

Вопросы ихтиологии, 2020, T. 60, № 5, стр. 572-583
Взаимоотношения хищник–жертва на примере судака Sander lucioperca и тюльки Clupeonella cultriventris Рыбинского водохранилища в условиях потепления климата
М. Н. Иванова 1, А. Н. Свирская 1, *, М. И. Базаров 1
1 Институт биологии внутренних вод РАН − ИБВВ РАН
Ярославская область, Борок, Россия
* E-mail: svirs@ibiw.yaroslavl.ru
Поступила в редакцию 01.08.2019
После доработки 06.11.2019
Принята к публикации 08.11.2019
Аннотация
Исследован состав пищи судака Sander lucioperca в Рыбинском водохранилище в период потепления климата в регионе и значительных колебаний уловов южного вселенца – тюльки Clupeonella cultriventris. В период относительно низкой численности тюльки (2007−2014 гг.) по сравнению со следующим после её массовой вспышки численности годом (2016) доля этого вида в пище судака была в два раза меньше (28.3 против 60.7% числа съеденных жертв). В 2007−2016 гг. тюлька занимала первое место в рационе судака: её доля в его рационе на русловых и пойменных участках водоёма составляла соответственно 39.5 и 33.9%. В 2010-е гг. период интенсивного нагула судака длился до декабря включительно. По сравнению с периодом климатической нормы у младших возрастных групп судака (3+−6+) изменились сроки и места нагула: они нагуливаются на биотопах открытой и закрытой литорали, питаясь молодью плотвы Rutilus rutilus. Основным кормовым объектом особей старше 6+ остаётся тюлька − обитатель русловых зон речных плёсов.
Потепление климата в XXI в. становится важным фактором, влияющим в целом на экосистему Рыбинского водохранилища и на отдельные её звенья (Рыбы …, 2015; Лазарева и др., 2018). Так, за 30-летний период (1976−2005) средняя температура поверхностного слоя воды в водоёме возросла на 3.1°С, при максимальный скорости повышения в июле − 0.7−1.2°С/10 лет (Литвинов, Рощупко, 2010). В 2000-е гг. средняя продолжительность вегетационного периода увеличилась на 20 сут (с 193 до 213) за счёт более позднего наступления ледостава (конец ноября) (Литвинов и др., 2012). На изменение климата среагировали все звенья экологических цепей водоёма и, прежде всего, первичное − фитопланктон (Девяткин и др., 2012; Литвинов и др., 2012, 2014; Минеева и др., 2016; Сахарова, Корнева, 2018). В зоопланктонном сообществе в 2004−2010 гг. зарегистрировано увеличение в 1.5−3.0 раза численности всех копепод, большинства крупных кладоцер, а также сдвиг структуры сообщества в сторону увеличения доли крупных организмов (Лазарева, Соколова, 2016). Вследствие потепления увеличилась продолжительность периода массового развития летнего зоопланктона и сформировался мощный пик его обилия во второй половине лета. Это послужило важным фактором для успешной натурализации нового планктофага − тюльки Clupeonella cultriventris (Лазарева, Соколова, 2016).
Черноморско-каспийская тюлька появилась в Рыбинском водохранилище в конце 1990-х гг. Первые две фазы её натурализации (проникновение и начало воспроизводства) прошли в течение шести лет (Кияшко, Слынько, 2012). Третья фаза натурализация вселенца длилась три года (2000−2002); в течение этого периода наблюдался интенсивный рост его численности, в результате чего он стал доминирующим видом пелагиали (Рыбы …, 2015). В эти годы тюлька стала одним из основных объектов питания всех хищных рыб в водоёме и, прежде всего, пелагического хищника − судака Sander lucioperca (Степанов, Кияшко, 2008). Она составляла 61% общего числа рыб-жертв судака, при этом значение двух традиционных местных видов (окуня Perca fluviatilis и ерша Gymnocephalus cernuus) в его пище уменьшилось (Герасимов и др., 2013; Иванова, Свирская, 2014). После 2003 г. популяция вселенца перешла в IV фазу развития, когда динамика её численности приобрела колебательный характер. Межгодовые флуктуации численности были значительными: средние уловы тюльки в 2007 г., например, отличались от таковых в 2004 г. в 5.4 раза, в 2012 и 2015 гг. − в 12 раз. Но и в этот период тюлька по-прежнему сохраняла статус доминирующего вида, её доля в уловах составляла 34−78% (Герасимов и др., 2009, 2018; Кияшко и др., 2012).
Цель работы − изучить питание судака Рыбинского водохранилища в разных биотопах его обитания в условиях изменяющейся численности вселенца и продолжающегося потепления климата.
МАТЕРИАЛ И МЕТОДИКА
Материал собран в 2007−2016 гг. в Рыбинском водохранилище на трёх русловых участках (Центральный, Шекснинский и Моложский плёсы) и в пойменном участке Волжского плёса. Центральный плёс представляет собой затопленное междуречье рек Шексна, Молога и Волга; Шекснинский плёс расположен в северо-западной части водохранилища в бывшей долине р. Шексна, Моложский − в северо-восточной его части вдоль русла р. Молога; Волжский − в южной части водохранилища. Русловые участки этих плёсов имеют глубину от 11 до 23−30 м; пойменный участок Волжского плёса − до 9 м (Рыбинское водохранилище …, 1972).
На русловых участках водоёма судаков отлавливали донным (горизонтальное раскрытие 18 м, вертикальное – 2 м, ячея в кутке 22 мм) и разноглубинным (соответственно 17.3 и 1.8 м, 4 мм) тралом на глубине 10−14 м во время ежегодных осенних рейсов. Параллельно с тралением проводили гидроакустическую съёмку с целью определения обилия рыб на биотопе по методике, разработанной Юдановым с соавторами (1984). Горизонт лова определяли по данным эхолота. Всего на русловых участках выловлено 247 судаков (в Центральном плёсе 79 экз., в Шекснинском − 82, в Моложском – 86). На пойменном участке Волжского плёса, находящемся на расстоянии 500 м от затопленного русла Волги, судаков отлавливали ежегодно в марте–мае и сентябре–декабре сетями (ячея 40−80 мм), выставляемыми на глубине 5−10 м. Всего на пойме было выловлено 258 судаков.
У всех рыб измеряли стандартную длину (SL) и массу, определяли пол, стадию зрелости половых продуктов, возраст (Правдин, 1966); извлекали желудочно-кишечный тракт и замораживали. Камеральную обработку содержимого желудка хищника проводили по методике Фортунатовой и Поповой (1973). В пищевом комке подсчитывали число и видовой состав заглоченных рыб, их длину. Вид и размеры переваренных рыб-жертв определяли по форме и величине глоточных зубов (для Cyprinidae) и предкрышечных костей (для Percidae) по таблицам Ковалева (1958). Всего исследовано содержимое желудков 505 судаков. Значение отдельных видов рыб в питании хищника оценивали в процентах общего числа съеденных жертв.
Отлов тюльки проводили во время тралово-акустической съёмки на тех же станциях, что и отлов судака. Численность тюльки в водоёме оценивали с помощью показателя улова на усилие исследовательского трала (экз/10 мин траления). Данные по уловам тюльки, собранные на 15 стандартных станциях, расположенных по всей акватории водоёма, суммировали и подсчитывали среднее значение показателя для водоёма в каждый год наблюдений. Плотность рыб (экз/м2) на отдельных станциях рассчитывали как по траловым уловам, так и по данным, полученным с эхолота EY-500 с помощью программы Echoview 5.0 методом эхоинтеграции.
РЕЗУЛЬТАТЫ
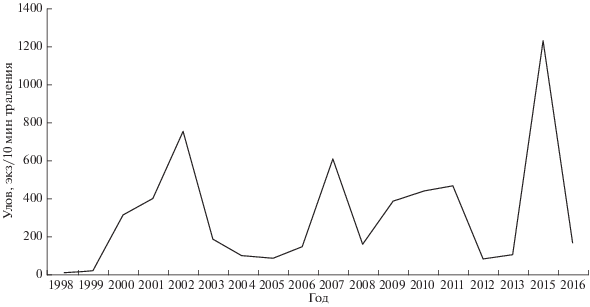
Динамика уловов тюльки (рис. 1). Вселившись в Рыбинское водохранилище, вид достиг высокой численности к 2002 г. (за 10 мин траления вылавливали в среднем 757 экз.). В течение последующих 12 лет (2003−2014) наблюдали значительные колебания уловов вселенца от 46 до 544 экз/10 мин траления. В 2015 г. была отмечена мощная вспышка численности тюльки в водоёме (1229 экз/10 мин траления), а на следующий год − её резкий спад (в 7.4 раза).
Скопления тюльки обычно имеют шаровидную форму, занимают наиболее продуктивные биотопы пелагиали, концентрируясь на горизонте 4−8 м (Рыбы …, 2015). На эхограмме, сделанной в августе 2015 г. (рис. 2), отчётливо видны скопления тюльки, различающиеся по объёму (7.67, 27.25, 4.17 м3) и плотности (соответственно 75.83, 19.52, 28.51 экз/м2). Они находятся на расстоянии 5−10 м друг от друга. Рядом со скоплениями тюльки часто фиксировали эхоотметки крупных рыб. В годы с низкой численностью вселенца его скопления были расположены на большем расстоянии друг от друга.
Рис. 2.
Эхограмма распределения тюльки Clupeonella cultriventris в Рыбинском водохранилище в августе 2015 г.: 1 − эхоотметки крупных рыб, 2 − скопления тюльки.

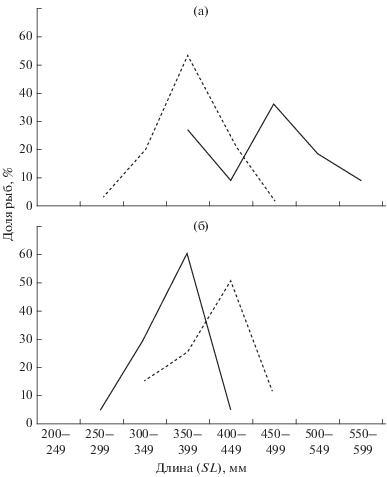
Размерный и возрастной состав судака. На русловых участках водоёма встречались судаки SL 185−750 мм; основу улова (51.3%) составляли особи SL 300−499 мм (рис. 3а). Кривая распределения рыб по длине тела сильно вытянута вправо за счёт наличия крупных особей: SL 450−599 мм (23.4%) и ≥600 мм (6.5%). Доля мелких рыб (SL < < 300 мм) составляла 18.8%. Размерный ряд судаков, выловленных на пойме, был менее широким − SL 167−600 мм – с модальным классом SL 350− 399 мм (33.3%). На долю рыб SL 400−449 мм приходилось 16.7%, SL 300−349 мм − 24.3%. Более крупные хищники (SL ≥ 450 мм) составляли в уловах всего 7.6%, а мелкие (SL < 300 мм) – 18.1%.
Рис. 3.
Размерный (а) и возрастной (б) состав судака Sander lucioperca на русловых (−⚫−) и пойменных (−⚪−) участках Рыбинского водохранилища в 2007−2016 гг. Число исследованных рыб: русло − 247 экз., пойма − 258 экз.
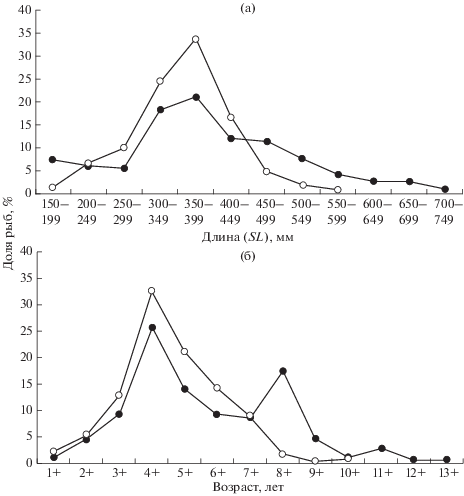
Возрастной состав судака на сравниваемых биотопах тоже различался (рис. 3б). На русловых участках отмечены особи в возрасте 1+−13+. Кривая их распределения имеет два модальных класса − 4+ (25.5%) и 8+ (17.4%). Возрастные группы 1+−3+ составляли 15.2% улова, 5+−7+ − 31.9%, 8+−11+ – 26.2%, 12+−13+ – 1.2%. На пойме встречались судаки в возрасте 1+−10+. Кривая их распределения была одновершинной с модальным классом 4+ (32.5%). По численности здесь доминировали особи в возрасте 5+−7+ (44%), более старые рыбы составляли 3.1%, а возрастные группы 1+−3+ − 20.4% улова.
Таким образом, в выборке из русла присутствовали более крупные по размерам и старшие по возрасту рыбы, чем в выборке из поймы.
Питание судака. Спектр питания судака Рыбинского водохранилища в 2007−2016 гг. включал 15 видов рыб: тюлька, окунь, ёрш, плотва Rutilus rutilus, синец Abramis ballerus, лещ A. brama, густера Blicca bjoerkna, чехонь Pelecus cultratus, карась Carassius auratus, уклейка Alburnus alburnus, молодь судака, бёрш Sander volgensis, щука Esox lucius, щиповка Cobitis taenia, ряпушка Coregonus albula.
На русловых участках водохранилища видовой состав пищи судака заметно различался. Так, в Центральном плёсе (рис. 4а) тюлька оставалась его постоянным и основным кормовым объектом (30−52% общего числа жертв) во все годы наблюдений11, несмотря на значительные колебания её численности в уловах пелагического трала. Заметную роль в питании судака имели также два вида местной фауны – ёрш и окунь. В Шекснинском плёсе тюлька входила в число основных кормовых объектов питания судака только в 2008, 2009 и 2016 гг. (рис. 4б). В 2011−2012 гг. потребление вселенца резко сократилось, несмотря на то, что самые высокие уловы тюльки в этом районе были зарегистрированы именно в 2011 г. На русловых участках Шексны хищники интенсивно откармливались местными видами: окунем, ершом и синцом, а лещ и плотва были второстепенными объектами питания. В Моложском плёсе (рис. 4в) тюлька вообще не встречалась в желудках судака (2008−2009 гг.) или была второстепенным объектом откорма (2011−2012 гг.). Состав его пищи характеризовался почти ежегодной сменой основных кормовых объектов: в 2008 г. доминировал окунь (63%), в 2009 г. – ёрш (75%), в 2011−2012 гг. – плотва (40−48%), в 2016 г. – ёрш (32%) и тюлька (29%).
Рис. 4.
Состав пищи судака Sander lucioperca на русловых участках Рыбинского водохранилища, осень 2007−2016 гг.: а − Центральный плёс, б − Шекснинский плёс, в − Моложский плёс; 1 − тюлька Clupeonella cultriventris, 2 − окунь Perca fluviatilis, 3 − ёрш Gymnocephalus cernuus, 4 − плотва Rutilus rutilus, 5 − синец Abramis balerus, 6 − лещ A. brama, 7 – судак, 8 − прочие виды рыб; (− ◼ −) − уловы тюльки; n − число исследованных рыб.
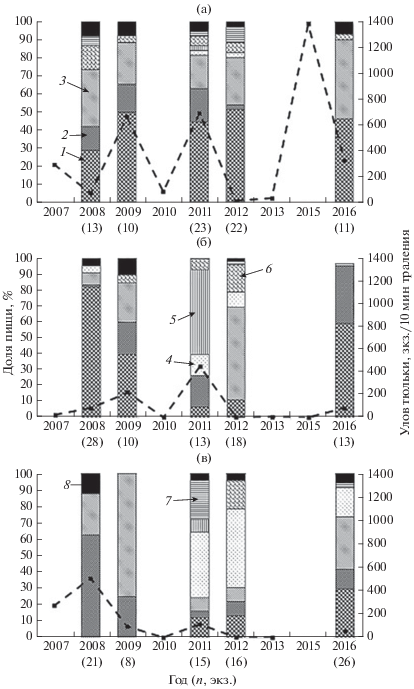
Из приведённых на рис. 4 данных видно, что в отдельные годы численность тюльки в учётных уловах малькового трала не совпадала с её долей в рационе судака. Это свидетельствует о его избирательности по отношению к тюльке, поскольку она встречалась в желудках хищников в годы минимальных уловов и, наоборот, отсутствовала в годы относительно высокой численности. На видовой состав пищи судака вспышка численности вселенца в 2015 г. продолжала оказывать влияние и на следующий год, особенно в Моложском и Шекснинском плёсах.
На пойменном участке Волжского плёса спектр питания судака более изменчивый и разнообразный, чем в русловых зонах водохранилища. Соотношение видов жертв в рационе судака в значительной степени зависит от обилия сеголеток того или иного вида в водоёме. По данным Столбунова (2016), в многоводные годы численность всей молоди и отдельных её видов в уловах в 1.5−2.0 раза выше, чем в маловодные; например, плотность рыб в скоплениях в многоводном 2013 г. превышала таковую в маловодном 2014 г. в 1.9 раза (3.9 против 2.1 экз/м2). В 2008 г. в связи с поздним нерестом фитофилов (акватории большинства прибрежных нерестилищ оказались незалитыми из-за низкого уровня воды в водоёме) наиболее многочисленными в стаях молоди были лещ и густера. Эти два вида и доминировали в пище судака в тот год, составляя в сумме >82%. Кроме них в его желудках встречался только окунь (рис. 5а). В многоводные годы, которые характеризуются ранним и эффективным нерестом фитофилов, спектр питания хищников был более широким: лещ, окунь, плотва, синец (2013 г.). В маловодный 2014 г. отчётливо прослеживалась сезонная смена рыб-жертв в рационе судака: весной он откармливался преимущественно на стаях плотвы, а осенью – на скоплениях тюльки, которая в это время проходила по русловому участку Волжского плёса на зимовку. Весной и осенью 2016 г. судак питался только тюлькой.
Рис. 5.
Состав пищи судака Sander lucioperca в Волжском плёсе Рыбинского водохранилища в весенний и осенний сезоны (а) и в декабре (б) 2007−2016 гг.; 9 − густера Blicca bjoerkna; 1–8 – см. на рис. 4.
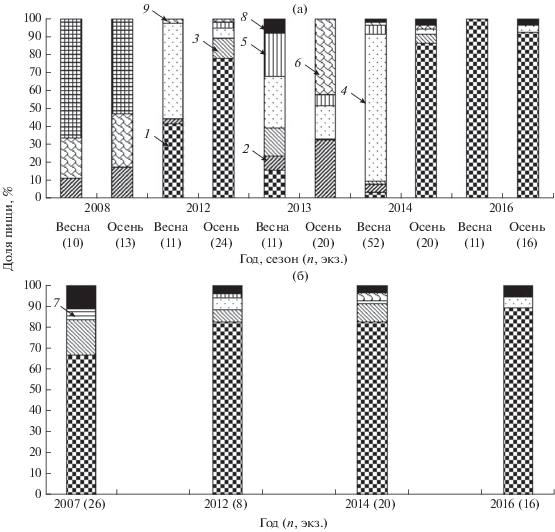
Определённый интерес представляют наши данные по питанию судака в декабре (рис. 5б). В 2007 г. пища была обнаружена в желудках 73% судаков, в 2012 г. − 75%, в 2014 г. − 100%, в 2016 г. − 92%. Среди жертв во все годы наблюдений доминировала тюлька (67−89%), преимущественно её 2-летки SL 60−80 мм; другие виды рыб имели второстепенное значение. Осенью тюлька начинает перемещаться к местам зимовки – в глубоководные районы речных плёсов с наличием стокового течения (Степанов, 2011). Во время перехода тюльки к местам зимовки, а также в декабре судак откармливался на её скоплениях. Питались тюлькой преимущественно крупные судаки (SL > 400 мм).
Размерный состав судака в годы разной водности. Весной многоводного 2013 г. (рис. 6а) на биотопах поймы доминировали по численности (72.8%) крупные (>400 мм) и старшие (≥7+) особи судака. Эту группу составляли уже отнерестившиеся самки (56%) и самцы (44%) с половыми продуктами VI−II стадии зрелости. Все они питались, но не очень активно. В их желудках находили по одному (редко два) экземпляру плотвы (33.2%), а также ерша, синца, карася и окуня (каждый вид по 16.7%).
Рис. 6.
Размерный состав судака Sander lucioperca в Волжском плёсе Рыбинского водохранилища в многоводный 2013 (—) и маловодный 2014 (· · ·) гг. : а – весна (соответственно 11 и 52 экз.), б – осень (20 и 20 экз.).
Молодые судаки (SL 300−400 мм, 4+−6+) в большом количестве (90%) встречались на биотопах поймы осенью 2013 г. (рис. 6б). Они начали интенсивно питаться молодью рыб в конце сентября, когда сеголетки достигли SL ≥ 30 мм, при этом в одном желудке хищника встречалось по пять–семь жертв. В сентябре–октябре основу их рациона составили лещ (41.8%), окунь (32.7%) и плотва (18.2%). Весной следующего 2014 г. хищники этой размерной группы откармливались преимущественно годовиками плотвы (81.5%). Размеры годовиков плотвы, в стаях которой судак выбирал жертв, варьировали от 50 до 80 мм. В основном хищники поедали рыб SL 60−70 мм (80.3%). Более мелкую (SL 50−60 мм) и крупную плотву (SL 80−100 мм) судак захватывал в меньшем количестве (соответственно 3.6 и 16.1%).
Весной маловодного 2014 г. крупных судаков (SL > 400 мм, ≥ 7+) в литорали водоёма не было (из-за низкого уровня воды).
Размеры жертв судака. На русловых участках водоёма в желудках судака была обнаружена тюлька SL 20−100 мм (рис. 7а). Чаще всего они потребляли рыб SL 40−79 мм (89%), это взрослые особи возрастом 1+−2+. Мелкими (SL < 40 мм, масса 1−3 г) и крупными экземплярами (SL ≥ 80 мм, 6−10 г), достигшими предельного возраста 3+ вселенца в водохранилище (Осипов, Кияшко, 2006; Кияшко и др., 2012), судаки питались крайне редко. На пойменных участках водоёма хищники потребляли тюльку SL 40−89 мм, доминировали среди них рыбы SL 60−79 мм (67.6%). Следует подчеркнуть, что мелкие особи этого вида (сеголетки SL < 50 мм) встречались в пище судаков ежегодно.
Рис. 7.
Размеры (SL) жертв судака Sander lucioperca на русловых (−⚫−) и пойменных (−⚪−) участках Рыбинского водохранилища: а − тюлька Clupeonella cultriventris, б − окунь Perca fluviatilis, в − ёрш Gymnocephalus cernuus, г − плотва Rutilus rutilus.
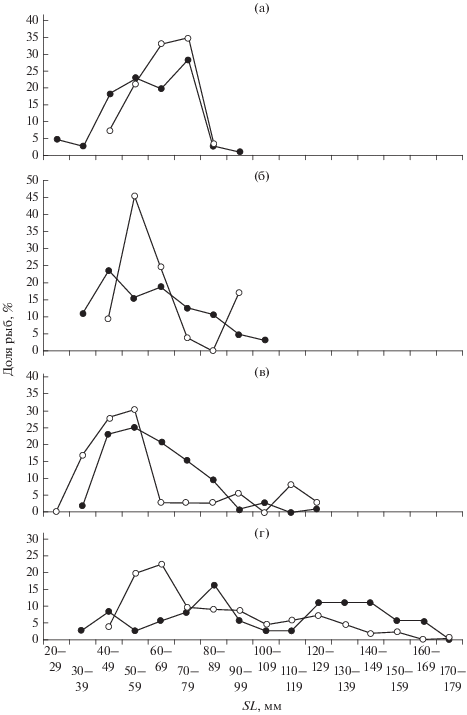
На русловых участках водоёма судак питался окунем разных размерных групп (SL 30−110 мм) и возраста (от сеголеток до взрослых рыб); с увеличением размеров жертвы её доля в пище хищника плавно уменьшалась (рис. 7б). Так, рыбы SL 40−49 мм составляли 23.5% общего числа съеденных окуней, SL 50−59 мм − 15.6%, SL 80−89 мм − 10.9%. Более крупные окуни (SL 90−110 мм) встречались в пище хищника в меньшем количестве (3.1−4.7%). На пойме судак питался в основном сеголетками окуня SL 50−59 мм (45.3%) и SL 60−69 мм (24.5%), а также его двухлетками SL 90−99 мм (17%).
Длина ерша в пище судака на русловых участках варьировала в пределах 30−130 мм (рис. 7в). Чаше всего в желудках хищников встречались особи SL 40−69 мм (68.9%). Размерный ряд ерша, изъятого из желудков судака, выловленного на пойменном участке, имеет сильно выраженную правостороннюю асимметрию с модальным классом SL 40−59 мм (58.3%). Доля более крупных ершей (SL ≥ 60 мм) в рационе хищника была в 5.4 раза меньше, чем на русле (8.4 против 45.6%).
Наиболее широкий размерный ряд у плотвы − SL 30−180 мм (рис. 7г). На русловых зонах обитания судак выбирал не только двухлеток плотвы SL 80−89 мм (16.4%), но и более крупных особей SL 120−149 мм (33.3%) в возрасте от 3+ и старше. А на пойменном участке от предпочитал (73.6%) сеголеток и годовиков плотвы SL 47−99 мм, ведущих стайный образ жизни. Модальный класс составляли особи SL 50−69 мм (42.3%), ими питался судак в литорали водохранилища. Крупные жертвы (SL ≥ 100 мм, возраст 2+ и 3+) встречались как в русловой зоне, так и на пойме; они, как правило, находились в желудках хищников поодиночке (редко по 2 экз.). Большей частью крупные жертвы разных видов встречались в желудках судака весной (после нереста) или летом, когда активность хищников понижена.
Приведённые на рис. 7 данные показывают, что на обоих исследованных биотопах водохранилища судак предпочитает поедать относительно мелкие кормовые объекты. Среди рыб-аборигенов жертвами хищника становились рыбы младших возрастных групп (наиболее многочисленная часть их популяций): среди окунёвых преобладали сеголетки, среди карповых – годовики. В скоплениях тюльки судаки поедали преимущественно взрослых особей.
ОБСУЖДЕНИЕ
Судак – пелагический хищник, обитающий в открытой зоне озёр, рек и водохранилищ, подстерегающий и преследующий добычу (Фортунатова, Попова, 1973; Попова, 1979). В Рыбинском водохранилище в период климатической нормы (1960−1970 гг.) судак образовывал многочисленные скопления вдоль затопленных русел рек, в эстуарных участках речных плёсов с глубинами до 14 м (Поддубный, 1971). Его основными кормовыми объектами в те годы были молодь окуня и ёрш, которыми питались особи всех размерных групп; важную роль в питании играли также плотва и корюшка Osmerus eperlanus (северный вселенец). В 1960-е гг. на долю этих четырёх видов приходилось 92% общего числа съеденных рыб-жертв (табл. 1). В этот период наблюдался рост численности данных видов рыб в водоёме (Рыбы …, 2015). Местом откорма судаков была пелагиаль Центрального и всех речных плёсов. Наиболее интенсивно питался хищник в летне-осенние месяцы (Иванова, 1966).
Таблица 1.
Состав пищи судака Sander lucioperca Рыбинского водохранилища в разные периоды, % общего числа жертв
| Компонент пищи | Годы наблюдений | |||
|---|---|---|---|---|
| 1960−1962 (климатическая норма) |
2007−2016 (потепление климата) | |||
| Весь водоём | Весь водоём | Русло | Пойма | |
| Тюлька | − | 35.7 | 39.5 | 33.9 |
| Окунь | 31.1 | 10.5 | 12.4 | 9.1 |
| Ерш | 28.0 | 13.3 | 24.7 | 5.0 |
| Плотва | 18.2 | 22.4 | 8.0 | 33.0 |
| Корюшка | 14.6 | − | − | − |
| Прочие виды рыб | 8.1 | 18.1 | 15.4 | 19.0 |
В годы наших наблюдений (2007−2016 гг.) первое место в рационе судака Рыбинского водохранилища заняла тюлька (35.7%), которая стала доминирующим видом в пелагических скоплениях рыб с 2002 г. (Рыбы …, 2015). Увеличение её численности произошло на фоне заметного снижения обилия таких значимых для судака видов жертв, как ёрш и молодь окуня (Герасимов и др., 2018). И, как следствие, их доли в пище хищника сократились соответственно в три и два раза по сравнению с годами климатической нормы (табл. 1). Судак интенсивно откармливался тюлькой как на русловых, так и на пойменных участках водоёма (соответственно 39.5 и 33.9% общего числа жертв). Доля представителей местной фауны изменялась в зависимости от района его нагула: в пелагиали судак в большей степени поедал окунёвых рыб (особенно ерша), а на пойме − плотву.
Пелагиаль Центрального и эстуарии речных плесов являются основным местом обитания и нагула особей судака старшего возраста (Рыбы …, 2015). Тюлька в составе пищи судака русловых участков Центрального плёса встречалась ежегодно и в большом количестве (30−52% общего числа съеденных рыб). В то время как в Шекснинском плёсе она входила в число основных кормовых объектов хищника лишь трижды (в 2008, 2009 и 2016 гг.), в Моложском − один раз (в 2016 г.). В остальные годы вселенец становился второстепенным объектом откорма судаков или же вообще не встречался в спектре их питания (рис. 4). Важную роль в рационе хищников в такие годы играли рыбы-аборигены: ёрш, окунь, плотва, синец и другие.
Доля тюльки в составе пище судаков не всегда находилась в прямой корреляции с её численностью в уловах малькового трала. Вселенец, например, встречался в желудках хищников и в годы минимальных уловов, что свидетельствует о наличии избирательности у судака по отношению к тюльке.
По мнению Карабанова (2013. С. 132) в отдельные годы в пелагиали Рыбинского водохранилища сохраняются зоны, на которых “могут нереститься тяготеющие к ним местные стада производителей”. Размножение даже незначительной части тюльки в отдельных точках пелагической зоны способствует сохранению всей популяции, а также объясняет появление инвазийного вида в пище судака в разных плёсах то в одном, то в другом году. Подтверждает это предположение ежегодное наличие мелких сеголеток тюльки (SL 30−50 мм) в желудках судаков, выловленных в Центральном плёсе. На долю этой возрастной группы приходилось от 20 (2016 г.) до 100% общего количества тюльки, съеденной хищниками, что свидетельствует о регулярном размножении вселенца в самом глубоководном участке водоёма.
В спектре питания судака тюльку начали отмечать ещё в 1996−1999 гг., несмотря на её низкую встречаемость в уловах (Герасимов и др., 2018). В 2000−2003 г., когда вселенец достиг первого пика своей численности, он стал основным объектом питания не только судака (61% общего числа съеденных рыб), но и других хищных рыб (Степанов, Кияшко, 2008).
Тюльке как короткоцикловому раносозревающему виду рыб свойственны межгодовые флуктуации численности (Никольский, 1974; Рыбы …, 2015). Период наших наблюдений охватывает годы как с относительно небольшими, так и с и очень высокими уловами тюльки в 2015 г. (рис. 1), что позволяет сопоставить соотношение отдельных видов жертв в пище судака в годы с разной численностью излюбленного кормового объекта в водоёме (табл. 2). В период невысокой численности тюльки (2007−2014 гг.) её доля в питании судака уменьшилась одновременно с ершом, но значительно увеличилась доля плотвы (30%), что свидетельствует об изменении мест нагула судака. На следующий год после самой мощной вспышки численности вселенца в Рыбинском водохранилище (в 2016 г.) в составе пищи судака доминирующая роль тюльки (60.7%) снова восстановилась; второе место занимал ёрш. Ранее временную замену тюльки в рационе судака на плотву и окуня отмечали Степанов и Кияшко (2008) летом 2003 г., когда резко сократилась численность вселенца в водоёме. Холодная зима 2002−2003 гг. в сочетании с низким уровнем воды привели к повсеместным заморам рыб, в том числе и тюльки; только к осени восстановилась её высокая доля в пище судака за счёт многочисленного поколения 2003 г. рождения.
Таблица 2.
Состав пищи судака Sander lucioperca Рыбинского водохранилища в годы с разной численностью тюльки Clupeonella cultriventris, % общего числа жертв
| Компонент пищи | Годы наблюдений | |||
|---|---|---|---|---|
| 1996−1999 | 2000−2003 | 2007−2014 | 2016 | |
| Герасимов и др., 2018 | Степанов, Кияшко, 2008 | Наши данные | ||
| Тюлька | 9.0 | 60.9 | 28.3 | 60.7 |
| Окунь | 28.0 | 5.3 | 13.0 | 9.7 |
| Ёрш | 2.0 | 27.0 | 14.7 | 18.1 |
| Плотва | 18.0 | 2.3 | 29.7 | 6.5 |
| Судак | 38.0 | – | 1.6 | 1.3 |
| Прочие виды | 5.0 | 4.5 | 12.7 | 3.7 |
Пойменные биотопы водохранилища, в которых размножаются фитофильные аборигенные виды рыб, являются важным источником кормовой базы судака. Сеголетки этих видов нагуливаются в районах нереста всё лето и отходят на глубину с падением уровня воды (Столбунов, 2007, 2016). Наиболее многочисленным видом в прибрежных группировках молоди является плотва, составляющая в отдельные годы до 90% скоплений (Столбунов, 2016). Особи младших возрастныхе групп судака (4+−6+, SL 300−400 мм) интенсивно питаются сеголетками фитофильных рыб в сентябре–октябре и годовиками плотвы апреле–мае. В результате биотопы открытой и закрытой литорали стали местом откорма самой многочисленной группы рыб (58−68%) в популяции судака (рис. 3) как осенью, так и весной, независимо от уровненного режима отдельных лет. Таким образом, интенсивный откорм судака прибрежной молодью (мелкими формами плотвы, окуня и других видов), не имеющими коммерческой ценности, включает этого хищника в потребителей прибрежного зоопланктона по цепочке: прибрежный зоопланктон → молодь рыб → судак (Гостев, Козловский, 1986).
Старшие возрастные группы судака (≥6 лет, SL > 400 мм) нагуливаются в пелагиали Центрального и речных плёсов, куда они возвращаются после нереста. Наибольшая интенсивность откорма рыб этой группы наблюдается с сентября по декабрь включительно, когда тюлька после окончания нагула поднимается в речные плёсы (Степанов, 2011). Для сравнения: в 1960−1970-е гг. (годы климатической нормы по: Литвинов, Рощупко, 2010) судак интенсивно питался в августе–сентябре, в октябре–ноябре с понижением температуры воды активность его питания заметно ослабевала (Иванова, 1968). Например, в декабре 1960 г. из 127 выловленных особей питались 54 (42.5%), тогда как в декабре 2007−2016 гг. доля питавшихся рыб была существенно выше – 73−100%. Подобное явление было отмечено в эти годы и для молоди рыб (Столбунов, 2016). Потепление климата в регионе (Литвинов, Законнова, 2012; Законнова, Литвинов, 2016) привело к более поздним срокам наступления ледостава и соответственно к увеличению продолжительности вегетационного периода. Удлинение вегетационного периода в водохранилище способствует продлению откорма старших особей в популяции хищников на скоплениях вселенца даже в зимний месяц.
Таким образом, полученные данные свидетельствуют о расхождении в сроках и местах откорма разных размерно-возрастных групп судака в середине второго десятилетия XXI в., что позволяет виду наиболее полно использовать кормовую базу водоёма. Занимая различные пищевые ниши, отдельные группы судака используют “имеющееся в водохранилище разнообразие местообитаний, что способствует устойчивому функционированию водных сообществ” (Герасимов и др., 2005. С. 108) в меняющихся условиях среды при продолжающемся потеплении климата в регионе.
Список литературы
Атлас пресноводных рыб России. 2002. Т. 2 / Под редакцией Решетникова Ю.С. М.: Наука, 253 с.
Герасимов Ю.В., Столбунов И.А., Павлов Д.Д. 2005. Роль поведенческого полиморфизма в процессе внутрипопуляционной сегрегации экологических ниш у рыб // Матер. докл. Междунар. конф. “Поведение рыб”. М.: Акварос. С. 104−109.
Герасимов Ю.В., Иванова М.Н., Стрельников А.С. 2009. Динамика пелагических скоплений рыб и изменения в составе пищи окуневых на разных этапах формирования ихтиофауны Рыбинского водохранилища (1950−2008 гг.) // Матер. XXVIII Междунар. конф. “Биологические ресурсы Белого моря и внутренних водоемов Европейского Севера”. Петрозаводск. С. 142−145.
Герасимов Ю.В., Стрельников А.С., Иванова М.Н. 2013. Динамика структурных показателей популяции судака Рыбинского водохранилища за период 1954−2010 гг. // Вопр. ихтиологии. Т. 53. № 1. С. 57−68. https://doi.org/10.7868/S0042875213010050
Герасимов Ю.В., Иванова М.Н., Свирская А.Н. 2018. Многолетние изменения роли местных и инвазийных видов рыб в питании хищных рыб Рыбинского водохранилища // Там же. Т. 58. № 5. С. 507−522. https://doi.org/10.1134/S0042875218040045
Гостев С.Н., Козловский С.В. 1986. К вопросу о роли тюльки в питании судака Куйбышевского водохранилища // Биология внутр. вод. № 69. С. 33−36.
Девяткин В.Г., Метелева Н.Ю., Вайновский П.А. 2012. О роли фитопланктона в формировании кислородного режима в связи с климатическими вариациями // Вода: химия и экология. № 12 (54). С. 68−74.
Законнова А.В., Литвинов А.С. 2016. Многолетние изменения гидроклиматического режима Рыбинского водохранилища // Тр. ИБВВ РАН. Вып. 75 (78). С. 16−22. https://doi.org/10.24411/0320-3557-2016-10016
Иванова М.Н. 1966. Питание и пищевые взаимоотношения хищных рыб в Рыбинском, Горьковском и Куйбышевском водохранилищах: Автореф. дис. … канд. биол. наук. М.: ЛГПИ, 17 с.
Иванова М.Н. 1968. О воздействии судака на популяции некоторых видов рыб // Тр. ИБВВ АН СССР. № 16 (19). С. 166−181.
Иванова М.Н., Свирская А.Н. 2014. Динамика питания хищных рыб Рыбинского водохранилища за период 1949−2012 гг. // Матер. докл. II Всерос. конф. “Современное состояние биоресурсов внутренних водоемов”. Т. 1. М.: Полиграф-Плюс. С. 204−212.
Карабанов Д.П. 2013. Генетические адаптации черноморско-каспийской тюльки Clupeonella cultriventris (Nordmann, 1840) (Actinopterygii: Clupeidae). Воронеж: Науч. книга, 179 с.
Кияшко В.И., Слынько Ю.В. 2012. Динамика структуры и состояния популяций черноморско-каспийской тюльки верхневолжских водохранилищ // Матер. докл. Всерос. конф. “Бассейн Волги в XXI веке: структура и функционирование экосистем водохранилищ”. Борок: Изд-во ИБВВ РАН. С. 117−119.
Кияшко В.И., Карабанов Д.П., Яковлев В.Н., Слынько Ю.В. 2012. Становление и развитие популяции черноморско-каспийской тюльки Clupeonella cultriventris (Clupeidae) в Рыбинском водохранилище // Вопр. ихтиологии. Т. 52. № 5. С. 571−580.
Ковалев И.Н. 1958. Справочные материалы по определению веса и длины тела некоторых видов рыб Дельты Волги по нижнеглоточным и нижнечелюстным костям // Тр. Астрахан. заповедника. Вып. 4. С. 237−267.
Лазарева В.И., Соколова Е.А. 2016. Обеспеченность пищей планктофагов в Рыбинском водохранилище в условиях потепления климата: динамика и продуктивность зоопланктона // Тр. ИБВВ РАН. Вып. 74 (77). С. 77−92. https://doi.org/10.24411/0320-3557-2016-10012
Лазарева В.И., Степанова И.Э., Цветков А.И. и др. 2018. Кислородный режим водохранилищ Волги и Камы в период потепления климата: последствия для зоопланктона и зообентоса // Там же. № 81 (84). С. 47−84.https://doi.org/10.24411/0320-3557-2018-10005
Литвинов А.С., Законнова А.В. 2012. Термический режим Рыбинского водохранилища при глобальном потеплении // Метеорология и гидрология. № 9. С. 91–96.
Литвинов А.С., Рощупко В.И. 2010. Многолетние изменения гидрометеорологического режима Рыбинского водохранилища // Там же. № 7. С. 75–84.
Литвинов А.С., Пырина И.Л., Законнова А.В. и др. 2012. Изменение термического режима и продуктивности фитопланктона Рыбинского водохранилища в условиях потепления климата // Матер. докл. Всерос. конф. “Бассейн Волги в XXI веке: структура и функционирование экосистем водохранилищ”. Борок: Изд-во ИБВВ РАН. С. 167−169.
Литвинов А.С., Пырина И.Л., Законнова А.В. и др. 2014. Термический режим и продуктивность фитопланктона Рыбинского водохранилища в условиях изменения климата // Вода: химия и экология. № 12 (78). С. 108−112.
Минеева Н.М., Корнева Л.Г., Соловьева В.В. 2016. Влияние факторов среды на фотосинтетическую активность фитопланктона водохранилищ реки Волга // Биология внутр. вод. № 3. С. 47−56. https://doi.org/10.7868/S0320965216030165
Никольский Г.В. 1974. Экология рыб. М.: Высш. шк., 357 с.
Осипов В.В., Кияшко В.И. 2006. Особенности воспроизводства тюльки Clupionella cultriventris (Clupeiformes Clupeioidei)) при вселении в пресноводные водоемы // Вопр. ихтиологии. Т. 46. № 4. С 574−576.
Поддубный А.Г. 1971. Экологическая топография популяций рыб в водохранилищах. Л.: Наука, 312 с.
Попова О.А. 1979. Питание и пищевые взаимоотношения судака, окуня и ерша в водоемах разных широт // Изменчивость рыб пресноводных экосистем. М.: Наука. С. 93−112.
Правдин И.Ф. 1966. Руководство по изучению рыб. М.: Пищ. пром-сть, 376 с.
Рыбинское водохранилище и его жизнь. 1972. Л.: Наука, 364 с.
Рыбы Рыбинского водохранилища: популяционная динамика и экология. 2015. Ярославль: Филигрань, 418 с.
Сахарова Е.Г., Корнева Л.Г. 2018. Фитопланктон литорали и пелагиали Рыбинского водохранилища в годы с разными температурным и уровенным режимами // Биология внутр. вод. № 1. С. 11−18. https://doi.org/10.7868/S0320965218010023
Степанов М.В. 2011. Морфобиологическая характеристика черноморско-каспийской тюльки Clupionella cultriventris (Nordman) в Рыбинском водохранилище: Автореф. дис. … канд. биол. наук. Борок: ИБВВ РАН, 23 с.
Степанов М.В., Кияшко В.И. 2008. Роль тюльки (Clupionella cultriventris Nordman) в питании хищных рыб Рыбинского водохранилища // Биология внутр. вод. № 4. С. 86−89.
Столбунов И.А. 2007. Особенности распределения молоди рыб в прибрежной зоне Рыбинского водохранилища // Там же. № 4. С. 55−61.
Столбунов И.А. 2016. Распределение, видовой состав и численность молоди рыб в мелководной зоне Рыбинского водохранилища в разные по уровненному режиму годы // Вестн. АГТУ. Сер. Рыб. хоз-во. № 2. С. 64−69.
Фортунатова К.Р., Попова О.А. 1973. Питание и пищевые взаимоотношения хищных рыб в дельте Волги. М.: Наука, 298 с.
Юданов К.И., Калихман И.Л., Теслер В.Д. 1984. Руководство по проведению гидроакустических съемок. М.: Изд-во ВНИРО, 124 с.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии