

Вопросы ихтиологии, 2021, T. 61, № 1, стр. 88-99
Распределение и некоторые черты биологии рогатковых рода Triglops (Cottidae) в Японском море
О. И. Пущина 1, В. В. Панченко 2, *, М. И. Бойко 1, А. И. Галеев 1
1 Тихоокеанский филиал Всероссийского научно-исследовательского института рыбного хозяйства
и океанографии – ТИНРО
Владивосток, Россия
2 Национальный научный центр морской биологии Дальневосточного отделения РАН − ННЦМБ ДВО РАН
Владивосток, Россия
* E-mail: vlad-panch@yandex.ru
Поступила в редакцию 25.10.2019
После доработки 07.11.2019
Принята к публикации 07.11.2019
Аннотация
Выявлено, что южная граница ареала большеглазого триглопса Triglops scepticus проходит в районе 37°23′ с.ш. Подтверждено расположение южной границы ареала триглопса Джордена T. jordani в водах Южной Кореи, а остроносого T. pingelii – в северокорейских. В российских водах Японского моря триглопс Джордена встречается на глубинах 14–230 м при температуре –1−13.8°C, остроносый – соответственно 30–424 м и –0.3−9.7°C, большеглазый – 57–586 м и 0.3−4.3°C; в тёплый период концентрации несколько сдвинуты в сторону мелководья, в холодный – в сторону глубоководья. Длина и масса тела одноразмерных особей повышаются со степенью глубоководности видов. Триглопс Джордена достигает длины 20 см, остроносый – 23 см и большеглазый – 26 см. У все трёх видов самки достигают больших размеров; у триглопсов Джордена и остроносого они доминируют в популяции, а у большеглазого триглопса выше доля самцов. В северо-западной части Японского моря большеглазый и остроносый триглопсы являются планктофагами, потребляющими в основном гипериид и эвфаузиид, а триглопс Джордена – нектобентофагом, в питании которого доминируют мизиды и мелкие креветки. В весенне-летний период средние величины суточных рационов взрослых особей триглопсов Джордена, остроносого и большеглазого составляют соответственно 3.3, 2.4 и 1.8% массы тела. Биомасса видов рода Triglops в российской зоне Японского моря составляет ~2.1 тыс. т.
Рыбы рода Triglops (Cottidae) широко распространены в северо-западной части Тихого океана (Линдберг, Красюкова, 1987; Pietsch, 1993; Amaoka et al., 1995; Борец, 2000; Шейко, Федоров, 2000; Федоров и др., 2003; Парин и др., 2014; Fricke et al., 2019; Froese, Pauly, 2019). В Японском море обитают три представителя этого рода: триглопс Джордена Triglops jordani, остроносый T. pingelii и большеглазый T. scepticus триглопсы. В российских водах Японского моря они являются обычными видами (Новиков и др., 2002; Соколовский и др., 2007), далее на юг встречаются до п-ова Корея (Mori, 1952; Сон Ён Хо, 1986; Kim, Yoon, 1992).
В настоящее время триглопсов добывают в незначительном количестве в качестве прилова при промысле других более ценных гидробионтов. Между тем в некоторых районах обитания виды рода Triglops образуют скопления, которые могут успешно осваиваться промышленностью. Так, в тихоокеанских водах северных Курильских о-вов и Камчатки большеглазый триглопс входит в тройку наиболее массовых промысловых видов рогатковых (Орлов, Токранов, 2008). Биология этих рыб изучена недостаточно. Первые данные о размерно-возрастных характеристиках, сроках нереста и плодовитости триглопсов приведены в работе Паракецова (1962). Позднее были довольно подробно исследованы их размерно-половой состав и питание у побережья Камчатки (Токранов 1991, 1995), а также особенности распределения у Юго-Восточной Камчатки и в тихоокеанских водах северных Курильских о-вов (Tokranov, Orlov, 2008; Токранов, Орлов, 2009). В этих районах общими с Японским морем являются остроносый и большеглазый триглопсы. По российским водам Японского моря имеются в основном фрагментарные сведения о распределении представителей этого рода (Вдовин, Зуенко, 1997; Калчугин, 1998; Ким Сен Ток, 2001, 2004; Соломатов, 2008; Панченко, Зуенко, 2009; Панченко и др., 2016) и питании большеглазого триглопса (Ким Сен Ток, 2001; Пущина, 2005).
Цель настоящей работы – на основании обобщения имеющихся у авторов данных уточнить границы ареалов, охарактеризовать сезонное батиметрическое и пространственное распределение, размерно-половую структуру, питание, а также оценить уровень запасов рогатковых рода Triglops в российских водах Японского моря.
МАТЕРИАЛ И МЕТОДИКА
В работе использованы материалы донных траловых съёмок и контрольных тралений, выполненных ТИНРО в российских водах Японского моря в разные сезоны 1984−2016 гг. Проанализированы данные 8459 тралений на шельфе и материковом склоне российской зоны Японского моря на глубинах от 2 до 935 м, из которых 3727 сопровождались измерением придонной температуры воды. Привлечены также результаты совместных работ с учёными Национального института исследований рыбного хозяйства Республики Корея (National Institute of Fisheries Science) у япономорского побережья Южной Кореи на НИС “Тамгу–5”, “Тамгу–20” и “Тамгу–21”. По этому району мы располагаем данными по 129 тралениям на глубинах 70−400 м, полученными в 14 съёмках в июне−ноябре 1999−2017 гг.
Основная масса тралений ниже 5-метровой изобаты выполнена донными тралами с мягким грунтропом, длиной верхней подборы от 20 до 69 м (горизонтальное раскрытие от 13 до 38 м) и ячеёй в кутце от 10 до 30 мм. Скорость тралений составляла 1.5−4.5 (в среднем 2.6) узлов. На меньших глубинах орудием лова служил донный трал с длиной верхней подборы 14.6 м (горизонтальное раскрытие 6 м), а также модифицированный в ТИНРО бим-трал (Вдовин и др., 2009) с ячеёй в кутце 10 мм и горизонтальным раскрытием 3 м; скорость траления 1.3−3.4 (2.4) узлов. Для получения сравнимых результатов при использовании тралов разных конструкций уловы рыб пересчитывали на плотность по формуле: P = B/S, где P – плотность, кг/км2; B – улов, кг; S – площадь траления, км2; коэффициенты уловистости при пересчёте на плотность не вводили.
Деление на гидрологические сезоны принято по классификации Зуенко (1994): зима – январь−февраль, весна – март−апрель, лето – июнь−сентябрь, осень – ноябрь−декабрь; май является переходным месяцем между весенним и летним сезонами, октябрь – между летним и осенним. Первая половину мая мы относим к весеннему сезону, вторую – к летнему; первую половину октября – к летнему, вторую – к осеннему. Поскольку батиметрическое распределение триглопсов в зимний, весенний и осенний периоды оказалось сходным, эти данные были объединены.
Измерили длину по Смитту (FL) 14543 особей триглопса Джордена, 16892 – остроносого и 5817 – большеглазого триглопсов, из них с определением пола соответственно 2059, 2405 и 1719 экз., индивидуальной массы − 672, 300 и 305 экз.
Материал на питание (357 желудков большеглазаго триглопса, 106 – остроносого и 44 – триглопса Джордена) собран в водах Приморья в апреле−августе. Обработку проб вели в соответствии с “Методическим пособием …” (1974). Величину суточного рациона оценивали по методу Новиковой (1949) в модификации Чучукало и Напазакова (1999) путём определения продолжительности переваривания исходной (восстановленной) массы пищевых объектов в зависимости от температуры придонного слоя воды. Средние величины рационов видов и соотношение в них компонентов вычислены с учётом биомассы исследованных размерных групп и их вклада в общее потребление.
Анализ пространственного распределения выполнен с помощью программного пакета CHARTMASTER методом сплайн-аппроксимации.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Северные границы ареалов рассматриваемых видов располагаются значительно выше Японского моря, а южные проходят по его акватории: у материкового побережья южная граница ареала большеглазого и остроносого триглопсов находится в водах Северной Кореи (Mori, 1952; Сон Ён Хо, 1986), триглопса Джордена – в северной части вод Республики Корея (Kim, Yoon, 1992). Полученные нами при работе с южнокорейскими учёными данные о распространении триглопсов Джордена и остроносого соответствуют литературным: на акватории Южной Кореи, недалеко от её северной границы, отмечены четыре одиночных поимки особей триглопса Джордена FL 15.5–16.2 см на глубинах 135−163 м, тогда как остроносый триглопс в южнокорейских водах в уловах отсутствовал. Имеющиеся у нас данные по распределению большеглазого триглопса позволяют расширить южную зону его обитания. В октябре 2016 г. при тралении в япономорских водах Республики Корея у 37°23′ с.ш. на глубине 147 м при температуре воды у дна 1.6°С были отмечены две особи этого вида FL 16.5 и 17.1 см. Следовательно, южная граница ареала большеглазого триглопса располагается не в водах Северной Кореи, как считалось ранее, а несколько ниже северной оконечности Южной Кореи.
По данным Соломатова (2008), в северо-западной части Японского моря триглопс Джордена входит в состав элиторальной группировки, обитающей на глубинах 20−300 м, но в основном тяготеющей к шельфовой зоне; остроносый триглопс − элиторально-мезобентальной группировки, придерживающейся шельфа и верхнего отдела материкового склона, а большеглазый – мезобентально-элиторальной группировки, предпочитающей материковый склон. Наши данные в целом подтверждают отнесение этих представителей рода Triglops к указанным группировкам: в российских водах Японского моря триглопс Джордена был отмечен нами в диапазоне глубин 14–230 м, остроносый – 30–424 м, большеглазый – 57–586 м.
Предпочитаемый батиметрический диапазон обитания триглопса Джордена в течение всего года располагался в пределах глубин 55−120 м; в тёплый период наибольшие плотности скоплений отмечены на 80−100 м, а в холодный – ниже 100-метровой изобаты (рис. 1а). Сезонные различия обусловлены выходом части особей на относительно небольшие глубины, подверженные наибольшим сезонным колебаниям температуры. В тёплый период минимальная глубина обнаружения триглопса Джордена в интенсивно прогреваемой мелководной зоне составила 14 м, тогда как в холодный период он избегал охлаждённых вод на глубинах <22 м.
Рис. 1.
Средние плотности исследованных видов рода Triglops в разных диапазонах глубин в российских водах Японского моря в тёплый (−◼−) и в холодный (- - × - -) периоды: а – триглопс Джордена T. jordani, б – остроносый триглопс T. pingelii, в – большеглазый триглопс T. scepticus.
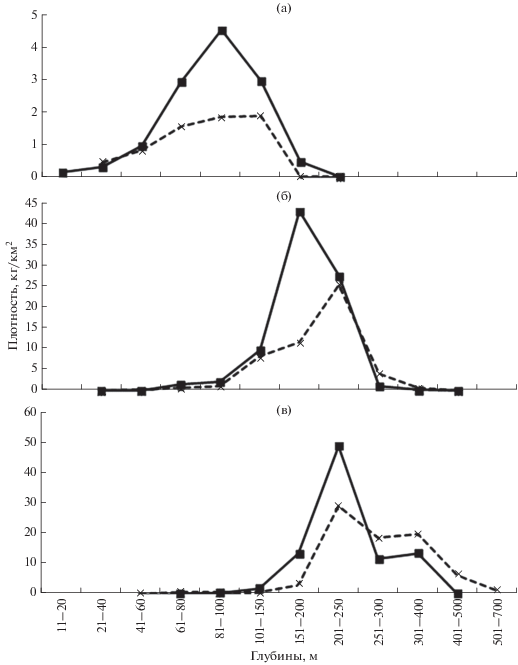
У остроносого триглопса различия в сезонном распределении выражены в меньшей степени: сходными оказались не только максимальные (411 м в тёплый период и 424 м в холодный) и предпочитаемые, но и минимальные глубины обитания (30 и 35 м) (рис. 1б). Во все сезоны основные скопления рыб отмечены в диапазоне глубин 150−250 м с той лишь разницей, что в холодный период они несколько смещались в сторону глубоководья.
Батиметрический диапазон обитания большеглазого триглопса в тёплый период составил 72−471 м, в холодный – 57−586 м, т.е. летом он был несколько меньшим, особенно со стороны глубоководья. Некоторые различия наблюдались и в глубинах предпочтения: в течение года величина уловов была наибольшей в диапазоне 200−250 м, однако в холодный период в отличие от тёплого значительные скопления формировались и в прилегающих глубоководных диапазонах (рис. 1в).
Таким образом, сезонные батиметрические различия у рассматриваемых видов выражены слабо. Отличие состоит в некотором смещении предпочитаемых глубин в сторону мелководной зоны в тёплый период. Все три вида в течение всего года обитают в основном в нижней части шельфа и в верхнем отделе материкового склона, где гидрологический режим не претерпевает значительных изменений и сохраняется стабильная слабоположительная температура (Зуенко, 1994, 2008).
В тралениях, проведённых с измерением фоновых показателей, придонная температура варьировала в пределах −1.8…22.6°C. Триглопс Джордена, обитающий на относительно небольших глубинах, подверженных значительному сезонному колебанию температуры, является самым эвритермным: в уловах он отмечен в температурном диапазоне −1.0…13.8°C (табл. 1). Остроносый триглопс обитает при температуре −0.3…9.7°C. Наиболее стенотермен глубоководный большеглазый триглопс, отмеченный только при положительных фоновых значениях – 0.3−4.3°C. Наибольшая частота встречаемости первого вида наблюдалась при температуре 1.6−7.0°С, второго – 1.0−2.2°С, третьего – 0.8−1.7°C. В тихоокеанских водах северных Курильских о-вов и Юго-Восточной Камчатки у остроносого и большеглазого триглопсов предпочитаемый диапазон температуры близок (Орлов, Токранов, 2008; Tokranov, Orlov, 2008; Токранов, Орлов, 2009).
Таблица 1.
Частота встречаемости рассматриваемых видов рода Triglops в российских водах Японского моря в зависимости от придонной температуры, %
| Вид | Температура,°С | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ≤ 0 | 0.01−0.20 | 0.21−0.40 | 0.41−0.60 | 0.61−0.80 | 0.81−1.00 | 1.01−1.20 | 1.21−1.40 | 1.41−1.60 | 1.61−1.80 | 1.81−2.00 | 2.01−2.20 | 2.21−2.50 | 2.51−3.00 | 3.01−4.00 | 4.01−5.00 | 5.01−7.00 | 7.01−10.00 | 10.01−14.00 | >14 | |
| T. jordani | 11.5 | 6.1 | 6.5 | 3.6 | 2.5 | 8.4 | 7.1 | 17.7 | 24.5 | 44.3 | 45.2 | 43.4 | 38.9 | 43.3 | 35.3 | 44.9 | 45.7 | 22.3 | 12.1 | 0 |
| T. pingelii | 9.0 | 6.1 | 7.7 | 6.4 | 13.4 | 21.9 | 41.4 | 40.3 | 49.5 | 35.8 | 38.9 | 31.3 | 20.2 | 23.0 | 3.5 | 1.3 | 1.3 | 2.2 | 0 | 0 |
| T. scepticus | 0 | 0 | 4.1 | 7.3 | 16.2 | 23.4 | 33.9 | 28.0 | 22.4 | 18.8 | 13.9 | 9.0 | 8.5 | 8.2 | 1.4 | 3.8 | 0 | 0 | 0 | 0 |
| Число тралений | 78 | 33 | 169 | 357 | 439 | 342 | 292 | 293 | 277 | 240 | 144 | 144 | 129 | 122 | 142 | 80 | 75 | 92 | 146 | 133 |
Пространственное распределение исследуемых видов в российских водах Японского моря различается. Скопления наиболее мелководного триглопса Джордена наблюдались, как правило, лишь на юге района – в зал. Петра Великого и в прилегающих водах Северного Приморья (рис. 2а), что объясняется наличием здесь обширного шельфа. В пределах этой зоны выделяются два центра плотности: один локализован в западной, другой – в восточной части зал. Петра Великого. Обусловлено это прохождением в центральной части залива, у 132° в.д., зоны раздела циркуляции вод (Зуенко, 2008). Сходная картина пространственного распределения отмечена здесь и для других рыб, в частности, для представителя семейства Cottidae – нитчатого шлемоносца Gymnocanthus pistilliger (Вдовин и др., 1994; Панченко и др., 2020). Подобная зал. Петра Великого и даже более обширная шельфовая зона расположена и на севере района, в Татарском проливе, однако скоплений триглопса Джордена, в особенности у материкового побережья, здесь не наблюдали. Объяснить это можно особенностями гидрологии: в Татарском проливе, в особенности в его западной части, за счёт зимней конвекции формируется подповерхностный слой вод с пониженной температурой и солёностью, нередко сохраняющийся даже летом (Зуенко, 2008).
Рис. 2.
Пространственное распределение исследованных видов рода Triglops в российских водах Японского моря: а – триглопс Джордена T. jordani, б – остроносый триглопс T. pingelii, в – большеглазый триглопс T. scepticus.
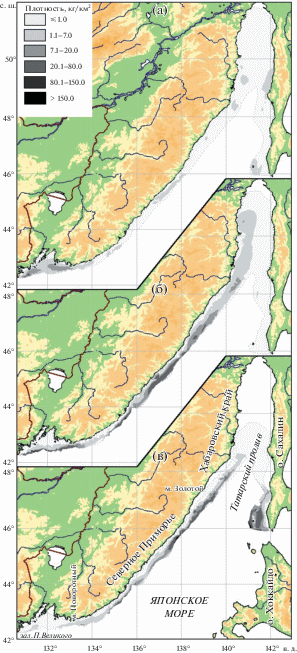
Повышенными концентрациями остроносого триглопса характеризовалась обширная зона центральной части материкового побережья (рис. 2б). Помимо этого, как и у триглопса Джордена, скопления формировались и на юге района − в зал. Петра Великого и прилегающей части Северного Приморья. Однако, возможно, за счёт предпочтения остроносым триглопсом более глубоководной зоны снижение его уловов в районе 132° в. д. не наблюдалось. Выраженный спад плотности между южным и центральным районами прослеживался восточней 134° в.д. Разрыв в распределении рыб, в том числе представителей Cottidae, связанный с системой вергенций вод (Яричин, Покудов, 1982), отмечался в этом районе и ранее (Калчугин, 1998; Соломатов, 2008). Минимальные уловы остроносого триглопса на предпочитаемых глубинах наблюдались в Татарском проливе у западного побережья Сахалина.
Районы повышенных концентраций большеглазого триглопса у материкового побережья отмечены в центральной области, однако располагались они несколько выше, чем у остроносого триглопса; далее на север плотность постепенно уменьшалась (рис. 2в). Помимо скоплений с материковой стороны значительные концентрации большеглазого триглопса отмечены у юго-западного побережья Сахалина. Формирование в Татарском проливе у островного побережья более плотных, чем со стороны материка, скоплений этого вида может быть обусловлено тем, что юго-западное побережье Сахалина находится под воздействием тёплого Цусимского течения, проникающего вдоль Японских о-вов и заносящего тёплые субтропические водные массы (Зуенко, 2008). Видимо, большеглазый триглопс более остальных представителей рода избегает расположенных северней выхоложенных и распреснённых вод. В Татарском проливе этот вид отмечался лишь до 49° с.ш., хотя и севернее располагаются глубины его обитания, в том числе и предпочитаемые на остальной акватории. Триглопсы же остроносый и Джордена в Татарском проливе, хотя и не образовывали концентраций, отмечались практически на всей его акватории. Возможно, для большеглазого триглопса в большей степени, чем для других описываемых видов, имеет значение наличие стабильных не только температурных, но и других фоновых условий. В данном случае речь может идти о неблагоприятном для него понижении солёности в кутовой части пролива. Фоновые условия в этом районе неблагоприятны для обитания и многих других видов гидробионтов, в том числе для представителей семейства рогатковых – пёстрого получешуйника Hemilepidotus gilberti и двурогого бычка Enophrys diceraus (Панченко, Пущина, 2018, 2019).
В наших уловах встречены особи триглопса Джордена FL 4−20 см, преобладала размерная группа 14−15 см; остроносого – соответственно 6−23 см и 17−18 см, большеглазого – 9−26 см и 20−22 см (рис. 3). Таким образом, предельные достигаемые размеры повышались со степенью глубоководности видов. Подобное мы отмечали и для других родов семейства рогатковых (Панченко, Пущина, 2004; Панченко, 2012). Логично, что у видов мелкого размера и половая зрелость наступает при меньшей длине, чем у крупноразмерных. По данным Паракецова (1962), триглопс Джордена достигает половой зрелости при FL 11−12 см, остроносый – 12−14 см, большеглазый – 16−18 см.
Рис. 3.
Размерный состав триглопса Джордена T. jordani (−◼−), остроносого триглопса T. pingelii (−×−) и большеглазого триглопса T. scepticus (−△−) в уловах донного трала в российских водах Японского моря.
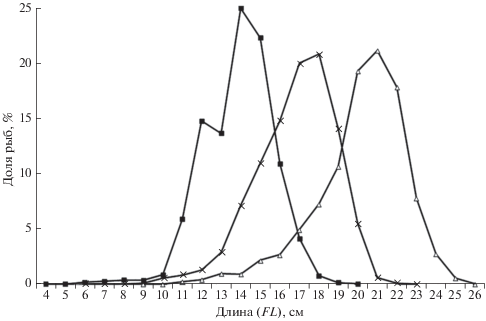
По нашим данным, связь между длиной (FL, см) и массой (W, г) триглопса Джордена описывается степенной зависимостью W = 0.0389 × × FL2.3317 (R2 = 0.7799), остроносого – W = 0.0110 × × FL2.8675 (R2 = 0.6415), большеглазого – W = = 0.0025 × FL3.4871 (R2 = 0.8838). Из представленных аллометрических зависимостей видно, что среди одноразмерных особей трёх видов (FL > 11 см, на основе которых по большей части и выведены зависимости) наименьшую массу имеет триглопс Джордена, наибольшую – большеглазый триглопс, а остроносый занимает промежуточную позицию. Следовательно, масса тела одноразмерных рыб также повышается с возрастанием степени глубоководности видов.
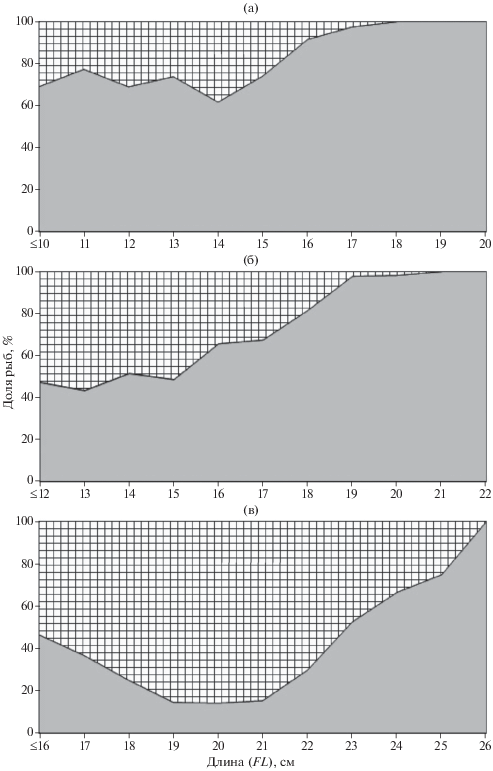
Длина промеренных с определением пола особей триглопса Джордена варьировала в пределах 6−20 см, остроносого – 10−22 см, большеглазого – 14−26 см. Для многих видов семейства Cottidae характерен половой диморфизм, проявляющийся, в частности, в различии размеров самцов и самок (при этом у одних видов большей длины достигают самки, у других – самцы) и соотношении полов. Например, в прикамчатских водах в популяциях остроносого, большеглазого и вильчатого T. forficatus триглопсов самки достигают больших размеров, чем самцы; в целом по численности они преобладают, хотя среди мелких рыб до определённых размеров доминируют самцы (Токранов, 1995). По нашим данным, в Японском море у трёх изученных видов предельные размеры самок также превышают таковые самцов: у триглопса Джордена − FL 20 против 17 см (рис. 4а), у остроносого триглопса – 22 против 20 см (рис. 4б) (очевидно, что промеренные без определения пола три особи FL 23 см также были самками), у большеглазого – 26 против 25 см (рис. 4в). Однако соотношение полов у этих видов различалось. У триглопсов Джордена и остроносого доля самок в уловах составляла соответственно 76 и 77%. Различие состояло в том, что у триглопса Джордена доминирование самок проявлялось во всех размерных группах (хотя среди особей FL ≤ 14 см оно выражено в меньшей степени), тогда как у остроносого среди рыб FL ≤ 15 см соотношение полов оказалось близко 1 : 1. У большеглазого триглопса, наоборот, в целом доминировали самцы. Доля их в уловах составила 76%. Наибольшей (85%) она была среди рыб FL 19−21 см; в следующих размерных группах доля самцов постепенно уменьшалась вплоть до полного их отсутствия среди рыб предельного размера. У мелких особей FL ≤ 16 см соотношение полов приближалось 1 : 1. Таким образом, у молоди большеглазого и остроносого триглопсов соотношение полов приблизительно равное. Возможно, и у триглопса Джордена среди не попавших в выборку особей FL ≤ 6 см соотношение полов значительно не различается. Поскольку уловистость мелкоразмерных рыб использованными орудиями лова меньше, чем более крупных особей, можно полагать, что при достоверном учёте молоди общая доля самцов и самок в популяциях оказалась бы ближе, чем в анализируемых уловах. Однако несомненно, что и в этом случае сохраняется преобладание в япономорских водах у триглопсов Джордена и остроносого самок, а у большеглазого – самцов.
Рис. 4.
Соотношение размерных групп самцов (  ) и самок (
) и самок ( ) исследованных видов рода Triglops в российских водах Японского моря: а – триглопс Джордена T. jordani, б – остроносый триглопс T. pingelii, в – большеглазый триглопс T. scepticus.
) исследованных видов рода Triglops в российских водах Японского моря: а – триглопс Джордена T. jordani, б – остроносый триглопс T. pingelii, в – большеглазый триглопс T. scepticus.
В трофологическом отношении к настоящему времени наиболее изучен большеглазый триглопс. В прибрежных водах Камчатки он является бентомакропланктофагом: потребляет в основном эвфаузиид (Euphausiacea) и пелагических амфипод (Hyperiidea) летом и донных амфипод (Gammaridea) − зимой (Токранов, 1991; Чучукало, 2006). В других районах ареала этот вид питается преимущественно зоопланктоном как в тёплый (Wakabayashi, 1986; Mito et al., 1999; Пущина, 2005), так и в холодный период года (Ким Сен Ток, 2001). Остроносый триглопс – нектобентофаг, использующий в прикамчатских водах главным образом мизид (Mysidacea) и молодь креветок (Pandalidae, Hippolytidae, Crangonidae) (Токранов, 1991; Чучукало, 2006). О питании триглопса Джордена известно лишь, что в декабре на тихоокеанском шельфе Хоккайдо основным кормом ему служат мизиды и креветки (Macrura), второстепенным – амфиподы (Amphipoda), изоподы (Isopoda) и многощетинковые черви (Polychaeta) (Nishikawa et al., 2000).
По нашим наблюдениям, в северо-западной части Японского моря большеглазый и остроносый триглопсы являются планктофагами. В весенне-летний период >90% рациона первого вида и >85% второго составляют гиперииды Themisto japonica и эвфаузииды рода Thysanoessa, при этом в пище самого глубоководного большеглазого триглопса выше доля гипериид, а остроносого – эвфаузиид (табл. 2). Кроме того, важную роль в питании молоди первого вида играют веслоногие ракообразные (Copepoda, преимущественно Calanus glacialis и Metridia pacifica), а у взрослых особей второго – мизиды (в основном Hemiacanthomysis dimorpha). В размерно-возрастной изменчивости состава пищи большеглазого триглопса в исследуемом районе прослеживаются те же тенденции, что и в прикамчатских водах летом (Токранов, 1991): доминирование копепод в желудках молоди FL 8−10 см сменяется преобладанием эвфаузиид у более крупной FL 11−15 см, однако с увеличением размеров взрослых рыб FL 16−25 см доля этих ракообразных в рационе постепенно сокращается, уступая место гипериидам. Некоторое снижение потребления эвфаузиид по мере роста происходит и у взрослого остроносого триглопса, но главные изменения в его пищевом спектре связаны с замещением мизид, составляющих значительную часть рациона особей FL 11−15 см, гипериидами у более крупных FL 16−20 см.
Несмотря на ограниченный материал по питанию триглопса Джордена, а также различия в районах и сезонах сбора проб, полученные нами данные оказались довольно близки к таковым японских авторов (Nishikawa et al., 2000). Как и на шельфе Хоккайдо, в северо-западной части Японского моря основными компонентами рациона взрослых особей этого вида являются придонные ракообразные – мизиды (в основном Inusitatomysis insolita, Xenacanthomysis pseudomacropsis) и креветки рода Eualus (в сумме 76.2%), второстепенными – полихеты, амфиподы и эвфаузииды. Хотя в желудках наиболее крупных рыб FL 16−20 см и доминирует зоопланктон (преимущественно гиперииды), трофический статус триглопса Джордена определяют особенности питания преобладающей в уловах группы FL 11−15 см (рис. 3), характеризуя его как нектобентофага.
Таблица 2.
Состав пищи исследованных видов рода Triglops в северо-западной части Японского моря (апрель–август), % массы
| Компонент пищи и другие показатели | T. scepticus | T. pingeli | T. jordani | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| FL, см | FL, см | FL, см | |||||||||
| 8−10 | 11−15 | 16−20 | 21−25 | в среднем | 11−15 | 16−20 | в среднем | 11−15 | 16−20 | в среднем | |
| Polychaeta | – | 1.2 | 1.1 | 0.8 | 1.0 | 0.5 | – | 0.2 | 5.8 | 0.9 | 5.5 |
| Polynoidae gen. sp. | – | – | – | – | – | – | – | – | 3.6 | – | 3.4 |
| Nothria sp. | – | – | – | – | – | – | – | – | 2.2 | – | 2.1 |
| Polychaeta varia | – | 1.2 | 1.1 | 0.8 | 1.0 | 0.5 | – | 0.2 | – | 0.9 | + |
| Copepoda | 70.7 | 15.7 | – | – | 0.3 | – | – | – | 1.5 | 13.1 | 2.1 |
| Calanus glacialis | 35.4 | 4.9 | – | – | 0.1 | – | – | – | – | – | – |
| Neocalanus plumchrus | – | 4.9 | – | – | 0.1 | – | – | – | – | – | – |
| Bradyidius pacificus | – | 2.5 | – | – | + | – | – | – | – | – | – |
| Metridia pacifica | 35.3 | 3.4 | – | – | 0.1 | – | – | – | 1.5 | 13.1 | 2.1 |
| Isopoda | 9.2 | 6.2 | – | – | 0.1 | – | – | – | – | – | – |
| Amphipoda | 18.3 | 19.1 | 48.6 | 59.7 | 55.0 | 3.6 | 36.5 | 23.8 | 5.7 | 66.2 | 8.9 |
| Jassa falcata | – | – | – | – | – | – | – | – | – | 5.6 | 0.3 |
| Dulichia sp. | – | – | – | – | – | – | 0.9 | 0.6 | 5.0 | 5.6 | 5.0 |
| Anonyx nugax | – | – | 0.6 | 5.4 | 3.6 | – | – | – | – | – | – |
| Paroediceros lynceus | – | 5.1 | – | – | 0.1 | – | – | – | – | – | – |
| Gammaridea varia | 9.1 | 1.7 | – | 0.1 | 0.1 | 1.3 | 0.1 | 0.5 | 0.7 | – | 0.7 |
| Primno macropa | – | – | 2.0 | 0.7 | 1.1 | – | – | – | – | – | – |
| Themisto japonica | 9.2 | 12.3 | 46.0 | 53.5 | 50.1 | 2.3 | 35.5 | 22.7 | – | 55.0 | 2.9 |
| Mysidacea | – | – | 0.8 | 0.6 | 0.6 | 26.7 | 2.0 | 11.5 | 59.5 | – | 56.2 |
| Hemiacanthomysis dimorpha | – | – | – | – | – | 20.9 | – | 8.0 | 9.9 | – | 9.4 |
| Inusitatomysis insolita | – | – | – | – | – | 0.8 | 0.1 | 0.4 | 20.9 | – | 19.7 |
| Xenacanthomysis pseudomacropsis | – | – | – | – | – | 5.0 | 1.9 | 3.1 | 28.7 | – | 27.1 |
| Mysidacea varia | – | – | 0.8 | 0.6 | 0.6 | – | – | – | – | – | – |
| Euphausiacea | – | 54.0 | 45.2 | 37.7 | 40.7 | 67.2 | 61.5 | 63.7 | 4.0 | 15.2 | 4.7 |
| Euphausia pacifica | – | – | – | – | – | – | – | – | 4.0 | 10.6 | 4.4 |
| Thysanoessa inermis | – | 54.0 | 36.5 | 30.4 | 33.0 | – | – | – | – | – | – |
| Th. longipes | – | – | 8.7 | 7.3 | 7.7 | 67.2 | 61.5 | 63.7 | – | 4.6 | 0.3 |
| Decapoda | 1.8 | 0.7 | 0.1 | 0.3 | 0.2 | 2.0 | – | 0.8 | 21.1 | – | 20.0 |
| Eualus fabricii | – | – | – | 0.2 | 0.1 | – | – | – | 15.1 | – | 14.3 |
| E. macilentus | – | – | – | – | – | – | – | – | 6.0 | – | 5.7 |
| Crangon dalli | – | – | – | – | – | 2.0 | – | 0.8 | – | – | – |
| Mezocrangon intermedia | – | – | 0.1 | 0.1 | 0.1 | – | – | – | – | – | – |
| Decapoda larvae | 1.8 | 0.7 | – | – | + | – | – | – | – | – | – |
| Cephalopoda | – | – | 3.6 | – | 1.3 | – | – | – | – | – | – |
| Chaetognatha | – | 2.5 | – | – | + | – | – | – | – | – | – |
| Прочие | – | 0.6 | 0.6 | 0.9 | 0.8 | – | – | – | 2.4 | 4.6 | 2.6 |
| Средний суточный рацион, % массы тела | 3.8 | 3.5 | 2.0 | 1.7 | 1.8 | 3.8 | 1.9 | 2.4 | 3.7 | 1.1 | 3.3 |
| Число желудков, шт. | 10 | 15 | 185 | 147 | 357 | 42 | 64 | 106 | 27 | 17 | 44 |
| Доля пустых желудков, % | – | 6.7 | 14.6 | 10.2 | 12.0 | 2.4 | 25.0 | 16.0 | 14.8 | 41.2 | 25.0 |
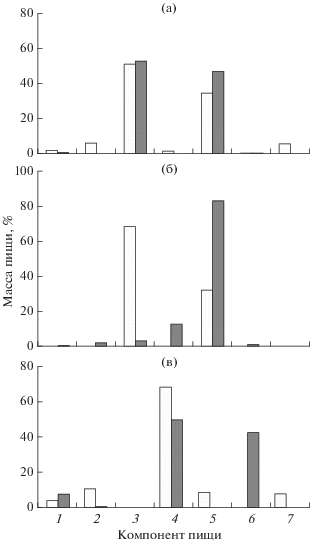
Сезонная динамика пищевого спектра взрослых особей менее всего выражена у большеглазого триглопса, основная масса которого в процессе миграций не покидает материковый склон. От весны к лету в его рационе несколько повышается доля эвфаузиид (с 34.5 до 49.6%), но гиперииды остаются практически на одном уровне (51.0−52.6%) (рис. 5а). В то же время смещение основных концентраций остроносого триглопса с верхней части склона на шельф сопровождается резким падением доли гипериид в его пище (с 68.1% в апреле−мае до 2.7% в июне−августе) и значительным ростом эвфаузиид (с 31.9 до 82.9%) и отчасти мизид (до 11.8%) (рис. 5б). В рационе постоянно обитающего на шельфе триглопса Джордена с апреля по август преобладают мизиды (68.8−50%), но летом их доля (как и суммарная доля второстепенных кормовых организмов) заметно сокращается за счёт активного потребления креветок (до 42.4%) (рис. 5в).
Рис. 5.
Состав пищи исследованных видов рода Triglops в северо-западной части Японского моря в апреле−мае (◻) и июне−августе ( ): а − большеглазый триглопс T. scepticus FL 16−25 см, б − остроносый триглопс T. pingeli FL 11−20 см, в − триглопс Джордена Triglops jordani FL 11−15 см. Компонент пищи: 1 – Polychaeta, 2 – Gammaridea, 3 – Hyperiidea, 4 – Mysidacea, 5 – Euphausiacea, 6 – Decapoda, 7 – прочие.
): а − большеглазый триглопс T. scepticus FL 16−25 см, б − остроносый триглопс T. pingeli FL 11−20 см, в − триглопс Джордена Triglops jordani FL 11−15 см. Компонент пищи: 1 – Polychaeta, 2 – Gammaridea, 3 – Hyperiidea, 4 – Mysidacea, 5 – Euphausiacea, 6 – Decapoda, 7 – прочие.
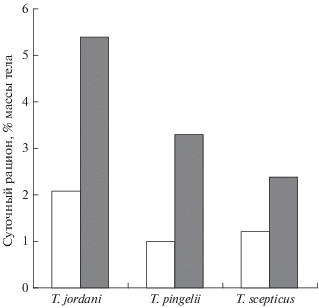
Сведений об интенсивности питания триглопсов крайне мало. Известно, что у побережья Камчатки индексы наполнения желудков большеглазого и остроносого триглопсов летом выше, чем зимой (Токранов, 1991), а суточный рацион остроносого FL 10−20 см в июле составляет 2.8% массы тела (Чучукало, 2006). По нашим наблюдениям, кормовая активность взрослых особей рассматриваемых видов находится в обратной зависимости от степени их глубоководности. В весенне-летний период наибольшую среднюю величину суточного рациона имеет триглопс Джордена (3.3% массы тела), за ним в порядке уменьшения этого показателя следуют остроносый (2.4%) и большеглазый (1.8%) (табл. 2). Интенсивность питания триглопсов в июне−августе в 2.0−3.3 раза выше, чем в апреле−мае (рис. 6).
Рис. 6.
Средняя величина суточного рациона триглопса Джордена Triglops jordani FL 11−15 см, остроносого триглопса T. pingeli FL 11−20 см и большеглазого триглопса T. scepticus FL 16−25 см в северо-западной части Японского моря в апреле−мае и июне−августе; обозначения см. на рис. 5.
По данным учётных траловых съёмок ТИНРО, в последние 10 лет у материкового побережья российской зоны Японского моря биомасса видов рода Triglops, рассчитанная с использованием коэффициента уловистости 0.3−0.4, составляет в среднем 1.50 тыс. т. Наибольший вклад здесь вносит триглопс остроносый (0.66 тыс. т), за которым следуют большеглазый (0.44 тыс. т) и Джордена (0.18 тыс. т). У западного побережья Сахалина среднее значение биомассы представителей рода Triglops составляет 0.63 тыс. т. Основу запаса здесь слагает большеглазый триглопс (0.58 тыс. т), при незначительной биомассе триглопсов Джордена (0.04 тыс. т) и остроносого (0.01 тыс. т). Таким образом, общая биомасса видов рода Triglops в российской зоне Японского моря составляет ~2.1 тыс. т.
ВЫВОДЫ
1. Южная граница ареала большеглазого триглопса проходит не в водах Северной Кореи, как считалось ранее, а в водах Южной Кореи в районе 37°23′ с.ш. Южная граница ареала триглопса Джордена также находится в япономорских водах Южной Кореи, а остроносого – в северокорейских водах.
2. В российских водах Японского моря триглопс Джордена встречается в диапазоне глубин 14–230 м, остроносый – 30–424 м, большеглазый – 57–586 м; во все сезоны глубины распространения каждого из них значительно не различаются. Предпочитаемый диапазон первого вида располагается на глубинах 55–120 м, второго – 150–250 м, третьего – 200–400 м. В тёплый период в пределах глубин предпочтения концентрации несколько сдвинуты в сторону мелководья, в холодный – в сторону глубоководья.
3. Максимальные размеры особей повышаются со степенью глубоководности видов. Триглопс Джордена достигает FL 20 см, остроносый – 23 см и большеглазый – 26 см. Среди особей одинакового размера масса тела также выше у более глубоководных видов.
4. Триглопс Джордена, обитающий и на относительно небольших, подверженных значительному сезонному колебанию температуры глубинах, – самый эвритермный; остроносый триглопс занимает промежуточное положение, глубоководный большеглазый триглопс – стенотермный. Первый отмечается в температурном диапазоне −1.0…13.8°С, второй в диапазоне –0.3…9.7°С, третий – 0.3−4.3°C. Триглопс Джордена предпочитает температуру 1.6−7.0°С, остроносый – 1.0−2.2°С, большеглазый – 0.8−1.7°C.
5. У всех трёх видов в результате полового диморфизма самки достигают бóльших размеров. У триглопсов Джордена и остроносого они и доминируют в популяции, тогда как у большеглазого триглопса выше доля самцов.
6. В северо-западной части Японского моря большеглазый и остроносый триглопсы являются планктофагами, потребляющими в основном гипериид и эвфаузиид, а триглопс Джордена – нектобентофагом, в питании которого доминируют мизиды и мелкие креветки. В весенне-летний период средние величины суточных рационов взрослых особей триглопсов Джордена, остроносого и большеглазого составляют соответственно 3.3, 2.4 и 1.8% массы тела. Интенсивность питания триглопсов в июне−августе в 2.0−3.3 раза выше, чем в апреле−мае.
7. По данным учётных траловых съёмок ТИНРО, биомасса видов рода Triglops в российской зоне Японского моря составляет ~2.1 тыс. т. В целом основу биомассы слагает большеглазый триглопс, далее следует остроносый, затем – триглопс Джордена.
Список литературы
Борец Л.А. 2000. Аннотированный список рыб дальневосточных морей. Владивосток: Изд-во ТИНРО-центр, 192 с.
Вдовин А.Н., Зуенко Ю.И. 1997. Вертикальная зональность и экологические группировки рыб залива Петра Великого // Изв. ТИНРО. Т. 122. С. 152−176.
Вдовин А.Н., Швыдкий Г.В., Афанасьева Н.И. и др. 1994. Пространственно-временная изменчивость распределения нитчатого шлемоносца в заливе Петра Великого // Экология. № 4. С. 53−59.
Вдовин А.Н., Мизюркин М.А., Пак А. 2009. Возможности использования бим-трала для прямых учетов гидробионтов // Вопр. рыболовства. Т. 10. № 1(37). С. 150−160.
Зуенко Ю.И. 1994. Типы термической стратификации вод на шельфе Приморья // Комплексные исследования морских гидробионтов и условий их обитания. Владивосток: Изд-во ТИНРО. С. 20−39.
Зуенко Ю.И. 2008. Промысловая океанография Японского моря. Владивосток: Изд-во ТИНРО-центр, 227 с.
Калчугин П.В. 1998. Распределение рогатковых (Cottidae) у северного Приморья // Изв. ТИНРО. Т. 123. С. 82–88.
Ким Сен Ток. 2001. Зимние миграции шельфовых рыб в зону материкового склона юго-западного Сахалина // Вопр. ихтиологии. Т. 41. № 5. С. 593−604.
Ким Сен Ток. 2004. Сезонные особенности вертикальной структуры ихтиоценов западносахалинского шельфа и островного склона // Там же. Т. 44. № 1. С. 77−88.
Линдберг Г.У., Красюкова З.В. 1987. Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Ч. 5. Л.: Наука, 526 с.
Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях. 1974. М.: Наука, 254 с.
Новиков Н.П., Соколовский А.С., Соколовская Т.Г., Яковлев Ю.М. 2002. Рыбы Приморья. Владивосток: Изд-во Дальрыбвтуз, 552 с.
Новикова Н.С. 1949. О возможности определения суточного рациона рыб в естественных условиях // Вестн. МГУ. № 9. С. 107−111.
Орлов А.М., Токранов А.М. 2008. Особенности распределения и динамика уловов некоторых потенциально промысловых видов рогатковых рыб (Cottidae) в тихоокеанских водах северных Курильских островов и юго-восточной Камчатки // Матер. Междунар. конф. “Современное состояние водных биоресурсов”. Новосибирск: Агрос. С. 173−178.
Панченко В.В. 2012. Возраст и рост шлемоносных бычков рода Gymnocanthus (Cottidae) в заливе Петра Великого и прилегающих районах Приморья // Вопр. ихтиологии. Т. 52. № 2. С. 234−247.
Панченко В.В., Зуенко Ю.И. 2009. Распределение бычков семейства Cottidae в заливе Петра Великого Японского моря в летний период // Вопр. рыболовства. Т. 10. № 4 (40). С. 750−763.
Панченко В.В., Пущина О.И. 2004. Биологическая характеристика керчаковых рыб рода Myoxocephalus (Cottidae) зал. Петра Великого Японского моря // Изв. ТИНРО. Т. 138. С. 120−153.
Панченко В.В., Пущина О.И. 2018. Распределение и некоторые черты биологии пёстрого получешуйника Hemilepidotus gilberti (Cottidae) в российских водах Японского моря // Вопр. ихтиологии. Т. 58. № 4. С. 439–449. https://doi.org/10.1134/S0042875218040148
Панченко В.В., Пущина О.И. 2019. Распределение и некоторые черты биологии двурогого бычка Enophrys diceraus (Cottidae) в российских водах Японского моря // Там же. Т. 59. № 2. С. 163–173. https://doi.org/10.1134/S0042875219020176
Панченко В.В., Калчугин П.В., Соломатов С.Ф. 2016. Уточнение глубин обитания и максимальных размеров донных и придонных видов рыб в российских водах Японского моря // Там же. Т. 56. № 3. С. 264–283. https://doi.org/10.7868/S0042875216030152
Панченко В.В., Матвеев А.А., Панченко Л.Л. 2020. Сезонное распределение нитчатого шлемоносца Gymnocanthus pistilliger (Cottidae) в российских водах Японского моря // Там же. Т. 60. № 2. С. 174−182. https://doi.org/10.31857/S0042875220020174
Паракецов И.А. 1962. О некоторых защитных приспособлениях у разных популяций и видов тихоокеанских подкаменщиков (Cottidae) // Тр. ИМЖ АН СССР. Вып. 42. С. 146−154.
Парин Н.В., Евсеенко С.А., Васильева Е.Д. 2014. Рыбы морей России: аннотированный каталог. М.: Т-во науч. изд. КМК, 733 с.
Пущина О.И. 2005. Питание и пищевые взаимоотношения массовых видов донных рыб в водах Приморья в весенний период // Изв. ТИНРО. Т. 142. С. 246−269.
Соколовский А.С., Дударев В.А., Соколовская Т.Г., Соломатов С.Ф. 2007. Рыбы российских вод Японского моря: аннотированный и иллюстрированный каталог. Владивосток: Дальнаука, 200 с.
Соломатов С.Ф. 2008. Состав и многолетняя динамика донных ихтиоценов северного Приморья: Автореф. дис. … канд. биол. наук. Владивосток: ТИНРО-центр, 24 с.
Сон Ён Хо. 1986. О распределении рыб в прибрежных водах Восточного моря // Тр. Вонсан. ин-та рыб. хоз-ва Восточного моря. № 1. С. 132−150.
Токранов А.М. 1991. Особенности питания рогатковых рыб рода Triglops Reinhardt (Cottidae) в прибрежных водах Камчатки // Бюл. МОИП. Отд. биол. Т. 96. Вып. 5. С. 46−52.
Токранов А.М. 1995. Размерно-половая структура рогатковых рыб рода Triglops (Cottidae) в прибрежных водах Камчатки // Вопр. ихтиологии. Т. 35. № 1. С. 134–136.
Токранов А.М., Орлов А.М. 2009. Особенности распределения и динамика уловов рогатковых рыб рода Triglops Reinhardt, 1830 (Cottidae) в тихоокеанских водах северных Курильских островов и юго-восточной Камчатки // Докл. IX Междунар. науч. конф. “Сохранение биоразнообразия Камчатки и прилегающих морей”. Петропавловск-Камчатский: Камчатпресс. С. 125−140.
Федоров В.В., Черешнев И.А., Назаркин М.В. и др. 2003. Каталог морских и пресноводных рыб северной части Охотского моря. Владивосток: Дальнаука, 204 с.
Чучукало В.И. 2006. Питание и пищевые отношения нектона и нектобентоса в дальневосточных морях. Владивосток: Изд-во ТИНРО-центр, 483 с.
Чучукало В.И., Напазаков В.В. 1999. К методике определения суточных рационов питания и скорости переваривания пищи у хищных и бентосоядных рыб // Изв. ТИНРО. Т. 126. С. 160−171.
Шейко Б.А., Федоров В.В. 2000. Глава 1. Рыбообразные и рыбы. Класс Cephalaspidomorphi – Миноги. Класс Chondrichthyes – Хрящевые рыбы. Класс Holocephali – Цельноголовые. Класс Osteichthyes – Костные рыбы // Каталог позвоночных Камчатки и сопредельных морских акваторий. Петропавловск-Камчатский: Камчат. печат. двор. С. 7−69.
Яричин В.Г., Покудов В.В. 1982. Формирование структурных особенностей гидрофизических полей и течений в северной глубоководной части Японского моря // Тр. ДВНИГМИ. Вып. 96. С. 86–95.
Amaoka K., Nakaya K., Yabe M. 1995. The fishes of Northern Japan. Sapporo: Kita-Nihon Kaiyo Center Co. Ltd, 391 p.
Fricke R., Eschmeyer W.N., Van der Laan R. (eds.). 2019. Eschmeyer’s catalog of fishes: genera, species, references. (http://researcharchive.calacademy.org/research/Ichthyology/catalog/ fishcatmain.asp.Version 06/2019)
Froese R., Pauly D. (eds.). 2019. Fishbase. World Wide Web electronic publication. (www.fishbase.de/search.php. Version 04/2019)
Kim I.-S., Yoon C.-H. 1992. Synopsis of the Family Cottidae (Pisces: Scorpaeniformes) from Korea // Kor. J. Ichthyol. V. 4. № 1. P. 54−79.
Mito K., Nishimura A., Yanagimoto T. 1999. Ecology of groundfishes in the Eastern Bering Sea, with emphasis on food habits // Dynamics of the Bering Sea / Eds. Loughlin T.R., Ohtani K. Fairbanks: Alaska Sea Grant College Program. P. 537−580.
Mori T. 1952. Check-list of the fishes of Korea // Mem. Hyogo Univ. Agric. V. 1. № 3. P. 1−228.
Nishikawa J., Sonoda T., Sakurai I. et al. 2000. Diets of demersal fishes and macrobenthos in the coastal water off Tomakomai, Hokkaido // Bull. Jpn. Soc. Sci. Fish. V. 66. № 1. P. 33−43.
Pietsch T.W. 1993. Systematics and distribution of cottid fishes of the genus Triglops Reinhardt (Teleostei:Scorpaeniformes) // Zool. J. Linn. Soc. V. 109. № 4. P. 335–393.
Tokranov A.M., Orlov A.M. 2008. Patterns of distribution and catch dynamics of sculpins of the genus Triglops (Cottidae) in the Pacific waters off the northern Kuril Islands and southeastern Kamchatka // Marine biodiversity and bioresources of the North-Eastern Asia: book and abstracts. Jeju, Korea: Cheju Nat. Univ. P. 129−132.
Wakabayashi K. 1986. Interspecific feeding relationships on the continental shelf of the eastern Bering Sea, with special reference to yellowfin sole // Bull. N. Pacif. Fish. Comm. V. 47. P. 3−30.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии