

Вопросы ихтиологии, 2020, T. 60, № 4, стр. 400-406
Новые данные по морфологии и распространению двух редких видов глубоководных удильщиков из семейств Linophrynidae и Himantolophidae
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН – ИПЭЭ РАН
Москва, Россия
2 Институт океанологии им. П.П. Ширшова РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 19.08.2019
После доработки 06.09.2019
Принята к публикации 18.09.2019
Аннотация
Приведены новые сведения о морфологии и распространении двух недостаточно изученных видов цератиоидных удильщиков – Linophryne pennibarbata и Himantolophus appelii. Вид L. pennibarbata впервые встречен в Индийском океане. Новый экземпляр отличается от рыб типовой серии по деталям строения придатков эски и задней ветви гиоидного усика. Обсуждена онтогенетическая изменчивость в строении эски L. рennibarbata; показано, что длина дистального придатка эски и его боковых ответвлений, а также число отростков задней (непарной) ветви гиоидного усика у этого вида подвержены значительной индивидуальной изменчивости, не связанной с ростом. Выявлено, что отмечавшиеся ранее различия в размерах и пигментации дистальных придатков эски у H. appelii не могут быть связаны с онтогенетической изменчивостью, как это предполагалось, а свидетельствуют о сборной природе этого вида. Приводятся основания для закрепления названия H. appelii за формой с короткими, интенсивно пигментированными (кроме самых кончиков) дистальными придатками.
Цератиоидные удильщики (подотряд Ceratioidei) представляют наиболее разнообразную в таксономическом отношении группу батипелагических рыб, населяющих глубины свыше 1000 м. Этот подотряд включает 11 семейств, 35 родов и около 170 видов (Pietsch, 2009; Fricke et al., 2019), надежная видовая диагностика которых в большинстве случаев возможна только по самкам, прошедшим метаморфоз, и только по одной морфологической структуре – эске, представляющей собой тканевое дистальное утолщение иллиция (модифицированной первой спинной колючки), обычно заключающее внутри светящийся орган и несущее на своей поверхности разнообразные придатки. Иллиций отсутствует у монотипического семейства Neoceratiidae, а светящийся орган внутри эски – у представителей семейства Caulophrynidae и у рода Rhynchactis из Gigantactinidae. В некоторых семействах (Caulophrynidae, Melanocetidae) эска имеет довольно простое и однообразное строение, и в видовой диагностике используются другие морфологические структуры; наконец, у видов рода Linophryne имеется своеобразный гиоидный усик, строение которого также видоспецифично (Pietsch, 2009). Во всех остальных родах Ceratioidei видовая диагностика основывается исключительно на особенностях строения придатков эски, специфичных для каждого вида и в то же время подверженных онтогенетической и индивидуальной изменчивости. Хотя с момента описания первого представителя этой группы (Reinhardt, 1837 – цит. по: Pietsch, 2009) накоплен обширный материал, ряд видов остаются известными только по голотипам или единичным экземплярам, что осложняет оценку вариабельности диагностически значимых структур. Как и другие бати- и абиссопелагические рыбы, преимущественно связанные с глубинами 1500–2000 м и более, цератиоидные удильщики характеризуются очень обширными видовыми ареалами и крайне слабой степенью эндемизма, однако многие виды известны по единичным находкам из двух или трёх океанов, из-за чего их ареалы продолжают оставаться окончательно не выясненными.
В настоящей работе сообщается о новых экземплярах двух редких видов удильщиков из семейств Linophrynidae и Himantolophidae. Некоторые детали их морфологии позволяют существенно уточнить индивидуальную и онтогенетическую изменчивость диагностически значимых структур; кроме того, рассматриваемый вид линофрины впервые указывается для Индийского океана.
МАТЕРИАЛ И МЕТОДИКА
Этикеточные данные приведены при видовых описаниях. Изученный материал хранится в коллекциях Зоологического музея Московского государственного университета (ЗММУ) и Института океанологии РАН, Москва (ИО РАН). Методика изучения и терминология соответствуют общепринятым (Bertelsen, 1980; Bertelsen, Krefft, 1988; Pietsch, 2009). Показатели счётных значений одних и тех же признаков на разных сторонах тела одной рыбы разделены знаком “/”. В тексте использованы следующие сокращения: D, A, P, С – соответственно спинной, анальный, грудные и хвостовой плавники, э/с – экспедиционное судно.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
(рис. 1–3)
Рис. 1.
Linophryne pennibarbata ЗММУ № 21690, SL 55 мм: а – общий вид; б – эска (масштаб – 3.5 мм): db – концевые ответвления дистального придатка, lb1–4 – боковые ответвления дистального придатка, pa – задний придаток.
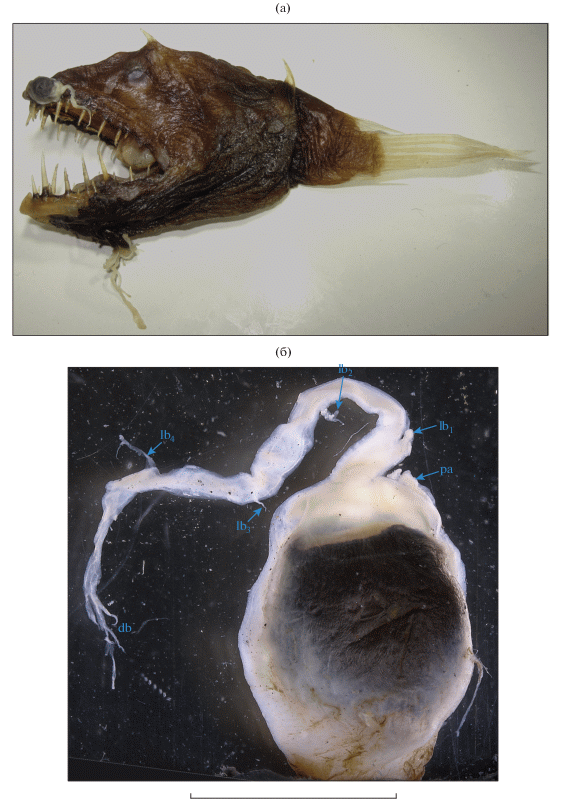
Рис. 2.
Linophryne pennibarbata ЗММУ № 21690, SL 55 мм, гиоидный усик: а, б – общий вид (а – вид спереди и сверху, б – вид сзади); в – концевое разветвление передней ветви; г – то же задней ветви. Масштаб: а, б – 3, в, г – 1 мм.
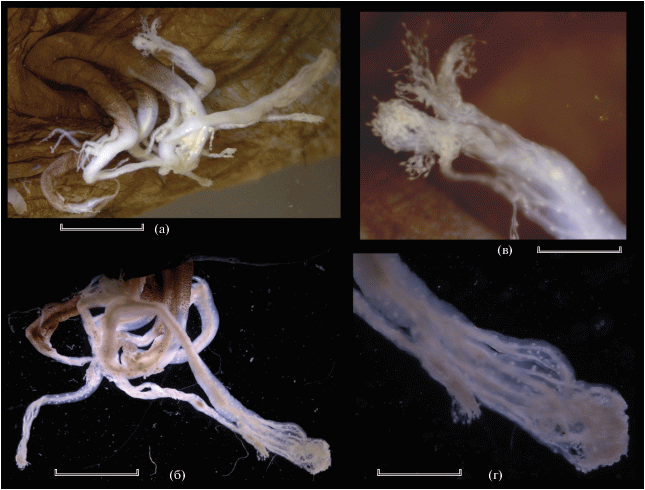
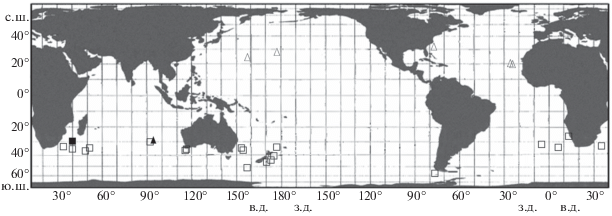
Рис. 3.
Известные места ловов экземпляров Linophryne pennibarbata (▲, △) и Himantolophus appelii (◼, ◻), описанных в статье (▲, ◼) и по литературным данным (△, ◻). В некоторых случаях один значок может отображать более одной поимки.
Материал. ЗММУ № 21690 (из ИО РАН), 1 экз. SL 55 мм (рис. 1а), э/с “Фиолент”, рейс 7(9), трал 38, 03.08.1977 г., 31°09′ ю.ш., 94°49′ в.д., глубина 1050 м.
Описание. D 3, A 3, P 15/16, C 4 + 4 (3 + 3 ветвистых). Зубов на praemaxillare 13/14, dentale 12/13, vomer 1 (центральный), palatinum 0, фарингобранхиальных зубов 5/5. Зубы на praemaxillare расположены в один ряд, на dentale – в два.
Некоторые измерения, в % SL: длина рыла 21.8, горизонтальный диаметр глаза 6.4, ширина головы на уровне sphenotica 27.3, длина сфенотикального шипа 7.3, длина верхней челюсти 40.0, длина наибольшего зуба praemaxillare и dentale соответственно 8.2 и 12.7, длина иллиция с птеригиофором и эской 43.6, длина эски и её дистального придатка соответственно 10.0 и 18.2, длина Р и С соответственно 27.3 и 41.8.
Эска с длинным дистальным придатком, несущим четыре пары очень коротких супротивных боковых ответвлений и пучок длинных филаментов на вершине, и очень коротким утолщённым задним придатком; на заднем крае эски имеется тонкий одиночный филамент (рис. 1б). Гиоидный усик состоит из трёх ветвей, правая и левая передние ветви которых разделяются на две у самого основания, а задняя ветвь не подразделена (рис. 2а, 2б). От этих ветвей отходят немногие боковые веточки второго порядка, в их толще заключены многочисленные светящиеся тельца; дистальные концы ветвей мелко ветвятся и густо покрыты светящимися тельцами (рис. 2в, 2г). Ветви гиоидного усика пигментированы лишь в их основной части (немногим далее уровня разветвления передних ветвей, где сплошная пигментация распадается на разрозненный меланофорный крап).
Замечания. Изученный экземпляр отнесён к L. pennibarbata на основании наличия у него длинного дистального придатка эски, несущего боковые и дистальные филаменты, двух парных (передних) и единственной (задней) непарной ветви гиоидного усика (Bertelsen, 1980; Pietsch, 2009). До сих пор данный вид был известен только по 11 самкам SL 35–52 мм на разных стадиях метаморфоза из западной и восточной частей Северной Атлантики и из центральной части Северной Пацифики (Pietsch, 2009). В Индийском океане этот вид отмечается впервые (рис. 3). Он несколько крупнее ранее известных экземпляров. Питч (Pietsch, 2009) указывает 1 экз. SL 52 мм, но особенности строения его эски остались неописанными. Согласно Бертельсену (Bertelsen, 1980), данный вид при завершённом метаморфозе (SL 40–47 мм) характеризуется очень длинным дистальным придатком эски (45–60% SL), несущим до пяти пар довольно длинных (около 3–6 раз в длине придатка, судя по рис. 6 А из цитируемой работы) супротивных боковых ответвлений и на вершине неветвящимся, либо несущим пару филаментов; два боковых ответвления в основании придатка всегда длиннее и толще остальных (Bertelsen, 1980. Fig. 7 ). В онтогенезе дистальный придаток сначала имеет вид трёх бугорков (при SL 35–36.5 мм), затем становится трёхветвистым (два более коротких (около половины–трети длины основного ствола), но толстых боковых ответвления) (SL 36.5–43 мм), его длина составляет 5–13% SL (Bertelsen, 1980. Figs. 6E, 7 ). Таким образом, метаморфоз у L. pennibarbata довольно растянут и не совсем коррелирует с SL, поскольку один из паратипов при SL 40 мм имеет дефинитивное строение эски, а другой при SL 43 мм проявляет лишь начальные стадии дифференцировки дистального придатка эски (Bertelsen, 1980). Изученный экземпляр по строению дистального придатка имеет ряд бросающихся в глаза отличий: его боковые ответвления очень короткие (менее 10 раз в длине придатка), хотя первая пара, как и у рыб типовой серии, заметно толще последующих; на вершине он разделяется на несколько тонких филаментов, заметно более длинных, чем боковые ответвления, а длина самого придатка гораздо меньше таковой у ранее известных рыб с дефинитивным строением эски (18.2% SL против 45–60%). Хотя короткий дистальный придаток и слабое развитие боковых ответвлений можно было бы посчитать ювенильной особенностью, я полагаю, что данный экземпляр всё же завершил метаморфоз, так как первая пара боковых ответвлений у него сильно укорочена по сравнению с основным стволом. В таком случае приходится полагать, что длина боковых ответвлений и самого дистального придатка, а также степень ветвления его вершины у L. pennibarbata подвержена значительной индивидуальной изменчивости, не коррелирующей с ростом.
По строению гиоидного усика описываемый экземпляр хорошо соответствует первоописанию, за исключением гораздо более слабого развития ответвлений второго порядка на задней ветви усика (рис. 2б; Bertelsen, 1980. Figs. 6C, 6D ), что, на мой взгляд, следует трактовать как индивидуальную вариацию.
(рис. 3–5)

Рис. 5.
Himantolophus appelii SL 215 мм, ИО РАН № 3597, иллиций и эска с придатками: а – вид сбоку, б – эска дорсолатерально; da, pa, pla – соответственно дистальные, задние и заднебоковые придатки эски; el – лопасти эски. Масштаб: а – 10, б – 5 мм.

Материал. ИО РАН № 3597, 1 экз. SL 215 мм (рис. 4), э/с ?, 17.04.1971 г., 30°18′ ю.ш., 40°34′ в.д., сборщик Юхов.
Описание. D 6, A 4, P 16/17, C 5 + 5 (3 + 3 ветвистых). Шипов на боках тела 75, на основании Р 6/6. Зубов на praemaxillare 30/30, dentale 48/53, в присимфизных частях этих костей зубы расположены соответственно в три и четыре сильно неправильных ряда, далее назад переходящих в два и затем в один ряд. Папиллы на вершине нижней челюсти округлые, плоские, едва выступающие.
Некоторые измерения, в % SL: длина головы 58.1, предорсальное и преанальное расстоянияе соответственно 74.4 и 86.1, длина оснований D и А соответственно 18.6 и 7.9, длина Р и С соответственно 26.1 и 14.0, диаметр основания наибольшего кожного шипа 9.3, горизонтальный диаметр глаза 1.9, длина рыла 14.0, ширина межглазничного промежутка 21.4, длина иллиция с эской 32.6, длина дистального, заднего и наибольшего заднебокового придатка эски соответственно 3.3, 27.9 и 16.3, продольный диаметр и высота (вместе с лопастями) эски соответственно 6.1 и 4.7.
Иллиций и эска с придатками изображены на рис. 5. Задний край задних придатков эски с 2/3 ответвлениями (расположенными на расстоянии соответственно 10/10, 22/22, 30/– мм от основания придатка при длине последнего в 60 мм), длина наименьшего и наибольшего ответвления составляет соответственно 15 и 22 мм (7.0 и 10.2% SL). Заднебоковых придатков две пары, они расположены по заднему краю эски сразу под её задними придатками; на иллиции кожных придатков нет. Вершины заднебоковых и задних придатков и их ответвлений ветвятся, светлые (рис. 5а). Дистальные придатки каждый с двумя короткими (около 4.7 раза в длине придатка) супротивными (передним и задним) отростками примерно на середине их длины; вершины этих придатков и их отростков простые, закруглённые. Дистальные придатки интенсивно-чёрные, но их кончики и кончики их отростков ярко-белые. Дорсальные лопасти эски длинные, плоские, округлые, их передние и задние отделы отделены очень слабо (рис. 5б). Вершина эски (лопасти и кожа вокруг поры фотофора) ярко-белые, без какого-либо пигмента.
Замечания. Вид распространен циркумглобально в Южном полушарии между 27° и 56° ю.ш. Изученный экземпляр полностью соответствует описанным в литературе признакам вида (Clarke, 1878; Bertelsen, Pietsch, 1983; Bertelsen, Krefft, 1988; Anderson, Leslie, 2001; Pietsch, 2009) и пойман в пределах известного для него ареала (рис. 3). Вместе с тем наблюдаемые у него особенности строения позволяют существенно уточнить характер онтогенетической изменчивости придатков эски. По имевшимся представлениям (Bertelsen, Krefft, 1988), дистальные придатки эски при SL 20–167 мм короткие и не проявляют тенденцию к увеличению с ростом, составляя 1.3–5.1% SL, тогда как у рыб SL 192–270 мм они гораздо длиннее и составляют около 10–20% SL. Однако изученный экземпляр не подтверждает тенденции к резкому увеличению длины дистальных придатков у очень крупных рыб: при SL 215 мм они составляют всего 3.3% SL. На мой взгляд, связывать изменчивость в строении дистальных придатков с ростом нельзя. По-видимому, правильным является предположение Питча (Pietsch, 2009) о возможном смешении под названием H. appelii двух видов, различающихся длиной и степенью аллометрии дистальных придатков эски. Необходимо отметить, что помимо гораздо большей длины и толщины дистальных придатков, сопоставимых с таковыми задних придатков эски, вторая форма отличается также полным отсутствием их пигментации (Bertelsen, Krefft, 1988. Fig. 16C ) в отличие от интенсивно пигментированных дистальных придатков (но с белыми вершинами всех отростков) у изученного экземпляра и у рыб SL 20–167 мм, исследованных Бертельсеном и Креффтом.
Голотип H. appelii (SL 287 мм) не сохранился (Pietsch, 2009; Fricke et al., 2019), а в первоописании (Clarke, 1878) дистальные придатки не были описаны и изображены. Однако вид был описан по выброшенному на берег экземпляру, вероятно, помятому и подсохшему (на что указывает, например, его изображение в профиль: Clarke, 1878. Pl. VI). В центре дорсальной поверхности эски Кларк изображает крупный чёрный диск – с большой долей вероятности можно предположить, что это остатки маленьких примятых и подсохших дистальных придатков, так как вершина самой эски у H. appelii совершенно не пигментирована. Без сомнения, для H. appelii требуется выделение неотипа (из-за отсутствия материала из новозеландских вод в настоящем сообщении я воздерживаюсь от этого), я считаю также, что это название должно быть закреплено за формой с короткими и пигментированными на всех стадиях онтогенеза дистальными придатками эски.
Список литературы
Anderson M.E., Leslie R.W. 2001. Review of the deep-sea anglerfishes (Lophiiformes: Ceratioidei) of southern Africa // Ichthyol. Bull. J.L.B. Smith Inst. Ichthyol. № 70. 32 p.
Bertelsen E. 1980. Notes on Linophrynidae V: A revision of the deepsea anglerfishes of the Linophryne arborifera-group (Pisces, Ceratioidei) // Steenstrupia. V. 6. № 6. P. 29–70.
Bertelsen E., Krefft G. 1988. The ceratioid family Himantolophidae (Pisces, Lophiiformes) // Ibid. V 14. № 2. P. 9–89.
Bertelsen E., Pietsch T.W. 1983. The ceratioid anglerfishes of Australia // Rec. Aust. Mus. V. 35. P. 77–99.
Clarke F.E. 1878. On two new fishes // Trans. Proc. N. Z. Inst. V. 10 (1877). P. 243–246.
Fricke R., Eschmeyer W.N., van der Laan R. (Eds.) 2019. Catalog of Fishes: Genera, Species, References. (http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Version 08/2019)
Pietsch T.W. 2009. Oceanic anglerfishes: extraordinary diversity in the deep-sea. Berkeley; Los Angeles: University of California Press, xii + 557 p., 310 figs.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии