

Вопросы ихтиологии, 2020, T. 60, № 6, стр. 655-664
Изменчивость сибирского тайменя Hucho taimen (Salmonidae) реки Амур
Н. С. Романов 1, *, П. Б. Михеев 2
1 Национальный научный центр морской биологии Дальневосточного отделения РАН − ННЦМБ ДВО РАН
Владивосток, Россия
2 Хабаровский филиал Всероссийского научно-исследовательского института рыбного хозяйства и океанографии − ХабаровскНИРО
Хабаровск, Россия
* E-mail: n_romanov@inbox.ru
Поступила в редакцию 12.11.2019
После доработки 05.03.2020
Принята к публикации 11.03.2020
Аннотация
Исследована морфологическая изменчивость девяти признаков в шести выборках сибирского тайменя Hucho taimen из р. Амур. Различия уровня флуктуирующей асимметрии рыб из разных притоков невелики, что говорит о сходстве условий эмбрионального и раннего постэмбрионального развития; достоверные отличия между выборками отмечены лишь в шести случаях. Максимальные колебания коэффициента вариации характерны для числа заглазничных костей и каналов сейсмосенсорной системы на 1-й заглазничной кости, а минимальные − для числа ветвистых лучей в грудных плавниках и каналов сейсмосенсорной системы на предкрышечных костях. Наибольшим значением суммы коэффициентов вариации по отдельным признакам отличается выборка из р. Бикин, а наименьшим - из р. Лимури, что положительно коррелирует с длиной и площадью бассейна этих притоков. Только по числу заглазничных костей отмечается закономерное уменьшение средних значений в выборках по мере удаления от устья р. Амур. При кластеризации по средним значениям признаков выборки образуют три кластера: 1) реки Большая Уссурка и Тырма (притоки 2-го порядка, наиболее удалённые от устья Амура), 2) р. Лимури (наименее удалённая от устья Амура), 3) реки Анюй, Хор и Бикин (географически близкие популяции), что может говорить о схожести условий раннего развития и вероятности обмена генами между последними тремя группировками.
Изменчивость - это реально существующие различия между организмами и группами организмов по степени выраженности их качественных и количественных признаков и свойств (Simpson, 1944). Исследование морфологической изменчивости представляет значительный интерес, так как позволяет оценить норму реакции того или иного вида, его адаптивные возможности. Результаты масштабных исследований морфологической изменчивости демонстрируют реальные диапазоны варьирования значений тех или признаков, а значит, позволяют внести изменения или дополнения в диагностические таблицы и таксономические описания видов. Особенно значимым представляется исследование флуктуирующей асимметрии билатеральных признаков, которая является показателем общей стабильности развития и определяется как следствие несовершенства онтогенетических процессов. Отражая особенности индивидуального развития, она требует для своего анализа не только собственно феногенетического, но и популяционного подхода (Waddington, 1940; Mather, 1953; Захаров, 1987). При необычных условиях развития уровень флуктуирующей асимметрии изменяется в сторону увеличения дисперсии, что обусловлено снижением стабильности развития (Tebb, Thoday, 1954a, 1954b; Beardmore, 1960; Valentine, Soule, 1973; Захаров, 1987; Романов, 2001). Это позволяет использовать данный показатель в мониторинговых исследованиях состояния популяций, особенно в местах антропогенного воздействия на них, по краям ареала (Mather, 1953; Захаров, 1987; Захаров, Кларк, 1993; Захаров и др., 2000), при искусственном воспроизводстве (Romanov, 1995; Романов, 2001; Михеев и др., 2014) и гибридизации (Захаров, 1987; Parsons, 1992; Романов, Скирин, 2011).
Сибирский, или обыкновенный, таймень Hucho taimen - самый крупный представитель семейства Salmonidae − обитает только в пресных водах. Его ареал простирается от рек Кама и Вятка на восток по всей Сибири, охватывает также реки Северного Китая, Монголии и Казахстана (Берг, 1948, Золотухин и др., 2000; Атлас …, 2003; Сафронов, Никифоров, 2003). Для вида характерна низкая плотность по всему ареалу (Берг, 1948; Кириллов, 1972; Dulmaa, 1999; Золотухин и др., 2000; Vander Zanden et al., 2007; Hogan, Jensen, 2013). Таймень - типичный хищник; рыба в желудках отмечается уже у сеголеток, а с 3-го года жизни он полностью переходит на питание рыбой (Леванидов, 1951; Никольский, 1956; Кириллов, 1972; Атлас …, 2003; Hogan, Jensen, 2013). По нашим данным, максимальный размер жертв составляет 47% длины хищника. Массовая половая зрелость у сибирского тайменя наступает в возрасте 6−7 лет при длине 65−83 см. Нерестится при температуре 6.0−7.5°С в зонах инфильтрации речных вод в аллювиях (зоны даунвеллинга) на плёсах с галечным грунтом и глубинами >1 м (Михеев и др., 2018). Плодовитость тайменя в наших сборах варьировала от 5.0 до 13.2 (в среднем 10.4) тыс. икринок. Продолжительность эмбрионального развития в зависимости от температуры варьирует от 28 до 38 сут (Кифа, 1974). Сибирский таймень может совершать довольно протяжённые нагульные и зимовальные миграции (Никольский, 1956; Атлас …, 2003; Gilroy et al., 2010).
Морфологическая изменчивость сибирского тайменя мало освещена в литературе. Данные по его внешней морфологии представлены в описаниях меристических и пластических признаков (Борисов, 1928; Берг, 1948; Карантонис и др., 1956; Никольский, 1956; Кириллов, 1962, 1972; Шапошникова, 1968). Первое подробное описание скелета тайменей выполнила Шапошникова (1968).
Цель работы − проанализировать флуктуирующую асимметрию, популяционную изменчивость, а также изменчивость тех признаков сибирского тайменя р. Амур, которые ранее не исследовали. Эти данные необходимы для познания морфологической изменчивости вида в пределах бассейна такой крупной реки, как Амур, что позволит уточнить таксономическое описание вида, оценить стабильность условий постэмбрионального развития и взаимосвязь между удалённостью мест обитания от русла Амура и морфологической дифференциацией проанализированных особей.
МАТЕРИАЛ И МЕТОДИКА
Материалом послужили сборы сибирского тайменя из притоков нижнего Амура: 1-го порядка − реки Лимури и Анюй, 2-го порядка − реки Хор, Бикин и Большая Уссурка; а также среднего Амура: 2-го порядка − р. Тырма (рис. 1, табл. 1).
Рис. 1.
Карта-схема района исследований и места сбора проб сибирского тайменя Hucho taimen; здесь и на рис. 2, 3: 1 - р. Лимури, 2 - р. Анюй, 3 − р. Хор, 4 - р. Бикин, 5 - р. Большая Уссурка, 6 - р. Тырма. Масштаб: 100 км.
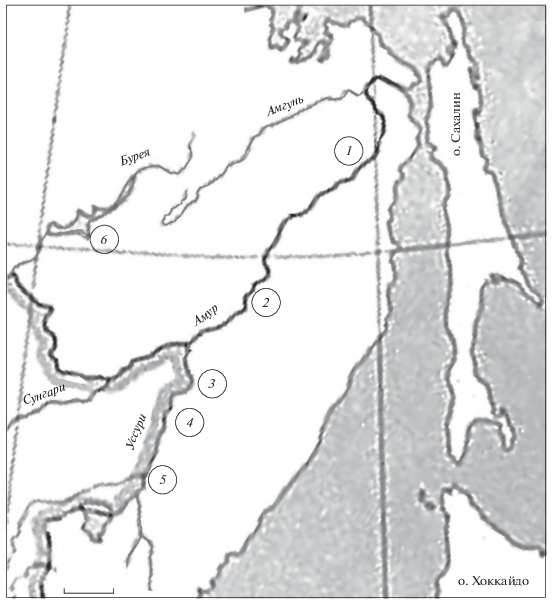
Таблица 1.
Объём собранного материала и доля асимметричных по числу признаков особей сибирского тайменя Hucho taimen из бассейна р. Амур
| Река | Число рыб, экз. |
Асимметричные по числу признаков особи, % | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| Лимури | 17 | - | 5.9 | 11.8 | 23.5 | 29.4 | 17.6 | 5.9 | 5.9 |
| Анюй | 60 | 3.3 | 11.7 | 23.3 | 16.7 | 23.3 | 16.7 | 5.0 | - |
| Хор | 28 | - | 21.4 | 17.9 | 35.6 | 17.9 | 3.6 | 3.6 | - |
| Бикин | 17 | - | 11.8 | 29.4 | 23.5 | 11.8 | 23.5 | - | - |
| Большая Уссурка | 8 | - | 12.5 | - | 50.0 | 25.0 | 12.5 | - | - |
| Тырма | 12 | - | 8.4 | 33.3 | 25.0 | 33.3 | - | - | - |
Анализировали девять билатеральных признаков: число ветвистых лучей в грудных плавниках (P); число заглазничных костей (porb), по которым проходит подглазничный канал сейсмосенсорной системы головы; число каналов 2-го порядка на 1-й заглазничной (porb-I), слёзной (lacr) и предкрышечной (pop) костях; число зубов на предчелюстной (pmx), зубной (dent), нёбной (pal) и язычной (glos) костях.
Флуктуирующую асимметрию оценивали по следующим показателям: 1) доле асимметричных рыб в выборке, 2) доле асимметричных особей по разному числу признаков от числа асимметричных рыб в выборке (число рыб, асимметричных по одному признаку, делится на число асимметричных рыб; число рыб, асимметричных по двум признакам, делится на число асимметричных рыб и т. д.); 3) доле асимметричных особей по каждому из признаков от общего числа случаев асимметрии в выборке (число случаев асимметрии в выборке представляет собой сумму случаев асимметрии по всем признакам); 4) дисперсии флуктуирующей асимметрии, которую рассчитывали по формуле, предложенной Палмером и Штробеком (Palmer, Strobeck, 1986):
Изменчивость признаков оценивали с помощью коэффициента вариации, который рассчитывали по формуле: CV = 100σ/M; его ошибку вычисляли по формуле:
Достоверность отличий средних значений (М) и коэффициентов вариации признаков определяли по величине t-критерия Стьюдента (Плохинский, 1970). Колебания значений того или иного показателя рассчитывали как разницу между минимумом и максимумом. Связь между параметрами оценивали посредством непараметрического коэффициента корреляции Спирмена (rs) (Поллард, 1982). Дендрограмму сходства строили методом полного сцепления. В качестве характеристики различий использовали Эвклидово расстояние с уровнем значимости р = 0.05. Была применена статистическая программа Systat 5.0 (Wilkinson et al., 1992a, 1992b).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Флуктуирующая асимметрия
Во всех выборках сибирского тайменя все особи были асимметричными, что можно объяснить большим числом использованных признаков. Только в р. Анюй встречались таймени асимметричные по одному, а в р. Лимури - по восьми признакам, что обусловило образование максимального числа групп (7) по доле рыб, асимметричных по числу признаков. Минимальное число таких групп (4) характерно для выборок из рек Тырма и Большая Уссурка; в четырёх выборках больше половины рыб были асимметричны по четырём и пяти признакам (табл. 1).
Доля асимметричных особей от числа асимметричных случаев варьирует в довольно широких пределах. Максимальная разница между наибольшим и наименьшим значениями этого показателя характерна для числа зубов на язычной кости (10.5%), а минимальная - для числа заглазничных костей (3.2%). Средняя доля рыб, асимметричных по каждому признаку, варьирует от 5.7% (Р) до 14.7% (pal) числа случаев асимметрии (табл. 2).
Таблица 2.
Флуктуирующая асимметрия девяти билатеральных признаков сибирского тайменя Hucho taimen из бассейна р. Амур
| Река | Доля асимметричных рыб, % числа случаев асимметрии | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| P | porb | porb-1 | lacr | pop | pmx | dent | pal | glos | |
| Лимури | 9.8 | 11.0 | 7.3 | 11.0 | 13.4 | 9.8 | 9.8 | 14.6 | 13.4 |
| Анюй | 9.6 | 9.6 | 12.0 | 11.6 | 13.2 | 10.4 | 10.8 | 13.6 | 9.2 |
| Хор | 5.7 | 9.5 | 15.2 | 9.5 | 14.3 | 13.3 | 10.5 | 12.4 | 9.5 |
| Бикин | 2.9 | 8.6 | 11.4 | 14.3 | 12.9 | 8.6 | 15.7 | 15.7 | 10.0 |
| Большая Уссурка | 5.9 | 11.8 | 17.6 | 11.8 | 14.7 | 8.8 | 11.8 | 14.7 | 2.9 |
| Тырма | 0.0 | 10.9 | 13.0 | 13.0 | 10.9 | 15.2 | 10.9 | 17.4 | 8.7 |
| В среднем | 5.7 | 10.2 | 12.8 | 11.9 | 13.2 | 11.0 | 11.6 | 14.7 | 9.0 |
Примечание. Здесь и в табл. 3−6: P − число ветвистых лучей в грудных плавниках, porb − число заглазничных костей; porb-1, lacr, pop − число каналов сейсмосенсорной системы соответственно на 1-й заглазничной, слёзной и предкрышечной костях; pmx, dent, pal, glos - число зубов на предчелюстной, зубной, нёбной и язычной костях.
Наибольшая средняя дисперсия флуктуирующей асимметрии характерна для числа заглазничных костей и числа каналов сейсмосенсорной системы на 1-й заглазничной кости, а наименьшая - для числа ветвистых лучей в грудных плавниках и зубов на зубных и нёбных костях (табл. 3). Наибольшие колебания дисперсии флуктуирующей асимметрии отмечены для числа заглазничных костей (5.42), каналов сейсмосенсорной системы на 1-й заглазничной кости (3.51) и зубов на язычной кости (2.84), а наименьшие - для числа зубов на зубной и нёбной костях (соответственно 0.15 и 0.20). Таким образом, можно сделать заключение об отрицательной корреляции между мерностью признака и дисперсией флуктуирующей асимметрии (rs = −0.97, p < 0.01). Достоверные различия по дисперсии флуктуирующей асимметрии отмечены только по числу ветвистых лучей в грудных плавниках между выборкой р. Бикин и таковыми рек Анюй (р > 0.99) и Лимури (р > 0.95), а также по числу зубов на язычной кости между выборкой из р. Большая Уссурка и рек Анюй и Хор (р > 0.95 обеих выборок), что может быть результатом небольшого объёма большинства выборок.
Таблица 3.
Дисперсия флуктуирующей асимметрии (×10−3) девяти билатеральных признаков сибирского тайменя Hucho taimen из бассейна р. Амур
| Река | Признак | Сумма | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| P | porb | porb-1 | lacr | pop | pmx | dent | pal | glos | ||
| Лимури | 1.04 | 23.40 | 13.38 | 3.95 | 2.50 | 1.93 | 0.78 | 0.94 | 4.58 | 52.50 |
| Анюй | 1.06 | 21.02 | 13.74 | 4.05 | 2.77 | 2.81 | 0.85 | 1.05 | 4.57 | 51.92 |
| Хор | 0.75 | 17.98 | 14.21 | 3.97 | 2.62 | 3.24 | 0.93 | 1.01 | 4.88 | 49.59 |
| Бикин | 0.42 | 18.21 | 16.04 | 3.87 | 3.23 | 2.48 | 0.86 | 1.01 | 4.86 | 50.98 |
| Большая Уссурка | 0.89 | 21.42 | 17.53 | 3.57 | 2.74 | 2.98 | 0.79 | 0.85 | 2.04 | 52.81 |
| Тырма | 0 | 19.90 | 14.02 | 3.69 | 2.90 | 2.20 | 0.78 | 1.01 | 4.34 | 48.84 |
| В среднем | 0.69 | 20.32 | 14.82 | 3.85 | 2.79 | 2.61 | 0.83 | 0.98 | 4.21 | 51.10 |
Если оценивать флуктуирующую асимметрию по интегрирующему показателю - сумме дисперсий по отдельным признакам, то можно отметить, что различия между исследованными выборками сибирского тайменя, а значит, и в стабильности развития невелики, что говорит о сходстве условий эмбрионального и раннего постэмбрионального развития. Это может быть следствием как низкой численности репродуктивной части популяций, что позволяет производителям сибирского тайменя выбирать для нереста места с оптимальными условиями развития, так и узкой специализации вида к условиям нереста и раннего развития, что проявляется в низкой фенотипической пластичности.
Сумма дисперсий флуктуирующей асимметрии (табл. 4) у сибирского тайменя из р. Амур по сравнению с крупночешуйной краснопёркой Tribolodon hakonensis (Романов, 2019) достоверно меньше (без учёта P и porb: р > 0.99), что может говорить либо о значительно лучших условиях развития в период эмбриогенеза и раннего постэмбриогенеза сибирского тайменя, либо о его высокой специализированности к этим условиям. Сравнение тайменя с обыкновенной малоротой корюшкой Hypomesus olidus (Романов, 2017) по двум признакам (P и porb), наоборот, демонстрирует значительно меньшую сумму дисперсий у последней (7.21, р > 0.999), что может указывать на лучшие условия её раннего развития или быть результатом более высокого темпа развития в связи с коротким эмбриогенезом. Более корректным мы считаем сравнение с тупорылым ленком Brachimystax tumensis ввиду схожести экологии размножения и раннего развития этих близкородственных видов (Глубоковский, 1995; Balakirev et al., 2013): при сравнении рыб из р. Анюй сумма дисперсий флуктуирующей асимметрии заметно меньше у сибирского тайменя (без учёта P и porb: р > 0.95). Полученные данные могут свидетельствовать о том, что условия размножения и раннего развития тайменя изученных популяций близки к оптимальным для вида.
Таблица 4.
Значения дисперсии флуктуирующей асимметрии (σd2) и коэффициента вариации (CV) сибирского тайменя Hucho taimen, тупорылого ленка Brachimystax tumensis, малоротой корюшки Hypomesus olidus и крупночешуйной краснопёрки Tribolodon hakonensis
| Признак | H. taimen | B. tumensis | H. olidus | T. hakonensis | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (наши данные) | (наши данные) | (Романов, 2017) | (Романов, 2019) | |||||||||||
| р. Анюй (60 экз.) |
р. Хор (28 экз.) |
в среднем по 6 выборкам (142 экз.) | р. Анюй (67 экз.) |
р. Хор (30 экз.) |
13 выборок (915 экз.) | 10 выборок (627 экз.) | ||||||||
| $\sigma _{d}^{2}$ | CV | $\sigma _{d}^{2}$ | CV | $\sigma _{d}^{2}$ | CV | $\sigma _{d}^{2}$ | CV | $\sigma _{d}^{2}$ | CV | $\sigma _{d}^{2}$ | CV | $\sigma _{d}^{2}$ | CV | |
| P | 1.06 | 4.67 | 0.75 | 4.20 | 0.69 | 4.40 | 0.78 | 3.74 | 0.79 | 3.64 | 1.35 | 5.19 | 0.86 | 4.78 |
| porb | 21.02 | 17.18 | 17.98 | 14.12 | 20.32 | 15.90 | 18.92 | 14.92 | 22.62 | 16.80 | 5.86 | 5.57 | 9.76 | 8.11 |
| porb-I | 13.74 | 15.85 | 14.21 | 15.47 | 14.82 | 15.60 | 21.37 | 17.22 | 20.98 | 16.91 | 41.19 | 24.15 | ||
| pop | 2.77 | 9.18 | 2.62 | 8.77 | 2.79 | 8.50 | 5.16 | 12.41 | 4.49 | 9.57 | 4.88 | 12.95 | ||
| lacr | 4.05 | 9.95 | 3.97 | 8.75 | 3.85 | 9.20 | 9.42 | 11.90 | 7.43 | 15.02 | 14.08 | 14.84 | ||
| Все признаки | 42.64 | 56.83 | 39.53 | 51.31 | 42.47 | 53.60 | 55.65 | 60.19 | 56.31 | 61.93 | 70.77 | 64.83 | ||
| Σ porb-I, pop и lacr | 20.56 | 34.98 | 20.80 | 32.99 | 21.46 | 33.36 | 36.95 | 41.53 | 32.90 | 41.50 | 60.15 | 52.74 | ||
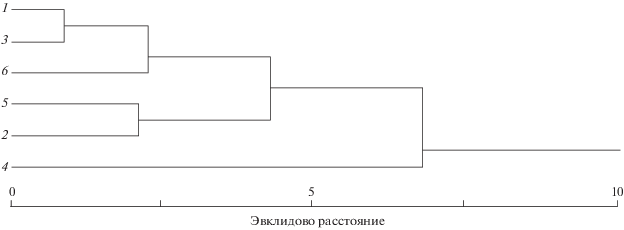
При кластеризации по сумме дисперсий по отдельным признакам выборки тайменя разбиваются на три кластера: в один входят выборки с высоким уровнем этого показателя (реки Большая Уссурка, Лимури и Анюй), в другой - с наименьшим (реки Тырма и Хор), а третий образует выборка из р. Бикин (рис. 2).

Изменчивость признаков
Максимальные диапазоны колебания коэффициента вариации характерны для числа заглазничных костей (6.44) и каналов сейсмосенсорной системы на 1-й заглазничной кости (4.78), а минимальные − для числа ветвистых лучей в грудных плавниках (1.79) и каналов сейсмосенсорной системы на предкрышечных костях (2.58) (табл. 5). Следует отметить отрицательную связь между мерностью признака и величиной коэффициента вариации (rs = -0.93, p < 0.01). Признаки с высокой мерностью (Р, dent и pal) имеют низкий уровень этого показателя, а с самой низкой мерностью (porb и porb-1) отличаются самым высоким уровнем фенотипической изменчивости. Наибольшим значением суммы коэффициентов вариации по отдельным признакам отличается выборка из р. Бикин (приток 2-го порядка), а наименьшим - из р. Лимури (приток 1-го порядка), что положительно коррелирует с длиной (rs = 0.60, p < < 0.05) и площадью бассейна этих притоков. Большая длина р. Бикин и площадь её бассейна предполагают наличие большего разнообразия условий обитания, что вынуждает рыб, в том числе и сибирского тайменя, адаптироваться к ним, это проявляется в более высоком уровне общей фенотипической изменчивости как по большинству признаков, так и по сумме коэффициентов вариации у тайменя р. Бикин.
Таблица 5.
Коэффициент вариации значений девяти билатеральных признаков сибирского тайменя Hucho taimen из бассейна р. Амур (среднее значение ± ошибка средней)
| Река | Признак | Сум-ма | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| P | porb | porb-1 | lacr | pop | pmx | dent | pal | glos | ||
| Лимури | 4.59 ± 0.56 | 14.02 ± 1.73 | 14.53 ± 1.80 | 7.90 ± 0.96 | 7.22 ± 0.88 | 8.22 ± 1.00 | 6.23 ± 0.76 | 6.14 ± 0.75 | 12.03 ± 1.48 | 80.88 |
| Анюй | 4.67 ± 0.31 | 17.18 ± 1.14 | 15.85 ± 1.05 | 9.95 ± 0.65 | 9.18 ± 0.60 | 9.30 ± 0.61 | 6.88 ± 0.45 | 6.61 ± 0.43 | 10.55 ± 0.69 | 90.17 |
| Хор | 4.20 ± 0.40 | 14.12 ± 1.36 | 15.47 ± 1.50 | 8.75 ± 0.83 | 8.77 ± 0.83 | 8.08 ± 0.77 | 6.00 ± 0.57 | 6.72 ± 0.64 | 9.59 ± 0.91 | 81.70 |
| Бикин | 5.09 ± 0.62 | 20.46 ± 2.58 | 16.78 ± 2.09 | 10.90 ± 1.34 | 9.36 ± 1.15 | 10.48 ± 1.29 | 7.18 ± 0.87 | 7.71 ± 0.94 | 8.74 ± 1.07 | 96.70 |
| Большая Уссурка | 3.30 ± 0.58 | 14.79 ± 2.67 | 17.90 ± 3.26 | 9.67 ± 1.73 | 7.52 ± 1.34 | 10.20 ± 1.82 | 9.04 ± 1.61 | 5.90 ± 1.05 | 9.80 ± 1.75 | 88.12 |
| Тырма | 4.31 ± 0.62 | 14.50 ± 2.14 | 13.12 ± 1.93 | 8.08 ± 1.17 | 8.96 ± 1.30 | 11.06 ± 1.62 | 5.75 ± 0.83 | 8.48 ± 1.23 | 9.70 ± 1.41 | 83.96 |
| В среднем | 4.40 | 15.90 | 15.60 | 9.20 | 8.50 | 9.60 | 6.90 | 6.90 | 10.10 | 86.90 |
Сопоставление сибирского тайменя с другими представителями костистых рыб показало, что от крупночешуйной краснопёрки и тупорылого ленка он отличается меньшей изменчивостью признаков, а от обыкновенной малоротой корюшки − значительно большей, особенно по числу заглазничных костей (табл. 4).
По коэффициентам вариации отдельных признаков достоверные различия между выборками из разных рек по большинству признаков отсутствуют, что может быть обусловлено малым объёмом выборок, в результате чего ошибка коэффициента вариации оказывается большой. Исключение составляют только два признака − число ветвистых лучей в грудных плавниках и число заглазничных костей: выборка из р. Большая Уссурка значимо отличается меньшим значением коэффициента вариации Р от выборок из рек Анюй (р > 0.95) и Бикин (р > 0.95); выборка из р. Бикин − большей величиной этого показателя porb от рыб из рек Лимури (р > 0.95) и Хор (р > 0.95). Кластеризация выборок сибирского тайменя по сумме коэффициентов вариации признаков демонстрирует разбиение на три кластера: в один объединяются выборки с наименьшим значением этого показателя (реки Лимури, Хор и Тырма), к нему присоединяется кластер рек Анюй и Большая Уссурка; кластер р. Бикин располагается обособленно (рис. 3).
Популяционная изменчивость
Колебания значений исследованных признаков неодинаковы (табл. 6): для числа зубов на зубной кости разница между максимальным и минимальным значением равна 6, для числа каналов на слёзной кости и зубов на нёбной − 5, для числа лучей в грудных плавниках, каналов на предкрышечной кости и зубов на предчелюстной и язычной костях − 4, для числа заглазничных костей и каналов на них − 3. Сравнение этих цифр с мерностью признаков демонстрирует положительную корреляцию, т. е. чем больше мерность признака, тем больше его колебание (rs = 0.82, p < 0.01).
Таблица 6.
Значения девяти билатеральных признаков сибирского тайменя Hucho taimen из бассейна р. Амур
| Река | Признак | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| P | porb | porb-1 | lacr | pop | pmx | dent | pal | glos | |
| Лимури | $\frac{{15.40 \pm 0.12}}{{14{\kern 1pt} - {\kern 1pt} 17}}$ | $\frac{{3.71 \pm 0.09}}{{2{\kern 1pt} - {\kern 1pt} 4}}$ | $\frac{{4.68 \pm 0.12}}{{3{\kern 1pt} - {\kern 1pt} 6}}$ | $\frac{{7.97 \pm 0.11}}{{7{\kern 1pt} - {\kern 1pt} 9}}$ | $\frac{{9.97 \pm 0.12}}{{9{\kern 1pt} - {\kern 1pt} 12}}$ | $\frac{{9.00 \pm 0.13}}{{8{\kern 1pt} - {\kern 1pt} 11}}$ | $\frac{{18.62 \pm 0.20}}{{17{\kern 1pt} - {\kern 1pt} 20}}$ | $\frac{{16.62 \pm 0.17}}{{15{\kern 1pt} - {\kern 1pt} 19}}$ | $\frac{{7.15 \pm 0.15}}{{6{\kern 1pt} - {\kern 1pt} 9}}$ |
| Анюй | $\frac{{14.78 \pm 0.06}}{{13{\kern 1pt} - {\kern 1pt} 17}}$ | $\frac{{3.55 \pm 0.06}}{{2{\kern 1pt} - {\kern 1pt} 5}}$ | $\frac{{4.29 \pm 0.06}}{{3{\kern 1pt} - {\kern 1pt} 6}}$ | $\frac{{8.24 \pm 0.07}}{{6{\kern 1pt} - {\kern 1pt} 11}}$ | $\frac{{9.48 \pm 0.08}}{{8{\kern 1pt} - {\kern 1pt} 11}}$ | $\frac{{9.25 \pm 0.08}}{{7{\kern 1pt} - {\kern 1pt} 11}}$ | $\frac{{16.72 \pm 0.10}}{{15{\kern 1pt} - {\kern 1pt} 20}}$ | $\frac{{15.88 \pm 0.10}}{{14{\kern 1pt} - {\kern 1pt} 19}}$ | $\frac{{7.11 \pm 0.07}}{{6{\kern 1pt} - {\kern 1pt} 10}}$ |
| Хор | $\frac{{15.00 \pm 0.08}}{{14{\kern 1pt} - {\kern 1pt} 16}}$ | $\frac{{3.54 \pm 0.07}}{{3{\kern 1pt} - {\kern 1pt} 4}}$ | $\frac{{4.46 \pm 0.09}}{{3{\kern 1pt} - {\kern 1pt} 6}}$ | $\frac{{8.46 \pm 0.10}}{{7{\kern 1pt} - {\kern 1pt} 10}}$ | $\frac{{9.70 \pm 0.11}}{{8{\kern 1pt} - {\kern 1pt} 12}}$ | $\frac{{8.91 \pm 0.10}}{{7{\kern 1pt} - {\kern 1pt} 10}}$ | $\frac{{17.18 \pm 0.14}}{{15{\kern 1pt} - {\kern 1pt} 19}}$ | $\frac{{16.07 \pm 0.14}}{{13{\kern 1pt} - {\kern 1pt} 18}}$ | $\frac{{6.88 \pm 0.09}}{{6{\kern 1pt} - {\kern 1pt} 10}}$ |
| Бикин | $\frac{{15.12 \pm 0.13}}{{14{\kern 1pt} - {\kern 1pt} 16}}$ | $\frac{{3.47 \pm 0.12}}{{2{\kern 1pt} - {\kern 1pt} 4}}$ | $\frac{{4.41 \pm 0.13}}{{3{\kern 1pt} - {\kern 1pt} 6}}$ | $\frac{{8.26 \pm 0.15}}{{6{\kern 1pt} - {\kern 1pt} 10}}$ | $\frac{{9.29 \pm 0.15}}{{7{\kern 1pt} - {\kern 1pt} 11}}$ | $\frac{{9.35 \pm 0.17}}{{6{\kern 1pt} - {\kern 1pt} 10}}$ | $\frac{{17.14 \pm 0.21}}{{15{\kern 1pt} - {\kern 1pt} 19}}$ | $\frac{{16.09 \pm 0.21}}{{13{\kern 1pt} - {\kern 1pt} 18}}$ | $\frac{{7.09 \pm 0.11}}{{6{\kern 1pt} - {\kern 1pt} 8}}$ |
| Большая Уссурка | $\frac{{15.13 \pm 0.13}}{{14{\kern 1pt} - {\kern 1pt} 16}}$ | $\frac{{3.38 \pm 0.13}}{{3{\kern 1pt} - {\kern 1pt} 4}}$ | $\frac{{4.19 \pm 0.19}}{{2{\kern 1pt} - {\kern 1pt} 5}}$ | $\frac{{8.38 \pm 0.02}}{{7{\kern 1pt} - {\kern 1pt} 10}}$ | $\frac{{9.31 \pm 0.18}}{{8{\kern 1pt} - {\kern 1pt} 10}}$ | $\frac{{9.31 \pm 0.24}}{{8{\kern 1pt} - {\kern 1pt} 11}}$ | $\frac{{18.25 \pm 0.41}}{{15{\kern 1pt} - {\kern 1pt} 21}}$ | $\frac{{16.94 \pm 0.25}}{{16{\kern 1pt} - {\kern 1pt} 19}}$ | $\frac{{6.94 \pm 0.17}}{{6{\kern 1pt} - {\kern 1pt} 8}}$ |
| Тырма | $\frac{{15.08 \pm 0.13}}{{14{\kern 1pt} - {\kern 1pt} 16}}$ | $\frac{{3.38 \pm 0.10}}{{3{\kern 1pt} - {\kern 1pt} 4}}$ | $\frac{{4.42 \pm 0.12}}{{3{\kern 1pt} - {\kern 1pt} 5}}$ | $\frac{{8.67 \pm 0.14}}{{7{\kern 1pt} - {\kern 1pt} 10}}$ | $\frac{{9.04 \pm 0.16}}{{8{\kern 1pt} - {\kern 1pt} 11}}$ | $\frac{{9.04 \pm 0.20}}{{6{\kern 1pt} - {\kern 1pt} 10}}$ | $\frac{{18.42 \pm 0.22}}{{17{\kern 1pt} - {\kern 1pt} 20}}$ | $\frac{{16.75 \pm 0.29}}{{14{\kern 1pt} - {\kern 1pt} 19}}$ | $\frac{{7.42 \pm 0.15}}{{6{\kern 1pt} - {\kern 1pt} 9}}$ |
Число случаев достоверных различий между выборками тайменя по средним значениям неодинаково для разных признаков: оно максимально для числа зубов на зубной (9) и нёбной костях (8) и минимально для числа заглазничных костей и каналов на них, а также зубов на предчелюстных костях (по 2). Сравнение с колебаниями значений признаков демонстрирует наличие положительной корреляции (rs = 0.85, p < 0.01).
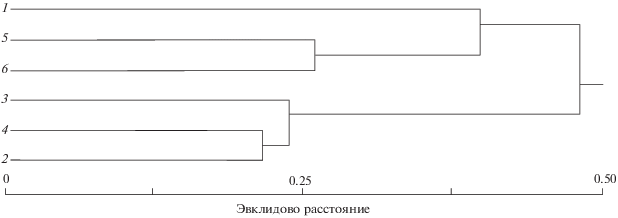
Исследованные выборки сибирского тайменя в разной степени достоверно различаются между собой по средним значениям признаков (рис. 4): Анюй - в 17 случаях, Хор и Лимури - в 14, Тырма - в 13, Бикин - в 9 и Большая Уссурка − в 8. Наиболее значимые отличия (р > 0.999) от других выборок отмечены для тайменя р. Лимури − 6 случаев.
Рис. 4.
Дендрограмма сходства выборок сибирского тайменя Hucho taimen по средним значениям исследованных признаков.
Отмеченные в литературе колебания числа ветвистых лучей в грудных плавниках (14−16: Кириллов, 1972; Атлас …, 2003) отличаются от наших результатов - 13−17 (табл. 6), что позволяет внести поправку в таксономическое описание этого вида.
Только по числу заглазничных костей фиксируется закономерное уменьшение средних значений в выборках тайменя по мере удаления от устья р. Амур. При кластеризации по средним значениям признаков выборки тайменя группируются следующим образом: один кластер образуют реки Большая Уссурка и Тырма (притоки 2-го порядка, наиболее удалённые от устья Амура), к нему присоединяется кластер р. Лимури (наименее удалённая от устья Амура), а в третий входят географически наиболее близкие выборки рек Анюй, Хор и Бикин, что может говорить о схожести условий раннего развития и вероятности обмена генами между последними тремя группировками.
ЗАКЛЮЧЕНИЕ
Различия в уровне флуктуирующей асимметрии сибирского тайменя из разных притоков р. Амур, а значит, и в стабильности развития невелики, что указывает на сходство условий эмбрионального и раннего постэмбрионального развития. Это может быть следствием как низкой численности репродуктивной части популяций, что позволяет производителям сибирского тайменя выбирать для нереста места с оптимальными условиями развития, так и узкой специализации вида к условиям нереста и раннего развития, что проявляется в низкой фенотипической пластичности. Подтверждением этого также могут служить меньшие значения суммы дисперсий флуктуирующей асимметрии сибирского тайменя по сравнению с тупорылым ленком - более многочисленным видом лососёвых, близким по экологии с тайменем; положительная связь между суммой коэффициентов вариации по отдельным признакам с длиной обследованных рек и, как следствие, с численностью в них тайменя. Нельзя не отметить отрицательную связь между мерностью признака и величиной как дисперсии флуктуирующей асимметрии, так и коэффициента вариации.
Только для числа заглазничных костей отмечается закономерное уменьшение средних значений в выборках тайменя по мере удаления от устья Амура. Выявлена положительная корреляция мерности признаков с диапазонами варьирования их значений. Обнаруженные более широкие колебания числа ветвистых лучей в грудных плавниках позволяют внести изменения в таксономическое описание сибирского тайменя. Таймени из р. Анюй в наибольшем, а из р. Большая Уссурка в наименьшем числе случаев достоверно отличаются от рыб из других рек. При кластеризации по средним значениям признаков большое значение, на наш взгляд, играет географический фактор: это наибольшая удалённость от устья Амура - реки Тырма и Большая Уссурка - и исток с западных склонов Сихтэ-Алиня - реки Анюй, Хор и Бикин.
Список литературы
Атлас пресноводных рыб России: в 2 томах. 2003. Т. 1 / Под ред. Решетникова Ю.С. М.: Наука, 379 с.
Берг Л.С. 1948. Рыбы пресных вод СССР и сопредельных стран. Ч. 1. М.; Л.: Изд-во АН СССР, 466 с.
Борисов П.Г. 1928. Рыбы реки Лены. Л.: Изд-во АН СССР, 181 с.
Глубоковский М.К. 1995. Эволюционная биология лососевых рыб. М.: Наука, 343 с.
Захаров В.М. 1987. Асимметрия животных. M.: Наука, 216 с.
Захаров В.М., Кларк Д.М. (ред.). 1993. Биотест: интегральная оценка здоровья экосистем и отдельных видов. М.: Биотест, 68 с.
Захаров В.М., Баранов А.С., Борисов В.И. и др. 2000. Здоровье среды: методика оценки. М.: Центр экол. политики России, 68 с.
Золотухин С.Ф., Семенченко А.Ю., Беляев В.А. 2000. Таймени и ленки Дальнего Востока России. Хабаровск: Хабаров. краев. тип., 128 с.
Карантонис Ф.Э., Кириллов Ф.Н., Мухамедеяров Ф.Б. 1956. Рыбы среднего течения реки Лены // Тр. ИБ СО АН СССР. Вып. 2. С. 3−130.
Кириллов Ф.Н. 1962. Ихтиофауна бассейна реки Вилюя // Тр. ИБ СО АН СССР. Вып. 8. С. 5−71.
Кириллов Ф.Н. 1972. Рыбы Якутии. М.: Наука, 359 с.
Кифа М.И. 1974. Материалы по экологии нереста ленка (Brachymystax lenok) и тайменя (Hucho taimen) в бассейне Амура // Исследования по биологии рыб и промысловой океанографии. Вып. 5. С. 105−108.
Леванидов В.Я. 1951. Питание тайменя в предгорных притоках Амура // Бюл. МОИП. Отд. биол. Т. LVI. Вып. 6. С. 31−38.
Михеев П.Б., Петренко Н.Г., Огородов С.П., Михеева О.И. 2014. Об изменчивости числа жучек стерляди Аcipenser ruthenus в ареале и аквакультуре // Рыбоводство и рыб. хоз-во. № 10. С. 25−31.
Михеев П.Б., Миронова Т.Н., Никифоров А.И. 2018. Экология нереста жилых лососевидных рыб бассейна Амура // Матер. Всерос. науч.-практ. конф. “Рациональная эксплуатация биоресурсов: проблемы и возможности в контексте целей устойчивого развития ООН”. М.: Перо. С. 270−278.
Никольский Г.В. 1956. Рыбы бассейна Амура. М.: Изд-во АН СССР, 552 с.
Плохинский Н.А. 1970. Биометрия. М.: Изд-во МГУ, 367 с.
Поллард Дж. 1982. Справочник по вычислительным методам статистики. М.: Финансы и статистика, 344 с.
Романов Н.С. 2001. Флуктуирующая асимметрия лососей заводского и естественного воспроизводства // Чтения памяти В.Я. Леванидова. Вып. 1. С. 328−335.
Романов Н.С. 2017. Морфологическая изменчивость обыкновенной малоротой корюшки Hypomesus olidus (Osmeridae) из некоторых водоемов Дальнего Востока // Вопр. ихтиологии. Т. 57. № 1. С. 15−23.https://doi.org/10.7868/S004287521701012X
Романов Н.С. 2019. Морфологическая изменчивость крупночешуйной красноперки Tribolodon hakonensis (Cyprinidae) // Там же. Т. 59. № 3. С. 258−267. https://doi.org/10.1134/S004287521902022X
Романов Н.С., Скирин В.И. 2011. Морфологическая изменчивость некоторых осетровых рыб и их искусственных гибридов // Изв. ТИНРО. Т. 165. С. 283−296.
Сафронов С.Н, Никифоров С.Н. 2003. Список рыбообразных и рыб пресных и солоноватых вод Сахалина // Вопр. ихтиологии. Т. 43. № 1. С. 42−53.
Шапошникова Г.Х. 1968. Сравнительно-морфологическое изучение тайменей (Hucho Günther) и ленка (Brachymystax Günther) // Там же. Т. 8. С. 440−464.
Balakirev E.S., Romanov N.S., Mikheev P.B., Ayala F.J. 2013. Mitochondrial DNA variation and introgression in Siberian taimen Hucho taimen // PLoS ONE. V. 8. № 8. P. 1−10. https://doi.org/10.1371/journal.pone.0071147
Beardmore J.A. 1960. Developmental stability in constant and fluctuating temperatures // Amer. J. Phys. Anthrop. V. 14. P. 411−422.
Dulmaa A. 1999. Fish and fisheries in Mongolia // Fish and fisheries at higher altitudes: Asia. FAO Fish. Tech. Pap. № 385. P. 187−236.
Gilroy D.J., Jensen O.P., Allen B.C. et al. 2010. Home range and seasonal movement of taimen, Hucho taimen, in Mongolia // Ecol. Freshwat. Fish. № 19. № 4. P. 545−554. https://doi.org/10.1111/j.1600-0633.2010.00434.x
Graham J., Freeman D.C., Emlen J. 1993. Antisymmetry, directional asymmetry and dynamic morphogenesis // Genetica. V. 89. P. 121−137. https://doi.org/10.1007/BF02424509
Hogan Z., Jensen O. 2013. Hucho taimen // The IUCN Red List of Threatened Species 2013. Article e.T188631A22605180.
Mather K. 1953. Genetical control of stability in development // Heredity. V. 7. № 3. P. 297−336.
Palmer A.R., Strobeck C. 1986. Fluctuating asymmetry: measurement, analysis, patterns // Ann. Rev. Ecol. Syst. № 17. P. 391−421.
Parsons P.A. 1992. Fluctuating asymmetry: a biological monitor of environmental and genomic stress // Heredity. V. 68. P. 361−364.
Romanov N.S. 1995. Fluctuating asymmetry in chum salmon, Oncorhynchus keta, from the Martime Province // J. Ichthyol. V. 35. № 9. P. 171−182.
Simpson G.G. 1944. Tempo and mode in evolution. N.Y.: Columbia Univ. Press, 234 p.
Sokal R.R., Rohlf F.J. 1981. Biometry. N.Y.: Freeman and Co., 859 p.
Tebb G., Thoday J.M. 1954a. Genetic effects of diurnal temperature change in laboratory populations of Drosophila melanogaster // Proc. IX Int. Congr. Genet. Caryol. V. 1. P. 789−791.
Tebb G., Thoday J.M. 1954b. Stability in development and relational balance of X-chromosomes in Drosophila melanogaster // Nature. V. 174. P. 1109−1110.
Valentine D.V., Soule M. 1973. Effect of p,p'-DDT on developmental stability of pectoral fin rays in the grunion, Leuresthes tenuis // Fish. Bull. V. 71. № 4. P. 921−926.
Vander Zanden M.J., Joppa L.N., Allen B.C. et al. 2007. Modeling spawning dates of Hucho taimen in Mongolia to establish fishery management zones // Ecol. Appl. V. 17. № 8. P. 2281-2289. https://doi.org/10.1890/06-1781.1
Waddington C.H. 1940. Organisers and genes. Cambridge: Univ. Press, 240 p.
Wilkinson L., Hill M.-A., Welna J.P., Birkenbeuel G.K. 1992a. SYSTAT for Windows: statistics, version 5. Evanston: Systat Inc., 750 p.
Wilkinson L., Hill M.-A., Miceli S. et al. 1992b. SYSTAT for Windows: graphics, version 5. Evanston: Systat Inc., 636 p.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии