

Вопросы ихтиологии, 2021, T. 61, № 1, стр. 46-51
О нахождении двух редких видов трахиноидных рыб – Chrionema furunoi (Percophidae) и Parapercis compressa (Pinguipedidae) – в Южно-Китайском море
1 Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
2 Институт океанологии РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 04.06.2020
После доработки 10.07.2020
Принята к публикации 10.07.2020
Аннотация
Перкофовая рыба Chrionema furunoi, ранее известная из вод Южной Японии и Северо-Восточного Тайваня, впервые обнаружена в Южно-Китайском море у берегов Вьетнама. Редкий вид параперковых рыб, Parapercis compressa, ранее известный только по голотипу от о. Сумбава (Малые Зондские о-ва), найден в Южно-Китайском море между о. Калимантан и полуостровом Малакка. Приведены детальные морфологические описания и иллюстрации новых экземпляров. Составлена таблица для определения видов рода Chrionema.
Настоящее сообщение посвящено новым нахождениям двух редких видов трахиноидных рыб из семейств Percophidae и Pinguipedidae в Западной тропической Пацифике. Chrionema furunoi (Percophidae) до сих пор считался эндемиком вод Южной Японии и прилежащего северо-восточного побережья Тайваня (Nakabo, 2002; https://www.tbn.org.tw. Version 05/2020), тогда как Parapercis compressa (Pinguipedidae) был известен только по голотипу, пойманному в водах Индонезии (Randall, 2008). Новые находки этих видов позволяют не только существенно расширить представления об ареалах этих видов, но и уточнить пределы их морфологической изменчивости, что особенно актуально для P. compressa, описанного по единственному экземпляру.
МАТЕРИАЛ И МЕТОДИКА
Исследованный материал хранится в коллекции Института океанологии РАН, Москва (ИО РАН). Методика изучения и терминология соответствуют общепринятым для группы (Cantwell, 1964; Iwamoto, Staiger, 1976; Randall, 2008). Последний, разделённый до основания луч считался как 1½. Маленькие чешуи в основании вертикальных плавников считались как 0.5. Подсчёт предорсальных чешуй начинался от заднего края черепа, в счёт включена последняя раздвоенная чешуйка перед основанием первого луча ID. Счётные признаки, различающиеся на разных сторонах тела одной и той же рыбы, разделены знаком “/”. В тексте использованы следующие сокращения: ID, IID, A, P, V, C – соответственно первый и второй спинные, анальный, грудные, брюшные и хвостовой плавники; LL, LL-D, LL-A, preD, crcp – соответственно число чешуй в боковой линии, между боковой линией и началом IID и А, предорсальных чешуй и чешуй вокруг хвостового стебля; sp. br, pbr – соответственно число жаберных тычинок в наружном ряду на первой жаберной дуге и лепестков ложножабры; SL – стандартная длина; lc, wc – соответственно длина и максимальная высота головы; H, h – соответственно наибольшая и наименьшая высота тела; w – ширина тела на уровне оснований Р; lcp – длина хвостового стебля; aID, aIID, aV, aA, iD – соответственно первая и вторая предорсальная, превентральная, преанальная и интердорсальная длина; lP, lV, lC – соответственно длина Р, V и С; hID, hIID, hA – соответственно длина наибольшего колючего и ветвистого луча D и наибольшего ветвистого луча А; lID, lIID, lA – соответственно длина основания колючего и мягкого D и А; ao – длина рыла; oo – горизонтальный диаметр глаза; io – ширина межглазничного промежутка; lmx – длина верхней челюсти; НИС, НПС – соответственно научно-исследовательское и научно-промысловое судна; ст. – океанографическая станция; BSKU – биологический факультет Университета Коти (Япония).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
(рис. 1, 2)

Рис. 2.
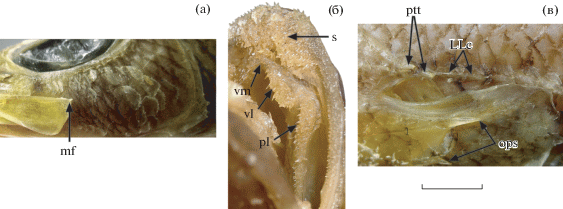
Chrionema furunoi ИО РАН № 3621, детали строения: а – пластинка maxillare, щека и praeoperculum; б – озубление крыши рта, в – область верхнего края жаберного отверстия. mf – кожный вырост maxillare, LLc – килеватые передние чешуи LL, ops – оперкулярные шипы, pl – нёбная лента зубов, ptt – посттемпоральные шипы, s – симфизная площадка зубов на praemaxillare; vm, vl – соответственно медиальная и латеральная группы зубов vomer. Масштаб (линейка общая): 5 мм.
Материал. ИО РАН № 3621, 1 экз. SL 152 мм (рис. 1), ~ 10°–15° с.ш. ~ 110° в.д., НПС “Одиссей”, рейс 1/84, трал № 70.
Описание. ID VI, IID 17, A 25, P 21, V I + 5; LL 69, LL-D 6.5, LL-A 6.0, preD 15, crcp 16; sp. br 4 + 16, pbr 15.
Тело дорсовентрально уплощённое, голова большая, 2.9 раза в SL; максимальная высота тела, приходящаяся на затылок, 8 раз содержится в SL. Рыло в 1.15 раза превышает диаметр глаза; межглазничный промежуток очень узкий (6.5 раза в диаметре глаза), мостовидный. Ноздри поровидные. Нижняя челюсть выступает вперёд, верхняя оканчивается посередине промежутка между передним краем зрачка и серединой глаза. Верхнезадний угол пластинки maxillare с очень короткой кожной мочкой (рис. 2а). На симфизе praemaxillaria широкая площадка прижатых назад игловидных зубов, назад переходящая в полоску мелких щетинковидных зубов, самые внутренние из которых немного крупнее более наружных (рис. 2б). На симфизе dentalia два ряда игловидных зубов, далее назад переходящих в полоску мелких щетинковидных зубов, в самом внутреннем ряду замещающихся на заметно более длинные, игловидные. Озубление симфиза dentalia и наружные зубные ряды praemaxillaria на большей части их длины видны при закрытом рте. На сошнике и нёбных костях полоски мелких игловидных зубов, заметно увеличенных в размере в самом внутреннем ряду. Боковые крылья головки сошника, несущие зубные скопления, заметно выступают в ротовую полость, в промежутке между ними на вершине головки сошника имеется ряд из трёх игловидных зубов, соединяющих правое и левое зубные скопления (рис. 2б). Praeoperculum с 3/4 шипиками в угловой части (рис. 2а). Operculum с двумя сильными расходящимися шипами (верхний почти в полтора раза длиннее) (рис. 2в). Верхний край suboperculum оканчивается шипом. Задний край posttemporate с двумя небольшими шипами (рис. 2в). Первые две−три чешуи LL с резким срединным килем, но без шипа (рис. 2в).
Спинные плавники далеко разобщены, величина интердорсального промежутка равна длине основания ID, прижатые назад колючки ID далеко не достигают начала IID. Концы части колючих и ветвистых лучей ID и IID обломаны, из-за чего с уверенностью установить длину наибольших из них невозможно. Конец прижатого к телу Р достигает вертикали основания второго луча А. Длина V составляет около двух третей длины вентроанального промежутка; наибольший луч этого плавника – третий. С усечённый. Анус расположен вблизи начала А. Чешуя на туловище почти полностью облетела, но хорошо сохранилась на голове. На верхней поверхности головы чешуя циклоидная, вперёд заходит за линию, соединяющую передние края глаз, но не доходит до вершин восходящих отростков praemaxillaria, не переходит на боковые стороны рыла, так что бóльшая часть его поверхности голая. Щека от вертикали переднего края зрачка и жаберная крышка полностью покрыты чешуей; чешуя ктеноидная, сохранилась только в передней половине щеки (рис. 2а). На вентральной поверхности головы чешуйный покров полностью отсутствует. На истмусе чешуя редуцирована в размерах, вросла в кожу, между основаниями V и на основании Р хорошо развита. Отдельные сохранившиеся чешуи LL ктеноидные (рис. 2в), кроме них отдельные циклоидные чешуйки сохранились у основания А. LL образует плавный изгиб на уровне конца Р.
Измерения, в % SL: lc 34.2, wc 17.1, H 12.5, h 4.6, lcp 7.2, aID 36.2, aIID 51.3, aV 23.7, aA 49.3, iD 10.9, lP 16.5, lV 14.5, lC 15.1, hA 6.6, lID 10.9, ao 9.9, oo 8.6, io 1.3, lmx 13.8.
Окраска. Общий фон окраски фиксированного экземпляра светлый, желтоватый, чисто-белый на груди и брюхе; края чешуйных карманов тёмные, намного более интенсивно окрашенные посередине боков, где эта пигментация образует узкие косые тёмные линии, перемежающиеся с плохо отграниченными тёмными пятнами неправильной формы (числом до восьми). На боках головы нечёткие поперечные и диагональные тёмные полоски; вершина рыла, крыша черепа за глазами и нижняя губа темнее фона остающейся части головы. Ротовая полость, включая зубные ленты челюстей, целиком светлая (рис. 2б). Перепонка между лучами ID, IID и А сохранилась плохо, но по меньшей мере была пигментирована между первой и второй колючками ID. IID, по-видимому, с продольными рядами пятнышек. Конец С затемнён. Р с пятью широкими поперечными тёмными полосами. V и А не пигментированы.
Замечания. Изученный экземпляр не имеет значимых отличий от рыб из вод Японии (Okamura, Yamachi, 1982; Masuda et al., 1985; Nakabo, 2002). Для вьетнамского экземпляра характерны несколько более длинные Р по сравнению с популяциями из более северных акваторий (как у голотипа, происходящего из зал. Тоса, так и у экземпляра из вод Тайваня, изображенного на веб-ресурсе Taiwan Biodiversity Network: https:// www.tbn.org.tw. Version 05/2020, их концы не заходят за вертикаль начала А). Также впервые для рода Chrionema Gilbert, 1905 отмечается наличие кожной мочки на пластинке maxillare, хотя бы и крошечной (рис. 2а). Характерными признаками C. furunoi являются наличие поперечных полос на Р и сочетание на боках тела узких косых тёмных линий и пятен. По окраске вьетнамский экземпляр хорошо соответствует голотипу этого вида (BSKU № 12430), изученному мною по фотографии. Виды рода Chrionema могут быть разделены на две группы по степени очешуенности головы, числу лучей А и числу sp. br. К группе видов, имеющих развитый чешуйный покров головы, малое число лучей А (18) и большое число sp. br (25–30), принадлежат C. squamentum (Ginsburg, 1955), C. squamiceps Gilbert, 1905 и C. pallidum Parin, 1990 c разобщёнными локальными ареалами (Карибское море, Гавайские о-ва, хребет Сала-и-Гомес). Возможные пути образования таких ареалов рассматривались ранее (Прокофьев, 2019). Другую группу представляют виды (C. furunoi, C. chlorotaenia McKay, 1971 и C. chryseres Gilbert, 1905) с частично перекрывающимися ареалами в Индо-Пацифике, характеризующиеся почти голым рылом и полностью бесчешуйной нижней поверхностью головы, бóльшим числом А, но меньшим – sp. br (соответственно 23–28 и 20–24). Вероятно, эти группы являются естественными, и все виды второй группы представлены в Западной Пацифике. По сравнению с другими западнотихоокеанскими видами, у C. furunoi меньше чешуй в LL (67–78 против 78–89) и больше лучей в А (25–28 против 23–26, как правило, 24 или 25) (Iwamoto, Staiger, 1976; Okamura, Yamachi, 1982; Nakabo, 2002). По особенностям пигментации рта C. furunoi соответствует C. chryseres, а по степени развития чешуи на верху головы занимает промежуточное положение между этим видом и C. chlorotaenia (см. ниже определительную таблицу).
До сих пор вид был известен только от берегов Южной Японии (к югу от Кумано Нада): как в тихоокеанских водах, так и в водах Восточно-Китайского моря, − и с хребта Кюсю-Палау (Okamura, Yamachi, 1982; Masuda et al., 1985; Shinohara et al., 2001; Nakabo, 2002), а также от северо-восточного побережья о. Тайвань (Восточно-Китайское море) (https://www.tbn.org.tw. Version 05/2020). Для Вьетнама и Южно-Китайского моря в целом он указывается впервые. Ранее уже отмечалось, что ряд видов, считавшихся ранее эндемичными или субэндемичными (южнояпонскими) для района залива Тоса и хребта Кюсю-Палау, оказались представленными в водах присвалового участка прибрежья Центрального и Южного Вьетнама к югу от 10° с.ш. (Prokofiev, 2014).
Со времени последней ревизии рода Chrionema (Iwamoto, Staiger, 1976) прошло более 40 лет, за которые были добавлены два новых вида и отмечены (Парин, 1990) некоторые сложности с разграничением видов C. chlorotaenia и C. chryseres. Поэтому я считаю целесообразным предложить новую таблицу для определения видов рода Chrionema, составленную на основании непосредственного изучения экземпляров видов C. chlorotaenia, C. chryseres и C. pallidum из коллекции ИО РАН и ряда литературных источников (Iwamoto, Staiger, 1976; Okamura, Yamachi, 1982; Щербачев и др., 1986; Парин, 1990; Bañón et al., 2001; Nakabo, 2002).
1(2) LL 50–58; Западная Атлантика (Карибское море и Флоридский пролив) ...........C. squamentum
2(1) LL > 60; Индо-Пацифика на восток до хребта Сала-и-Гомес и Юго-Восточная Атлантика ……………....................................................... 3
3(4) LL 60–78 .………………………………...............5
4(3) LL 78–89 [большая часть рыла и вся нижняя поверхность головы не покрыты чешуей; A 23–26; sp. br 21–24] ..…………………………............ 9
5(6) Бóльшая часть рыла и вся нижняя поверхность головы не покрыты чешуей; A 25–28; sp. br. 20; грудной плавник с поперечными тёмными полосами, на боках тела тёмные пятна и узкие косые полосы; воды Южной Японии, Тайваня и Вьетнама ……………….…………………………........ C. furunoi
6(5) Голова почти полностью покрыта чешуёй (её нет только на губах, подбородке, жаберной перепонке и на узкой полосе перед глазом); A 18; sp. br 25–30; грудной плавник без поперечных полос; на боках только округлые или неправильной формы пятна, иногда (C. pallidum) плохо выраженные ……………………………………….................................. 7
7(8) Во втором спинном плавнике 14–15 (обычно 15) лучей; хребет Сала-и-Гомес …………C. pallidum
8(7) Во втором спинном плавнике 16 лучей; Гавайские о-ва …………….…................... C. squamiceps
9(10) Зубные полосы на praemaxillare и верхняя губа с интенсивно-чёрной пигментацией; чешуя на дорсальной поверхности головы вперёд не заходит за линию, соединяющую передний край глаз; Индо-Вест-Пацифика на восток до Японии и Австралии, Юго-Восточная Атлантика (банка Вальдивия) ……………………………….. C. chlorotaenia
10(9) Зубные полосы на praemaxillare не пигментированы, верхняя губа лишь немного затемнена; чешуя на дорсальной поверхности головы вперёд почти достигает задних ноздрей; Тихий океан от Южной Японии и Тайваня до Гавайских о-вов и хребта Сала-и-Гомес ……………………… C. chryseres
(рис. 3)

Материал. ИО РАН № 3622, 1 экз. SL 90 мм, 01° с.ш. 107° в. д., НИС “Витязь”, рейс 57, ст. 7248, проба 105, трал Сигсби, глубина ~60 м, время траления 21.30–21.50, 6.03.1975 г.
Описание. ID V, IID 21½, A 18½, P 17, V I + 5; LL 55, LL-D 4, LL-A 12, preD 8, crcp 23; sp. br 3 + 10, pbr 16.
Тело заметно сжато с боков на всем протяжении, его ширина на уровне оснований Р в 1.3 раза меньше наибольшей высоты; голова 3.75 раза в SL, слабо уплощённая, её максимальная ширина в 1.6 раза меньше длины и в 1.1 раза меньше наибольшей высоты. Длина рыла равняется диаметру глаза, 3.7 раза в длине головы. Передняя ноздря с коротким треугольным клапаном, задняя открывается простой порой. Нижняя челюсть слабо выступает вперёд, верхняя оканчивается на вертикали переднего края зрачка. На симфизе praemaxillaria с каждой стороны по одному загнутому клыковидному зубу, за которым следует ряд конических зубов, более крупных в задней половине ряда. На симфизе dentalia между двумя сильными загнутыми клыковидными зубами (более крупными, чем на praemaxillaria) расположено четыре зуба, позади клыка зубы сидят в один ряд, в задней части ряда они заметно крупнее. На сошнике поперечный ряд из восьми сильных конических зубов; нет зубов на нёбных костях. Praeoperculum несёт около 18 коротких притуплённых шипиков по свободному краю, лучше выраженных над перегибом кости; suboperculum с 8−10 слабо выраженными шипиками, большей частью не выступающими за задний край кости; свободный край interoperculum гладкий. Operculum с одним плоским шипом, не достигающим конца оперкулярной лопасти на 1.5 своей длины. Оперкулярная лопасть хорошо выражена, треугольная, узкая. Сеть вторичных пор на голове слабо развита; в супраорбитальном канале пять простых пор; имеется корональная пора. Инфраорбитальный сенсорный канал с короткими простыми вентральными ответвлениями в передней половине lacrimale (широкие) и в заглазничной части (узкие); темпоральный канал открывается серией немногочисленных вторичных пор, супратемпоральная комиссура с единственным срединным передним ответвлением и множественными простыми задними ветвями. Нижнечелюстные поры простые, по четыре с каждой стороны, подбородочные поры широко расставлены и соединены низким поперечным кожным гребешком. Четыре нижние преоперкулярные поры простые, две верхние открываются серией вторичных пор на коротких задних ответвлениях. На щеке шесть рядов чешуй.
ID и IID соединены, перепонка между последним колючим и первым ветвистым лучом с неглубокой выемкой, верхний край перепонки расположен на уровне первой трети длины первого мягкого луча. Наибольший колючий луч ID – четвёртый. Концы прижатых назад Р едва не достигают концов V, конец прижатого к брюху V достигает промежутка между третьим и четвёртым лучами А. Наибольший луч V – четвёртый. C усечённый, его верхняя лопасть совершенно не выражена. Анус расположен у начала А. Чешуя повсеместно ктеноидная, включая область щеки, жаберную крышку, истмус, основание Р и предорсальный участок тела; ктении очень мелкие.
Измерения, в % SL: lc 26.7, wc 16.7, H 18.9, h 8.9, w 14.4, lcp 12.2, aID 26.7, aIID 35.6, aV 25.6, aA 44.4, lP 18.9, lV 22.2, lC 16.7, hID 6.7, hIID 13.3, hA 12.2, ao 7.2, oo 7.2, io 3.1, lmx 10.6.
Окраска. Экземпляр полностью депигментирован, лишь на С прослеживается серия поперечных тёмных полос, нижняя половина С выглядит темнее верхней (за счёт меланофорного крапа, различимого только при большом увеличении).
Замечания. Изученный экземпляр хорошо соответствует признакам голотипа этого вида (Randall, 2008), за исключением полного отсутствия у него верхней лопасти С, хорошо развитой у голотипа и единственного ранее известного экземпляра этого вида. Это отличие нельзя объяснить онтогенетической изменчивостью, поскольку изученный экземпляр и голотип имеют близкие размеры (SL соответственно 90 и 87 мм), однако оно вряд ли имеет существенное значение. К сожалению, особенности пигментации не могут быть сравнены из-за её практически полной утраты у описываемого экземпляра. P. compressa принадлежит к комплексу видов “P. somaliensis”, характеризующемуся большим разнообразием в Индо-Вест-Пацифике (Randall, 2008; Прокофьев, 2010), но хорошо отличается от всех видов сильно сжатым с боков телом (ширина в 1.3 раза меньше высоты против 1.1 раза) и полным отсутствием циклоидных чешуй где-либо на голове и теле. Новый экземпляр подтверждает диагностическую значимость первого признака и отсутствие какой-либо связи его с деформацией или аномалией. Голотип этого вида был пойман при тралении на глубине 40–60 м у южного берега о. Сумбава (Малые Зондские о‑ва). Новый экземпляр был добыт с тех же глубин в южной части Южно-Китайского моря между о. Калимантан и полуостровом Малакка, что существенно расширяет известный ареал вида.
ЗАКЛЮЧЕНИЕ
Новые нахождения документируют присутствие Chrionema furunoi и Parapercis compressa в ихтиофауне Южно-Китайского моря. Поимка C. furunoi в акватории Вьетнама существенно расширяет известный ареал этого вида к югу. Подобно ряду других элиторальных видов с преимущественно южнояпонскими ареалами (Prokofiev, 2014), C. furunoi представляет собой северо-западнотихоокеанский элемент в ихтиофауне Южно-Китайского моря, принадлежащего к Тропическому региону Индийско-Западнотихокеанской подобласти. Присутствие таких видов в водах Центрального и Южного Вьетнама связывают с притоком вод Североэкваториального течения через Лусонский пролив во время зимнего муссона (Prokofiev, 2014). P. compressa, напротив, является тропическим видом, возможно, лишь краем ареала заходящим в южную часть Южно-Китайского моря. Отсутствие сколько-нибудь существенных морфологических отличий новых экземпляров от ранее известных свидетельствует об отсутствии изолирующих барьеров в пределах известных ареалов обоих видов. Обнаружение у исследованного экземпляра C. furunoi маленького кожного выроста на пластинке maxillare (рис. 2а) позволяет предполагать, что этот вырост вторично отсутствует у представителей этого рода (по сравнению с крупным выростом у видов близкого рода Bembrops Steindachner, 1876). Сходные пропорции тела нового экземпляра и голотипа P. compressa подтверждают, что необычно сильное для параперковых сжатие тела с боков у этого вида является не аномалией, а видоспецифичной особенностью.
Список литературы
Парин Н.В. 1990. Перкофовые рыбы (Percophidae) с подводного хребта Сала-и-Гомес (юго-восточная часть Тихого океана) // Вопр. ихтиологии. Т. 30. Вып. 1. С. 1–12.
Прокофьев А.М. 2010. Два новых вида рыб из семейств Muraenidae (Anguilliformes) и Mugiloididae (Perciformes) из вод Вьетнама // Там же. Т. 50. № 5. С. 603–612.
Прокофьев А.М. 2019. Новый вид серрановых окуней рода Pseudogramma из Индийского океана (Serranidae) // Там же. Т. 59. № 3. С. 350–354. https://doi.org/10.1134/S0042875219030184
Щербачев Ю.Н., Парин Н.В., Пахоруков Н.В., Пиотровский А.С. 1986. Мезобентальные и мезобентопелагические рыбы некоторых подводных поднятий в западной тропической части Индийского океана // Тр. ИO АН СССР. Т. 121. С. 195–214.
Bañón R., Armesto A., Garabana D. 2001. First record of Chrionema chlorotaenia (Perciformes, Percophidae) from the southeastern Atlantic // Cybium. V. 25. № 3. P. 278–280.
Cantwell G.E. 1964. A revision of the genus Parapercis family Mugiloididae // Pac. Sci. V. 18. № 3. P. 239−280.
Iwamoto T., Staiger J.C. 1976. Percophidid fishes of the genus Chrionema Gilbert // Bull. Mar. Sci. V. 26. № 4. P. 488–498.
Masuda H., Amaoka K., Araga C. et al. 1985. The fishes of the Japanese Archipelago. Pt. 1. Tokyo: Tokai Univ. Press, i-xxii + 437 p.
Nakabo T. 2002. 297. Percophidae duckbills // Fishes of Japan with pictorial keys to the species. V. 2. Tokyo: Tokai Univ. Press. P. 1065–1068.
Okamura O., Yamachi F. 1982. Chrionema furunoi Okamura et Yamachi // Fishes of the Kyushu-Palau Ridge and Tosa Bay. The intensive research of unexploited fishery resources on continental slopes. Tokyo: Jpn Fish. Res. Conserv. Assoc. P. 254–257.
Prokofiev A.M. 2014. Erythrocles microceps (Emmelichthyidae) in the waters of Vietnam // J. Ichthyol. V. 54. № 10. P. 781–785.
Randall J.E. 2008. Six new sandperches of the genus Parapercis from the western Pacific, with description of a neotype for P. maculata (Bloch et Schneider) // Raffles Bull. Zool. Suppl. № 19. P. 159–178.
Shinohara G., Endo H., Matsuura K. et al. 2001. Annotated checklist of the deepwater fishes from Tosa Bay, Japan // Monographs Nat. Sci. Mus.Tokyo. № 20. Deep-sea fauna and pollutants in Tosa Bay. P. 283–343.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии