

Вопросы ихтиологии, 2020, T. 60, № 5, стр. 545-551
Особенности пространственного распределения разных экологических групп ихтиопланктона в северной части Центрально-Восточной Атлантики
А. Г. Архипов 1, *, Р. А. Пак 1
1 Атлантический филиал Всероссийского научно-исследовательского института рыбного хозяйства
и океанографии − АтлантНИРО
Калининград, Россия
* E-mail: arkhipov@atlantniro.ru
Поступила в редакцию 15.08.2019
После доработки 14.10.2019
Принята к публикации 30.10.2019
Аннотация
С использованием многомерных методов анализа данных рассмотрено распределение рыб ранних стадий развития в слое воды 0−100 м (0−дно) в северной части Центрально-Восточной Атлантики (северная и южная части Марокко и Мавритания). Для анализа распределения разных экологических групп рыб на ранних стадиях развития можно использовать данные по численности икринок и личинок рыб, для которых наблюдаются определённые закономерности в локализации. В рассматриваемых районах часто выделяются две самостоятельные экологические группы: мезопелагическая (удалённая от побережья) – представители семейств Myctophidae, Gonostomatidae и неритическая (прибрежная) – Clupeidae, Sparidae. Использованная методика анализа материала позволяет объективно оценить сходство и различие видовой структуры и особенности распределения массовых рыб на ранних стадиях онтогенеза.
Воды, прилегающие к северной части Центрально-Восточной Атлантики (ЦВА), в поверхностном слое (до 150−200 м) формируются, как правило, субтропической и тропической водными массами. Атлантическое побережье Марокко (32°−21° с.ш.) омывается водами Канарского течения, которое следует в юго-западном направлении. Прибрежные воды Мавритании (21°−16° с.ш.) находятся под воздействием северной ветви Межпассатного (Экваториального) противотечения, которое идёт в северном направлении. На участках, расположенных у выступающих мысов, образуются квазистационарные круговороты. На шельфе бóльшую часть года наблюдаются апвеллинги (Доманевский, 1998; Берников и др., 2002).
На этой акватории происходит интенсивный нерест и нагул молоди стайных неритических и мезопелагических рыб. В верхнем 100-метровом слое над шельфом отмечается более 170 видов пелагических икринок и личинок рыб (Blache et al., 1970; Калинина, 1981; Доманевский, 1998; Берников и др., 2002; Архипов, 2015а). Места наибольших скоплений ихтиопланктона у берегов северо-западного побережья Африки привязаны к динамическим процессам поверхностных вод (зонам вергенции), происходящим в этом районе, и сравнительно постоянны (Архипов, 2006, 2015а). Массовыми представителями ихтиоцена ЦВА являются такие промысловые виды, как европейская сардина Sardina pilchardus, европейская ставрида Trachurus trachurus, западноафриканская ставрида T. trecae, восточная скумбрия Scomber colias, круглая сардинелла Sardinella aurita. Распространёнными являются и некоторые другие промысловые виды рыб (плоская сардинелла Sardinella maderensis, европейский анчоус Engraulis encrasicolus, каранкс Caranx rhonchus, пеламида Sarda sarda и другие), однако повышение их численности наблюдается не каждый год и не на всей рассматриваемой акватории. Разные виды морских карасей (Sparidae), миктофид (Myctophidae) и гоностоматид (Gonostomatidae) также часто встречаются в ихтиофауне практически круглый год (Калинина, 1981; Доманевский, 1998; Берников и др., 2002; Архипов, 2006, 2015а).
В результате многолетних исследований получены данные по качественному и количественному составу и пространственному распределению ихтиопланктона в разные сезоны, установлены районы и периоды нереста массовых видов рыб в северной части ЦВА (Калинина, 1981; Седлецкая, 1983; Rodriguez et al., 2001; Nellen, Ruseler, 2004; Архипов, 2006, 2015а). Исследования изменений численности массовых рыб на ранних стадиях их развития являются важными составляющими рациональной эксплуатации водных биоресурсов, основанной на прогнозировании величины будущих поколений с различной заблаговременностью (Ahlstrom, Moser, 1976; Дехник и др., 1985; Архипов, 2006, 2015б).
Цель работы – проанализировать особенности распределения разных экологических групп ихтиопланктона северной ЦВА, используя статистические методы многомерного анализа данных.
МАТЕРИАЛ И МЕТОДИКА
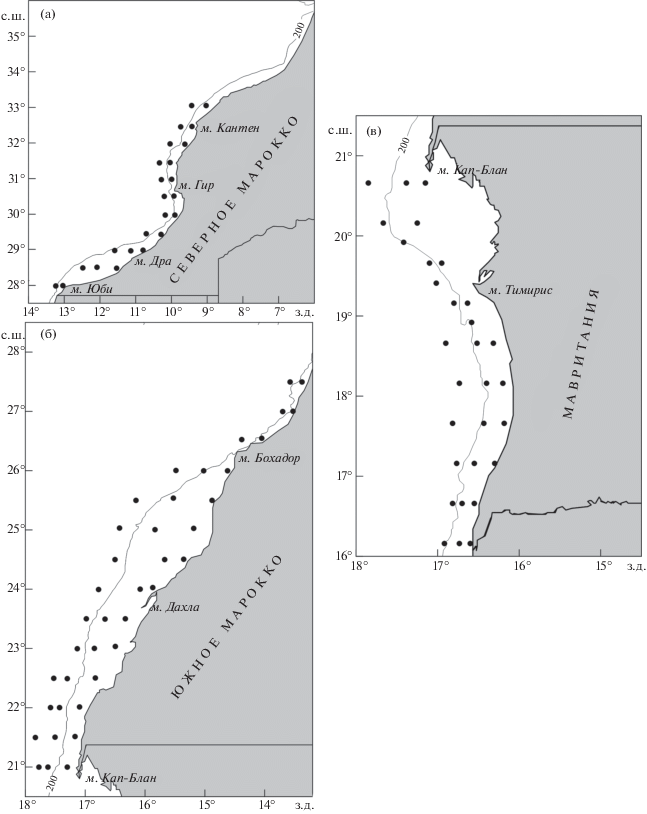
Материалы по распределению ихтиопланктона в слое воды 0−100 м (0−дно) получены в ходе комплексных съёмок в ноябре 2007 г. в северной части Марокко (19 станций), в ноябре−декабре 2012 г. в Мавритании (26 станций) и в октябре−ноябре 2018 г. в южной части Марокко (33 станции). Планктонные станции на акваториях съёмок располагались над глубинами от 20 до 1000 м (рис. 1). Пробы отбирали планктоносборщиком “Бонго-20” с площадью входного отверстия 0.03 м2, газом с ячеёй 417−333 мкм. Ступенчато-косой лов выполняли на горизонтах 100, 50, 35, 25, 10 и 0 м по 1.5−3.0 мин на каждом горизонте при скорости судна 2−3 узла (Smith, Richardson, 1977; Методические указания …, 1983). В ходе камеральной обработки под бинокулярным микроскопом МБС-10 (увеличение 8 × 2−4) определяли качественный (Калинина, 1981; Mater, Çoker, 2002) и количественный состав икринок и личинок рыб.
Рис. 1.
Сетки станций в районах исследований: а – северная часть Марокко, б – южная часть Марокко, в – Мавритания; (⚫) – станции, (——) − изобаты.
Для статистического анализа полученных данных выбраны непараметрический метод Краскала и процедура многомерного шкалирования с использованием коэффициента сходства Брея−Кертиса. Метод визуализации данных в виде дендрограмм и диаграмм рекомендован в экологических исследованиях и интегрирован в пакете программ PRIMER®6 (Clarke, Warwick, 2001; Clarke, Gorley, 2006). Так как решения отнесения пограничных объектов к тому или иному кластеру могут быть до некоторой степени произвольными даже в ситуации, когда объекты хорошо дифференцированы на группы, кластерный анализ рекомендуют использовать совместно с многомерным шкалированием. Согласованность результатов двух этих методов показывает их адекватность. Многомерное шкалирование позволяет изобразить совокупность объектов в виде набора точек в пространстве небольшой размерности и более или менее адекватно отражает исходное взаимоотношение между ними. В результате объекты, которым в исходной матрице соответствуют меньшие меры сходств, находятся дальше друг от друга, а объекты, которым соответствуют большие меры сходств, – ближе. Для проведения кластерного анализа стандартизированных и трансформированных данных применён иерархический метод с использованием группового осреднения. В ходе анализа выделяли группы ихтиопланктона, приуроченные к определённым глубинам. Такой анализ даёт возможность наглядно представить полученные результаты и выявить скрытые признаки и свойства пространственного распределения планктонных объектов (Clarke, Warwick, 2001; Clarke, Gorley, 2006; Лидванов, 2014; Дмитриева, 2017).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Динамическая структура вод в неритической зоне северной части ЦВА влияет на расположение нерестовых скоплений, пути переноса икринок и личинок и местонахождение молоди стайных пелагических рыб. Наличие синоптических вихрей разного знака в зоне прибрежных апвеллингов обусловливает “пятнистость” распределения икры и личинок. Эти пятна часто наблюдаются на периферии вихревых образований. Однако выноса икринок за пределы синоптических вихревых образований, как правило, не происходит ввиду короткого периода эмбрионального развития (1.5−2.0 сут). Дрейфовые миграции более характерны для личиночного и малькового этапов развития рыб (Калинина, 1981; Седлецкая, 1983; Архипов, Седлецкая, 2000; Rodriguez et al., 2001; Nellen, Ruseler, 2004; Архипов, 2006, 2015а).
В рассматриваемых районах можно выделить две экологические группы ихтиопланктона: мезопелагическую (океаническое сообщество) – представители семейств Myctophidae, Gonostomatidae и неритическую (прибрежное сообщество) – Clupeidae, Sparidae. Подобные группировки выявлены и для мезозоопланктона: неритическая, биотопически ассоциированная с водами прибрежного апвеллинга; и дальненеритическая, удалённая от берегов и связанная с водами Канарского течения (Лидванов, 2014; Лидванов и др., 2018).
В северной части Марокко в конце осени начинается нерест субтропических видов рыб, в ихтиопланктоне преобладают икринки и личинки европейской сардины (Clupeidae). Нерест разных видов миктофид, гоностоматид и спаровых с разной интенсивностью продолжается бóльшую часть года. В ноябре 2007 г. в рассматриваемом районе личинки сельдевых и спаровых видов рыб распределялись на шельфе над глубинами <100 м; икринки и личинки светящихся анчоусов (Myctophidae) и гоностоматид − в удалённых районах континентального шельфа и над свалом глубин (рис. 2, 3). Индекс сходства в экологических группах составил 65%. Ранее были описаны подобные закономерности в распределении разных групп ихтиопланктона и мезозоопланктона северной части Марокко для других периодов (Лидванов и др., 2018; Пак и др., 2018).
Рис. 2.
Дендрограмма кластерного анализа распределения ихтиопланктона разных экологических
групп в прибрежных водах северной части Марокко, ноябрь 2007 г.; здесь и на рис. 3–7: (▼) – прибрежное сообщество (Myctophidae, Gonostomatidae), (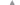 ) – океаническое сообщество (Clupeidae, Sparidae), (- - -) - показатель индекса сходства
в экологических группах, цифрами обозначена глубина в районе станции.
) – океаническое сообщество (Clupeidae, Sparidae), (- - -) - показатель индекса сходства
в экологических группах, цифрами обозначена глубина в районе станции.
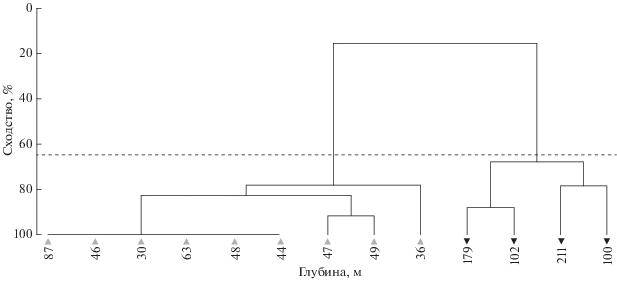
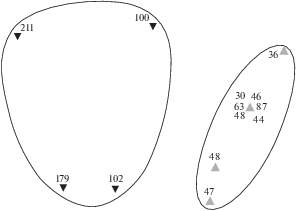
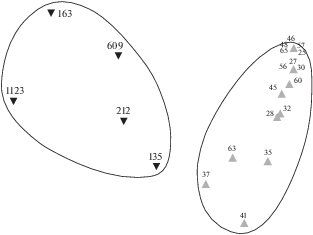
Рис. 3.
Диаграмма распределения ихтиопланктона разных экологических групп в прибрежных водах северной части Марокко, ноябрь 2007 г.
В южной части Марокко осенью заканчивается нерест у тропических видов рыб (в основном – сардинелл) и начинается у субтропических (в ихтиопланктоне преобладают европейские сардины ранних стадий развития). Продолжается нерест разных видов спаровых, миктофид и гоностоматид. В октябре−ноябре 2018 г. икринки и личинки сельдевых и спаровых видов рыб распределялись на шельфе, как правило, над глубинами <100 м; икринки и личинки миктофид и гоностоматид встречались в небольшом количестве в конце континентального шельфа и над свалом глубин (рис. 4, 5). Индекс сходства в экологических группах ихтиопланктона составил 63%. Близкие закономерности в распределении мезозоопланктона и рыб ранних стадий развития для южной части Марокко отмечены и в другие годы (Архипов и др., 2015; Лидванов и др., 2018; Пак и др., 2018).
Рис. 4.
Дендрограмма кластерного анализа распределения ихтиопланктона разных экологических групп в прибрежных водах южной части Марокко, октябрь−ноябрь 2018 г.
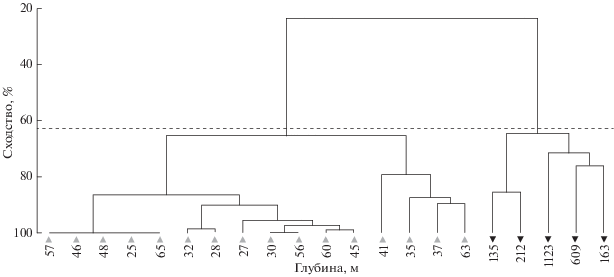
Рис. 5.
Диаграмма распределения ихтиопланктона разных экологических групп в прибрежных водах южной части Марокко, октябрь−ноябрь 2018 г.
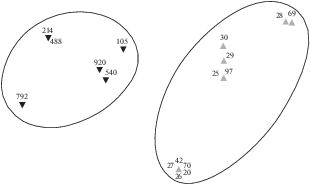
В районе Мавритании проанализировано распределение ихтиопланктона по данным ноябрьско-декабрьского рейса 2012 г. Полученные результаты представлены на рисунках 6−7. В конце осени−начале зимы в Мавритании практически заканчивается нерест у тропических видов рыб (круглой и плоской сардинелл) и начинается у субтропических (в основном у европейской сардины). Наблюдается нерест разных видов спаровых, миктофид и гоностоматид. В ноябре−декабре 2012 г. икринки и личинки сельдевых и спаровых видов рыб распределялись на шельфе, как правило, над глубинами <100 м; икринки и личинки миктофид и гоностоматид встречались в небольшом количестве у границ континентального шельфа и над свалом глубин (рис. 6, 7). Индекс сходства в экологических группах ихтиопланктона составил 33%.
Рис. 6.
Дендрограмма кластерного анализа распределения ихтиопланктона разных экологических групп в прибрежных водах Мавритании, ноябрь−декабрь 2012 г.
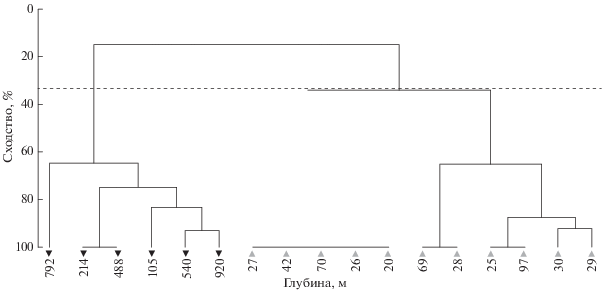
Рис. 7.
Диаграмма распределения ихтиопланктона разных экологических групп в ноябре−декабре 2012 г. в прибрежных водах Мавритании, ноябрь−декабрь 2012 г.
Из представленных данных и материалов предыдущих публикаций (Архипов и др., 2015; Пак и др., 2018) следует, что икринки и личинки Myctophidae, Gonostomatidae и некоторых других семейств в основном встречаются в океанической части съёмки на глубоководных станциях; икринки и личинки ранних стадий развития Clupeidae, Sparidae и других семейств – в прибрежной части. В средней части континентального шельфа (над глубинами 100−200 м) присутствуют икринки и личинки неритического и мезопелагического комплексов. Подтверждено представление о том, что мезопелагические виды рыб (миктофовые, гоностомовые) чаще нерестятся на удалении от берегов, неретические (сельдевые и спаровые) – ближе к побережью. Результаты многомерного анализа наглядно показали, что в районах исследований, как правило, можно выделить две экологические группы ихтиопланктона: мезопелагическую (удалённую от побережья) – Myctophidae, Gonostomatidae и неритическую (прибрежную) – Clupeidae, Sparidae.
ВЫВОДЫ
1. Использование многомерных методов анализа данных по численности разных экологических групп ихтиопланктона позволяет объективно оценить сходство и различие видовой структуры массовых рыб на ранних стадиях развития, наблюдающиеся в разные периоды исследований, типизировать структурные состояния и на этой основе оценить их сезонные и межгодовые перестройки.
2. Для анализа распределения различных экологических групп ихтиопланктона можно использовать данные по численности икринок, так как выноса икры в процессе онтогенеза за пределы синоптических вихревых образований, как правило, не происходит ввиду короткого эмбрионального периода. Дрейфовые миграции более характерны для личиночной и мальковой фаз развития рыб, но и для этих этапов наблюдаются определённые закономерности в распределении. Можно также рассматривать распределение икринок и личинок совместно.
3. В северной части ЦВА (северная и южная части Марокко и Мавритания) часто можно выделить две экологические группы ихтиопланктона: мезопелагическую (удалённую от побережья) – Myctophidae, Gonostomatidae и неритическую (прибрежную) – Clupeidae, Sparidae.
Список литературы
Архипов А.Г. 2006. Динамика численности и особенности распределения ихтиопланктонных сообществ северной части центрально-восточной Атлантики и морей Средиземноморского бассейна. Калининград: Изд-во АтлантНИРО, 232 с.
Архипов А.Г. 2015а. Динамика численности икринок и личинок массовых видов рыб северной части центрально-восточной Атлантики // Вопр. ихтиологии. Т. 55. № 2. С. 173−179. https://doi.org/10.7868/S0042875215020010
Архипов А.Г. 2015б. Применение результатов изучения раннего онтогенеза морских промысловых рыб в рыбохозяйственной деятельности // Тр. ВНИРО. Т. 156. С. 14−35.
Архипов А.Г., Седлецкая В.А. 2000. Межгодовые и сезонные изменения численности и распределения ихтиопланктона у атлантического побережья Африки от мыса Спартель до мыса Кап-Блан // Сб. науч. тр. АтлантНИРО. Гидробиологические исследования в бассейне Атлантического океана. Т. 2. Морская гидробиология. С. 48−65.
Архипов А.Г., Лидванов В.В., Пак Р.А. 2015. Особенности пространственного распределения разных таксономических групп ихтиопланктона южной части Марокко // Тез. докл. II Междунар. конф. “Актуальные проблемы планктонологии”. Калининград: Изд-во КГТУ. С. 34−35.
Берников Р.Г., Доманевский Л.Н., Кудерский С.К., Яковлев В.Н. 2002. Центрально-восточная Атлантика // Промыслово-океанологические исследования в Атлантическом океане и южной части Тихого океана Т. 1 / Под ред. Яковлева В.Н. Калининград: Изд-во АтлантНИРО. С. 146−195.
Дехник Т.В., Серебряков В.П., Соин С.Г. 1985. Значение ранних стадий развития в формировании численности поколений // Теория формирования численности и рационального использования стад промысловых рыб. М.: Наука. С. 56−72.
Дмитриева О.А. 2017. Исследование закономерностей пространственно-временных изменений структурных и количественных показателей фитопланктона в различных районах Балтийского моря: Автореф. дис. … канд. биол. наук. М.: ВНИРО, 24 с.
Доманевский Л.Н. 1998. Рыбы и рыболовство в неритической зоне центрально-восточной Атлантики. Калининград: Изд-во АтлантНИРО, 195 с.
Калинина Э.М. 1981. Ихтиопланктон района Канарского течения. Киев: Наук. думка, 116 с.
Лидванов В.В. 2014. Мезозоопланктон в районе Канарского апвеллинга: Автореф. дис. … канд. биол. наук. СПб.: ЗИН РАН, 26 с.
Лидванов В.В., Грабко О.Г., Кукуев Е.И., Королькова Т.Г. 2018. Структура мезозоопланктонных сообществ прибрежных вод Марокко // Океанология. Т. 58. № 2. С. 230–245. https://doi.org/10.7868/S0030157418020077
Методические указания по сбору проб зоо- и ихтиопланктона планктоносборщиком “Бонго” и их обработке. 1983. Калининград: Изд-во АтлантНИРО, 36 с.
Пак Р.А., Архипов А.Г., Лидванов В.В. 2018. Пространственное распределение различных экологических групп ихтиопланктона в северной и южной частях Марокко // Изв. КГТУ. № 51. С. 25−34.
Седлецкая В.А. 1983. Ихтиопланктон Атлантического океана у северо-западных берегов Африки // Вопр. ихтиологии. Т. 23. Вып. 5. С. 862−865.
Ahlstrom E.N., Moser H.G. 1976. Eggs and larvae of fishes and their roles in systematic investigations and in fisheries // Rev. Trav. Inst. Peches Mar. V. 40. № 3−4. P. 379−398.
Blache J., Cadenat J., Stauch A. 1970. Faune tropicale // XVIII Cles de determination des poissons de mer signales dans l’Atlantique oriental. Paris: ORSTOM, 479 p.
Clarke K.R., Gorley R.N. 2006. PRIMER version 6: user manual tutorial. Plymounth: PRIMER-E Ltd., 190 p.
Clarke K.R., Warwick R.M. 2001. Change in marine communities: an approach to statistical analysis and interpretation. Plymouth: PRIMER-E Ltd., 175 p.
Mater S., Çoker T. 2002. Turkiye denizleri ihtiyoplankton atlasi. Izmir: Ege Univ. Basimevi, 211 p.
Nellen W., Ruseler S. 2004. Composition, horizontal and vertical distribution of ichthyoplankton in the Great Meteor Seamount area in September 1998 // Arch. Fish. Mar. Res. V. 51. № 1−3. P. 132−164.
Rodriguez J.M., Barton E.D., Eve L., Hernandez-Leon S. 2001. Mesozooplankton and ichtyoplankton distribution around Gran Canaria, an oceanic island in the NE Atlantic // Deep-Sea Res. Pt. I. V. 48. P. 2161−2183.
Smith P.E., Richardson S.L. 1977. Standard techniques for pelagic fish egg and larvae surveys // FAO Tech. Pap. № 175. Rome: FAO, 95 p.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии