

Вопросы ихтиологии, 2021, T. 61, № 1, стр. 52-58
Новые данные о распространении окуня-клювача Sebastes mentella (Sebastidae) в Гренландском море
О. В. Карамушко 1, *, Й. Ш. Христиансен 2, 3
1 Мурманский морской биологический институт Кольского научного центра РАН – ММБИ КНЦ РАН
Мурманск, Россия
2 Арктический университет Норвегии
Тромсё, Норвегия
3 Академический университет Обо
Турку, Финляндия
* E-mail: karamushko_o@mmbi.info
Поступила в редакцию 29.01.2020
После доработки 06.02.2020
Принята к публикации 06.02.2020
Аннотация
В результате ихтиологических исследований, выполненных на шельфе Северо-Восточной Гренландии в 2002–2017 гг., обнаружены новые районы обитания окуня-клювача Sebastes mentella. Показано, что распространение сеголеток не ограничивается европейским шельфом, а простирается гораздо дальше к западу, на глубоководную часть пролива Фрама. Молодь окуня-клювача постоянно встречается у Северо-Восточной Гренландии, где ветви Гольфстрима достигают её шельфа, а также далее к югу в район формирования кругового течения в восточной части Гренландского и западной части Норвежского морей. По-видимому, данная циклоническая система и холодное Гренландское течение представляют собой неотъемлемую часть нагульного и миграционного пути личинок, молоди и подросших особей окуня-клювача в район его воспроизводства на европейском шельфе, а также в море Ирмингера.
В результате многолетних исследований биологии окуня клювача Sebastes mentella сложилось мнение, что выростные районы личинок и ранней молоди его норвежско-баренцевоморской популяции находятся на европейском шельфе Норвежского и Гренландского морей между Скандинавским п-овом и архипелагом Шпицберген (Мельников, 2013). Ареал окуня-клювача на этой акватории выходит далеко за пределы шельфа и ограничивается линией от прибрежья северной части о-ва Западный Шпицберген до юго-восточной окраины шельфа о-ва Ян-Майен и далее приблизительно до м. Густав-Хольм, чуть южнее зал. Кангердлугссуак Юго-Восточной Гренландии (Мельников, 2013, 2016; Melnikov, 2016; Мельников, Глубоков, 2017).
Однако в ряде работ приведённые границы ареала окуня-клювача заметно различаются. В одних случаях показаны только основные районы распределения, которые практически не выходят за пределы европейского шельфа (Nedreaas, 1995; Saborido-Rey, Nedreaas, 2000), а в других у побережья Гренландии северная граница доходит до зал. Скорсбисунн, что приблизительно на 700 км севернее м. Густав-Хольм (Cadrin et al., 2009). В некоторых работах указывается распределение окуня-клювача ещё севернее − вдоль изобаты 500 м до траверза м. Мари-Вальдемар в Северо-Восточной Гренландии (Cadrin et al., 2010; Planque et al., 2013; Saha et al., 2017) и даже значительно обширнее (Roques et al., 2001). Имеются данные, что в Северо-Восточной Атлантике окунь-клювач распространён до Бискайского залива (Павлов, 1992; Древетняк, 1999), а также противоречивые сведения, когда в одной и той же работе карты распространения видов рода Sebastes в районе Северо-Восточной Гренландии единственного для этих мест окуня-клювача существенно различаются (Garabana, 2005).
В 2002–2017 гг. при выполнении ихтиологических исследований на шельфе Северо-Восточной Гренландии были обнаружены новые районы обитания окуня-клювача, расположенные за пределами установленной ранее области распространения вида (Мельников, 2016), которая у Гренландии простирается до границы между юго-восточным и северо-восточным прибрежьем (Møller et al., 2010). При этом некоторые исследователи не допускают возможности расширения ареала окуня-клювача в направлении Гренландии даже при потеплении климата (Hollowed et al., 2013).
В настоящей работе представлены сведения о поимках окуня-клювача на шельфе Северо-Восточной Гренландии обсуждаются вероятные пути переноса сеголеток и миграций молоди.
МАТЕРИАЛ И МЕТОДИКА
Молодь окуня-клювача собрана в 2002−2017 гг. в ходе проведения экспедиционных исследований ихтиофауны фьордов, прибрежных вод и прилегающего шельфа Восточной Гренландии по программе Арктического университета Норвегии TUNU–MAFIG (Marine fishes of northeast Greenland – diversity and adaptation). Лов вели с борта НИС “Хельмер Хансен” (RV Helmer Hanssen) мелкоячейным донным тралом Campelen-1800/96 NOFI (ячея в кутовой части 10 мм) и пелагическим тралом Harstad (ячея в кутовой части 4 мм); скорость траления соответственно 2.5 и 3.0 узла. Траления пелагическим тралом выполняли над большими глубинами прол. Фрама (2608–3182 м) в слое 20–47 (в среднем 30.7 ± 6.3) м от поверхности. Всего в девяти рейсах у Гренландии и о-ва Ян-Майен выполнено 92 траления и в уловах 14 из них обнаружен окунь-клювач.
Основная задача исследований заключалась в количественной оценке биоразнообразия рыбной части сообществ и в большинстве случаев в процессе работ определяли только массу рыб. Поэтому длину окуня-клювача рассчитывали на основе зависимости длина−масса11, описываемой степенной функцией, используя следующее уравнение:
где х – общая длина (TL), см; у – масса рыбы, г; а и b – коэффициенты. В расчётах приняты значения коэффициентов а и b уравнения степенной зависимости длина−масса, полученные непосредственно для норвежско-баренцевоморской популяции окуня-клювача (Берестовский и др., 1989). Именно эти показатели оказались максимально близки к фактическим данным, полученным нами у Северо-Восточной Гренландии. Например, расчётная длина особи TL 9.5 см и массой 8 г составила 9.3 см (табл. 1). Кроме того, по данным генетического анализа, особи, собранные в прол. Фрама и на шельфе Северо-Восточной Гренландии, принадлежат к норвежско-баренцевоморской популяции (Andrews et al., 2019).Таблица 1.
Параметры степенного уравнения, описывающего зависимость длина−масса окуня-клювача Sebastes mentella из разных районов ареала
| Район | Коэффициент | Расчётная длина особи TL массой 8 г | Источник информации | |
|---|---|---|---|---|
| а | b | |||
| Баренцево и Норвежское моря | 0.00656 | 3.184 | 9.32 | Берестовский и др., 1989 |
| Море Ирмингера | 0.00780 | 3.160 | 8.97 | Павлов, 1992 |
| Море Ирмингера, прибрежье юго-восточной Гренландии | 0.01270 | 2.977 | 8.71 | Wilhelms, 2013 |
РЕЗУЛЬТАТЫ
В 2017 г. на двух станциях над большими глубинами в средней части прол. Фрама в уловах пелагического трала были обнаружены сеголетки окуня-клювача (рис. 1, табл. 2). На расположенной южнее станции (77°13′37″ с.ш. 02°08′41″ в.д.) над глубинами 3178–3182 м при глубине траления 34.4 ± 4.5 м и температуре воды 5.4–5.6°С за 24 мин были пойманы 40379 сеголеток общей массой 23.42 кг. Их длина варьировала в пределах 36−50 (44 ± 0.3) мм, коэффициент вариации – 6.1%; модальную группу (31%) составляли особи TL 44–45 мм (рис. 2). На этой же станции была зарегистрирована молодь пикши Melanogrammus aeglefinus TL 117 мм и массой 12 г., информация о которой частично отражена в атласе морских рыб арктического региона (Mecklenburg et al., 2018). На более северной станции (78°31′49″ с.ш. 00°23′01″ з.д.) над глубиной 2608–2609 м при глубине траления 26.4 ± 5.3 м и температуре воды 0.5–1.2°С за 20 мин были пойманы 615 сеголеток общей массой 0.554 кг.
Рис. 1.
Распространение и вероятные пути переноса сеголеток и миграций молоди (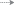 ) норвежско-баренцевоморской популяции окуня-клювача Sebastes mentella в системе тёплых (
) норвежско-баренцевоморской популяции окуня-клювача Sebastes mentella в системе тёплых (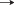 ) и холодных (
) и холодных (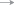 ) течений континентального склона Cеверо-Восточной Гренландии: прежние представления
об ареале вида (
) течений континентального склона Cеверо-Восточной Гренландии: прежние представления
об ареале вида (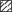 ) (Мельников, 2016) и о районах распространения сеголеток (
) (Мельников, 2016) и о районах распространения сеголеток ( ) (Мельников, 2013), а также новый район распространения молоди, открытый в 2002–2017
гг. (
) (Мельников, 2013), а также новый район распространения молоди, открытый в 2002–2017
гг. ( ), и сеголеток, открытый в 2017 г. (
), и сеголеток, открытый в 2017 г. ( ). Станции, где были обнаружены: (▲) − сеголетки в 2017 г., (⚫) – молодь в 2002–2017
гг., (◆) – особи TL ≤ 44 см в 1981−2007 гг. (Weinerroither et al., 2011).
). Станции, где были обнаружены: (▲) − сеголетки в 2017 г., (⚫) – молодь в 2002–2017
гг., (◆) – особи TL ≤ 44 см в 1981−2007 гг. (Weinerroither et al., 2011).
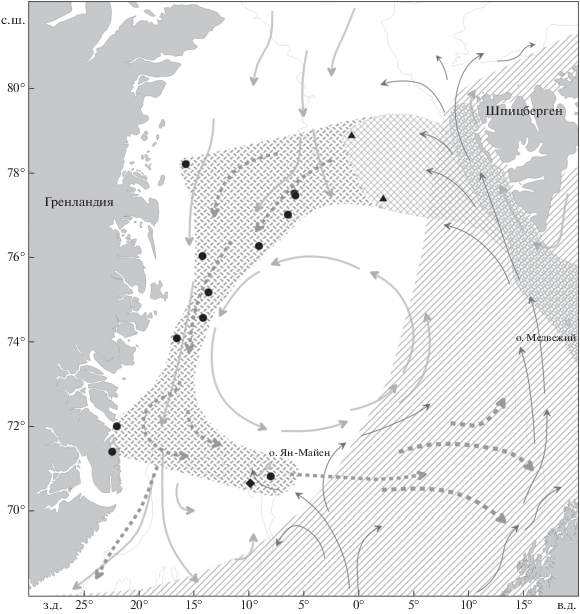
Таблица 2.
Поимки окуня-клювача Sebastes mentella у Северо-Восточной Гренландии о-ва Ян-Майен
| Район | Год | Номер станции | Координаты | Глубина лова, м | Темпера-тура, °С | Число рыб, экз. |
Масса, г | Длина (TL), см | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| с.ш. | з.д. | в.д. | общая | средняя | расчётная | факти-ческая | ||||||
| Гренландский шельф | 2002 | 1265 | 76°59′31" | 06°25′19" | 400–508 | 1.2–1.4 | 3 | 380 | 126.7 | 22.2 | ||
| 1273 | 78°10′15" | 15°44′48" | 342–360 | 0.5 | 1 | 228 | 228.0 | 26.7 | ||||
| 2003 | 900 | 73°01′15" | 16°18′47 | 500–509 | 0.8–0.9 | 1 | 288 | 288.0 | 28.7 | |||
| 2010 | 1 | 72°00′45" | 21°01′43" | 351–358 | 1.5 | 1 | 36 | 36.0 | 14.9 | |||
| 3 | 71°23′35" | 22°24′29" | 293–332 | 0.6 | 1 | 16 | 16.0 | 11.6 | ||||
| 2015 | 1312 | 74°33′02" | 14°08′19" | 294–300 | 4.8–4.9 | 7 | 86 | 12.3 | 8.0–12.5 | |||
| 1321 | 75°09′17" | 13°38′26" | 211–213 | 0.7–0.8 | 2 | 36 | 18.0 | 12.0 | ||||
| 1339 | 76°14′32" | 09°03′27" | 280–282 | 1.7–1.8 | 6 | 114 | 19.0 | 12.2–13.5 | ||||
| 1350 | 77°28′00" | 05°50′47" | 375–385 | 0.2–0.3 | 2 | 164 | 82.0 | 19.4 | ||||
| 1353 | 77°28′58" | 05°49′40" | 376–386 | 0.2–0.5 | 9 | 604 | 67.1 | 18.2 | ||||
| 2017 | 1338 | 76°00′26" | 14°07′21" | 349–351 | –0.1–0.0 | 1 | 8 | 8.0 | 9.5 | |||
| Прол. Фрама | 2017 | 1278 | 77°13′37" | 02°08′41" | 29–47 | 5.4–5.5 | 40379 | 23420 | 0.6 | 3.6–5.0 | ||
| 1381 | 78°31′49" | 00°23′01" | 20–34 | 0.5–1.1 | 615 | 554 | 0.9 | 4.7 | ||||
| Район о-ва Ян-Майен | 2003 | 902 | 70°40′17" | 08°13′32" | 303 | 1 | 66 | 66.0 | 18.1 | |||
| 1981–2007* | 1–52 | 214 | 26–44 | |||||||||
Рис. 2.
Размерный состав (TL) сеголеток окуня-клювача Sebastes mentella в прол. Фрама, сентябрь 2017 г.
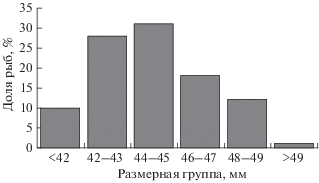
Расстояние этих двух станций до традиционного района распространения сеголеток окуня-клювача с глубиной до 1000 м составляет соответственно 203 и 212 км (110 и 115 миль). Таким образом, наши данные свидетельствуют, что область распространения сеголеток окуня-клювача не ограничивается европейским шельфом, а простирается гораздо дальше к западу.
Более крупные особи окуня-клювача впервые были обнаружены нами на шельфе Северо-Восточной Гренландии в начале октября 2002 г. В период исследований они встречались практически на всей акватории, но в 2005, 2007 и 2013 гг. в уловах отсутствовали. Самая северная точка поимки в пределах гренландского шельфа находится в районе банки Бельгика на траверзе о-ва Сторёен, самая южная – в Карсберг-фьорде. Только на самых южных станциях окунь-клювач встречался в непосредственной близости от побережья. Глубины, на которых выполнены результативные донные траления, составляли 211−509 м, температура придонных вод от −0.1 до +4.9°С (табл. 2). В уловах на шельфе Северо-Восточной Гренландии у о-ва Ян-Майен отмечены особи TL 8.0−28.7 см; максимальные размеры наблюдались только в 2002–2003 гг. (26.7–28.7 см), а в другие годы длина рыб не превышала 19.4 см. Максимальная масса составила 288 г.
ОБСУЖДЕНИЕ
Среди арктических рыб, обычно наблюдаемых у Северо-Восточной Гренландии, в 2002–2017 гг. обнаружены и виды, ареал которых по современным представлениям ограничивается склоном европейского шельфа и ближайшими сопредельными районами: окунь-клювач, мойва Mallotus villosus, треска Gadus morhua, синяя зубатка Anarhichas denticulatus. Из них наиболее часто встречался окунь-клювач, хотя в прибрежных и сопредельных водах Северо-Восточной Гренландии этот вид, как, впрочем, и другие представители рода Sebastes, ранее не отмечался (Møller et al., 2010; Мельников, 2016; Melnikov, 2016). Судя по материалам, собранным нами в этом районе, окунь-клювач, по-видимому, является здесь хотя и не многочисленным, но постоянно присутствующим видом.
Обнаружение сеголеток окуня-клювача в средней части прол. Фрама также не соответствует традиционным представлениям о районах их распространения. Предшествующими исследованиями установлено, что область расселения сеголеток приурочена к шельфовой зоне и прилегающему району с глубинами ≤1000 м (Melnikov, 2016; Saha et al., 2017), а сама область расселения молоди TL 5–20 см шире, чем у рыб старшего возраста (Древетняк, 1999). Очевидно, сеголетки окуня-клювача оказались в прол. Фрама вдали от шельфовой зоны благодаря одной из ветвей Западно-Шпицбергенского течения, отклоняющейся к западу в направлении Гренландии. Удивительно, что плотность их распределения в приповерхностном слое была чрезвычайно высока по сравнению с приведёнными в литературе данными. Так, например, в 1976−1992 гг. концентрация сеголеток норвежско-баренцевоморской популяции окуня-клювача не превышала 107 экз/ч траления (Drevetnyak, 1995), тогда как в 2017 г. в прол. Фрама она составляла 1.8−100.9 тыс. экз/ч траления. У нас нет данных, до какой глубины от поверхности распределялись сеголетки, хотя известно, что особи TL 28–67 мм встречаются в слое 0–100 м (Павлов, 1992). Сеголетки окуня-клювача питаются относительно мелкими ракообразными (Dolgov, Drevetnyak, 1995), для чего используют зрение, поэтому наиболее плотные концентрации молоди, возможно, были приурочены к верхнему 50-метровому слою.
В результате дальнейшего пассивного переноса в западном направлении молодь достигает гренландского шельфа, где через определённое время опускается в придонные слои. Массовое оседание происходит на 2-м году жизни или несколько позже (Drevetnyak, 1995). То, что сеголетки окуня-клювача достигают гренландского шельфа, теперь не вызывает сомнений, поскольку в течение многих лет молодь встречалась в этом районе (табл. 2, рис. 1) и, по данным генетического анализа, она относится к норвежско-баренцевоморской популяции (Andrews et al., 2019). Молодь окуня-клювача (за исключением 2 экз.) была заметно меньшего размера, чем особи в начале созревания, длина которых в пределах ареала составляет 26–30 см (Древетняк, 1999; Drevetnyak et al., 2004; Мельников и др., 2010, Филина и др., 2017). В то же время рано созревающие особи могут массово (50%) становиться половозрелыми уже при TL 29 см (Мельников, Глубоков, 2017).
Молодь окуня-клювача норвежско-баренцевоморской популяции встречается на глубине до 700 м (Древетняк, 1999), поэтому глубины, на которых были отловлены особи у Северо-Восточной Гренландии (211–509 м), соответствуют общему представлению о батиметрическом распространении вида на данном этапе онтогенеза. А вот минимальная температура, при которой встречалась молодь в районе исследований, оказалась отрицательной (−0.1°С), тогда как в Норвежском и Баренцевом морях этот показатель не бывает ниже 0.5°С (Drevetnyak et al., 2004).
Перенос сеголеток к северо-восточному шельфу и склону Гренландии, а также последующее распределение молоди окуня-клювача на обширной акватории данного района и к югу от него позволяют предположить, что дальнейшая миграция особей может проходить как минимум в двух направлениях. Часть особей может продолжать движение вдоль Восточной Гренландии и через Датский пролив попадать в море Ирмингера, а часть рыб по круговому течению к северу от о-ва Ян-Майен возвращаться на шельф и континентальные склоны Норвежского и Баренцева морей. Первое направление миграции подтверждается генетическим сходством окуня-клювача из районов северной части Норвежского моря и из пелагической группы моря Ирмингера (Stefansson et al., 2009; Строганов и др., 2009; Мельников, 2013), а это, несомненно, указывает на возможность такого проникновения. Относительно второго направления мы пока можем лишь предполагать, что именно так и происходит; для получения достоверной подтверждающей информации необходимо провести сравнительные генетические исследования окуня-клювача из районов шельфа о-ва Ян-Майен и из верхних слоёв пелагиали Норвежской и Лофотенской котловин.
Подобные переносы и миграции не являются исключительным событием для данного вида: перенос молоди далеко за пределы области обитания взрослой части популяции наблюдается и в Северо-Западной Атлантике, где по достижении половой зрелости особи постепенно возвращаются на акваторию воспроизводства (Мельников, 2005).
Следует также отметить, что кроме окуня-клювача прибрежные районы Гренландии могут играть определённую роль в формировании поколений и других промысловых видов рыб баренцевоморских популяций. Так, у Гренландии в 2013 г. была отмечена молодь синей зубатки (Карамушко и др., 2017), атлантической трески и мойвы (Christiansen et al., 2016), а также молодь пикши, обнаруженная нами в сентябре 2017 г. над глубоководными районами в 200 км к западу от границы европейского шельфа. Ранее считалось, что глубоководный прол. Фрама между архипелагом Шпицберген и Гренландией является барьером в распространении рыб (Briggs, 1974), но данные, полученные на шельфе и верхнем участке континентального склона, указывают на то, что в действительности проникновение окуня-клювача и других видов к Северо-Восточной Гренландии имеет место и, возможно, данный район является традиционной акваторией при формировании поколений.
Список литературы
Берестовский Е.Г., Анисимова Н.А., Денисенко С.Г. и др. 1989. Зависимость между размерами и массой тела некоторых беспозвоночных и рыб северо-восточной Атлантики. Апатиты: Изд-во КНЦ РАН, 23 с.
Древетняк К.В. 1999. Биология и промысел окуня-клювача норвежско-баренцевоморской популяции: Автореф. дис. … канд. биол. наук. М.: ВНИРО, 24 с.
Карамушко О.В., Биркъедал И., Христиансен Й.Ш., Лингхаммар А. 2017. Первая поимка молоди синей зубатки Anarhichas denticulatus на континентальном склоне Северо-Восточной Гренландии // Вопр. ихтиологии. Т. 57. № 4. С. 475–480. https://doi.org/10.7868/S0042875217040051
Мельников С.П. 2005. Биологические основы регулирования промысла окуня-клювача (Sebastes mentella Travin) в пелагиали Северной Атлантики: Автореф. дис. … канд. биол. наук. М.: ВНИРО, 25 с.
Мельников С.П. 2013. Окунь-клювач Sebastes mentella Атлантического и Северного ледовитого океанов (популяционная структура, биология, промысел): Автореф. дис. … докт. биол. наук. М.: ВНИРО, 49 с.
Мельников С.П. 2016. О дифференциации запасов окуня-клювача Sebastes mentella Атлантического и Северного ледовитого океанов // Вопр. рыболовства. Т. 16. № 2. С. 189–202.
Мельников С.П., Глубоков А.И. 2017. Направления внутривидовой дифференциации окуня-клювача Sebastes mentella (Scorpaenidae) Атлантического океана и сопредельных вод // Журн. общ. биологии. Т. 78. № 6. С. 66–75.
Мельников С.П., Строганов А.Н., Шибанов В.Н. 2010. Популяционно-генетические исследования окуня-клювача Sebastes mentella моря Ирмингера // Тр. ВНИРО. Т. 149. С. 237–248.
Павлов А.И. 1992. Биология, состояние запаса и промысел окуня-клювача (Sebastes mentella Travin) в море Ирмингера: Автореф. дис. … канд. биол. наук. М.: ВНИРО, 23 с.
Строганов А.Н., Лепесевич Ю.М., Мельников С.П. 2009. Биолого-генетическая характеристика окуня-клювача Sebastes mentella (Scorpenidae) открытой части Норвежского моря // Вопр. ихтиологии. Т. 49. № 3. С. 333–340.
Филина Е.А., Рольский А.Ю., Бакай Ю.И. и др. 2017. Особенности репродуктивного цикла самок окуня-клювача Sebastes mentella (Sebastidae) // Там же. Т. 57. № 1. С. 89–95. .https://doi.org/10.7868/S0042875216060047
Andrews A.J., Christiansen J.S., Bhat S. et al. 2019. Boreal marine fauna from the Barents Sea disperse to Arctic Northeast Greenland // Sci. Rept. V. 9. P. 1–8. https://doi.org/10.1038/s41598-019-42097-x
Briggs J.C. 1974. Marine zoogeography. N.Y.: McGraw-Hill, 475 p.
Cadrin S., Bernreuther M., Danielsdottir A.K. et al. 2009. Mechanisms and consequences of life cycle diversity of beaked redfish, Sebastes mentella // ICES CM 2009/H:01. Copenhagen: ICES Headquarters, 36 p.
Cadrin S.X., Bernreuther M., Danielsdottir A.K. et al. 2010. Population structure of beaked redfish, Sebastes mentella: evidence of divergence associated with different habitats // ICES J. Mar. Sci. V. 67. № 8. P. 1617–1630. https://doi.org/10.1093/icesjms/fsq046
Christiansen J.S., Bonsdorff E., Birkjedal I. et al. 2016. Novel biodiversity baselines outpace models of fish distribution in Arctic waters // Sci. Nature. V. 103. № 1–2. P. 103–108. https://doi.org/10.1007/s00114-016-1332-9
Dolgov A.V., Drevetnyak K.V. 1995. Feeding of young Sebastes mentella Travin in the Barents and Norwegian seas // Proc. 6-th IMR−PINRO Symp. Bergen, Norway. P. 129–132.
Drevetnyak K.V. 1995. Distribution and abundance of young Sebastes mentella in the Barents and Norwegian seas in 1991 and 1992 // Ibid. P. 219–228.
Drevetnyak K.V., Nedreaas K.H., Kluev A. 2004. Spatial migration pattern of deep-sea redfish (Sebastes mentella Travin) of the Norwegian-Barents population from survey data // ICES Ann. Sci. Conf. CM 2004/K:31. Copenhagen: ICES Headquarters, 28 p.
Garabana D. 2005. The genus Sebastes Cuvier, 1829 (Pisces, Scorpaenidae) in the North Atlantic: species and stock discrimination using traditional and geometric morphometrics: Ph. D. Thesis Univ. Vigo: Univ. Vigo, 306 p.
Hollowed A.B., Planque B., Loeng H. 2013. Potential movement of fish and shellfish stocks from sub-Arctic to the Arctic Ocean // Fish. Oceanogr. V. 22. № 5. P. 355–370. https://doi.org/10.1111/fog.12027
Mecklenburg C.W., Lynghammar A., Johannesen E. et al. 2018. Marine fishes of the Arctic Region. V. 1. Akureyri, Iceland: CAFF, 454 p.
Melnikov S.P. 2016. Intraspecies structure of beaked redfish Sebastes mentella of the Atlantic and Arctic oceans // J. Ichthyol. V. 56. № 1. P. 52–71. .https://doi.org/10.1134/S0032945216010082
Møller P.R., Nielsen J.G., Knudsen S.W. et al. 2010. A checklist of the fish fauna of Greenland waters // Zootaxa. Ser. Monograph. V. 2378. № 1. 84 p. https://doi.org/10.11646/zootaxa.2378.1
Nedreaas K.H. 1995. Short note on abundance of Sebastes mentella in the 0-group and youngfish surveys as indicator of recruitment overfishing // Proc. 6-th IMR−PINRO Symp. Bergen, Norway. P. 215–218.
Planque B., Kristinsson K., Astakhov A. et al. 2013. Monitoring beaked redfish (Sebastes mentella) in the North Atlantic, current challenges and future prospects // Aquat. Liv. Resour. V. 26. № 4. P. 293–306. https://doi.org/10.1051/alr/2013062
Roques S., Sevigny J.-M., Bernatchez L. 2001. Evidence for broadscale introgressive hybridization between two redfish (genus Sebastes) in the North-west Atlantic: a rare marine example // Mol. Ecol. V. 10. № 1. P. 149–165. https://doi.org/10.1046/j.1365-294X.2001.01195.x
Saborido-Rey F., Nedreaas K.H. 2000. Geographic variation of Sebastes mentella in the Northeast Arctic derived from morphometric approach // ICES J. Mar. Sci. V. 57. № 4. P. 965–975. https://doi.org/10.1006/jmsc.2000.0703
Saha A., Johansen T., Hedeholm R. et al. 2017. Geographic extent of introgression in Sebastes mentella and its effect on genetic population structure // Evol. Appl. V. 10. № 1. P. 77–90. https://doi.org/10.1111/eva.12429
Stefansson M.O., Reinert J., Sigurðsson T. et al. 2009. Depth as a potential driver of genetic structure of Sebastes mentella across the North Atlantic Ocean // ICES J. Mar. Sci. V. 66. № 4. P. 680–690. https://doi.org/10.1093/icesjms/fsp059
Weinerroither R.M., Nedreaas K.H., Uiblein F. et al. 2011. The marine fishes of Jan Mayen Island, NE Atlantic – past and present // Mar. Biodiv. V. 41. № 3. P. 395–411. https://doi.org/10.1007/s12526-010-0055-y
Wilhelms I. 2013. Atlas of length-weight relationships of 93 fish and crustacean species from the North Sea and the North-East Atlantic // Thünen Working Paper. № 12. 551 p. https://doi.org/10.3220/WP_12_2013
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии