

Вопросы ихтиологии, 2021, T. 61, № 2, стр. 153-166
Распространение и биологические особенности хариуса Thymallus thymallus (Thymallidae) на Европейском Северо-Востоке России
В. И. Пономарев 1, *, А. Б. Захаров 1
1 Институт биологии Коми научного центра Уральского отделения РАН − ИБ Коми НЦ УрО РАН
Сыктывкар, Россия
* E-mail: ponomarev@ib.komisc.ru
Поступила в редакцию 10.02.2020
После доработки 28.05.2020
Принята к публикации 29.05.2020
Аннотация
Изучены распространение и некоторые особенности экологии европейского хариуса Thymallus thymallus крайнего Европейского Северо-Востока России. Во многих водных системах этого региона сохраняется близкое к естественному состояние вида. Выявлена существенная экологическая и генетическая разнокачественность популяций. По соотношению частот генотипов полиморфной двухаллельной системы трансферринов сыворотки крови популяции хариуса речной системы Печоры можно объединить в два кластера – уральский и тиманский, которые отличаются от группировок Кольского п-ова. Предполагается, что широкий спектр экологических условий разных ландшафтных комплексов обеспечивает успешное существование всего видового разнообразия европейского хариуса северо-востока европейской части России.
Европейский хариус Thymallus thymallus широко распространён: его ареал простирается от Британских о-вов и Франции на западе до Полярного и Приполярного Урала на востоке, от Скандинавии и Большеземельской тундры на севере до Балканского п-ова и Южного Урала на юге (Анацкий, 1996; Новоселов, 2000; Зиновьев, 2007; Богданов, Мельниченко, 2010; Рыбы …, 2010; Аськеев и др., 2016). В последние десятилетия этот вид стал редким во многих западноевропейских странах: здесь он отнесён к числу исчезающих и подлежит особой охране (Northcote, 1995; Persat, 1996; Koskinen, Primmer, 2001; Uiblein et al., 2001; Carlstein, 2004; Fieseler, Wolter, 2006; Maire at al., 2017). Популяции европейского хариуса р. Урал и верховьев Волги включены в Красную книгу Российской Федерации. Вид занесён в Красные книги целого ряда субъектов Российской Федерации (Вологодской, Ивановской, Костромской, Оренбургской, Челябинской, Ярославской областей, Пермского края, республик Марий Эл и Татарстан).
При этом в большинстве бассейнов рек северо-востока европейской части России европейский хариус играет важную роль в рыбной части водных сообществ (Сидоров, Решетников, 2014). В частности, на Южном, Среднем и Северном Тимане его доля в промысловой части рыбного населения составляет 24−96% (Захаров, Бознак, 2019). Хариус наряду с гольяном Phoxinus phoxinus является одним из доминирующих в лососёвых реках Европейского Севера России (Мартынов, 2007). Однако биология вида в этой чрезвычайно обширной части его ареала изучена весьма фрагментарно (Сидоров, 1974; Соловкина, 1975; Зиновьев, 1979, 2007; Анацкий, 1996; Пономарев и др., 1998; Павлов и др., 2000; Коротаева, 2003; Скоринова и др., 2014; Захаров, Бознак, 2019). Между тем актуальность подобного рода информации трудно переоценить с позиций назревшей необходимости разработки комплекса мероприятий по сохранению и восстановлению многочисленных популяций вида, подверженных риску исчезновения.
Цель работы − изучить распространение, структуру популяций и особенности экологии европейского хариуса в горных и равнинных реках и озёрах крайнего Европейского Северо-Востока России.
МАТЕРИАЛ И МЕТОДИКА
Материал собирали в 1979−2019 гг. в бассейнах рек Печора, Мезень, Северная Двина, а также водотоков Большеземельской тундры, впадающих в губы Баренцева и Карского морей (рис. 1, табл. 1).
Рис. 1.
Карта-схема основных речных систем северо-востока Европы с указанием мест сбора материалов: 1−20 − западные склоны Северного, Приполярного и Полярного Урала; 21−27 − Печорская низменность, 28−42 – Тиманский кряж, 43−53 – Большеземельская тундра, 54 – р. Тулома, Кольский п-ов.
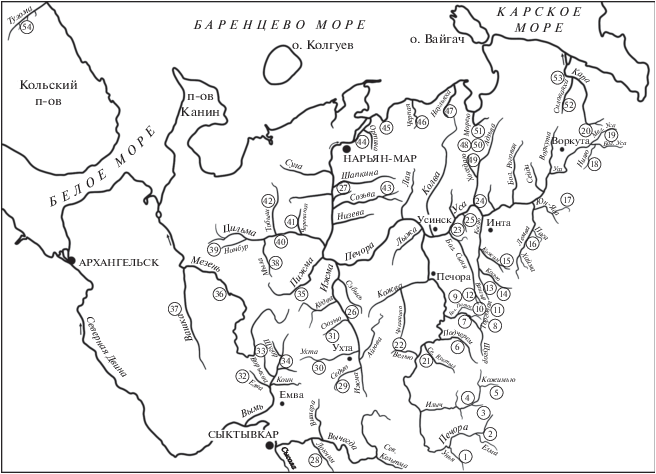
Таблица 1.
Характеристика собранного и исследованного материала
| Выборка, № | Водоём | Бассейн | Период отлова (месяц, год) | Число рыб, экз. |
|---|---|---|---|---|
| Западные склоны Северного, Приполярного и Полярного Урала | ||||
| 1 | р. Унья | Печора | IX.2013 | 20 |
| 2 | р. Елма | То же | V.2012 | 13 |
| 3 | Истоки р. Печора | Баренцево море | VII.2003 | 100 |
| 4 | р. Илыч | Печора | IX.2008 | 41 |
| 5 | р. Кожимью | Печора, приток р. Илыч | IX.2015 | 51 |
| 6 | р. Подчерем | Печора | VII.2017 | 31 |
| 7 | р. Щугор | То же | 1986−1990 | 137 |
| 8 | р. Торговая | Печора, приток р. Щугор | 2010−2014 | 169 |
| 9 | р. Большой Паток | То же | 1998−2018 | 162 |
| 10 | оз. Номты | Печора, водосбор р. Большой Паток | VII.2006 | 10 |
| 11 | оз. Сыняты | Печора, исток р. Войвож-Сыня | IX.2010 | 20 |
| 12 | р. Войвож-Сыня | Печора, приток р. Большая Сыня | 2013−2014 | 182 |
| 13 | р. Вангыр | Печора, приток р. Косью | 2010-2012 | 116 |
| 14 | р. Косью | Печора, приток р. Уса | VIII.2014 | 35 |
| 15 | р. Кожим | Печора, приток р. Косью | 1982−1993 | 95 |
| 16 | оз. Пагаты | Печора, приток р. Лемва | VI.2007 | 32 |
| 17 | оз. Хойлаты | Печора, приток р. Юн-Яга | VIII.2009 | 13 |
| 18 | р. Нияю | Печора, приток р. Большая Уса | 2008−2009 | 65 |
| 19 | оз. Усваты | Печора, исток р. Малая Уса | VII.2008 | 18 |
| 20 | оз. Плаунты | То же | VII.2008 | 5 |
| Печорская низменность | ||||
| 21 | р. Светлый Вуктыл | Печора | VI.2001 | 31 |
| 22 | р. Велью | То же | VI.2003 | 20 |
| 23 | р. Большая Сыня | Печора, приток р. Уса | VII.2002 | 62 |
| 24 | р. Уса | Печора | IX.2008 | 38 |
| 25 | р. Шарью | Печора, приток р. Уса | VIII.1999 | 48 |
| 26 | р. Ижма | То же | VI.2001 | 9 |
| 27 | р. Енва | » | III.1998 | 23 |
| Тиманский кряж | ||||
| 28 | р. Локчим | Северная Двина, приток р. Вычегда | VI.2005 | 68 |
| 29 | р. Седью | Печора, приток р. Ижма | VI.1993 | 45 |
| 30 | р. Ухта | То же | VII.1997 | 61 |
| 31 | р. Сюзью | » | VII.1994 | 37 |
| 32 | р. Елва | Северная Двина, приток р. Вымь | IX.2007 | 56 |
| 33 | р. Ворыква | То же | VI.1979 | 56 |
| 34 | р. Вымь | Северная Двина, приток р. Вычегда | VII.2000 | 61 |
| 35 | р. Пижма | Печора | VIII.1993 | 39 |
| 36 | р. Мезень | Белое море | VII.2006 | 109 |
| 37 | р. Вашка | Мезень | VII.2000 | 13 |
| 38 | р. Мыла | Печора, приток р. Цильма | VII.1994 | 66 |
| 39 | р. Номбур | То же | III.1995 | 7 |
| 40 | р. Цильма | Печора | III.2002 | 25 |
| 41 | р. Черепанка | Печора, приток р. Цильма | III.1996 | 30 |
| 42 | р. Тобыш | То же | VIII.2009 | 7 |
| Большеземельская тундра | ||||
| 43 | р. Ваглейвис | Печора, приток р. Созьва | VII.2001 | 44 |
| 44 | р. Ортина | Печора | VI.1999 | 43 |
| 45 | Безымянное озеро | Баренцево море, сток в оз. Большой Торавей | VII.1992 | 31 |
| 46 | р. Чёрная | Баренцево море | VII.2013 | 24 |
| 47 | оз. Наульто | Наульяха | VII.1992 | 32 |
| 48 | р. Хоседаю | Печора, приток р. Адзьва | VII.2000 | 13 |
| 49 | оз. Лангутаты | Печора, водосбор р. Адзьва | IX.2018 | 94 |
| 50 | оз. Ватьярты | То же | VIII.2001 | 39 |
| 51 | р. Морею | Баренцево море | VI.2001 | 39 |
| 52 | р. Силоваяха | Кара | VIII.2018 | 62 |
| 53 | р. Кара | Карское море | VIII.2018 | 73 |
| Кольский п-ов | ||||
| 54 | р. Тулома | Баренцево море | VIII.1980 | 47 |
В реках рыб отлавливали закидными промысловыми (ячея в приводе 24 мм, длина 60−150 м) и мальковыми (ячея 5−18 мм, длина 10−25 м) неводами, спиннингами, корабликом (распространённая среди рыбаков Севера специальная снасть для лова рыбы, с помощью которой можно доставить оснастку с мушками к нужной точке) и поплавочными удочками. В озерах использовали стандартный набор ставных жаберных сетей с ячеёй 10, 20, 30, 40, 50 и 60 мм, длиной 30 м и высотой 1.8 м.
Биологический анализ проводили общепринятыми методами (Правдин, 1966; Мина, Клевезаль, 1976; Решетников, 1980). У рыб в свежем состоянии измеряли длину по Смитту (FL), определяли общую массу тела, пол и стадию зрелости гонад. Возраст определяли по чешуе. Всего исследовали 2767 экз.
Фракционный состав сывороточных белков изучен у хариуса из бассейнов Печоры (реки Кожим, Щугор и Пижма) и Северной Двины (Вымь), а также из р. Тулома, расположенной на Кольском п-ове. Белки сыворотки крови разделяли в камере для вертикального электрофореза в полиакриламидном геле. Систему гелей и буферные растворы готовили по Аотсука и Асами (Aotsuka, Asami, 1979). Время электрофореза 3.5 ч при начальном напряжении 200 В и силе тока 20 мА. После электрофореза белковые фракции на геле выявляли красителем амидочёрным 10В. Полиморфную систему белков, расположенную в зоне β-глобулинов, идентифицировали видоспецифичным красителем как трансферрины.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Распространение и экология. Европейский хариус обитает в большинстве водотоков бассейнов уральских, тиманских, равнинных таёжных и равнинных тундровых притоков р. Печора. Как правило, он распространён от самых истоков до устьевых участков печорских притоков, а также на большей части магистрального русла этой крупнейшей североевропейской реки. Вид населяет речные системы крупных притоков Печоры: Унья, Илыч, Подчерем, Щугор, Большая и Малая Сыня, Косью, Лемва, Уса, Большая Роговая, Адзьва, Большая Макариха, Колва, Кожва, Ижма, Пижма, Цильма, Созьва, Шапкина, Сула и тысячи средних и малых притоков разного порядка.
Европейский хариус обитает в большинстве притоков р. Вычегда (бассейн Северной Двины). Вид широко распространён в бассейнах р. Мезень и многих водотоков, впадающих в Баренцево и Карское моря, в том числе в реках Кара, Коротаиха, Морею, Чёрная и целом ряде других (Соловкина, 1975; Новоселов, 2000). Хариус обычен в большинстве населённых рыбами озёр Большеземельской тундры. Среди них Вашуткины (бассейн р. Адзьва), Харбейские (бассейн р. Большая Роговая) и Падимейские (бассейн р. Сейда) озёра, образующие крупнейшие в регионе озёрно-речные системы (Сидоров, 1974), а также оз. Амбар-ты из бассейна р. Коротаиха (Сидоров, Решетников, 2014). Вид также отмечен нами во многих других озёрах восточноевропейских тундр, в частности, Хоседаты, Лангутаты, Ватьярты (бассейн р. Адзьва), Молотовей Ямботы и Никэрэматы (бассейн р. Коротаиха), Писяты (бассейн р. Колва) и в некоторых безымянных тундровых озерах бассейна р. Морею.
Как правило, в тундре хариус постоянно населяет глубокие ледниковые озёра, в промерзающих термокарстовых пока он не отмечен. Тем не менее в период открытой воды разновозрастные группировки хариуса используют для нагула и мелководные озёра. В горной области Приполярного Урала даже глубокие озёра не всегда пригодны для обитания здесь хариуса. Ранее нами было показано, что этот вид не обитает ни в одном из озёр, расположенных в долине р. Косью и соединённых постоянными протоками с руслом водотока (Пономарев, 2019). Эти озёра площадью 1.0−8.4 га и глубиной 4−14 м находятся на высоте более 225 м над уровнем моря.
В вегетационный период европейский хариус горных и предгорных рек держится территориально, преимущественно на перекатах и углублённых участках с ускоренным течением, а также в малых реках и ручьях. В подлёдный период хариус концентрируется главным образом на плёсах (Пономарев и др., 1998).
Европейский хариус обнаружен нами и во многих горных озёрах западных склонов Урала (Пономарев, 2017). Во многих глубоководных уральских озёрах хариус обитает постоянно. В частности, это крупнейшие озёра западных склонов Урала: Длинное, Торговое, Паток, Номты, Сыняты, Падежаты, Большое и Малое Балбанты, Пагаты, Хойлаты, Естото, Чаньты, Усваты, Гнетьты. В горные озёра бассейнов ряда рек, в частности Малый Паток, Войвож-Сыня и Вангыр, молодь хариуса заходит на нагул, в результате чего в верховьях этих рек в летнее время отмечаются преимущественно особи старшего возраста. В других мелководных озёрах (например, в бассейне р. Большой Паток) хариус встречается только транзитом: в период его нерестовой миграции во впадающие в эти озёра ручьи. В некоторых как глубоководных, так и промерзающих в зимний период озёрах, которые хариус использует для летнего нагула, он является единственным обитающим здесь видом рыб. В промысловой части рыбного населения большинства горных озёр европейский хариус является супердоминантом наряду либо с сибирским хариусом Th. arcticus, либо с арктическим гольцом Salvelinus alpinus, а также в некоторых случаях с окунем Perca fluviatilis.
В тундровой зоне излюбленными местообитаниями хариуса являются протоки между озёрами и участки литорали и сублиторали озёр, хотя в годы с экстремально тёплыми погодными условиями мы его отмечали на глубине до 20 м (тундровое озеро Хоседаты). На таёжной равнине хариус предпочитает углублённые плёсовые участки водотоков с погружёнными в них фрагментами стволов и корней деревьев и нависшими над поверхностью воды кронами древостоев и крутыми берегами, которые можно рассматривать в качестве дополнительных укрытий.
В водотоках и водоёмах Крайнего Европейского Северо-Востока хариус образует три хорологические формы – речную, озёрно-речную и озёрную. Одни группировки обитают только в реках (например, без озёр на водосборе), другие заходят из рек в озёра на нагул, третьи постоянно живут в озёрах. В частности, в безымянном предгорном озере на водосборе р. Большой Паток, в которое впадают относительно крупные ручьи, половозрелые особи хариуса появляются только в период нереста. В сетных уловах на протоках, соединяющих озёра бассейнов рек Торговая, Малый Паток, Вангыр и Войвож-Сыня, неоднократно отмечали как заходящих в озеро рыб (как правило, весной), так и мигрирующих в водотоки (обычно осенью). В оз. Сыняты и в нескольких безымянных озёрах на водосборе рек Войвож-Сыня и Торговая хариус живёт постоянно, а вытекающие из этих озёр протоки служат физическим барьером для захода сюда рыб из нижерасположенных участков рек в силу большого перепада высот. В таких случаях возможен только выход хариуса из озёр. Ручьевая форма этого вида (Зиновьев, 2007) в регионе не выявлена, а её существование вызывает серьёзные сомнения; практически все ручьи и малые водотоки региона в осенне-весенний период (до 9 мес в году) перемерзают.
Весной и осенью хариус совершает местные, хотя и довольно протяжённые сезонные миграции − до нескольких десятков километров (Захарченко, 1973), временно переходя с территориального на стайный образ жизни. В предшествующий льдообразованию период он образует довольно крупные скопления, включающие от нескольких десятков до сотен и тысяч особей. Такие скопления мы визуально наблюдали в среднем течении р. Щугор в начале октября 1985 г., в верховьях р. Косью в 1993 г., на предгорных участках рек Большой Паток в 1999 г. и Малый Паток в 2002 г. Эти группировки перемещаются на зимние стации нижерасположенных участков рек в пределах одной речной системы. Можно предположить наличие хоминга у хариуса северо-востока Европы, подобно верхневолжским популяциям (Павлов и др., 2000).
Размерно-возрастной состав и рост. Биологические показатели европейского хариуса из наших сборов разных лет в водотоках и водоёмах четырёх ландшафтных комплексов Крайнего Европейского Северо-Востока различаются как при сопоставлении группировок в пределах одного ландшафтного комплекса, так и при сравнении между собой этих комплексов (табл. 2). Наибольшие средние и максимальные значения длины и массы, как правило, свойственны выборкам из рек Уральского Припечорья и Большеземельской тундры. В более доступных для рыболовства равнинных таёжных и тиманских реках эти показатели существенно меньше. Аналогичные закономерности проявляются при сопоставлении среднего возраста рыб из разных ландшафтных комплексов, доли половозрелых особей и возраста массового полового созревания. В водотоках западных склонов Урала и Большеземельской тундры доля половозрелых особей в целом достаточно велика и варьирует в зависимости от наличия/отсутствия на их водосборах нагульных озёр и конкретного модельного участка речных систем, в частности, излюбленных стаций молоди в устьях рек и мелководьях, отличающихся от предпочитаемых старшевозрастными особями. В свою очередь европейский хариус равнинных рек характеризуется невысокой долей воспроизводительной части популяций, очевидно, изымаемой рыболовами. В тиманских реках сохраняется относительно высокая доля половозрелых особей, что отражает нормальное соотношение различных возрастных классов.
Таблица 2.
Биологические показатели европейского хариуса Thymallus thymallus из разных рек Европейского Северо-Востока России
| Выборка № | Река | Число рыб, экз. | Длина (FL), мм | Масса, г | Число возрастных групп | Возрастной состав, лет | Возраст | Доля половозре-лых особей, % | Соотноше-ние самцы : : самки, % | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| средний, годы |
мода, лет | массового созревания, лет | |||||||||
| Западные склоны Северного, Приполярного и Полярного Урала | |||||||||||
| 1 | Унья | 20 | $\frac{{223.6 \pm 8.3}}{{133{\kern 1pt} - {\kern 1pt} 302}}$ | $\frac{{108.4 \pm 11.9}}{{21{\kern 1pt} - {\kern 1pt} 234}}$ | 4 | 1+−4+ | 2.3 | 2+ | 3+ | 10.0 | 40 : 60 |
| 4 | Илыч | 41 | $\frac{{344.7 \pm 8.7}}{{248{\kern 1pt} - {\kern 1pt} 445}}$ | $\frac{{364.3 \pm 26.5}}{{116{\kern 1pt} - {\kern 1pt} 814}}$ | 8 | 3+−10+ | 6.1 | 5+ | 4+ | 85.4 | 53 : 47 |
| 6 | Подчерем | 31 | $\frac{{280.8 \pm 9.5}}{{173{\kern 1pt} - {\kern 1pt} {\kern 1pt} 374}}$ | $\frac{{247.1 \pm 23.6}}{{54{\kern 1pt} - {\kern 1pt} 554}}$ | 6 | 2+−7+ | 4.4 | 5+ | 6+ | 35.5 | 68 : 32 |
| 12 | Войвож-Сыня | 43 | $\frac{{402.9 \pm 6.8}}{{254{\kern 1pt} - {\kern 1pt} 490}}$ | $\frac{{666.2 \pm 31.8}}{{147{\kern 1pt} - {\kern 1pt} 1176}}$ | 8 | 4+−14+ | 9.1 | 8+ | 6+ | 83.8 | 42 : 58 |
| 13 | Вангыр | 53 | $\frac{{416.2 \pm 4.6}}{{367{\kern 1pt} - {\kern 1pt} 480}}$ | $\frac{{706.3 \pm 21.8}}{{488{\kern 1pt} - {\kern 1pt} 1058}}$ | 7 | 5+−11+ | 7.1 | 7+ | 5+ | 100.0 | 51 : 49 |
| Печорская низменность | |||||||||||
| 21 | Светлый Вуктыл | 31 | $\frac{{181.6 \pm 7.5}}{{93{\kern 1pt} - {\kern 1pt} 292}}$ | $\frac{{76.9 \pm 9.4}}{{8{\kern 1pt} - {\kern 1pt} 275}}$ | 5 | 1+−5+ | 3.0 | 3+ | 5+ | 3.2 | 68 : 32 |
| 22 | Велью | 20 | $\frac{{193.2 \pm 9.8}}{{141{\kern 1pt} - {\kern 1pt} 325}}$ | $\frac{{88.7 \pm 19.4}}{{29{\kern 1pt} - {\kern 1pt} 364}}$ | 5 | 2+−6+ | 3.3 | 3+ | 5+ | 10.0 | 45 : 55 |
| 24 | Уса | 38 | $\frac{{256.7 \pm 7.2}}{{174{\kern 1pt} - {\kern 1pt} 373}}$ | $\frac{{195.4 \pm 15.3}}{{55{\kern 1pt} - {\kern 1pt} 507}}$ | 6 | 2+−7+ | 3.7 | 4+ | 4+ | 52.6 | 61 : 39 |
| 25 | Шарью | 48 | $\frac{{217.9 \pm 8.8}}{{148{\kern 1pt} - {\kern 1pt} 328}}$ | $\frac{{122.6 \pm 14.4}}{{29{\kern 1pt} - {\kern 1pt} 358}}$ | 6 | 2+−7+ | 3.4 | 2+ | 6+ | 16.7 | 56 : 44 |
| 26 | Ижма | 9 | $\frac{{228.2 \pm 15.1}}{{131{\kern 1pt} - {\kern 1pt} 268}}$ | $\frac{{147 \pm 22.1}}{{22{\kern 1pt} - {\kern 1pt} 226}}$ | 4 | 2+−5+ | 4.3 | 5+ | 5+ | 11.1 | 33 : 67 |
| Тиманский кряж | |||||||||||
| 28 | Локчим | 68 | $\frac{{253.7 \pm 7.6}}{{161{\kern 1pt} - {\kern 1pt} 455}}$ | $\frac{{244.7 \pm 34.9}}{{41{\kern 1pt} - {\kern 1pt} 1089}}$ | 8 | 2+−9+ | 4.9 | 5+ | 5+ | 26.5 | 27 : 73 |
| 30 | Ухта | 61 | $\frac{{171.7 \pm 7.0}}{{73{\kern 1pt} - {\kern 1pt} 341}}$ | $\frac{{43.7 \pm 8.2}}{{21{\kern 1pt} - {\kern 1pt} 185}}$ | 5 | 1+−5+ | 2.2 | 2+ | 3+ | 23.0 | 39 : 61 |
| 34 | Вымь | 61 | $\frac{{291.4 \pm 5.7}}{{210{\kern 1pt} - {\kern 1pt} 401}}$ | $\frac{{261.5 \pm 16.8}}{{101{\kern 1pt} - {\kern 1pt} 656}}$ | 6 | 3+−9+ | 4.8 | 5+ | 4+ | 36.1 | 33 : 67 |
| 37 | Вашка | 13 | $\frac{{188.1}}{{125{\kern 1pt} - {\kern 1pt} 225}}$ | $\frac{{75.1}}{{15{\kern 1pt} - {\kern 1pt} 154}}$ | 4 | 1+−4+ | 2.8 | 3+ | 3+ | 23.1 | 38 : 62 |
| 41 | Черепанка | 30 | $\frac{{256.9 \pm 4.8}}{{186{\kern 1pt} - {\kern 1pt} 318}}$ | $\frac{{180.5 \pm 10.6}}{{60{\kern 1pt} - {\kern 1pt} 330}}$ | 6 | 3+−8+ | 5.7 | 5+ | 5+ | 73.3 | 47 : 53 |
| Большеземельская тундра | |||||||||||
| 46 | Ортина | 43 | $\frac{{381.1 \pm 7.1}}{{285{\kern 1pt} - {\kern 1pt} 480}}$ | $\frac{{613.8 \pm 37.7}}{{105{\kern 1pt} - {\kern 1pt} 1244}}$ | 8 | 5+−13+ | 8.5 | 8+ | 7+ | 81.4 | 49 : 51 |
| 47 | Чёрная | 24 | $\frac{{385.5 \pm 7.7}}{{315{\kern 1pt} - {\kern 1pt} 458}}$ | $\frac{{641.1 \pm 39.5}}{{355{\kern 1pt} - {\kern 1pt} 1091}}$ | 3 | 5+−7+ | 5.9 | 6+ | 5+ | 100.0 | 67 : 33 |
| 48 | Хоседаю | 13 | $\frac{{355.3 \pm 9.6}}{{300{\kern 1pt} - {\kern 1pt} 409}}$ | $\frac{{594.6 \pm 43.2}}{{342{\kern 1pt} - {\kern 1pt} 840}}$ | 5 | 5+−9+ | 6.6 | 7+ | 5+ | 100.0 | 54 : 46 |
| 51 | Морею | 39 | $\frac{{274.6 \pm 17.0}}{{130{\kern 1pt} - {\kern 1pt} 455}}$ | $\frac{{342.2 \pm 52.6}}{{20{\kern 1pt} - {\kern 1pt} 1245}}$ | 9 | 2+−10+ | 4.9 | 2+ | 5+ | 46.2 | 56 : 44 |
| 53 | Кара | 73 | $\frac{{255.7 \pm 6.4}}{{92{\kern 1pt} - {\kern 1pt} 380}}$ | $\frac{{168.6 \pm 11.7}}{{8{\kern 1pt} - {\kern 1pt} 462}}$ | 10 | 1+−10+ | 4.9 | 4 | 6+ | 23.3 | 45 : 55 |
Примечание. Здесь и в табл. 3−5: над чертой – среднее значение и его ошибка, под чертой – пределы варьирования показателя; номера выборок см. в табл. 1 и на рис. 1. Максимальное число возрастных групп в популяциях хариуса характерно для удалённых от населённых пунктов и промышленных центров водоёмов и их участков. Исключением являются хотя и удалённые, но традиционно служащие базой для массового любительского нерегулируемого рыболовства водотоки, в частности, р. Унья (табл. 2).
Для рыб Большеземельской тундры характерно более позднее наступление половой зрелости (в возрасте 5+−7+) по сравнению с популяциями Урала, Тимана и Печорской равнины (3+−6+) (табл. 2). По данным Сидорова и Решетникова (2014), половое созревание хариуса в уральских и тиманских реках, как правило, происходит на 4−5-м годах жизни. Ещё более раннее достижение половой зрелости (в возрасте 1+) отмечено у ультра- и короткоцикловых популяций европейского хариуса Прикамья (Зиновьев и др., 2011). Относительно соотношения полов не наблюдается существенных различий ни между группировками, населяющими разные географические зоны, ни внутри них, аналогично другим регионам, в частности, на северо-западе России (Анацкий, 1996).
Закономерности распределения хариуса в разных водотоках северо-востока Европы хорошо иллюстрируют графики его возрастной структуры (рис. 2). Левая часть этих графиков отражает как отсутствие в некоторых водоёмах младших возрастных групп хариуса, так и известную селективность в отношении них использованных крючковых орудий лова. Практически полное отсутствие молоди хариуса в возрасте до 4+−5+ на участках водотоков, образующих озёрно-речные системы (реки Войвож-Сыня, Вангыр, Малый Паток, Хоседаю и Ортина), обусловлено летним нагулом неполовозрелых особей в связанных с руслом водотоков озёрах. Правая часть графиков демонстрирует наличие значительной доли рыб старшего возраста в тундровых и уральских реках (кроме Подчерема и Кожима, из которых половозрелая часть популяций в период открытой воды перемещается в малые притоки).
Рис. 2.
Возрастной состав уловов крючковыми снастями европейского хариуса Thymallus thymallus в водотоках четырёх ландшафтных комплексов Европейского Северо-Востока России: а – западные склоны Северного, Приполярного и Полярного Урала (1 – р. Унья, 6 – р. Подчерем, 13 − р. Вангыр, 18 − р. Нияю); б – Печорская низменность (21 − р. Светлый Вуктыл, 23 – р. Большая Сыня, 25 − р. Шарью, 27 − р. Енва); в – Тиманский кряж (31 – р. Сюзью, 36 − р. Мезень, 39 − р. Номбур, 40 − р. Цильма); г – Большеземельская тундра (44 − р. Ортина, 48 − р. Хоседаю, 52 − р. Силоваяха, 53 – р. Кара); номера выборок см. на рис. 1 и в табл. 1.
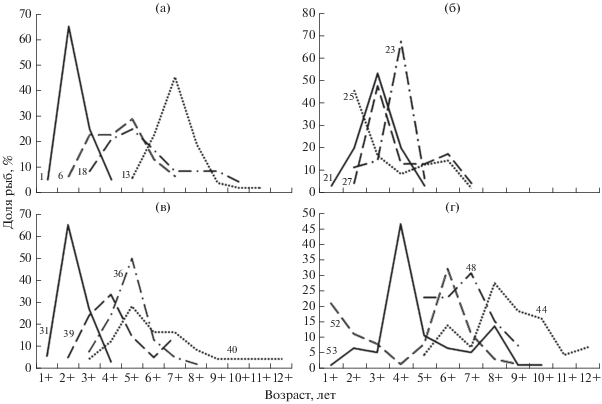
Общей чертой возрастной структуры группировок европейского хариуса, населяющего водотоки всех четырёх ландшафтных комплексов, является широкое варьирование модального возраста (табл. 2). Так, на Урале (рис. 2а) он изменяется от 2+ (в наиболее подверженной перелову р. Унья) до 7+−8+ на труднодоступных участках рек Вангыр и Войвож-Сыня (территория национального парка “Югыд ва”). В реках Большеземельской тундры (рис. 2г) модальный возраст варьирует в пределах 4+−8+. В наиболее доступных для рыболовства равнинных таёжных (рис. 2б) и тиманских реках (рис. 2в) этот показатель различается не столь значительно – от 2+ до 5+.
По темпу роста и размеру одновозрастных особей наблюдаются очевидные черты сходства исследованных популяций хариуса (рис. 3). В то же время установлен более высокий темп роста хариуса в реках северной части Приполярного Урала в бассейне р. Уса (правого и крупнейшего притока Печоры), с одной стороны, и остальных уральских, тиманских и равнинных рек, с другой стороны.
Рис. 3.
Темп роста европейского хариуса Thymallus thymallus в водоёмах Европейского Северо-Востока России: 4 − р. Илыч, 8 − р. Торговая, 9 − р. Большой Паток, 11 – оз. Сыняты, 12 − р. Войвож-Сыня, 13 − р. Вангыр, 18 − р. Нияю, 28 − р. Локчим, 30 − р. Ухта, 32 – р. Елва, 35 − р. Пижма, 38 − р. Мыла, 50 − оз. Ватьярты; номера выборок см. на рис. 1 и в табл. 1.

В ещё большей степени проявляются различия размерно-весовых показателей хариуса из разных рек и озёр при сравнении особей одного возраста. Так, длина и масса 5-летних, как правило, неполовозрелых рыб варьирует в весьма широких пределах (табл. 3). При этом величина показателей не зависит от формы хариуса – озёрной, озёрно-речной или речной. Прослеживается зависимость скорости роста рыб от района их обитания: максимальный темп роста характерен для группировок европейского хариуса, населяющих разного типа водоёмы Приполярного и Полярного Урала. Замедленный по сравнению с ними рост наблюдается главным образом в равнинных таёжных речных системах, а также в некоторых реках и озёрах Северного Урала. Случаи замедленного роста хариуса региона, в частности в р. Черепанка (бассейн р. Цильма), описаны в литературе (Сидоров, Решетников, 2014). При этом индивидуальная изменчивость размерно-весовых показателей оказалась более высокой в уральских реках.
Таблица 3.
Размерно-весовые показатели особей европейского хариуса Thymallus thymallus в возрасте 4+ в разных реках и озёрах Европейского Северо-Востока России
| Выборка № | Водоём | Число рыб, экз. | Длина (FL), мм | Масса, г |
|---|---|---|---|---|
| Западные склоны Северного, Приполярного и Полярного Урала | ||||
| 3 | р. Печора, истоки | 13 | $\frac{{253.6 \pm 4.1}}{{226{\kern 1pt} - {\kern 1pt} 281}}$ | $\frac{{159.8 \pm 9.3}}{{105{\kern 1pt} - {\kern 1pt} 229}}$ |
| 10 | оз. Номты | 36 | $\frac{{212.4 \pm 1.4}}{{201{\kern 1pt} - {\kern 1pt} 239}}$ | $\frac{{89.4 \pm 2.0}}{{73{\kern 1pt} - {\kern 1pt} 123}}$ |
| 12 | р. Войвож-Сыня | 32 | $\frac{{309.9 \pm 3.4}}{{273{\kern 1pt} - {\kern 1pt} 341}}$ | $\frac{{284.0 \pm 9.7}}{{189{\kern 1pt} - {\kern 1pt} 383}}$ |
| 14 | р. Косью | 21 | $\frac{{291.3 \pm 1.9}}{{275{\kern 1pt} - {\kern 1pt} 310}}$ | $\frac{{237.1 \pm 4.5}}{{203{\kern 1pt} - {\kern 1pt} 207}}$ |
| 15 | р. Кожим | 19 | $\frac{{299.3 \pm 5.3}}{{256{\kern 1pt} - {\kern 1pt} 380}}$ | $\frac{{291.2 \pm 18.6}}{{192{\kern 1pt} - {\kern 1pt} 530}}$ |
| 16 | оз. Пагаты | 32 | $\frac{{270.0 \pm 4.3}}{{220{\kern 1pt} - {\kern 1pt} 348}}$ | $\frac{{193.8 \pm 10.6}}{{87{\kern 1pt} - {\kern 1pt} 419}}$ |
| 17 | оз. Хойлаты | 13 | $\frac{{313.4 \pm 5.2}}{{280{\kern 1pt} - {\kern 1pt} 344}}$ | $\frac{{372.5 \pm 22.2}}{{234{\kern 1pt} - {\kern 1pt} 552}}$ |
| 18 | р. Нияю | 14 | $\frac{{246.0 \pm 5.4}}{{215{\kern 1pt} - {\kern 1pt} 280}}$ | $\frac{{178.8 \pm 11.9}}{{124{\kern 1pt} - {\kern 1pt} 254}}$ |
| 19 | оз. Усваты | 18 | $\frac{{278.1 \pm 5.1}}{{251{\kern 1pt} - {\kern 1pt} 333}}$ | $\frac{{224.4 \pm 15.3}}{{156{\kern 1pt} - {\kern 1pt} 413}}$ |
| 20 | оз. Плаунты | 5 | $\frac{{242.2 \pm 7.9}}{{227{\kern 1pt} - {\kern 1pt} 265}}$ | $\frac{{149.4 \pm 15.1}}{{116{\kern 1pt} - {\kern 1pt} 193}}$ |
| Тиманский кряж | ||||
| 39 | р. Номбур | 7 | $\frac{{208.0 \pm 8.1}}{{180{\kern 1pt} - {\kern 1pt} 235}}$ | $\frac{{95.0 \pm 12.3}}{{55{\kern 1pt} - {\kern 1pt} 150}}$ |
| Большеземельская тундра | ||||
| 53 | р. Кара | 33 | $\frac{{238.2 \pm 2.0}}{{216{\kern 1pt} - {\kern 1pt} 263}}$ | $\frac{{122.1 \pm 3.3}}{{82{\kern 1pt} - {\kern 1pt} 165}}$ |
| 47 | оз. Наульто | 18 | $\frac{{232.8 \pm 4.1}}{{201{\kern 1pt} - {\kern 1pt} 259}}$ | $\frac{{121.5 \pm 7.2}}{{72{\kern 1pt} - {\kern 1pt} 171}}$ |
| 44 | р. Ортина | 7 | $\frac{{258.0 \pm 2.2}}{{248{\kern 1pt} - {\kern 1pt} 265}}$ | $\frac{{160.7 \pm 13.9}}{{125{\kern 1pt} - {\kern 1pt} 225}}$ |
Аналогичные закономерности наблюдаются и в отношении неоднократно принимавших участие в нересте 9-летних рыб (табл. 4). Весьма существенно различаются средние, минимальные и наибольшие значения длины и особенно массы хариуса как при попарном сравнении популяций разных водоёмов, так и при сопоставлении группировок из разных ландшафтных комплексов. Как и у 5-летних рыб, наибольшие размеры отмечены в популяциях Приполярного и Полярного Урала. Эти данные хорошо согласуются с результатами сопоставления темпа роста разных популяций европейского хариуса из бассейна р. Уса − крупнейшего печорского притока (Скоринова и др., 2014).
Таблица 4.
Размерно-весовые показатели особей европейского хариуса Thymallus thymallus в возрасте 8+ в разных реках и озёрах Европейского Северо-Востока России
| Выборка № | Водоём | Число рыб, экз. | Длина (FL), мм | Масса, г |
|---|---|---|---|---|
| Западные склоны Северного, Приполярного и Полярного Урала | ||||
| 2 | р. Елма | 5 | $\frac{{375.6 \pm 4.2}}{{360{\kern 1pt} - {\kern 1pt} 385}}$ | $\frac{{497.0 \pm 11.3}}{{468{\kern 1pt} - {\kern 1pt} 517}}$ |
| 6 | р. Подчерем | 7 | $\frac{{376.4 \pm 4.4}}{{360{\kern 1pt} - {\kern 1pt} 393}}$ | $\frac{{581.1 \pm 24.5}}{{503{\kern 1pt} - {\kern 1pt} 680}}$ |
| 8 | р. Торговая | 26 | $\frac{{364.6 \pm 2.6}}{{328{\kern 1pt} - {\kern 1pt} 379}}$ | $\frac{{463.3 \pm 8.6}}{{327{\kern 1pt} - {\kern 1pt} 519}}$ |
| 9 | р. Большой Паток | 13 | $\frac{{365.9 \pm 1.9}}{{356{\kern 1pt} - {\kern 1pt} 382}}$ | $\frac{{488.9 \pm 11.1}}{{411{\kern 1pt} - {\kern 1pt} 551}}$ |
| 10 | оз. Номты | 17 | $\frac{{307.0 \pm 2.9}}{{283{\kern 1pt} - {\kern 1pt} 324}}$ | $\frac{{280.4 \pm 8.8}}{{211{\kern 1pt} - {\kern 1pt} 338}}$ |
| 11 | оз. Сыняты | 20 | $\frac{{381.3 \pm 1.8}}{{370{\kern 1pt} - {\kern 1pt} 399}}$ | $\frac{{570.7 \pm 14.4}}{{493{\kern 1pt} - {\kern 1pt} 711}}$ |
| 12 | р. Войвож-Сыня | 17 | $\frac{{422.1 \pm 2.1}}{{410{\kern 1pt} - {\kern 1pt} 439}}$ | $\frac{{733.3 \pm 18.6}}{{612{\kern 1pt} - {\kern 1pt} 860}}$ |
| 13 | р. Вангыр | 10 | $\frac{{464.4 \pm 3.5}}{{432{\kern 1pt} - {\kern 1pt} 485}}$ | $\frac{{1040.3 \pm 28.3}}{{871{\kern 1pt} - {\kern 1pt} 1221}}$ |
| 14 | р. Косью | 16 | $\frac{{438.1 \pm 4.7}}{{405{\kern 1pt} - {\kern 1pt} 481}}$ | $\frac{{797.1 \pm 25.0}}{{631{\kern 1pt} - {\kern 1pt} 1012}}$ |
| 15 | р. Кожим | 6 | $\frac{{423.9 \pm 8.0}}{{388{\kern 1pt} - {\kern 1pt} 447}}$ | $\frac{{863.3 \pm 41.5}}{{700{\kern 1pt} - {\kern 1pt} 950}}$ |
| 18 | р. Нияю | 7 | $\frac{{461.3 \pm 4.7}}{{450{\kern 1pt} - {\kern 1pt} 479}}$ | $\frac{{1060.1 \pm 49.5}}{{923{\kern 1pt} - {\kern 1pt} 1290}}$ |
| Тиманский кряж | ||||
| 42 | р. Тобыш | 7 | $\frac{{311.1 \pm 10.5}}{{290{\kern 1pt} - {\kern 1pt} 365}}$ | $\frac{{306.9 \pm 33.1}}{{233{\kern 1pt} - {\kern 1pt} 478}}$ |
| Большеземельская тундра | ||||
| 43 | р. Ваглейвис | 8 | $\frac{{340.4 \pm 4.3}}{{324{\kern 1pt} - {\kern 1pt} 360}}$ | $\frac{{417.8 \pm 15.2}}{{360{\kern 1pt} - {\kern 1pt} 496}}$ |
| 49 | оз. Лангутаты | 5 | $\frac{{397.6 \pm 11.3}}{{376{\kern 1pt} - {\kern 1pt} 440}}$ | $\frac{{765.6 \pm 93.8}}{{613{\kern 1pt} - {\kern 1pt} 1135}}$ |
Не выявлена связь различий скорости роста хариуса в тех или иных речных системах с величиной биомассы бентоса и излюбленных групп кормовых организмов. Значения этих показателей в разных водотоках Урала и Тимана варьируют в весьма широких пределах и подвержены значительным сезонным и межгодовым изменениям (Шубина, 2006). Можно предположить, что темп роста хариуса контролируется не столько трофологическими, сколько наследственными и эколого-поведенческими механизмами (Михеев, 2006).
Данные по длине и массе 9-летних особей хариуса р. Большой Паток в в 1998−2018 гг. (табл. 5) свидетельствуют о том, что и межгодовые различия темпа роста весьма велики. В 2003−2015 гг. на Крайнем Северо-Востоке Европы наблюдалось незначительное повышение среднегодовой температуры воздуха; в этот период (за исключением 2010 и 2014 гг.) сумма положительных (летних) значений температуры стабильно превышала 1100°С/сут (Каверин и др., 2017). При этом размерно-весовые показатели хариуса проявляли в эти годы высокую индивидуальную изменчивость, а средние значения в целом соответствовали тому же уровню, что и в более холодные годы (1999, 2002, 2010 и 2014 гг.), когда сумма положительных значений температуры только один раз превысила 890°С/сут. Это свидетельствует об относительной независимости темпа роста европейского хариуса региона от погодных условий.
Таблица 5.
Размерно-весовые показатели особей европейского хариуса Thymallus thymallus в возрасте 8+ в р. Большой Паток в 1998−2018 гг.
| Период отлова (месяц, год) |
Число рыб, экз. | Длина (FL), мм | Масса, г |
|---|---|---|---|
| IV.1998 | 20 | $\frac{{381.0 \pm 2.8}}{{364{\kern 1pt} - {\kern 1pt} 416}}$ | $\frac{{606.5 \pm 14.7}}{{498{\kern 1pt} - {\kern 1pt} 803}}$ |
| V.1999 | 13 | $\frac{{378.4 \pm 3.7}}{{360{\kern 1pt} - {\kern 1pt} 406}}$ | $\frac{{540.5 \pm 14.9}}{{446{\kern 1pt} - {\kern 1pt} 615}}$ |
| VIII.2000 | 5 | $\frac{{405.8 \pm 7.1}}{{383{\kern 1pt} - {\kern 1pt} 420}}$ | $\frac{{673.0 \pm 40.0}}{{545{\kern 1pt} - {\kern 1pt} 768}}$ |
| V.2003 | 21 | $\frac{{366.6 \pm 3.2}}{{343{\kern 1pt} - {\kern 1pt} 395}}$ | $\frac{{503.0 \pm 17.9}}{{380{\kern 1pt} - {\kern 1pt} 701}}$ |
| XI.2009 | 9 | $\frac{{370.9 \pm 5.8}}{{350{\kern 1pt} - {\kern 1pt} 401}}$ | $\frac{{528.2 \pm 23.2}}{{451{\kern 1pt} - {\kern 1pt} 665}}$ |
| III.2010 | 8 | $\frac{{344.4 \pm 3.2}}{{332{\kern 1pt} - {\kern 1pt} 359}}$ | $\frac{{385.9 \pm 29.9}}{{230{\kern 1pt} - {\kern 1pt} 520}}$ |
| XI.2010 | 6 | $\frac{{366.5 \pm 7.9}}{{339{\kern 1pt} - {\kern 1pt} 388}}$ | $\frac{{536.2 \pm 31.7}}{{408{\kern 1pt} - {\kern 1pt} 608}}$ |
| III.2011 | 15 | $\frac{{346.9 \pm 7.7}}{{254{\kern 1pt} - {\kern 1pt} 378}}$ | $\frac{{434.0 \pm 23.1}}{{227{\kern 1pt} - {\kern 1pt} 592}}$ |
| III.2012 | 17 | $\frac{{365.6 \pm 3.5}}{{345{\kern 1pt} - {\kern 1pt} 392}}$ | $\frac{{474.1 \pm 13.1}}{{370{\kern 1pt} - {\kern 1pt} 573}}$ |
| IV.2013 | 16 | $\frac{{373.8 \pm 3.0}}{{355{\kern 1pt} - {\kern 1pt} 392}}$ | $\frac{{482.4 \pm 14.3}}{{404{\kern 1pt} - {\kern 1pt} 565}}$ |
| XI.2015 | 6 | $\frac{{376.6 \pm 7.4}}{{346{\kern 1pt} - {\kern 1pt} 401}}$ | $\frac{{554.7 \pm 33.0}}{{452{\kern 1pt} - {\kern 1pt} 667}}$ |
| IX.2016 | 13 | $\frac{{365.9 \pm 1.9}}{{356{\kern 1pt} - {\kern 1pt} 382}}$ | $\frac{{488.9 \pm 11.1}}{{411{\kern 1pt} - {\kern 1pt} 551}}$ |
| IV.2018 | 13 | $\frac{{373.8 \pm 3.2}}{{350{\kern 1pt} - {\kern 1pt} 391}}$ | $\frac{{518.5 \pm 16.8}}{{376{\kern 1pt} - {\kern 1pt} 600}}$ |
Полиморфизм трансферринов сыворотки крови. Европейский хариус в ареале представлен многими популяциями, обитающими в водоёмах с разной степенью географической изоляции. Генетическая структура его, как и большинства лососевидных рыб, неоднородна, о чём свидетельствуют данные электрофоретических спектров белков сыворотки, в зоне глобулинов которых обнаружены генетические варианты белков, идентифицированных как трансферрины. Анализ распределения частот генотипов полиморфной двухаллельной системы трансферринов сыворотки разных группировок показал, что популяции хариуса, по крайней мере, крупной речной системы Печоры можно объединить в два кластера – уральский и тиманский (табл. 6). Интересно отметить, что в сыворотке крови хариуса из р. Тулома Кольского п-ова присутствует лишь один генетический вариант трансферрина (АА), который у хариуса из бассейнов Печоры и Северной Двины является редким (2–4%). При этом на период отбора проб частоты аллелей трансферрина у печорских популяций хариуса соответствуют ожидаемым согласно уравнению Харди−Вайнберга значениям, что характерно для больших равновесных панмиктических популяций.
Таблица 6.
Доля разных генотипов трансферрина у европейского хариуса Thymallus thymallus из разных рек
| Регион, река | Выборка № | Число рыб, экз. | Тип трансферрина, % | χ2 (df = 1) |
||
|---|---|---|---|---|---|---|
| АА | АВ | ВВ | ||||
| Урал: | ||||||
| Щугор | 7 | 137 | 3.5/2.1 | 21.9/24.9 | 74.6/72.9 | 2.0 |
| Кожим | 15 | 61 | 3.3/2.4 | 24.6/26.3 | 72.1/71.2 | 0.3 |
| Тиман: | ||||||
| Вымь | 34 | 43 | 2.3/0.3 | 7.0/10.9 | 90.7/88.7 | 5.6 |
| Пижма | 35 | 68 | 2.0/0.5 | 10.3/13.3 | 87.7/86.2 | 3.4 |
| Кольский п-ов | ||||||
| Тулома | 54 | 47 | 100.0/100.0 | 0/0 | 0/0 | |
Эти данные хорошо согласуются с результатами анализа 37 аллозимных локусов, в том числе и трансферринов, и с оценкой степени генетической дифференциации европейского хариуса в восточной части ареала − в бассейнах Баренцева и Белого морей (Титов, Студенов, 2005).
ЗАКЛЮЧЕНИЕ
В результате многолетнего изучения распространения, популяционной структуры и особенностей экологии европейского хариуса в равнинных таёжных, тундровых и горных реках и озёрах Крайнего Северо-Востока России показано, что на большей части этой территории состояние и разнообразие единой по происхождению группировки популяций остаётся на близком к естественному уровню. Сокращение возрастного ряда и преобладание в популяциях медленнорастущих рыб как результат регулярного изъятия крупных и быстрорастущих особей (Коротаева, 2003; Зиновьев и др., 2011; Михеев и др., 2014) наблюдается только в относительно густонаселённых и доступных для отлова реках, тогда как в большинстве водотоков и озёр сохраняется нормальная структура популяций.
Европейский хариус, как и другие виды рода Thymallus, проявляют значительную внутривидовую разнокачественность (Павлов и др., 2000). В рассматриваемом регионе многочисленные популяции вида существенно различаются по целому ряду биологических особенностей и обнаруживают генетические различия. Дальнейшие исследования закономерностей разноуровневой дифференциации популяций европейского хариуса на Европейском Северо-Востоке России позволят углубить имеющиеся представления о микроэволюционных процессах у рыб. Особенно перспективным представляется изучение в качестве модельных относительно изолированных популяций ледниковых и подпружных озёр района водораздела бассейнов сибирских и европейских рек Приполярного и Полярного Урала.
Полученные данные не только существенно расширяют имеющуюся информацию о распространении и популяционной структуре вида, но и могут послужить базовым материалом при организации мероприятий по сохранению и восстановлению депрессивных и подверженных риску исчезновения популяций в Европе. Использование в рамках мероприятий по искусственному воспроизводству сохранившихся донорских популяций хариуса Европейского Северо-Востока России, включающего широкий спектр климатических и географических зон, позволит расширить возможности восстановления вида на ранее уже утраченных участках ареала.
Список литературы
Анацкий С.Ю. 1996. Фенотипическая изменчивость хариуса, Thymallus thymallus (L.), в водоемах Северо-Запада России: Автореф. дис. … канд. биол. наук. СПб.: СПбГУ, 16 с.
Аськеев А.О., Аськеев О.В., Аськеев И.О., Монахов С.П. 2016. Численность, встречаемость, историческое и современное распространение европейского хариуса и налима в градиентах окружающей среды в реках Республики Татарстан // Рос. журн. приклад. экологии. № 4. С. 17–22.
Богданов В.Г., Мельниченко И.П. 2010. Граница пресноводной европейской и азиатской ихтиофауны в арктической части Полярного Урала // Экология. № 5. С. 372–377.
Захаров А.Б., Бознак Э.И. 2019. Рыбное население водотоков Тимана. Сыктывкар: Изд-во ФИЦ Коми НЦ УрО РАН, 184 с.
Захарченко Г.М. 1973. О миграциях хариуса Thymallus thymallus (L.) в верховьях Печоры // Вопр. ихтиологии. Т. 13. Вып. 4 (81). С. 744–745.
Зиновьев Е.А. 1979. Морфологическая характеристика двух видов хариусов реки Кожим // Сб. науч. тр. Перм. лаб. ГосНИОРХ. Вып. 2. С. 69–78.
Зиновьев Е.А. 2007. Таксономическая и экологическая структура хариусовых рыб // Сб. науч. тр. ГосНИОРХ. Вып. 337. С. 425–452
Зиновьев Е.А., Бакланов М.А., Боталова И.Н. 2011. Суперкарликовая популяция хариуса реки Язовой // Вестн. УдмуртГУ. Биология. Науки о Земле. Вып. 4. С. 71–77.
Каверин Д.А., Пастухов А.В., Новаковский А.Б. 2017. Динамика глубины сезонного протаивания тундровых мерзлотных почв (на примере площадки циркумполярного мониторинга деятельного слоя в европейской России) // Криосфера Земли. Т. 21. № 6. С. 35−44. https://doi.org/10.21782/KZ1560-7496-2017-6(35-44)
Коротаева С.Э. 2003. Эколого-популяционные особенности хариусов Приуралья: Автореф. дис. … канд. биол. наук. Пермь: ПермГУ, 23 с.
Мартынов В.Г. 2007. Атлантический лосось (Salmo salar L.) на Севере России. Екатеринбург: Изд-во УрО РАН, 416 с.
Мина М.В., Клевезаль Г.А. 1976. Рост животных. М.: Наука, 292 с.
Михеев В.Н. 2006. Неоднородность среды и трофические отношения у рыб. М.: Наука, 191 с.
Михеев П.Б., Петренко Н.Г., Михеева О.И., Семенов В.В. 2014. Современное состояние поголовья европейского хариуса на особо охраняемом заповедном участке р. Вишера и в его притоках // Тр. заповедника “Вишерский” за 2011−2014 гг. Пермь: Изд-е Госзаповедника “Вишерский”. С. 109–123.
Новоселов А.П. 2000. Современное состояние рыбной части сообществ в водоемах европейского северо-востока России: Автореф. дис. … докт. биол. наук. М.: ИПЭЭ РАН, 50 с.
Павлов Д.С., Кузищин К.В., Легкий Б.П. и др. 2000. Сравнительный морфологический анализ природных популяций европейского хариуса Thymallus thymallus верхневолжского бассейна // Вопр. ихтиологии. Т. 40. № 4. С. 477–485.
Пономарев В.И. 2017. Рыбы озер западных склонов Приполярного и Полярного Урала // Изв. Коми НЦ УрО РАН. Вып. 2 (30). С. 16–29.
Пономарев В.И. 2019. Распространение рыб в малых озерах горной полосы бассейна реки Косью (Приполярный Урал) // Вестн. ПермГУ. Сер. Биология. Вып. 2. С. 187−196. https://doi.org/10.17072/1994-9952-2019-2-187-196
Пономарев В.И., Лоскутова О.А., Серегина Е.Ю. 1998. Биотопическое распределение хариуса реки Щугор в подледный период // Тр. Коми НЦ УрО РАН. № 157. С. 82–89.
Правдин И.Ф. 1966. Руководство по изучению рыб. М.: Пищ. пром-сть, 376 с.
Решетников Ю.С. 1980. Экология и систематика сиговых рыб. М.: Наука, 300 с.
Рыбы в заповедниках России. 2010. Т. 1. Пресноводные рыбы / Под ред. Решетникова Ю.С. М.: Т-во науч. изд. КМК, 627 с.
Сидоров Г.П. 1974. Рыбные ресурсы Большеземельской тундры. Л.: Наука, 164 с.
Сидоров Г.П., Решетников Ю.С. 2014. Лососеобразные рыбы водоемов европейского северо-востока. М.: Т-во науч. изд. КМК, 346 с.
Скоринова А.В., Зиновьев Е.А., Коротаева С.Э. 2014. О структуре популяций и росте европейского хариуса бассейна реки Усы // Вестн. ПермГУ. Сер. Биология. Вып. 1. С. 18–20.
Соловкина Л.Н. 1975. Рыбные ресурсы Коми АССР. Сыктывкар: Коми кн. изд-во, 168 с.
Титов С.В., Студенов И.И. 2005. Генетическая дифференциация европейского хариуса (Thymallus thymallus L.) из рек восточной части ареала (бассейны Баренцева и Белого морей) // Лососевидные рыбы Восточной Фенноскандии. Петрозаводск: Изд-во КарНЦ РАН. С. 178−190.
Шубина В.Н. 2006. Бентос лососевых рек Урала и Тимана. СПб.: Наука, 401 с.
Aotsuka T., Asami T. 1979. A simplified apparatus for vertical stab gel electrophoresis // Jpn. J. Genetic. V. 54. № 5. P. 397−400.
Carlstein M. 2004. Growth and survival of European grayling reared at different stocking densities // Aquacult. Int. V. 3. P. 260–264. https://doi.org/10.1007/BF00118108
Fieseler C.A., Wolter C. 2006. A fish-based typology of small temperate rivers in the Northestern lowlands of Germany // Limnologica. V. 36. № 1. P. 2−16.
Koskinen M.T., Primmer C.R. 2001. High throughput analysis of 17 microsatellite loci grayling (Thymallus spp. Salmonidae) // Conserv. Genetics. № 2. P. 173–177. https://doi.org/10.1023/A:1011886412830
Maire A., Laffaille P., Buisson A. 2017. Identification of priority areas for the conservation of stream fish assemblage: implications for river management in France // River Res. Appl. V. 33. № 4. P. 524−537. https://doi.org/10.1002/rra.3107
Northcote T.G. 1995. Comparative biology and management of Arctic and European grayling (Salmonidae, Thymallus) // Rev. Fish Biol. Fish. V. 5. P. 141–194. https://doi.org/10.1007/BF00179755
Persat H. 1996. Threatened populations and conservation of European grayling, Thymallus thymallus (L., 1758) // Conservation of endangered freshwater fish in Europe / Eds. Kirchhofer A., Hefti D. Basel: Birkhauser Verlag. P. 233–247. https://doi.org/10.1007/978-3-0348-9014-4_23
Uiblein F., Jagsch A., Honsig-Erlenburg W., Weiss S. 2001. Status, habitat use, and vulnerability of the European grayling in Austrian waters // J. Fish Biol. V. 59. P. 223–247. https://doi.org/10.1111/j.1095-8649.2001.tb01388.x
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии