

Вопросы ихтиологии, 2020, T. 60, № 5, стр. 611-616
Первая поимка рыбы-единорога Eumecichthys fiski (Lophotidae) в зоне Канарского апвеллинга с замечаниями по особенности географического распространения
Е. И. Кукуев 1, *, О. Ю. Краснобородько 1, В. П. Павлов 1, В. С. Сухорукова 1
1 Атлантический филиал Всероссийского научно-исследовательского института рыбного хозяйства и океанографии– АтлантНИРО
Калининград, Россия
* E-mail: efi-kukuev@yandex.ru
Поступила в редакцию 30.12.2019
После доработки 23.01.2020
Принята к публикации 30.01.2020
Аннотация
Документируется первая поимка рыбы-единорога Eumecichthys fiski у берегов Северо-Западной Африки в зоне Канарского апвеллинга. Это также первая поимка этого вида в восточной части Атлантического океана. Даётся краткое описание вида и сведения о его распространении в водах Мирового океана. Предлагается возможная схема трансокеанического дрейфа по системе течений из Северо-Западной Атлантики.
Род Eumecichthys (Lophotidae) включает единственный вид E. fiski, известный по очень редким поимкам в тропических и субтропических водах Мирового океана. Впервые вид был описан по взрослой особи от побережья Южной Африки − Калк-Бей у м. Доброй Надежды (Günther, 1890). В Атлантике E. fiski был известен только в западной части к северу от Флориды (Гольфстрим), у побережья Бразилии и Южной Африки (м. Доброй Надежды), а в Индо-Пацифике – в Мозамбикском проливе, у берегов Индии, в центральной части Тихого океана у Гавайских о-вов, в южной части Японского моря и у берегов Австралии (Линдберг, Легеза, 1965; Парин, Похильская, 1968; Masuda et al., 1984; Heemstra, 1986; Figueiredo et al., 2002; Hoese et al., 2006). Самая крупная из описанных особей достигала длины 1500 мм (Robins, Rey, 1986). В ноябре 2019 г. у побережья Северо-Западной Африки в зоне Канарского апвеллинга был пойман экземпляр E fiski. В сообщении приводится его описание и рассматривается возможный путь трансокеанического дрейфа по системе течений из Северо-Западной Атлантики.
МАТЕРИАЛ И МЕТОДИКА
В экспедиции сейнера-траулера морозильного (СТМ) “АТЛАНТНИРО” у побережья Марокко 14.11.2019 г. в координатах 22°57′ с.ш. , 17°17′ з.д. в ночное время над глубинами 680−705 м в слое 0−100 м был пойман экземпляр E. fiski общей длиной (TL) 2100 мм и массой 3.6 кг (рис. 1а). В этот же трал (№ 155) было поймано ~ 50 000 экз. миктофид (доминировал Diaphus dumerilii), 80 экз. кубоглавов Cubiceps gracilis (Nomeidae), 8 экз. гемпиловой рыбы Nealotus tripes (Gempylidae), 1 экз. Manducus maderensis (Gonostomatidae), 2 экз. Scopelosaurus sp. (Notosudidae) и рыба-меч Xiphius gladius (Xiphiidae). Экземпляр E. fiski доставлен в АтлантНИРО, где был обследован.
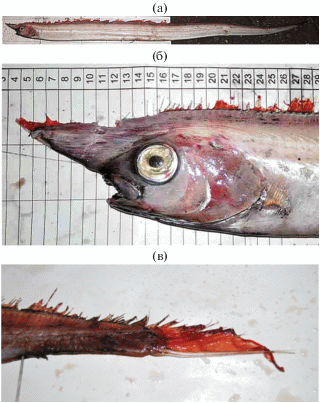
Рис. 1.
Рыба-единорог Eumecichthys fiski TL 2100 мм (14.11.2019 г., 22°57′ с.ш., 17°17′ з.д.): а − общий вид, б − голова, в − хвостовая часть.
Условия среды в районе поимки оценивали на основе данных, собранных 07−18.11.2019 г. в ходе выполнения комплексной съёмки с борта СТМ “АТЛАНТНИРО”. Гидрологические станции выполняли от поверхности до дна или до глубины 1000 м с отбором проб воды на стандартных горизонтах с помощью океанологического комплекса SeaBird Electronics (США), включающего CTD-профилограф SBE 19plusV2 и пробоотборник SBE 32. Содержание растворённого в воде кислорода определяли методом Винклера (Руководство …, 2003). Возможный маршрут переноса объекта исследования в системе течений на верхней границе мезопелагиали (~220 м) строили на основе среднемесячных модельных данных о характере циркуляции Мирового океана (зональная и меридиональная компоненты течений U и V) (CMEMS, 2019). Оценивали возможные среднемесячные траектории и дистанции переноса объекта на акватории Северной Атлантики между 0°−60° с.ш. 0°−90° з.д. Дистанцию и траекторию сноса рассчитывали в обратном порядке − от точки поимки объекта в водах Марокко к исходной точке начала его дрейфа в начале каждого месяца. За траекторию дрейфа принималась кривая, проходящая через точки акватории с наиболее высокими величинами скоростей течений (стрежни). Дистанцию сноса за месяц рассчитывали как произведение средней скорости потока в пределах выделенной траектории (в м/с) на 2592000 (число секунд в 30 сут).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Описание изученного экземпляра E. fiski. Тело сильно удлинённое, лентообразное, выступающее рыло, брюшные плавники отсутствуют (рис. 1а). Наибольшая высота тела (11 см) укладывается в длине тела 19 раз в TL, длина головы (20 см) − 10.5 раза в TL (таблица). Спинной плавник начинается колючим лучом (обломан) на самом конце рыла (рис. 1б), за ним следуют мягкие лучи, доходящие до хвостового плавника; всего в спинном плавнике 382 луча, из них 12 на верхней поверхности рыла до вертикали глаза. Спинной и хвостовой плавники практически не разделены промежутком. Нижний луч хвостового плавника утолщён в виде удлинённого шипа, его длина ~5 см (рис. 1в). Анальный плавник рудиментарный, расположен недалеко от заднего конца тела. Перед анальным плавником расположена своеобразная клоака с выходящим в неё чернильным мешком (после поимки рыбы из клоаки обильно выделялась чернильная жидкость). Диаметр глаза (3 см) укладывается 6.6 раза в длине головы (с) и составляет 15% с. Длина рыла (10 см) укладывается в длине головы 2 раза и составляет 50% с; длина рострума (8 см) − 40% с. На 1-й жаберной дуге 12 жаберных тычинок. Мелкие острые зубы имеются на верхней и нижней челюстях, на сошнике и нёбных костях. Окраска тела однотонно-серебристая без заметных вертикальных полосок. Непарные плавники красные. Наш экземпляр полностью соответствует описанию голотипа, его пропорции соответствуют возрастной изменчивости (Парин, Похильская, 1968. Таблица), но отличается более коротким рострумом и рылом, бóльшим числом лучей в спинном и анальном плавниках и отсутствием видимых поперечных полосок. Следует отметить, что у нашего экземпляра, в отличие от большинства известных, в хорошо сохранившемся хвостовом плавнике нижний луч удлинён, отделён от остальных перепонкой и имеет вид шипа (рис. 1в). По размерам (TL 2100 мм) он превосходит все когда-либо пойманные особи. Это также первая поимка вида в восточной части Атлантического океана, включая Северо-Западную Африку.
Некоторые морфометрические признаки рыбы-носорога Eumecichthys fiski из разных районов Мирового океана
| Признак | Марокко | Тихий океан | Тихий и Индийский океаны | ||||
| Наши данные | Matsubara, 1939 | Abe, 1954 | Парин, Похильская, 1968 | ||||
| TL, мм | 2100 | 881 | 752 | 678 | – | – | – |
| SL, мм | 2050 | 851 | 728 | 647 | 207 | 58 | 59 |
| В % SL | |||||||
| с | 9.7 | 10.9 | 10.8 | 10.6 | 11.0 | 12.9 | 12.8 |
| Lr | 3.9 | 5.4 | 4.8 | 4.7 | 4.0 | 3.2 | 3.3 |
| ао | 4.8 | 6.3 | 6.0 | 5.8 | 5.0 | 4.6 | 4.7 |
| о | 1.4 | 1.5 | 1.3 | 1.4 | 1.9 | 3.1 | 2.9 |
| Н | 5.36 | 3.1 | 3.1 | 3.3 | 3.8 | 6.3 | 5.9 |
| Меристические признаки | |||||||
| D | 382 | 315 | 310 | 317 | 297 | 315 | 314 |
| А | 9 | 5 | 5 | 7 | 7 | 5 | 5 |
| С | 12 | - | 13 | 12 | 12 | 12 | 12 |
| P | 13 | 13 | 14 | 14 | 14 | 14 | 14 |
Условия среды в месте поимки и возможная траектория дрейфа E. fiski. Океанологический режим морской экосистемы Канарского течения формируется сложным сочетанием водных масс, в котором наряду с водными массами местного происхождения (поверхностные и апвеллинговые, прибрежные и океанические) значительную роль играют сильно трансформированные адвективные водные массы, поступающие в район в системе подповерхностных и глубинных течений Атлантики как с севера (воды Северной Атлантики), так и с востока (средиземноморские воды) и с юга (южноатлантические и антарктические воды) (Emery, Meincke, 1986; Valdés, Déniz-González, 2015). Течения Канарской морской экосистемы (поверхностные и подповерхностные) являются частью единого многоуровнего циркуляционного процесса, развивающегося в северной части Атлантического океана. В своей северной части поток Канарского течения стыкуется с юго-восточными ветвями Северо-Атлантического течения, которое, в свою очередь, берёт начало в потоках Гольфстрима (Бурков, 1980). Таким образом, есть предпосылки для существования единого миграционного пути из западной в восточную часть Северной Атлантики.
Учитывая, что E. fiski мезопелагический вид, мы рассматриваем условия среды на нижнем горизонте траления (100 м). На рассматриваемом участке материкового склона и внешнего шельфа Марокко между м. Раймас (23°10′ с.ш.) и м. Барбас (22°20′ с.ш.) на горизонте 100 м отмечалось вторжение на шельф Марокко потока относительно тёплой (>18.2°C) высокосолёной (>36.15‰) океанической воды (рис. 2а, 2б) с высокой концентрацией растворённого кислорода (>4.0 мл/л) (рис. 2в). По своим термохалинным характеристикам воды на этом участке были ближе к трансформированным восточным североатлантическим центральным водным массам (ENACW − Eastern North Atlantic Central Water), которые характеризуются диапазоном температуры 8−18°C, солёности − 35.2−36.7‰ (Emery, Meincke, 1986). Район зарождения этой водной массы – Западно-Европейская котловина. Также существует вероятность того, что это воды из западного сектора Северной Атлантики – западные североатлантические центральные водные массы (WNACW − Western North Atlantic Central Water), принесённые сюда потоками северной части субтропического антициклонического круговорота Северной Атлантики. Характеристики WNACW близки к ENACW: температура – 7−20°C, солёность – 35.0−36.7‰. Граница между ENACW и WNACW на широте м. Барбас условно проходит по 40° з.д. (Emery, Meincke, 1986).
Рис. 2.
Условия среды на глубине 100 м в районе поимки рыбы-единорога Eumecichthys fiski, по данным океанологической съёмки СТМ “АТЛАНТНИРО” 07−18.11.2019 г.: а – температура, °С; б – солёность, ‰; в − содержание растворённого кислорода, мл/л; г − скорость и направление течений; (⚫) – место поимки, (· · ·) – изобаты.
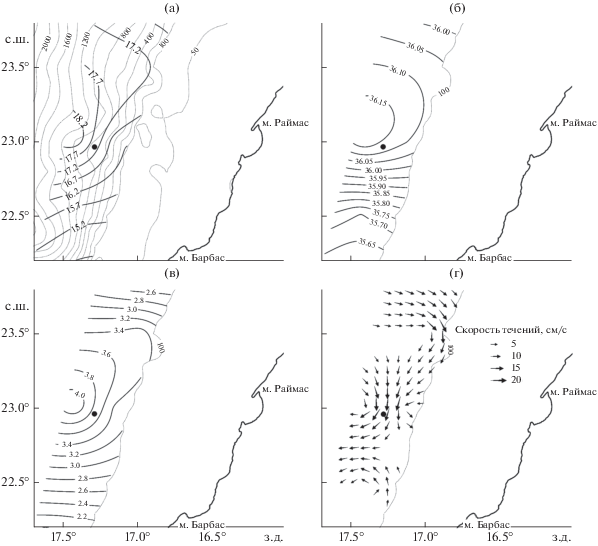
Скорости течений на участках предполагаемой траектории дрейфа объекта исследования в период с мая по ноябрь 2019 г. варьировали в пределах 0.1−1.0 м/с (рис. 2г) и в среднем составили: в мае – 0.8, в июне – 0.4, в июле−августе – 0.2, в сентябре−октябре – 0.2, в ноябре – 0.4 м/с. Исходя из этого, ежемесячная дистанция сноса объекта могла составлять от 400 до 1900 км. За 7 мес. суммарная покрытая дистанция дрейфа объекта из района действия Гольфстрима в южную часть атлантической рыболовной зоны Марокко составила ~5800 км (рис. 3а).
Рис. 3.
Предполагаемый маршрут переноса течениями в верхней части мезопелагиали (~ 220 м, по данным: CMEMS, 2019) пойманной у берегов Марокко особи из Западной в Восточную Атлантику в мае−ноябре 2019 г. (а) и места поимок рыбы-единорога Eumecichthys fiski в водах Мирового океана (б) по данным литературы (Froese, Pauly, 2019) (•) и нашим (⚫); (· · ·) − меандры течений, ($ \Rightarrow $) − генеральные направления адвекции восточных (ENACW) и западных (WNACW) центральных водных масс (пояснения см. в тексте).
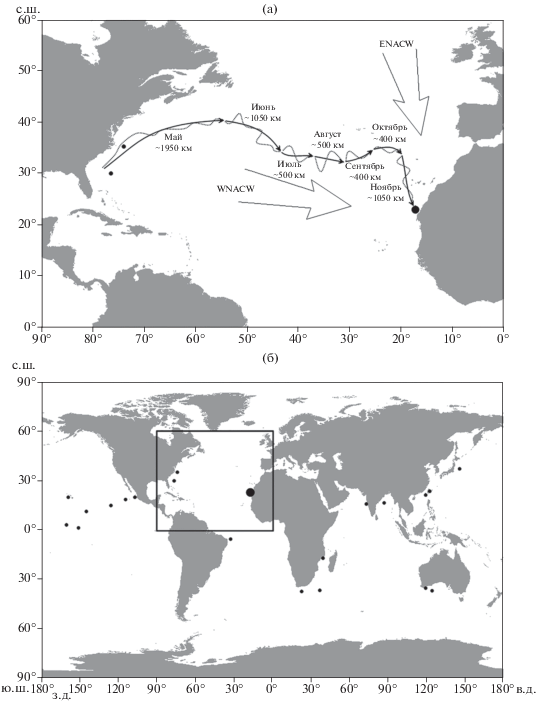
В водах Западной Атлантики E. fiski встречался в уловах у восточного побережья США в зоне действия Гольфстрима (рис. 3б) (Froese, Pauly, 2019). Будучи вовлечённой в интенсивный поток Гольфстрима, особь E. fiski теоретически могла быть вынесена в Северную Атлантику и далее по системе южных ветвей Северо-Атлантического течения могла проникнуть в открытые воды Северо-Восточной Атлантики. Существующая здесь система течений предполагает разные возможные миграционные пути.
Предполагаемая траектория дрейфа E. fiski характеризуется значительным меандрированием, особенно в районе восточнее 55° з.д., на участке юго-восточных ветвей Северо-Атлантического течения (рис. 3а). Сильное меандрирование является следствием влияния на течения в этом районе процессов вихреобразования. Фактически предполагаемая траектория дрейфа объекта пролегает по смежным перифериям мезомасштабных вихрей субтропических широт Северной Атлантики. На таких участках траектории скорости течений были невысокими (<0.2 м/с), упаковка вихрей плотная, поэтому в их пределах высока вероятность перехода переносимого течениями объекта из одного потока в другой. Поэтому предложенную траекторию переноса можно рассматривать лишь как один из возможных вариантов, но далеко не единственный. С учётом движения по меандрам общая протяжённость маршрута дрейфа может достигать 9000 тыс. км, а длительность дрейфа около года. Показателем длительности дрейфа также могут быть необычно крупные размеры пойманного экземпляра.
Судя по сопутствующим видам в улове, среди которых доминировали океанические мезопелагические тропико-субтропические виды, и гидрологической ситуации, имел место заток океанических вод в шельфово-склоновую зону Канарского апвелинга (рис. 2). Можно также предположить, что поимка в водах Марокко макропланктонного E. fiski, известного ранее только в Северо-Западной Атлантике в зоне Гольфстрима, связана с его длительным дрейфом в струях течений Северной Атлантики в направлении с запада на восток по предполагаемому маршруту (рис. 3а). Подобный дрейф (перенос) был показан на примере личинок камбал семейства Bothidae рода Bothus (Евсеенко, 2008) и атлантического гипероглифа Hyperogliphe perciformis, основной ареал которого находится в Западной Атлантике, а молодь дрейфует под куполами медуз и также входит в состав биоценоза океанского плáвника (Федоряко, 1982, 1985; Вилер, 1983; Karrer, 1986). Так как все поимки E. fiski связаны с контурами крупных течений (рис. 3б), можно также предположить, что всесветный ареал этого вида обусловлен его способностью к дрейфу в струях тёплых течений и носит тропико-субтропический характер в водах Мирового океана.
Список литературы
Бурков В.А. 1980. Общая циркуляция Мирового океана. Л.: Гидрометеоиздат, 253 с.
Вилер А. 1983. Определитель рыб морских и пресных вод северо-европейского бассейна. М.: Лег. и пищ. пром-сть, 432 с.
Евсеенко С.А. 2008. Распределение и пути дрейфовых миграций личинок трех видов камбал рода Bothus (Bothidae) в открытых водах северной Атлантики // Вопр. ихтиологии. Т. 48. № 6. С. 825–843.
Линдберг Г.У., Легеза М.И. 1965. Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Ч. 2. М.: Наука, 394 с.
Парин Н.В., Похильская Г.Н. 1968. О возрастной изменчивости редкой океанической рыбы Eumecichthys fiski (Pisces, Lophotidae) // Вопр. ихтиологии. Т. 8. Вып. 6 (53). С. 1015−1021.
Руководство по химическому анализу морских и пресных вод при экологическом мониторинге рыбохозяйственных водоемов и перспективных для промысла районов Мирового океана. 2003. М.: ВНИРО, 202 с.
Федоряко Б.И. 1982. Аннотированный список рыб, встречающихся с дрейфующим плавником // Малоизученные рыбы открытого океана. М.: Изд-во ИО АН СССР. С. 110−118.
Федоряко Б.И. 1985. Биоценоз океанского плавника: условия существования и прикладное значение: Автореф. дис. … канд. биол. наук. М.: ИО АН СССР, 24 с.
Abe T. 1954. New, rare or unrecorded fishes from Japanese waters. IV. Records of rare fishes of the families Lophotidae, Nomeidae and Icosteidae // Jpn. J. Ichthyol. V. 3. P. 90–95.
CMEMS. 2019. Copernicus Marine Environment Monitoring Service. (http://marine.copernicus.eu/services-portfolio/access-to- products/?option=om_csw&view=
details&product_id=GLOBAL_ANALYSIS_FORECAST_PHY_001_024. Version 12/2019)
Emery W.J., Meincke J. 1986. Global water masses: summary and review // Oceanol. Acta. V. 9. № 4. P. 383−391.
Figueiredo J.L., Santos A.P., Yamaguti N. et al. 2002. Peixes da zona econômica exclusiva da Região Sudeste-Sul do Brasil: Levantamento com rede de meia água. São Paulo: EDUSP, 244 p.
Froese R., Pauly D. (eds.). 2019. FishBase. World Wide Web electronic publication. (www.fishbase.org. Version 08/2019).
Günther A. 1890. Description of a new species of deep sea fish from the Cape (Lophotes fiski) // Proc. Zool. Soc. London. P. 224−247.
Heemstra P.C. 1986. Lophotidae // Smiths’ Sea fishes / Eds. Smith M.M., Heemstra P.C. Berlin: Springer-Verlag. P. 402−403.
Hoese D.F., Bray D.J., Paxton J.R., Allen G.R. 2006. Fishes // Zoological catalogue of Australia. V. 35 / Eds. Beasley O.L., Wells A. Melbourne: CSIRO Publ. P. 1473−2178.
Karrer C. 1986. The occurrence of the barrel-fish, Hyperoglyphe perciformis (Teleostei, Perciformes, Stromateoidei) in the Mediterranean Sea and off Portugal // Cybium. V. 10. № 1. P.77−83.
Masuda H., Amaoka K., Araga C. et al. 1984. The fishes of the Japanese Archipelago. V. 1. Tokyo: Tokai Univ. Press, 437 p.
Matsubara K. 1939. Studies on the deep-sea fishes of Japan. XI. On an imperfectly known ribbon-fish, Eumethichthys fiski (Günther), with a supplement to the genus Eumethichthys // Bull. Biogeogr. Soc. V. 9. P. 193–199.
Robins C.R., Ray G.C. 1986. A field guide to Atlantic coast fishes of North America. Boston: Houghton Mifflin Co., 354 p.
Valdés L., Déniz-González I. (eds.). 2015. Oceanographic and biological features in the Canary Current Large Marine Ecosystem // IOC Tech. Ser. № 115. Paris: UNESCO, 383 p.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии