio, полученные при использовании разных установок и методов расчёта
| Опыт | Индекс контранатантности (Ik)/перемещение, м | Индекс подвижности (Im)/общий путь, м | Частота смены типов реореакции, мин–1 |
|---|---|---|---|
| РГ1 | 0.90/н.о. | 0.96/н.о. | н.о. |
| РИ1 | 0.44/н.о. | 0.87/н.о. | н.о. |
| РГ2 | 0.88/+8.4 | н.о./9.6 | 1.01 |
| РИ2 | 0.51/+4.8 | н.о./9.4 | 0.85 |
| КИ3 | 0.45/+79.1 | н.о./174.8 | 2.91 |
| КлИ3 | 0.03/+1.7 | н.о./53.4 | 3.97 |
Примечание. Обозначения см. в табл. 2.
В кольцевых лотках перемещение рыб и индекс контранатантности различались: при наличии лимнозоны они были на порядок меньше. Величины индекса контранатантности в “рыбоходе” и “кольцевом лотке” (без лимнозоны) отличались мало, а наличие лимнозоны привело к резкому снижению этого индекса.
Подвижность рыб в “рыбоходе” оказалась в 5.6–18.6 раза меньше, чем в кольцевых лотках, что связано, скорее всего, с длительной задержкой рыб в крайних отсеках (табл. 1). Возможно, с этим же обстоятельством связана и значительно меньшая частота смены типов реореакции в “рыбоходе” (табл. 4).
Влияние ограниченной дистанции движения рыб в “рыбоходе” на показатель мотивационной компоненты реореакции – соотношение типов реореакции − отражает поведение рыб в отсеках, прилегающих к крайним. Возврат из 2-го и 8-го отсеков в крайние наблюдался достоверно (р $ \ll $ 0.001, критерий Стьюдента для долей) чаще, чем удаление от них – 73 против 27% (табл. 5); т.е. ограничение пространства для перемещений рыб в “рыбоходе” существенно не могло влиять на частоту проявления рыбами разных типов реореакции.
Таблица 5.
Направление перемещений данио Danio rerio из отсеков установки “рыбоход”, прилегающих к крайним
| Направление перемещений из отсеков № 2 и 8 | Последовательность посещения отсеков | Число наблюдений | Доля, % | В среднем, % |
|---|---|---|---|---|
| Возврат в крайний отсек | 1−2−1 | 58 | 63.0 | 72.8 |
| 9−8−9 | 81 | 81.8 | ||
| Уход в центральные отсеки | 1−2−3 | 34 | 37.0 | 27.2 |
| 9−8−7 | 18 | 18.2 |
Способ сохранения местоположения в установке. Известно, что рыбы могут сохранять место обитания двумя способами – статическим и динамическим (Пономарева и др., 2017). В использованных установках это соответствует способу сохранения местоположения. При статическом способе сохранения местоположения у рыб превалируют КТР и ФТР (статические типы реореакции). При динамическом способе превалируют ПТР и ОТР (динамические типы реореакции) при практически одинаковой частоте проявления ПТР и ОТР, что соответствует близкому к нулю индексу контранатантности.
В “рыбоходе” и “кольцевом лотке” у рыб превалировали динамические типы реореакции (77–100% времени), при этом индекс котранатантности был в пределах 0.44−0.90, что свидетельствует о том, что в этих установках сохранение местоположения не происходило.
В “кольцевом лотке с лимнозоной” реореакцию статических типов особи проявляют в среднем 74% времени опыта. Индекс контранатантности был близок к нулю, т.е. на порядок меньше, чем у рыб в других установках (табл. 4). Соответственно и перемещение рыб в этой установке составило всего 1.7 м, что также на порядок меньше, чем в “кольцевом лотке”. Всё это свидетельствует о том, что в “кольцевом лотке с лимнозоной” рыбы проявляли статический способ сохранения местоположения.
ОБСУЖДЕНИЕ
Различия между оценками соотношения типов реореакции в “рыбоходе” по конечному распределению особей в установке и по времени проявления типов реореакции не достоверны (табл. 2, 3), т.е. метод расчёта практически не изменяет величину этого показателя. Это позволяет корректно сравнивать результаты, полученные в “рыбоходе” при оценке по конечному положению особей, с результатами, полученными в кольцевых лотках.
Сравнительный анализ результатов, полученных на разных установках на примере данио, показал, что при предоставлении этим рыбам наиболее широкого спектра возможного поведения (“кольцевой лоток с лимнозоной”) они преимущественно демонстрировали ФТР – уход из потока в лимнозону, где они в среднем проводили 54% времени опыта. А динамические типы реореакции (ПТР и ОТР) особи проявляли только 26% времени опыта. Данио в природе обитают в медленнотекущих и стоячих водах, в прибрежье рек и ручьёв. Также они населяют неглубокие пруды со стоячей водой и, в частности, рисовые чеки (Жизнь животных, 1983; Talwar, Jhingran, 1991). Кроме того, эти рыбы в течение многих десятилетий воспроизводятся в аквариумных условиях. Поведение рыб этого вида в “кольцевом лотке с лимнозоной” адекватно отражает отношение к течению в привычных им условиях обитания и содержания.
В “кольцевом лотке с лимнозоной” у данио преобладает реореакция статических типов (74%), а в “кольцевом лотке” и “рыбоходе” – динамических (соответственно 77 и 100%). У диких особей других исследованных видов рыб такое преобладание соответствует миграционному состоянию (Pavlov et al., 2010; Zvezdin et al., 2015), которое трудно ожидать у данио, судя по их образу жизни и результатам, полученным в “кольцевом лотке с лимнозоной”. Это свидетельствует о том, что невозможность проявления ФТР в установках “рыбоход” и “кольцевой лоток” изменяет поведение рыб, в результате чего существенно меняется и соотношение типов реореакции у данио. В то же время в “рыбоходе” адекватно отражается поведение диких речных рыб разных видов в естественных условиях (табл. 6). И изменение соотношения типов реореакции у них выражено значительно в меньшей степени, чем у данио. Возможно, причина рассмотренного различия данио и диких рыб заключается в том, что аквариумные рыбы по сравнению с дикими испытывают больший дискомфорт в установках с течением при отсутствии лимнозоны.
Таблица 6.
Поведение рыб в естественных условиях и преобладающий тип реореакции по оценке в “рыбоходе” (расчёт по конечному положению группы особей в установке)
| Вид и возраст, лет | Место | Поведение в природе | Преоблада-ющий тип реореакции | Источник информации |
|---|---|---|---|---|
| Густера Blicca bjorkna, 0+ | Дельта Волги, протока Быстрая | Не мигрирует | КТР | Павлов и др., 2019 |
| Вобла Rutilus rutilus caspicus, 0+ | Авандельта Волги | То же | То же | То же |
| Разновидовые скопления рыб, 0+: краснопёрка Scardinius erythrophthalmus, густера, уклея Alburnus alburnus, вобла | Дельта Волги, протока Быстрая: | » | ||
| – день | Контранатантная миграция | ПТР | ||
| – ночь | Не мигрируют | КТР | ||
| Плотва Rutilus rutilus: | Р. Ильдь | Pavlov et al., 2010; Звездин, 2016 |
||
| – С2 | Покатная миграция | ОТР | ||
| – D1 | Не мигрирует | КТР | ||
| – D2–F | Контранатантная миграция | ПТР | ||
| – мальки | Не мигрируют | КТР | ||
| Нерка Oncorhynchus nerka | Притоки оз. Курильское | Покатная миграция | ОТР | Павлов и др., 2013 |
| Исток из оз. Курильское | Контранатантная миграция | ПТР | То же | |
| Атлантический лосось Salmo salar, 0+ (расселяющийся из нерестовых гнёзд) | Р. Варзуга | Скат и расселение в прибрежье | ОТР | Павлов и др., 2010б |
| Р. Ареньга | Контранатантная миграция | ПТР | То же |
Результаты экспериментов свидетельствуют, что для получения адекватной оценки мотивационной компоненты реореакции мигрирующих и немигрирующих рыб требуется рассматривать все типы реореакции. Без учёта ФТР соотношение типов реореакции у некоторых немигрирующих рыб может кардинально изменяться, вплоть до преобладания ПТР, что указывает на их предрасположенность (мотивацию) к контранатантной миграции. Кроме того, в природе немигрирующие рыбы проявляют КТР при питании, когда держатся на потоке на границе прибрежья и русла реки, а ФТР при отдыхе в заливах реки, в которых скорости течение малы и непостоянны по величине и направлению. В течение светлого времени суток особи неоднократно посещают оба указанных биотопа (Павлов и др., 2007). Количественно такое поведение можно о изучать только экспериментально при использовании “кольцевого лотка с лимнозоной”. В целом наиболее точно соотношение типов реореакции отражается при обеспечении рыбам возможности проявления реореакции всех типов, т.е. в “кольцевом лотке с лимнозоной”.
Преимущества и недостатки использования рассмотренных установок и методов расчёта по затратам времени на получение одинакового объёма данных и качеству результатов приведены в табл. 7. Наиболее точные и полные оценки типов реореакции можно получить при использовании “кольцевого лотка с лимнозоной”. Однако этот способ требует в 16 раз больше времени, чем опыты в “рыбоходе”, при оценке по конечному положению особей в опытах с группами рыб.
Таблица 7.
Сравнительный анализ методов определения показателей реореакции Danio rerio
| Показатель | Рыбоход | Кольцевой лоток | Кольцевой лоток с лимнозоной | |
|---|---|---|---|---|
| метод № 1 | метод № 2 | метод № 3 | ||
| Время получения результатов для 100 особей, ч: | ||||
| – опыты с группами рыб по10 экз. | ~10 | ~115 | – | – |
| – опыты с индивидами | ~100 | ~160 | ~160 | ~160 |
| Частота динамических типов реореакции | Завышена | Реальная | ||
| Частота статических типов реореакции | Сильно занижена, так как у рыб нет возможности проявлять факультативный тип реореакции | Реальная | ||
| Индекс контранатантности | Завышен | Реальный | ||
| Показатель подвижности | Резко завышен | Занижен | Завышен | Реальный |
| Частота смены типов реореакции | н.о. | Сильно занижена | Занижена | Реальная |
| Способ сохранения местоположения | Не сохраняют | Статический | ||
Пока методики с использованием кольцевых лотков не верифицированы на диких рыбах. Методика определения соотношения типов реореакции в “рыбоходе” с оценкой по конечному положению особей, хотя и вносит систематическую ошибку из-за отсутствия условий для проявления ФТР, адекватно отражает поведение диких речных рыб в естественных условиях. Эту ошибку необходимо учитывать при оценке миграционного или резидентного состояния одной группы рыб. В то же время надо отметить, что систематическая ошибка не влияет на результаты сравнительной оценки двух или более групп, например: контроль–опыт, голодные–сытые, мигрирующие–немигрирующие.
Проведённый сравнительный анализ показал, что нет оснований для использования как “кольцевого лотка” (без лимнозоны), так и метода расчёта № 2 по времени проявления реореакции разных типов в “рыбоходе”, так как эти методики уступают методике с использованием “кольцевого лотка с лимнозоной” по качеству результатов, а методике с использованием “рыбохода” с расчётом по конечному положению особей – по затратам времени на их получение. Таким образом, для дальнейшего применения можно рекомендовать две методики определения рассмотренного показателя мотивационной компоненты реореакции рыб: методику с использованием “рыбохода” при определении частот ПТР, ОТР и КТР по конечному положению особей (метод расчёта № 1) и новую методику с использованием установки “кольцевой лоток с лимнозоной”.
Первая методика с использованием “рыбохода” прошла верификацию на диких рыбах, она достаточно адекватно отражает их реореакцию и её изменение в естественных условиях. С её помощью успешно проведено более 10 исследований, результаты которых опубликованы (Павлов и др., 2010а, 2010б, 2013, 2019; Pavlov et al., 2010; Звездин, 2016). Область применения “рыбохода” – это экспресс-оценка частот реореакции трёх типов – ПТР, ОТР и КТР.
Вторая методика с использованием “кольцевого лотка с лимнозоной”, предоставляющая рыбам возможность проявить ФТР, расширяет экспериментальный инструментарий и делает, судя по первым данным, получаемые оценки более полными, точными и многосторонними. Использование этой методики даст возможность не только проверить ряд сформулированных ранее положений, но и продвинуться вперёд в изучении реореакции речных рыб. Эта установка нуждается в верификации на диких рыбах. Если верификация будет успешной, то областью применения этой методики станет определение частот реореакции четырёх типов, что даст более точное представление о мотивационной компоненте реореакции.
ВЫВОДЫ
1. Апробирована новая установка “кольцевой лоток с лимнозоной” для определения частот проявления рыбами реореакции разных типов (ПТР, ОТР, КТР, ФТР). На примере данио показано: возможность проявлять ФТР не только количественно, но и качественно изменяет реореакцию рыб; преобладающим становится ФТР, а не ПТР, как в установках без лимнозоны. В этой установке наиболее точно определяется соотношение типов реореакции рыб. Однако методика с использованием этой установки требует верификации на диких рыбах.
2. “Рыбоход” с расчётом по конечному распределению особей в установке вполне пригоден для экспресс-оценки соотношения трёх типов реореакции (ПТР, ОТР, КТР), а также для сравнительной оценки этого соотношения у разных групп рыб.
3. Не следует использовать “кольцевой лоток” (без лимнозоны) и метод расчёта по времени проявления реореакции разных типов в “рыбоходе”, поскольку при одинаковых затратах времени они дают менее точные результаты, чем “кольцевой лоток с лимнозоной”.
Список литературы
Жизнь животных. 1983. Т. 4. Рыбы / Под ред. Расса Т.С. М.: Просвещение, 575 с.
Звездин А.О. 2016. Реореакция ранней молоди нерки Oncorhynchus nerka (Walb.) в период расселения с нерестилищ: Автореф. дис. … канд. биол. наук. М.: ИПЭЭ РАН, 28 с.
Лакин Г.Ф. 1973. Биометрия. М.: Высш. шк., 352 с.
Павлов Д.С. 1979. Биологические основы управления поведением рыб в потоке воды. М.: Наука, 319 с.
Павлов Д.С., Лупандин А.И. 1994. Реакции рыб на потоки с различной степенью турбулентности // Докл. АН. Т. 339. № 3. С 427−430.
Павлов Д.С., Скоробогатов М.А. 2014. Миграции рыб в зарегулированных реках. М.: Т-во науч. изд. КМК, 413 с.
Павлов Д.С., Лупандин А.И., Костин В.В. 2007. Механизмы покатной миграции молоди речных рыб. М.: Наука, 213 с.
Павлов Д.С., Костин В.В., Пономарева В.Ю. 2010а. Поведенческая дифференциация сеголеток черноморской кумжи Salmo trutta labrax: реореакция в год, предшествующий смолтификации // Вопр. ихтиологии. Т. 50. № 2. С. 251−261.
Павлов Д.С., Пономарева В.Ю., Веселов А.Е., Костин В.В. 2010б. Реореакция как один из механизмов формирования фенотипических группировок сеголеток атлантического лосося Salmo salar // Там же. Т. 50. № 4. С. 548–553.
Павлов Д.С., Звездин А.О., Костин В.В. 2013. Тип реореакции ранней молоди нерки Oncorhynchus nerka в реке Озерная и озере Курильское // Там же. Т. 53. № 1. С. 87–95. https://doi.org/10.7868/s0042875213010104
Павлов Д.С., Костин В.В., Звездин А.О. и др. 2019. Реореакция молоди некоторых карповых рыб (Cyprinidae) в период осенней контранатантной миграции // Там же. Т. 59. № 6. С. 716−723. https://doi.org/10.1134/S0042875219060122
Пономарева В.Ю., Павлов Д.С., Костин В.В. 2017. Разработка и апробирование методики исследования соотношения типов реореакции рыб в кольцевом гидродинамическом лотке // Биология внутр. вод. № 1. С. 100–108. https://doi.org/10.7868/S0320965217010156
Johnston M.E., Kelly J.T., Lindvall M.E. et al. 2017. Experimental evaluation of the use of vision and barbels as references for rheotaxis in green sturgeon // J. Exp. Mar. Biol. Ecol. V. 496. P. 9–15. https://doi.org/10.1016/j.jembe.2017.04.002
Pavlov D.S., Kostin V.V., Zvezdin A.O., Ponomareva V.Yu. 2010. On methods of determination of the rheoreaction type in fish // J. Ichthyol. V. 50. № 11. P. 977–984. https://doi.org/10.1134/s0032945210110020
Talwar P.K., Jhingran A.G. 1991. Inland fishes of India and adjacent countries. V. 1–2. N. Delhi, Bombay, Calcutta: Oxford; IBH Publ., 1158 p. https://doi.org/10.1007/BF0004326
Zvezdin A.O., Pavlov D.S., Kostin V.V. 2015. On the mechanism of orientation and navigation of sockeye salmon underyearlings (Oncorhynchus nerka Walb.) during feeding migration in the inlet–lake–outlet system // Inland Water Biol. V. 8. №. 3. P. 287–295. https://doi.org/10.1134/s195082915030189
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии