

Вопросы ихтиологии, 2020, T. 60, № 6, стр. 624-635
Новый вид нетопырей рода Halicmetus из Вьетнама и новые данные по морфологии H. ruber (Ogcocephalidae)
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН – ИПЭЭ РАН
Москва, Россия
2 Институт океанологии им. П.П. Ширшова РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 23.03.2020
После доработки 10.05.2020
Принята к публикации 10.05.2020
Аннотация
Из Южно-Китайского моря описан новый вид Halicmetus odysseus, отличающийся от близких видов формой верхнего края иллициальной впадины, строением чешуйного покрова (модифицированного в щитовидные и игловидные шипы), пигментацией и другими признаками. Уточнена морфологическая характеристика вида H. ruber по результатам изучения восьми экземпляров из северо-западной части Индийского океана; отмечена неоднородность литературного материала, относимого к данному виду, на основании чего не исключается его сборная природа. Составлена таблица для определения известных видов Halicmetus из Южно-Китайского моря и Индийского океана (на восток до Сиамского залива и Больших Зондских островоов).
Нетопыри рода Halicmetus Alcock, 1891 характеризуются уплощённой головой со вздернутой ростральной частью; разрывом туловищной серии невромастов между диском и хвостом; иллициальной костью бабочковидной формы (с двумя длинными восходящими отростками); присутствием нёбных зубов; двумя жаберными дугами, несущими жаберные лепестки; наличием щитовидных шипов, по крайней мере, в хвостовом отделе; хорошо развитым “локтем” Р и в той или иной степени выраженной редукцией D (Bradbury, 1967). В настоящее время в составе этого рода описано семь видов, населяющих тропическую Индо-Вест-Пацифику от Южной Японии до Восточной Африки и Южной Австралии; таксономическая однородность некоторых из этих видов нуждается в уточнении (Bradbury, 2003; Ho et al., 2008; Ho, Last, 2018). Типовой вид рода − H. ruber Alcock, 1891− был описан из Андаманского моря (Wood-Mason, Alcock, 1891). Позже он был указан Брауером (Brauer, 1902) для восточного побережья Африки и западного берега о. Суматра, однако Хо и Ласт (Ho, Last, 2018) предполагают, что экземпляры Брауера принадлежат к виду H. marmoratus Weber, 1913, описанному из Мадурского моря (Индонезия) (Weber, 1913), а позднее указанному для вод Восточной Австралии (на юг до Тасмании) и (под вопросом) Мадагаскара (Ho, Last, 2018). В 17-м рейсе научно-исследовательского судна (НИС) “Витязь-II” нетопыри, морфологически сходные с H. marmoratus и западнотихоокеанским видом H. reticulatus Smith et Radcliffe, 1912, были собраны в северо-западной части Индийского океана (ИО РАН № 1795). Однако конспецифичность индоокеанских рыб вышеперечисленным видам вызывает сомнение, так как у них совершенно отсутствует пигментация, хорошо сохранившаяся у типовых экземпляров H. marmoratus и H. reticulatus после более чем 100 лет хранения. К сожалению, этот материал в 2007 г. был передан для изучения С.-Ч. Хо (Hsuan-Ching Ho, National Museum of Marine Biology and Aquarium, Pingtung, Taiwan) и до сих пор не возвращён, а запрос относительно его возврата остался без ответа.
Ареал и объём вида H. ruber на настоящем уровне изученности рода представляются неясными. Конспецифичность рыб из индоокеанских вод у берегов Северо-Западной Австралии, отнесённых к этому виду Хо и Ластом (Ho, Last, 2018), на мой взгляд, совершенно неочевидна, о чём будет сказано далее. Западнотихоокеанские популяции (от Южной Японии до Австралии), прежде относимые к H. ruber (Radcliffe, 1912; Masuda et al., 1984; Bradbury, 2003), предположительно относятся к отдельному, пока не описанному, виду (Ho et al., 2008; Ho, Shao, 2009). Кроме него в западной части Тихого океана известны еще три вида рода: H. reticulatus, описанный от Филиппин (Radcliffe, 1912), а затем обнаруженный в водах Южной Японии и Тайваня (Masuda et al., 1984; Yamada, 2002; Ho, Shao, 2009); H. niger Ho, Endo et Sakamaki, 2008, известный от Южной Японии, Тайваня и Северной Австралии (Ho et al., 2008); и H. drypus Ho et Last, 2018, известный только по голотипу из Кораллового моря (Ho, Last, 2018). Наконец, ещё один вид, H. westraliensis Ho et Last, 2018, был описан по массовому материалу из индоокеанских вод Австралии между 12° и 20° ю.ш. (Ho, Last, 2018).
Разбирая сохранившуюся в Институте океанологии РАН (ИО РАН) часть коллекции рыб, собранной в рейсе НИС “Одиссей” в Южно-Китайское море в 1984 г., я обнаружил необычный экземпляр Halicmetus, резко отличающийся от известных видов рода по строению чешуйного покрова и безусловно заслуживающий выделения в особый вид. Его описание составляет предмет настоящей статьи. Кроме того, по материалу из северо-западной части Индийского океана уточнена морфологическая характеристика вида H. ruber, пределы изменчивости которого по имеющимся литературным данным представляются недостаточно охарактеризованными.
МАТЕРИАЛ И МЕТОДИКА
Методика изучения и терминология соответствуют принятым для группы (Bradbury, 1967, 1980; Линдберг и др., 1997; Прокофьев, 2019). Материал по новому виду и H. ruber приведён при видовых описаниях. Помимо него были изучены голотип H. reticulatus (по фотографиям и рентгенограммам в разных проекциях) (USNM № 70271, 13°48′45″ с.ш. 120°41′51″ в.д., НИС “Альбатрос”, станция (ст.) 5118) и синтипы (5 экз.) H. marmoratus (ZMA № 101.893, 07°15′ ю.ш. 115°16′ в.д., НИС “Сибога”, ст. 12), хранящиеся в коллекциях Национального музея США (USNM, Вашингтон) и Зоологического музея Амстердама (ZMA, ныне коллекция помещена в Центр биоразнообразия г. Лейден, Нидерланды). В тексте использованы следующие сокращения: SL – стандартная длина; D, A, P, V, С – соответственно спинной, анальный, грудные, брюшные и хвостовой плавники; n – число экземпляров.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
(рис. 1, 2)

Рис. 2.
Halicmetus odysseus sp. nov., голотип, детали строения: а – голова сбоку; б – ростр и эска, вид сверху; в – эска, вид спереди; г – форма зубных пятен на сошнике (v), нёбных костях (pl) и ceratobranchialia-5 (crb5); д – краевые щитовидные шипы диска; е – шипики вентральной поверхности диска; ж – шипики дорсальной поверхности переднебоковой части диска между неврокранием и субоперкулярным шипом. Масштаб, мм: а, б – 6; в, д – 3; г – 2.5, е – 5, ж – 1.
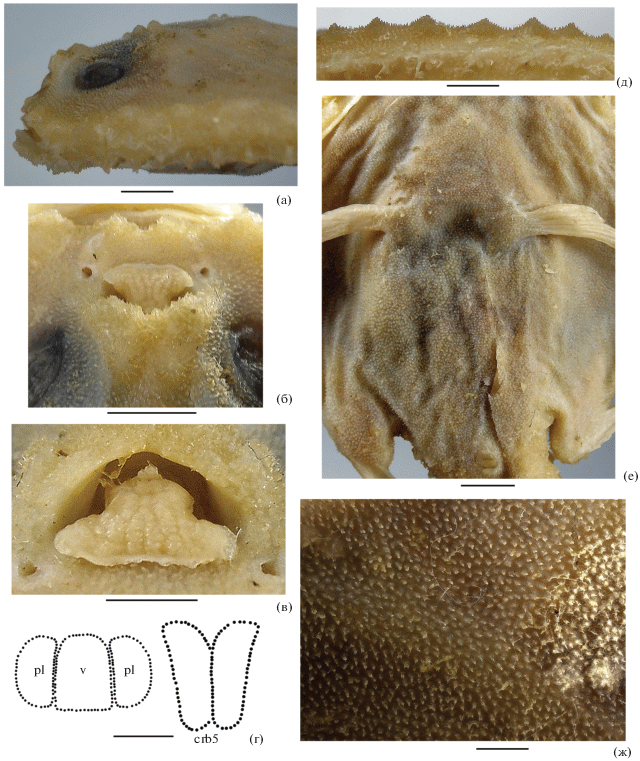
Материал. ИО РАН № 3617, голотип (рис. 1) SL 80 мм, 11°10′ с.ш. 110°02′ в.д., 680–750 м, НИС “Одиссей”, трал 52, 22.09.1984 г.
Диагноз. Вид рода Halicmetus без какой-либо пигментации тела у фиксированных особей; с коричневатым перитонеумом, несущим редкие крупные чёрные меланофоры; без щитовидных шипиков (даже мелких) на дорсальной (за пределами границ неврокрания) и вентральной поверхности диска; с низкими притуплёнными вершинами щитовидных шипов по краю диска и в хвостовом отделе; с игловидными шипиками, имеющими узкие основания с одной или двумя вершинами; с выемчатым верхним краем иллициальной впадины, в профиль не выступающим вперед; с коротким (около половины горизонтального диаметра глаза) субоперкулярным шипом, умеренной величины глазом (7.5% SL) и узким межглазничным промежутком (5.6% SL); с хвостовым отделом тела, составляющим 42.5% SL; без лучей D.
Описание. Счётные и пластические признаки приведены в таблице. Диск сужающийся кпереди, без переднебоковых углов (рис. 1), его максимальная ширина соизмерима с длиной (превышает её только в 1.02 раза); хвостовой отдел × 0.7 длины диска, 2.35 раза в SL. Нижняя челюсть выступает вперёд, рот целиком виден сверху; верхний край иллициальной впадины в профиль не выступает (рис. 2а), при взгляде сверху − V-образной формы, далеко не достигает нижнего края иллициальной впадины (рис. 2б); эска с тремя отчётливыми лопастями, из которых нижние сильно оттянуты вбок (рис. 2в). В челюстях мелкие шагреневидные зубы полосками, зубные пятна на сошнике, нёбных костях и ceratobranchialia-5 образованы такими же зубами; сошниковое зубное пятно прямоугольной формы, к его боковым сторонам прилегают нёбные зубные пятна, по форме близкие к трапециевидным, в 1.4 раза более узкие, чем сошниковое зубное пятно; зубные пятна на ceratobranchialia-5 удлинённо-прямоугольной формы с оттянутыми и расходящимися задними концами (рис. 2г). Верхнеглоточные зубные пластинки крупные, вытянутые по продольной оси. Субоперкулярный шип вдвое короче диаметра глаза, направлен латерально. Жаберных дуг 3½; жаберные лепестки хорошо развиты на второй и третьей дугах, на остальных отсутствуют; жаберные тычинки в один ряд, по три на первой и второй и четыре − на третьей дугах. D полностью отсутствует; конец прижатого А далеко (на 2/3 своей высоты) не достигает основания С; Р достигают субоперкулярного шипа, “локоть” Р хорошо обособлен; V сравнительно длинный (1.4 раза в длине Р), его лучи прогрессивно удлиняются от первого к последнему, разница в длине первого и последнего луча составляет 1.75.
Меристические и пластические признаки четырёх видов Halicmetus
| Признак | H. odysseus sp. nov., голотип | H. marmoratus, синтипы (n = 5) | H. ruber (n = 8) |
H. reticulatus, голотип* |
|---|---|---|---|---|
| SL, мм | 80 | 27.5–52 | 51–71 | 64 |
| В % SL | ||||
| Длина диска | 57.5 | 53.1–60.0 | 49.2–53.0 | 55 |
| Максимальная ширина диска | 58.8 | (61.8)69.4–75.3** | 52.2–68.2 | 74 |
| Длина хвостового отдела | 42.5 | 40.0–44.4(46.9) | 47.0–50.9 | 45 |
| Длина нейрокрания | 26.3 | 23.5–26.9 | 22.7–26.4 | 24 |
| Длина рыла | 5.0 | 4.7–5.8 | 3.4–4.5 | 4 |
| Горизонтальный диаметр глаза | 7.5 | (5.8)7.3–9.4 | 7.6–8.5 | 8 |
| Ширина костного межглазничного промежутка | 5.6 | 6.9–7.7(8.7) | 4.9–5.7 | 8 |
| Ширина рта | 17.5 | 15.3–17.3 | 13.4–15.9 | 16 |
| Ширина иллициальной впадины | 5.6 | (5.8)6.1–7.3 | 4.2–6.1 | 8 |
| Длина Р | 23.8 | 22.5–25.9(29.1) | 20.2–23.7 | 23 |
| Длина V | 17.5 | 12.3–16.5(18.1) | 13.4–14.7 | 14 |
| Длина С | 20.0 | 18.4–22.2 | 17.1–20.3 | 16 |
| Высота А | 15.0 | 11.0–13.0 | 11.9–13.6 | 11.7 |
| Меристические признаки | ||||
| D | 0 | 0–3 | 0 | 3 |
| A | 4 | 4 | 4 | 4 |
| P | 12 | 11–12 | 11–12 | 12 |
| V | 5 | 5 | 5 | 5 |
| C, главных (ветвистых верхних + нижних) | 9 (3 + 3) | 9 (3 + 3) | 9 (3 + 3) | 9 (–) |
| Число невромастов: | ||||
| в поперечном ряду на нижней челюсти | 11 | 10 | 10–11 | – |
| в щечной серии | 6 | 7 | 6–7 | – |
| в туловищной + хвостовой боковой линии | 22 (12 + 10) | 24–25 (13 + 11–12) | 22–25 (12–13 + 10–12) | – |
Супраорбитальный гребень с крупным бугорком в передней части (позади крыши иллициальной впадины) и двумя – в задней, его средняя часть спрямлена и несёт только мелкие шипики (рис. 2а). Дорсальная поверхность диска сплошь покрыта одно- и двухвершинными игловидными шипиками, такие же шипики покрывают всю поверхность хвостового отдела между щитовидными шипами. Единичные мелкие щитовидные шипы на дорсальной поверхности диска присутствуют только в заглазничной части неврокрания и совершенно отсутствуют на остальной поверхности диска. Боковой край диска несёт 12 крупных щитовидных шипов перед субоперкулярным шипом и три сильно редуцированных щитовидных шипика позади него. Хвостовой отдел с двумя параллельными рядами небольших щитовидных шипов на дорсальной и вентральной поверхностях, с парным латеральным рядом шипов, располагающихся над и под невромастами туловищной серии, и с промежуточным рядом щитовидных шипов между латеральным и дорсальным рядами. Вершины всех щитовидных шипов притуплённые (рис. 2д). Средние лучи С в ошипленных щиточках. Вентральная поверхность диска сплошь покрыта одно- и в меньшей степени двухвершинными шипиками, щитовидные шипы совершенно отсутствуют (рис. 2е). Как на дорсальной, так и на вентральной поверхностях шипики образуют сплошной, очень грубый на ощупь, покров; основания отдельных шипиков узкие (рис. 2ж).
Окраска фиксированного экземпляра однотонно-светлая, область ноздрей не пигментирована; ротожаберная полость и перитонеум плотно покрыты очень мелкими точечными буроватыми меланофорами, границы между которыми трудноразличимы невооружённым глазом; перитонемум, кроме того, в редко рассеянных крупных чёрных меланофорах; оральная сторона верхней и нижней челюстей затемнена. Плавники не окрашены, лишь дистальный край Р слегка затемнён.
Этимология. Вид назван в честь судна, в рейсе которого был пойман голотип и единственный известный в настоящее время экземпляр. Видовой эпитет является несклоняемым существительным.
Сравнение. Новый вид наиболее близок к H. marmoratus Weber, 1913 (рис. 3а–3в), но хорошо отличается от этого вида не выступающим вперед “ростром” (верхним краем иллициальной впадины), лишённым срединной выемки (рис. 2а, 2б, 3б), полным отсутствием щитовидных шипиков у заднего края диска на его дорсальной и вентральной поверхностях (рис. 1, 2е, 3а, 3в), отсутствием мраморно-пятнистого рисунка (рис. 3а) на дорсальной поверхности диска, возможно, полным отсутствием D, а также некоторыми пропорциями (таблица). Так как степень редукции D может быть подвержена индивидуальной и возрастной изменчивости, это отличие нуждается в верификации на более представительном материале и на одноразмерных рыбах, то же самое можно сказать о различиях в промерах. Однако бóльшая ширина диска и костного межглазничного промежутка у H. marmoratus по сравнению с H. odysseus sp. nov., очевидно, имеют диагностическое значение, так как у исследованных мною представителей рода эти показатели не коррелируют с величиной SL. От H. reticulatus (рис. 3г, 3д) новый вид хорошо отличается полным отсутствием щитовидных шипиков на дорсальной (за пределами неврокрания) и вентральной поверхностях диска (у H. reticulatus имеются мелкие щитовидные шипики, рис. 3д), выемчатым (против прямого) верхним краем иллициальной впадины (рис. 2б, 3г), отсутствием сетчатого рисунка (рис. 3г) на дорсальной поверхности тела, отсутствием D (всегда имеется у H. reticulatus: Ho et al., 2008. Tabl. 2), наличием на дорсальной поверхности диска двухвершинных шипиков и соотношением диаметра глаза и ширины межглазничного промежутка (равны у H. reticulatus, тогда как у нового вида межглазничный промежуток заметно ýже). Отличия от H. ruber и H. niger суммированы в определительной таблице. От известных мне только по описаниям австралийских видов H. drypus и H. westraliensis новый вид может быть отличен по отсутствию мелких щитовидных шипиков, по меньшей мере, на дорсальной поверхности диска позади неврокрания (у H. westraliensis они имеются и на вентральной стороне). Кроме того, новый вид отличается от H. westraliensis менее широким диском и межглазничным промежутком (соответственно 58.7 и 5.6% SL против 65.5–73.6 и 6.3–8.3), более широким ртом (17.5% SL против 15.5–16.8), пигментацией перитонеума (светлый с редкими меланофорами у H. westraliensis) и, возможно, отсутствием D (как правило, имеется у H. westraliensis); а от H. drypus – меньшей величиной глаза (7.5% SL против 9.3), присутствием двухвершинных шипиков (только одновершинные у H. drypus), низкими и тупыми щитовидными шипами бокового края диска и хвостового отдела тела (против высоких и островершинных у H. drypus) и, возможно, меньшим числом лучей Р (12 против 13) (Ho, Last, 2018). В первоописании H. drypus имеется расхождение в описании окраски перитонеума: в определительной таблице она указана как “greyish or black” (Ho, Last, 2018. P. 181), тогда как в диагнозе вида – “pale with many scattered melanophores (not visible from ventral surface)” (Ho, Last, 2018. P. 190). Судя по фотографии голотипа H. drypus (Ho, Last, 2018. P. 190. Fig. 5B), тёмный перитонеум просвечивает сквозь брюшную стенку тела, что является ещё одним отличием этого вида от H. odysseus sp. nov.
Рис. 3.
Halicmetus marmoratus, синтип SL 52 мм (а–в) и H. reticulatus, голотип SL 64 мм (г, д); общий вид (сверху) (а, г) и детали строения: б – ростр сбоку и сверху (масштабная линейка общая); в, д – щитовидные шипики вентральной поверхности между основаниями V и анусом; (▶) участки сохранившейся пигментации. Масштаб: б, в – 3; д – 5 мм.
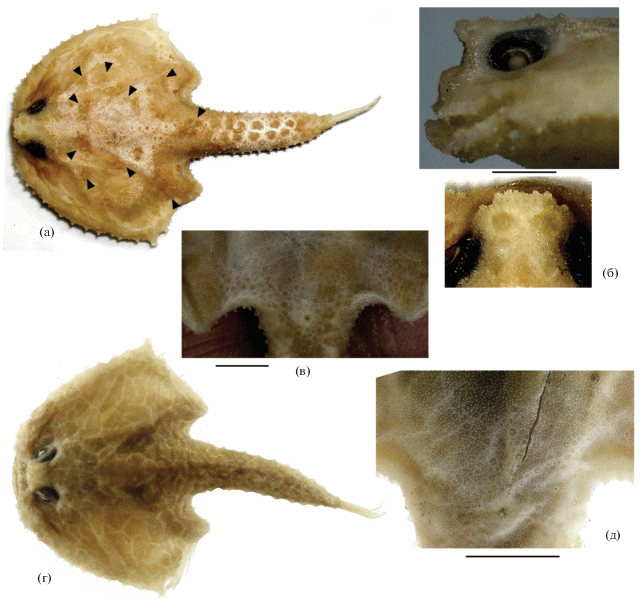
Замечания. Новый вид не может быть отождествлён с западнотихоокеанским H. cf. ruber из работ Хо с соавт. (Ho et al., 2008; Ho, Shao, 2009; Ho, Last, 2018), так как для последнего характерно наличие неправильной формы пятен на дорсальной поверхности тела и мелких щитовидных щитов (“small bucklers”: Ho, Shao, 2009. P. 303). В то же время форму, очевидно очень близкую к описываемому виду, представляют австралийские экземпляры, приводимые Хо и Ластом (Ho, Last, 2018) под названием “H. ruber”. Однако отождествлению последних с описываемым видом препятствует наличие у австралийских рыб широких оснований шипиков (на фотографии рыбы сверху (Ho, Last, 2018. Fig. 6 ) они имеют вид гранул) и прямого верхнего края иллициальной ямки.
(рис. 4, 5)

Рис. 5.
Halicmetus ruber ИО РАН № 1794, SL 71 мм, детали строения: а – голова сбоку; б – ростр и эска, вид сверху; в – форма зубных пятен на сошнике (v), нёбных костях (pl) и ceratobranchialia-5 (crb5); г – щитовидные шипики дорсальной поверхности диска (положение М-образного ряда показано пунктиром); д – краевые щитовидные шипы диска; е – щитовидные и игловидные шипики вентральной поверхности диска; ж – шипики дорсальной поверхности переднебоковой части диска между неврокранием и субоперкулярным шипом (bck – щитовидный шипик (“buckler”)). Масштаб, мм: а, г, е – 5; б, в – 2.5, д – 3, ж – 1.5.
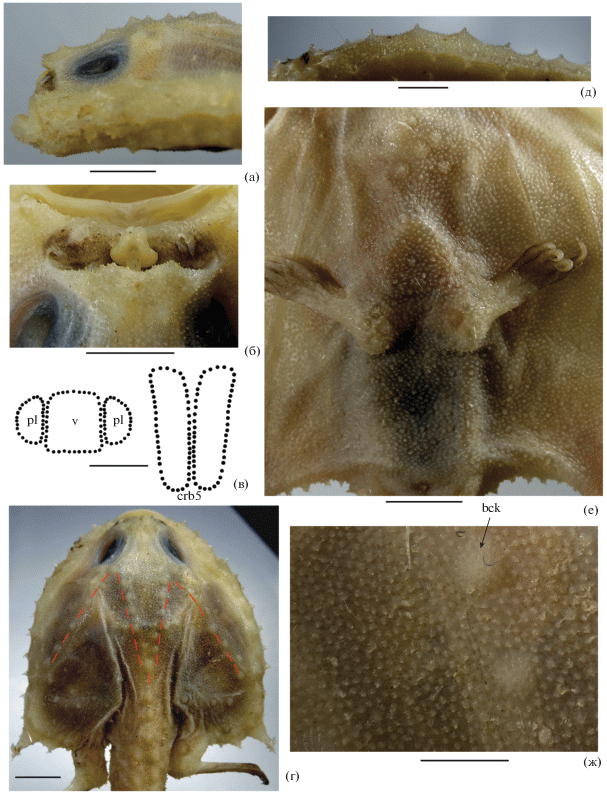
Материал. ИО РАН № 1793, 4 экз. SL 51–66 мм, 12°18′–12°14′ с.ш., 53°09′–53°06′ в.д., 375–380 м, НИС “Витязь-II”, рейс 17, ст. 2560. ИО РАН № 1794, 4 экз. SL 61–71 мм, 12°14′ с.ш., 53°06′ в.д., 420–435 м, НИС “Витязь-II”, рейс 17, ст. 2830.
Диагноз. Вид рода Halicmetus без отчётливого рисунка на теле, но с интенсивно пигментированной областью ноздрей и дистальными концами плавников у фиксированных особей, с пигментированным перитонеумом; с хорошо развитыми щитовидными шипиками на дорсальной и вентральной поверхностях диска, на дорсальной поверхности образующими М-образный ряд, на вентральной – неоднородными по величине, более крупными перед основаниями V; с высокими остроконечными вершинами щитовидных шипов по краю диска и в хвостовом отделе; с игловидными шипиками, имеющими широкие основания и одну или две вершины (как исключение, три); с дважды выемчатым (bisinuate) верхним краем иллициальной впадины, в профиль отчётливо выступающим вперёд; с длинным субоперкулярным шипом, направленным вбок и отчётливо назад; с длинным (47.0–50.9% SL) хвостовым отделом тела, умеренно крупным глазом (7.6–8.5% SL) и узким межглазничным промежутком (4.9–5.7% SL); без лучей D.
Описание. Счётные и пластические признаки приведены в таблице. Диск почти треугольной формы, без переднебоковых углов, его боковые края отчётливо сходятся к вершине рыла (рис. 4), максимальная ширина в 1.0–1.3 раза больше длины; хвостовой отдел × 0.89–1.04 длины диска, 2.0–2.1 раза в SL. Нижняя челюсть выступает вперёд, рот целиком виден сверху; верхний край иллициальной впадины в профиль выступает в виде конька (рис. 5а), при взгляде сверху дважды вогнутый (bisinuate) (рис. 5б), не достигает нижнего края иллициальной впадины; эска образует три неотчётливо подразделенные доли (рис. 5б). В челюстях мелкие шагреневидные зубы полосками, зубные пятна на сошнике, нёбных костях и ceratobranchialia-5 образованы такими же зубами; сошниковое зубное пятно прямоугольной формы, к его боковым сторонам прилегают вдвое меньшие нёбные зубные пятна, по форме близкие к трапециевидным; зубные пятна на ceratobranchialia-5 удлинённо-прямоугольной формы с оттянутыми и расходящимися задними концами (рис. 5в). Верхнеглоточные зубные пластинки крупные, овальные; пластинки правой и левой сторон плотно примыкают друг к другу. Субоперкулярный шип длинный, соизмерим с диаметром глаза, направлен назад и вбок. Жаберных дуг 3½; жаберные лепестки хорошо развиты на второй и третьей дугах, на остальных отсутствуют; жаберные тычинки в один ряд, три на первой и по четыре − на второй и третьей дугах. D полностью отсутствует у всех исследованных рыб (n = 8); конец прижатого А далеко (почти на свою высоту) не достигает основания С; Р относительно короткие, не заходят за субоперкулярный шип (иногда не достигают его); “локоть” Р хорошо обособлен; V сравнительно длинный (в 1.4–1.7 раза короче Р), его лучи прогрессивно (но не сильно) удлиняются от первого к последнему (длина первого луча содержится в длине последнего около 1.5 раза).
Супраорбитальный гребень с крупным бугорком в передней части (позади крыши иллициальной впадины) и двумя – в задней, его средняя часть полого вогнута (рис. 5а). Дорсальная поверхность диска сплошь покрыта игловидными шипиками (одно-, двух- и очень редко трёхвершинными, одновершинные преобладают), такие же шипики покрывают всю поверхность хвостового отдела между щитовидными шипами, здесь двухвершинные шипики более многочисленны. На дорсальной поверхности диска имеются умеренно крупные щитовидные шипы, образующие М-образный ряд (от преоперкулярных шипов к супраорбитальным гребням) и далее сходящиеся назад по срединно-спинной линии; помимо этого ряда на диске имеются разрозненные разноразмерные щитовидные шипы, преимущественно вдоль его заднего края (рис. 5г). Боковой край диска с 12–14 крупными щитовидными шипами перед субоперкулярным шипом и двумя или тремя мелкими (редуцированными) – позади него. Хвостовой отдел несёт по два параллельных ряда крупных щитовидных шипов на дорсальной и вентральной поверхностях, парный боковой ряд шипов, располагающихся над и под невромастами туловищной серии, и не организованные в ряды более мелкие щитовидные шипы между ними. Вершины щитовидных шипов на дорсальной поверхности диска притуплённые, образованы несколькими шипиками; щитовидные шипы бокового края диска и хвостового отдела высокие, остроконечные, одновершинные или с несколькими шипиками на вершине (рис. 5д). Средние лучи С покрыты ошипленными щиточками. Вентральная поверхность диска сплошь в мелких простых шипиках, близ среднебрюшной линии шипики более крупные, отчасти двухвершинные; вдоль заднего края диска, между анусом и основаниями V и впереди последних (достигая уровня гиоидных дуг), рассеяны некрупные, неоднородные по величине щитовидные шипы (наибольшие из них располагаются между и кпереди от оснований V) (рис. 5е). Основания шипиков дорсальной и вентральной поверхностей довольно широкие (рис. 5ж).
Окраска фиксированных рыб однотонно-светлая, но у части рыб на дорсальной поверхности диска при увеличении видны мелкие точечные буроватые меланофоры, занимающие от небольших участков до практически всей поверхности диска; ротожаберная полость светлая, в очень мелких точечных буроватых меланофорах, трудноразличимых невооружённым глазом; перитонеум более интенсивно пигментирован, меланофоры сливаются в сплошной коричневатый до черноватого фон, у некоторых рыб просвечивающий через брюшную стенку. Область вокруг и между ноздрями плотно пигментирована сливающимися меланофорами (рис. 5б). Дистальные края всех плавников черноватые или буроватые.
Сравнительные замечания. Я не имел возможности исследовать два синтипа, по которым был описан данный вид, но на оригинальных рисунках (Wood-Mason, Alcock, 1891. Pl. VIII. Figs. 1; Alcock, 1899. Pl. XIX. Fig. 5) чётко показан длинный (соизмеримый с глазом), направленный отчасти назад субоперкулярный шип. Щитовидные шипики дорсальной поверхности диска на оригинальном рисунке Олькока, очевидно, отображены не полностью, но отдельные шипики по ходу М-образного ряда, хорошо выраженного у всех исследованных мною особей, на нём показаны. Первоописание не содержит каких-либо расхождений с признаками изученных мною рыб, на основании чего я делаю вывод об их конспецифичности типовым экземплярам Олькока.
Вместе с тем следует отметить, что имеющиеся в литературе указания на последующие находки данного вида, вероятнее всего, к нему не относятся. Сомнения в конспецифичности этому виду экземпляров, описанных под названием H. ruber Брауэром (Brauer, 1902), равно как и рыб из вод западной части Тихого океана уже высказывались Хо и его соавт. (Ho et al., 2008; Ho, Shao, 2009; Ho, Last, 2018). Вместе с тем принадлежность к этому виду рыб с шельфа Северо-Западной Австралии (Ho, Last, 2018) также крайне сомнительна. У изображённой в указанной работе рыбы (Ho, Last, 2018. Fig. 6 ; отдельного описания австралийского материала не дано) совершенно отсутствуют щитовидные шипики на дорсальной поверхности диска за пределами границ неврокрания; очень короткий (менее половины диаметра глаза) субоперкулярный шип, направленный строго латерально; отсутствует пигментация в области ноздрей и на дистальных концах Р и С. Гораздо большее сходство австралийские рыбы проявляют с H. odysseus sp. nov., хотя их отождествление не бесспорно (см. выше).
Западнотихоокеанский H. cf. ruber плохо охарактеризован в литературе и я не располагаю экземплярами этой формы. Судя по информации, приводимой Хо с соавт. (Ho et al., 2008; Ho, Shao, 2009), и фотографиям в этих работах, западнотихоокеанская форма отличается от индоокеанской чётко оформленными неправильной формы участками коричневато-красного пигмента на дорсальной поверхности тела. Нужно, однако, отметить, что у изученных мною индоокеанских рыб на дорсальной поверхности диска также иногда прослеживаются участки меланофорной пигментации, хотя и неразличимые невооружённым глазом. Поскольку пигментация западнотихоокеанской формы оценивалась, по-видимому, по прижизненному состоянию, данное различие может оказаться артефактом. По морфометрическим показателям различия между западнотихоокеанской и индоокеанской формами незначительны (Ho et al., 2008. Tabl. 1), а хиатус имеется только по одному признаку (ширине рта: соответственно 13.0–14.6 и 15.4–18.2% SL). Однако мои экземпляры H. ruber по данному признаку ближе к западнотихоокеанской форме, чем к индоокеанским экземплярам, изученным Хо с соавт. (происхождение и место хранения последних в статье не указано). Вместе с тем на приводимых фотографиях чётко видно, что западнотихоокеанская форма характеризуется коротким субоперкулярным шипом, ориентированным латерально, что исключает её конспецифичность H. ruber. Качество фотографий не позволяет уверенно оценить характер щитовидных шипов дорсальной поверхности диска, но, кажется, они располагаются неупорядоченно. Из-за недостатка данных западнотихоокеанская форма H. cf. ruber в нижеприведенную определительную таблицу не включена.
Наиболее характерными признаками вида H. ruber являются максимально развитые в пределах рода щитовидные шипики на дорсальной и вентральной поверхностях диска, образующие на дорсальной поверхности чётко оформленный М-образный ряд (рис. 5г), длинный субоперкулярный шип, направленный не только вбок, но и отчётливо назад (рис. 4, 5г), и интенсивно пигментированная область ноздрей. По указанным признакам H. ruber может быть легко отличён от всех известных видов рода.
Таблица для определения видов Halicmetus Южно-Китайского моря и западной и северной частей Индийского океана (от Восточной Африки до Сиамского залива и Больших Зондских о-вов)
1(2) Окраска тела тёмная, без какого-либо рисунка; перитонеум чёрный; верхний край иллициальной впадины нависает над ртом; на дорсальной поверхности тела преобладают трехвершинные шипики..……………………………..... H. niger
2(1) Окраска тела светлая или с рисунком, перитонеум никогда не бывает сплошь чёрным; верхний край иллициальной впадины не достигает нижнего, рот целиком виден сверху; на дорсальной поверхности тела преобладают одно- или двухвершинные шипики …………………………....... 3
3(4) Хвостовой отдел тела составляет 47.0–50.9% SL; субоперкулярный шип длинный (соизмерим с продольным диаметром глаза), направлен назад и вбок; щитовидные шипики на дорсальной поверхности диска образуют М-образный ряд (от субоперкулярного шипа к затылку, а затем назад, сходясь по средней линии спины: рис. 5г); щитовидные шипики на вентральной поверхности перед основаниями V явственно крупнее шипиков между основаниями V и анусом (рис. 5е); область ноздрей интенсивно пигментирована, дистальные концы всех плавников тёмные [дорсальная поверхность диска без отчётливого рисунка; верхний край иллициальной впадины в профиль отчётливо выступает вперед; D отсутствует]………...... H. ruber
4(3) Хвостовой отдел тела составляет 40.0–46.9% SL; субоперкулярный шип короткий (около половины диаметра глаза или менее), направлен вбок перпендикулярно продольной оси тела или почти так; щитовидные шипики на дорсальной поверхности диска, если имеются, расположены неупорядоченно; щитовидные шипики на вентральной поверхности, если имеются, примерно одноразмерные; область ноздрей не пигментирована, плавники не окрашены или самое большее дистальные концы Р затемнены …………............... 5
5(6) Дорсальная поверхность диска с отчётливым рисунком (отсутствует у популяций H. cf. marmoratus из северо-западной части Индийского океана, ИО РАН № 1795); верхний край иллициальной впадины в профиль выступает вперёд в виде конька, при взгляде сверху прямой; мелкие щитовидные шипики присутствуют на дорсальной и вентральной поверхностях диска, основания игловидных шипиков широкие (как на рис. 5ж); ширина диска >60% SL (обычно 69–75%); D обычно имеется ………………………...................................... 7
6(5) Дорсальная поверхность диска без рисунка; верхний край иллициальной впадины в профиль не выступает вперёд, имеет срединную выемку; щитовидные шипики на дорсальной (кроме неврокрания) и вентральной поверхностях диска отсутствуют, основания игловидных шипиков узкие (рис. 2ж); ширина диска <60% SL; D отсутствует [межглазничный промежуток заметно меньше продольного диаметра глаза; игловидные шипики одно- и двухвершинные] ……….. H. odysseus sp. nov.
7(8) Дорсальная поверхность диска с тонким сетчатым рисунком (рис. 3г); межглазничный промежуток соизмерим с продольным диаметром глаза; игловидные шипики только одновершинные ……………………………………………… H. reticulatus
8(7) Дорсальная поверхность диска с пятнисто-мраморным рисунком (рис. 3а); межглазничный промежуток заметно меньше диаметра глаза; игловидные шипики одно- и двухвершинные….…………….………..……………….. H. marmoratus
Список литературы
Линдберг Г.У., Федоров В.В., Красюкова З.В. 1997. Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Ч. 7. Teleostomi. Osteichthyes. Actinopterygii. XXXII. Dactylopteriformes – XXXVII. Pegasiformes (CCII. Сем. Dactylopteridae – CCXIX. Сем. Pegasidae). СПб: Гидрометеоиздат, 350 с.
Прокофьев А.М. 2019. Новый вид рыб-лопат рода Halieutaea (Ogcocephalidae) из Вьетнама // Вопр. ихтиологии. Т. 59. № 6. С. 623–629. https://doi.org/10.1134/S0042875219060158
Alcock A.W. 1899. A descriptive catalogue of the Indian deep-sea fishes in the Indian Museum being a revised account of the deep-sea fishes collected by the Royal Indian Marine Survey Ship “Investigator.” Calcutta: Indian Museum, 211 p.
Bradbury M.G. 1967. The genera of batfishes // Copeia. № 2. P. 399–422.
Bradbury M.G. 1980. A revision of the fish genus Ogcocephalus with descriptions of new species from the western Atlantic Ocean (Ogcocephalidae; Lophiiformes) // Proc. Calif. Acad. Sci. Ser. 4. V. 42. № 7. P. 229–285.
Bradbury M.G. 2003. Family Ogcocephalidae Jordan 1895 batfishes // Annotated checklists of fishes. № 17. San Francisco: Calif. Acad. Sci., 17 p.
Brauer A. 1902. Diagnosen von neuen Tiefseefischen, welche von der Valdivia-Expedition gesammelt sind // Zool. Anz. Bd. 25. S. 277–298.
Ho H.-C., Last P.R. 2018. Review of the batfish genus Halicmetus Alcock, 1891 from Australian waters, with descriptions of two new species (Lophiiformes: Ogcocephalidae) // Zootaxa. V. 4508. № 2. P. 179–196.
Ho H.-C., Shao K.-T. 2009. The batfishes (Lophiiformes: Ogcocephalidae) of Taiwan, with descriptions of eight new records // J. Fish. Soc. Taiwan. V. 35. № 4. P. 289–313.
Ho H.-C., Endo H., Sakamaki K. 2008. A new species of Halicmetus (Lophiiformes: Ogcocephalidae) from the western Pacific, with comments on congeners // Zool. Stud. V. 47. № 6. P. 767–773.
Masuda H., Amaoka K., Araga C. et al. (Eds.). 1984. The fishes of the Japanese Archipelago. Tokyo: Tokai Univ. Press. Pt. 1. 437 p. + i–xxii.
Radcliffe L. 1912. New pediculate fishes from the Philippine Islands and contiguous waters // Proc. US Natl. Mus. V. 42. № 1896. P. 199–214. Pls. 16–27.
Weber M. 1913. Die Fische der Siboga-Expedition. Leiden: E.J. Brill, 710 p. + 12 pls.
Wood-Mason J., Alcock A.W. 1891. Natural history notes from H. M. Indian marine survey steamer “Investigator”, Commander R.F. Hoskyn, R.N., commanding. Series II. № 1. On the results of deep-sea dredging during the seasons 1890–1891 // Ann. Mag. Nat. Hist. Ser. 6. V. 8. № 43/44. P. 16–34, 119–138. Pls. VII, VIII.
Yamada U. 2002. 140. Ogcocephalidae batfishes // Fishes of Japan with pictorial keys to the species. V. 1 / Ed. Nakabo T. Tokyo: Tokai Univ. Press. P. 460–466.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии