

Вопросы ихтиологии, 2021, T. 61, № 3, стр. 314-325
Обнаружение арктического гольца Salvelinus alpinus (Salmonidae) и пестроногого подкаменщика Cottus cf. poecilopus (Cottidae) в бассейне Верхней Ангары на водоразделе с системой Лены: новые данные о путях вселения представителей ленской ихтиофауны в бассейн Байкала
С. С. Алексеев 1, *, В. П. Самусенок 1, 2, А. Н. Матвеев 1, 2, Р. С. Андреев 2, И. В. Самусенок 2, А. И. Вокин 2, А. Л. Юрьев 1, 2
1 Институт биологии развития РАН – ИБР РАН
Москва, Россия
2 Иркутский государственный университет
Иркутск, Россия
* E-mail: alekseyev@mail.ru
Поступила в редакцию 27.05.2020
После доработки 05.06.2020
Принята к публикации 08.06.2020
Аннотация
В оз. Амут (бассейн р. Верхняя Ангара) обнаружена третья в бассейне Байкала популяция арктического гольца Salvelinus alpinus – вида, предположительно вселившегося из бассейна Лены, а также другой представитель ленской ихтиофауны − пестроногий подкаменщик Cottus cf. poecilopus. Это озеро расположено на водоразделе с р. Правая Мама (бассейн Витима) вблизи её русла. Структура рельефа водораздельного участка, на котором расположено оз. Амут, указывает на высокую вероятность проникновения ленских видов в бассейн Байкала в этом месте. Приводятся данные о морфологии, возрасте, росте, питании арктического гольца из оз. Амут в сравнении с другими гольцами бассейна Байкала. В связи с новыми находками обсуждаются вопросы палеогеографии плейстоцена Байкальского региона, связанные с историей формирования его ихтиофауны.
В формировании гидрофауны оз. Байкал и водоёмов его бассейна важную роль играло вселение видов из соседних бассейнов (Петров, 2003; Матвеев, Самусенок, 2008; Ситникова, Прозорова, 2008; Пудовкина и др., 2014; Аров, Мишарина, 2018). К числу таких видов относится арктический голец Salvelinus alpinus. Обсуждались разные возможные варианты его проникновения в бассейн Байкала: из бассейнов Лены, Енисея и Амура (Савваитова и др., 1977). В последние десятилетия по мере накопления данных о его широком распространении в верховьях бассейнов Витима, Олёкмы и Чаи (притоки Лены) (Алексеев и др., 1999; Alekseyev et al., 2002, 2009) стало очевидным, что заселение происходило из бассейна Лены, скорее всего, через водораздел (Пронин и др., 2007), однако конкретные пути оставались неясными.
К настоящему моменту в Забайкалье обнаружено около 30 популяций арктического гольца в бассейне Лены, а в бассейне Байкала до недавнего времени были известны только две – в озёрах Фролиха и Светлинское. В оз. Фролиха голец был обнаружен почти два с половиной века назад (Georgi, 1775) и первоначально описан как вид Salmo erythrinus, а позже − как подвид арктического гольца Salvelinus alpinus erythrinus (даватчан) (Berg, 1907). Согласно современным данным, для выделения его в отдельный подвид нет оснований (Алексеев и др., 2000). Эта популяция арктического гольца до середины прошлого века оставалась единственной известной во всём Забайкалье и неоднократно изучалась (Мухомедияров, 1942; Редкозубов, Мовчан, 1974; Савваитова и др., 1977). Популяция оз. Светлинское открыта относительно недавно (Самусенок и др., 2006). Обе популяции удалены от границы между бассейнами Лены и Байкала, и их положение не позволяет реконструировать пути вселения гольцов в последний. В то же время в приближенных к водоразделу участках бассейна Байкала обнаружены популяции других представителей ленской ихтиофауны − байкалоленского хариуса Thymallus baicalolenensis (Матвеев и др., 2005, 2006; Weiss et al., 2006; Пронин и др., 2007; Раднаев, 2007; Вокин, 2008; Книжин и др., 2008) и пестроногого подкаменщика Cottus cf. poecilopus (Андреев и др., 2020).
В 2010 г. в расположенном в верховьях бассейна Верхней Ангары, на водоразделе с бассейном Лены, оз. Амут мы обнаружили третью в бассейне Байкала популяцию арктического гольца. Были изучены изменчивость митохондриальной ДНК (мтДНК) (Гордеева и др., 2018) и линейно-весовой рост (Матвеев и др., 2020) этих гольцов, иные сведения по ним не публиковались. В этом же озере отмечен пестроногий подкаменщик.
Цель настоящей работы – сообщить об этих находках, дать морфоэкологическое описание арктического гольца из оз. Амут и обсудить возможные пути проникновения рыб, населяющих верховья бассейна Лены, в бассейн Байкала через водораздел.
МАТЕРИАЛ И МЕТОДИКА
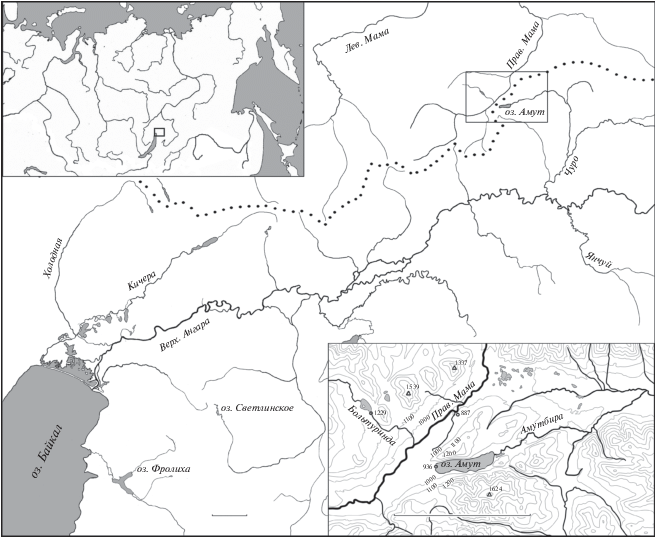
Озеро Амут (56°32ʹ с.ш. 112°00ʹ в.д., не путать с одноимённым озером в верховьях бассейна р. Баргузин) размером 4.1 × 0.7 км, глубиной до 29 м расположено в бассейне Верхней Ангары на высоте 936 м над уровнем моря, вблизи байкало-ленского водораздела (рис. 1). Оно вытянуто в широтном направлении, южный и северный берега крутые, покрытые лесом и курумниками. Мелководная зона практически не выражена, береговой свал начинается в 2−5 м от береговой линии, небольшое мелководье имеется только в западной оконечности озера. Максимальные глубины приурочены к восточной части озера, из неё вытекает р. Амутбира, которая впадает в р. Уклонь, а та – в р. Чуро, приток Верхней Ангары. Западная оконечность озера находится в 2.3 км от русла р. Правая Мама (бассейн Витима, правого притока Лены), от которого отделена выположенным водораздельным участком. Собственно линия водораздела между бассейнами пролегает примерно в 1 км от озера. Примерно в 5 км к северу от озера находится похожий низкий участок водораздела между Правой Мамой и притоком Амутбиры. Помимо арктического гольца в озере отмечены озёрный Rhynchocypris percnurus и речной Phoxinus phoxinus гольяны, сибирский голец Barbatula toni, налим Lota lota и пестроногий подкаменщик. В 6 км от оз. Амут на притоке Правой Мамы р. Больтуринда расположено мелководное (глубина до 5 м) и потому непригодное для арктического гольца озеро, в котором обитают байкалоленский хариус, тупорылый ленок Brachymystax tumensis, озёрный и речной гольяны, сибирский голец.
Рис. 1.
Карта-схема центрального участка байкало-ленского водораздела (···) и района оз. Амут. Масштаб: 10 км.
Отлов проводили 30.07−05.08.2010 г. донными жаберными сетями с ячеёй 14, 18, 20, 25, 30, 35 мм, у свежих гольцов (n = 88) измеряли длину тела по Смитту (FL), определяли массу, пол, стадию зрелости (Правдин, 1966). Возраст определяли по отолитам, у одной особи – по позвонку. У фиксированных формалином рыб оценивали 10 меристических признаков (Алексеев и др., 2000), длину наибольшей жаберной тычинки (lsb) и длину головы (c). Для сравнения с гольцами из других популяций бассейна Байкала определяли коэффициент различия Майра CD (Mayr, 1963), показатель сходства rzh, Животовского (1979) и значимость различий по критерию Манна−Уитни. Проводили анализ главных компонент (АГК) по 10 счётным признакам. Использовали стандартизованные данные, собственные векторы вычисляли по вариационно-ковариационной матрице, длина собственного вектора равнялась корню квадратному из собственного значения. Вычисления выполняли в программе StatSoft STATISTICA, версия 12. Также использованы наши данные по арктическому гольцу из оз. Фролиха: изученная ранее выборка (n = 20: Алексеев и др., 2000) дополнена 18 экз. из сборов 1998 и 2009 гг. и использована для оценки меристических признаков, длины жаберных тычинок и проведения АГК.
Морфоэкологический анализ пестроногих подкаменщиков провести не удалось, поскольку в озере этот вид малочислен и встречался только в полупереваренном состоянии в желудках гольцов, но не в сетных уловах.
РЕЗУЛЬТАТЫ
Длина арктических гольцов в выборке 14−48 см, масса 21−1022 г, возраст 3−13, преимущественно 7−8 лет. Также была найдена высушенная рыбаками голова гольца, по которой определили его примерную длину (FL ~ 55 см), и по позвонку − возраст (14 лет). Тело невысокое, прогонистое, плавники короткие, голова коническая, рот обычно конечный (рис. 2, 3). Средние и крупные особи сходны по внешнему виду с гольцами из оз. Фролиха (рис. 2б). У особей небольших размеров (FL < 30 см) челюсти не массивные, у некоторых рот полуверхний (рис. 3б) – вариант, характерный для форм-планктофагов и не отмеченный у гольцов из озёр Фролиха и Светлинское (рис. 2б–2г, 3в, 3г). Особи с полунижним ртом, характерным для многих карликовых (рис. 3г) и части мелких гольцов из оз. Светлинское, не обнаружены. Верхняя челюсть заходит за вертикаль заднего края глаза при FL > 33−35 см. Взрослые особи в период наблюдений неяркие, с серыми спиной и боками и серо-розовым брюшком, молодь серебристо-серая.
Рис. 2.
Арктические гольцы Salvelinus alpinus из популяций бассейна Байкала: а − оз. Амут (FL 45 см), б – оз. Фролиха (FL 37 см); в, г – оз. Светлинское (в − мелкая форма (FL 33 см), г – карликовая форма (FL 13 см)).

Рис. 3.
Положение рта у арктических гольцов Salvelinus alpinus из популяций бассейна Байкала: а, б − оз. Амут (а − конечный рот (типичный вариант), FL 24 см; б − полуверхний рот (редкий вариант), FL 24 см); в – оз. Фролиха, конечный рот (типичный вариант), FL 27 см; г – оз. Светлинское (карликовая форма), полунижний рот (распространённый вариант), FL 18 см.

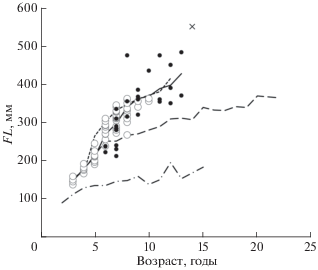
Темп роста выше, чем у гольцов мелкой формы из оз. Светлинское и у всех мелких форм гольцов из бассейна Лены, и близок к темпу роста гольцов из оз. Фролиха. При этом выделяются несколько отстающих в росте половозрелых или созревающих особей (самцы с гонадами III и самки с гонадами II−III стадий зрелости FL 21−24 см в возрасте 6−7 лет), а также несколько особей с наиболее быстрым ростом, достигших в возрасте 8−14 лет FL > 43 см (рис. 4). Последние крупнее всех гольцов, отмечавшихся в оз. Фролиха (Мухомедияров, 1942; Савваитова и др., 1977; наши данные), и сходны по размерам с гольцами крупной формы из таких забайкальских озёр, как Даватчан, Кирялта-3, 4, Токко и другие. Эти данные указывают на определённую внутрипопуляционную гетерогенность темпа роста, но, учитывая небольшое число в выборке как особей с низким темпом роста, так и особей старших возрастных групп, в которых намечается размерная дифференциация быстрорастущих рыб, не дают веских оснований предполагать наличие симпатрических размерных форм. Гольцов из оз. Амут можно рассматривать как мономорфную популяцию крупной формы с широкими пределами варьирования размеров половозрелых особей.
Рис. 4.
Темп роста (средняя длина (FL) в возрастных группах) арктического гольца Salvelinus alpinus в озёрах бассейна Байкала: (—) – оз. Амут (Матвеев и др., 2020); (− ⋅ −) – оз. Светлинское, карликовая форма; (– –) − оз. Светлинское, мелкая форма (Самусенок и др., 2006); (- - -) – оз. Фролиха (Матвеев и др., 2020); особи из оз. Амут: (⚪) − с гонадами I, I−II и II стадий зрелости, (⚫) – с гонадами II−III, III, III−IV и VI−II стадий, (×) – FL ≈ 550 мм, рассчитанная по длине высушенной головы.
Соотношение самцов и самок в выборке равное. Впервые созревают в возрасте 6−7 лет. Половозрелые рыбы имели гонады II−III, III и III−IV стадий зрелости; индивидуальная абсолютная плодовитость варьировала в пределах 620−1055 (864) икринок, относительная – 1.4−2.4 (1.95) шт/г массы (5 самок FL 31−43.5 (36.4) см и массой 291−745 (463) г). Отмечены также пропускающие нерест самки с остаточными икринками (диаметром ~5 мм) в полости тела, сохранившимися после прошлогоднего нереста.
Гольцы из оз. Амут однородны по меристическим признакам (какие-либо внутрипопуляционные группировки не выявлены) и имеют ряд отличий от гольцов из двух других озёр бассейна Байкала (табл. 1). В частности, они имеют большее число жаберных тычинок (особенно в сравнении с гольцами из оз. Фролиха) и пилорических придатков. Более 95% гольцов из оз. Фролиха имеют в брюшном плавнике девять ветвистых лучей, а из оз. Светлинское − восемь, у гольцов из оз. Амут оба варианта встречаются с высокой частотой. При анализе по меристическим признакам в пространстве первых двух ГК гольцы из оз. Амут в большей степени перекрываются с гольцами из оз. Светлинское, чем из оз. Фролиха, скаттеры гольцов из последних двух озёр перекрываются незначительно, что обусловлено в первую очередь разным числом лучей в брюшном плавнике (рис. 5). По длине жаберных тычинок гольцы из оз. Амут входят в число наиболее длиннотычинковых гольцов Забайкалья. Она намного больше, чем у короткотычинковых гольцов из оз. Светлинское; гольцы же из оз. Фролиха, несмотря на малое число тычинок, по их длине занимают промежуточное положение (табл. 1, рис. 6).
Таблица 1.
Меристические признаки и относительная длина жаберных тычинок у гольцов Salvelinus alpinus из озёр бассейна Байкала
| Признак | Озеро, форма гольца | CD | rzh | Значимость различий по критерию Манна−Уитни |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Амут, крупная n = 321 |
2. Фролиха, крупная n = 382 |
Светлинское | |||||||||||
| 3 – карликовая n = 253 |
4 – мелкая n = 254 |
1–2 | 1–3 | 1–4 | 1–2 | 1–3 | 1–4 | 1–2 | 1–3 | 1–4 | |||
| FL, см | 21−48 | 15−42 | 13−18 | 27−38 | |||||||||
| Sb | $\frac{{32 - 41;\,\,1.72}}{{35.4 \pm 0.22}}$ | $\frac{{28 - 34;\,\,1.34}}{{30.6 \pm 0.20}}$ | $\frac{{31 - 39;\,\,1.84}}{{34.8 \pm 0.37}}$ | $\frac{{31 - 36;\,\,1.46}}{{33.7 \pm 0.29}}$ | 1.52 | 0.15 | 0.52 | 0.192 | 0.932 | 0.760 | *** | – | *** |
| rb1 | $\frac{{12 - 14;0.59}}{{12.7 \pm 0.10}}$ | $\frac{{11 - 14;0.72}}{{12.3 \pm 0.12}}$ | $\frac{{12 - 14;0.58}}{{12.8 \pm 0.11}}$ | $\frac{{12 - 13;0.44}}{{12.8 \pm 0.09}}$ | 0.33 | 0.10 | 0.07 | 0.908 | 0.995 | 0.954 | * | – | – |
| rb2 | $\frac{{11 - 13;0.61}}{{12.1 \pm 0.11}}$ | $\frac{{11 - 13;0.53}}{{11.7 \pm 0.09}}$ | $\frac{{11 - 13;0.57}}{{11.9 \pm 0.11}}$ | $\frac{{11 - 13;0.64}}{{12.4 \pm 0.13}}$ | 0.42 | 0.18 | 0.19 | 0.911 | 0.983 | 0.979 | ** | – | – |
| D | $\frac{{9 - 10;0.50}}{{9.4 \pm 0.09}}$ | $\frac{{9 - 11;0.61}}{{10.1 \pm 0.10}}$ | $\frac{{9 - 10;0.51}}{{9.6 \pm 0.10}}$ | $\frac{{9 - 10;0.50}}{{9.6 \pm 0.10}}$ | 0.55 | 0.12 | 0.16 | 0.824 | 0.992 | 0.987 | *** | – | – |
| A | $\frac{{7 - 9;0.54}}{{8.2 \pm 0.09}}$ | $\frac{{8 - 10;0.57}}{{9.1 \pm 0.09}}$ | $\frac{{8 - 9;0.43}}{{8.2 \pm 0.09}}$ | $\frac{{8 - 9;0.51}}{{8.6 \pm 0.10}}$ | 0.86 | 0.06 | 0.40 | 0.714 | 0.968 | 0.924 | *** | – | * |
| P | $\frac{{12 - 14;0.59}}{{13.1 \pm 0.10}}$ | $\frac{{12 - 13;0.51}}{{12.5 \pm 0.08}}$ | $\frac{{12 - 14;0.49}}{{12.9 \pm 0.10}}$ | $\frac{{12 - 14;0.54}}{{13.0 \pm 0.11}}$ | 0.58 | 0.18 | 0.13 | 0.831 | 0.980 | 0.991 | *** | – | – |
| V | $\frac{{8 - 9;0.49}}{{8.6 \pm 0.09}}$ | $\frac{{8 - 9;0.16}}{{9.0 \pm 0.03}}$ | $\frac{{7 - 9;0.29}}{{8.0 \pm 0.06}}$ | $\frac{{8 - 8;0.00}}{{8.0 \pm 0.00}}$ | 0.72 | 1.08 | 2.17 | 0.879 | 0.745 | 0.612 | ** | *** | *** |
| Ll | $\frac{{129 - 144;3.64}}{{134.2 \pm 0.66}}$ | $\frac{{129 - 138;2.02}}{{132.7 \pm 0.33}}$ | $\frac{{124 - 136;2.57}}{{129.0 \pm 0.51}}$ | $\frac{{126 - 135;2.31}}{{131.1 \pm 0.46}}$ | 0.33 | 1.02 | 0.64 | 0.892 | 0.431 | 0.647 | – | *** | ** |
| pc | $\frac{{37 - 59;5.31}}{{46.7 \pm 0.61}}$ | $\frac{{33 - 50;3.97}}{{40.9 \pm 0.40}}$ | $\frac{{31 - 47;3.70}}{{37.1 \pm 0.74}}$ | $\frac{{34 - 49;3.64}}{{41.0 \pm 0.72}}$ | 0.63 | 1.31 | 0.79 | 0.768 | 0.501 | 0.639 | *** | *** | *** |
| vert. | $\frac{{64 - 67;{\text{ }}0.87}}{{66.1 \pm 0.15}}$ | $\frac{{64 - 67;0.75}}{{65.5 \pm 0.13}}$ | $\frac{{65 - 67;0.61}}{{66.0 \pm 0.12}}$ | $\frac{{66 - 67;0.44}}{{66.2 \pm 0.09}}$ | 0.49 | 0.07 | 0.11 | 0.882 | 0.944 | 0.877 | ** | – | – |
| Среднее | 0.64 | 0.43 | 0.52 | 0.780 | 0.847 | 0.837 | |||||||
| lsb/c, % | $\frac{{9.5 - 13.5;1.12}}{{11.9 \pm 0.23}}$ | $\frac{{8.6 - 11.9;0.89}}{{10.1 \pm 0.27}}$ | $\frac{{6.9 - 9.7;0.72}}{{8.2 \pm 0.14}}$ | $\frac{{7.1 - 9.2;0.72}}{{8.1 \pm 0.24}}$ | 0.90 | 2.57 | 2.86 | *** | *** | *** | |||
Примечание. Обозначения признаков: sb − число жаберных тычинок на 1-й дуге; rb1, rb2 − число жаберных лучей слева и справа; D, A, P, V – число ветвистых лучей соответственно в спинном, анальном, грудном и брюшном плавниках; ll − число чешуй в боковой линии, vert. – число позвонков, pc − число пилорических придатков, lsb/c – длина наибольшей жаберной тычинки, в % длины головы. 1Для sb n = 62, для ll n = 30, для pc n = 75, для lsb/c n = 24 (FL 14−25 см); 2для sb n = 45, для pc n = 100, для vert. n = 33, для lsb/c n = 11 (FL 15−29 см); 3для pc n = 36; 4для pc n = 62, для lsb/c n = 9 (FL 27−32 см). Над чертой – пределы варьирования признака и среднее квадратическое отклонение, под чертой – среднее значение и его ошибка. CD – коэффициент различия Майра, rzh − показатель сходства Животовского (при его вычислении значения ll сгруппированы в классы по два, pc – по три). Различия по критерию Манна−Уитни достоверны при p: *< 0.05, **< 0.01, ***< 0.001; “−” − p > 0.05. Данные по карликовой и мелкой формам из оз. Светлинское – по: Самусeнок и др., 2006.
Рис. 5.
Распределение арктических гольцов Salvelinus alpinus из озёр бассейна Байкала в пространстве первых двух главных компонент (ГК) по 10
меристически признакам: (⚫), (—) – оз. Амут; (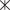 ), (– –) – оз. Фролиха; (∙ ∙ ∙) – оз. Светлинское ((△) − карликовая форма, (◇) − мелкая
форма).
), (– –) – оз. Фролиха; (∙ ∙ ∙) – оз. Светлинское ((△) − карликовая форма, (◇) − мелкая
форма).
Рис. 6.
Зависимость длины наибольшей жаберной тычинки (lsb) от длины головы (с) у арктических гольцов Salvelinus alpinus из озёр бассейна Байкала: (⚫) – оз. Амут; (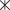 ) – оз. Фролиха; (△), (◇) – карликовая и мелкая форма из оз. Светлинское.
) – оз. Фролиха; (△), (◇) – карликовая и мелкая форма из оз. Светлинское.
Гольцы из бассейна Байкала не обособлены по морфологии от гольцов из забайкальской части бассейна Лены, отличающихся очень высокой межпопуляционной морфологической изменчивостью (Алексеев и др., 2000). Их меристические признаки укладываются в общие широкие пределы варьирования значений у последних, хотя при попарных сравнениях популяций в ряде случаев выявляются значительные различия. В бассейне Байкала пока не обнаружены столь многотычинковые гольцы (со средним числом жаберных тычинок >38), как в некоторых озёрах бассейнов верхнего Витима и Олёкмы. В этом байкальские гольцы сходны с гольцами из прилежащих бассейнов нижнего Витима и Чаи.
В питании преобладает рыба (гольяны и подкаменщики, случаи каннибализма не отмечены), обнаруженная в желудках 75% гольцов всех размеров (начиная с FL 14 см) и составлявшая 78% массы совокупного пищевого комка (табл. 2). В значительно меньшем количестве потребляются организмы макрозообентоса, наземные членистоногие и амфибиотические насекомые постларвальных стадий развития. Планктон не играет никакой роли даже в питании молоди, несмотря на относительно большое число жаберных тычинок и характерное для планктофагов строение ротового аппарата у некоторых молодых особей. Таким образом, в оз. Амут, как и в оз. Светлинское (Самусeнок и др., 2006), прослеживается несоответствие “трофической морфологии” и типа питания.
Таблица 2.
Питание арктического гольца Salvelinus alpinus из оз. Амут
| Компонент пищи и другие показатели | Доля массы пищевого комка, % | Частота встречаемости, % |
|---|---|---|
| Зоопланктон (Calanoida) | <0.1 | 1.6 |
| Макрозообентос: | 12.0 | 53.1 |
| – Chironomidae (личинки) | 0.6 | 31.3 |
| – Ephemeridae (личинки) | 2.7 | 25.0 |
| – Trichoptera (личинки) | <0.1 | 1.6 |
| – Euglesidae | 1.7 | 7.8 |
| – Hirudinea | 7.0 | 1.6 |
| Амфибиотические насекомые постларвальных стадий развития: | 2.8 | 59.4 |
| – Chironomidae (куколки) | 1.4 | 53.1 |
| – Trichoptera (имаго) | 1.4 | 12.5 |
| Наземные членистоногие: | 4.7 | 15.6 |
| – Hymenoptera (Formicidae) | 2.3 | 1.6 |
| – Aranea | 2.1 | 14.1 |
| – Heteroptera (Cicadellidae) | <0.1 | 1.6 |
| – Coleoptera: | ||
| – Cerambycidae | 0.3 | 1.6 |
| – Coccinellidae | <0.1 | 1.6 |
| Рыба: | 78.2 | 75.0 |
| – Cottus cf. poecilopus | 1.4 | 1.6 |
| – Phoxinus phoxinus | 36.0 | 18.8 |
| – переваренная | 40.8 | 79.7 |
| Прочие: | 2.3 | 34.4 |
| – фрагменты растительности | 1.5 | 34.4 |
| – Nostoc | 0.8 | 3.1 |
| Число рыб, экз. | 71 | |
| Доля пустых желудков, % | 7.8 | |
| Индекс наполнения желудков, ‱ | 0.1−169.5 (25.0) | |
ОБСУЖДЕНИЕ
Древнее позднеплиоцен–эоплейстоценовое соединение Байкала с бассейном Лены через долину Бугульдейки-Манзурки (Логачев и др., 1964; Нагорья …, 1974; Мац и др., 2001, 2010; Кононов, 2016) вряд ли можно рассматривать как путь распространения арктического гольца, поскольку на этом северо-западном направлении ни в бассейне Байкала, ни в прилежащей части бассейна верхней Лены этот вид не отмечен. К тому же сток в Лену прекратился более 600 тыс. лет назад (Кузьмин, 2001) − до наиболее вероятного времени проникновения гольца в бассейн Байкала. Не отмечен он и в бассейнах рек Баргузин и Ципа, через которые по одной из гипотез могло происходить более позднее соединение Байкала с бассейном Лены (Витима) (Ламакин, 1968; Кожов, 1972). Наиболее вероятным представляется его проникновение не через прямое соединение двух бассейнов, а через приводораздельные участки контакта в истоках ленских и байкальских притоков. Возможность такого расселения подтверждается геоморфологическими свидетельствами о вызванных комплексом экзогенных процессов плейстоценовых перехватах верховьев притоков Лены реками Байкальского бассейна (Базаров и др., 1981), а также находками характерного для притоков верхней Лены байкалоленского хариуса в северо-восточных притоках Байкала (Матвеев и др., 2005, 2006; Weiss et al., 2006; Пронин и др., 2007; Раднаев, 2007; Вокин, 2008; Книжин и др., 2008). Недавние находки пестроногого подкаменщика также сделаны в этих притоках: в Верхней Ангаре в районе устья р. Янчуй (в 18 км выше по течению от впадения в неё Чуро) и в низовьях р. Холодная (в бассейне соседнего с Верхней Ангарой крупного притока Северного Байкала – Кичеры) (Андреев и др., 2020). Все эти данные подтверждают гипотезу ингрессии ленских вод в бассейн Байкала (Ламакин, 1968; Кожов, 1972; Пронин и др., 2007).
Вопрос о времени проникновения арктического гольца в бассейн Байкала тесно связан с вопросом о длительности существования забайкальской популяционной группировки в бассейне Лены. По данным анализа мтДНК (Brunner et al., 2001; Alekseyev et al., 2009), гольцы Забайкалья, как и других континентальных районов Восточной Сибири, относятся к сибирской филогенетической группе (Brunner et al., 2001) или к сибирской подгруппе евроазиатской филогенетической группы (Гордеева и др., 2018) S. alpinus, возникновение которой с учётом данных разных генетических исследований (Brunner et al., 2001; Олейник и др., 2015; Lecaudey et al., 2018) можно датировать первой половиной неоплейстоцена. Поскольку Восточная Сибирь не покрывалась сплошным ледниковым покровом, как Северная Европа (Розенбаум, Шполянская, 2000), популяции арктического гольца могли существовать в ней длительное время, не ограниченное послеледниковьем. Согласно палеогеографическим данным, в плейстоцене на территории Забайкалья было четыре оледенения убывающей мощности: самаровское (мис-8) [(290 ± 60−260 ± 54)–(230 ± 54−200 ± 38) тыс. лет назад], тазовское (мис-6) (180 ± 40–130 ± 24 тыс. лет назад), муруктинское (мис-4) [(90−100)–(50−55) тыс. лет назад] и сартанское (мис-2) (25–10 тыс. лет назад) (Еникеев, 2009, 2011). Во время этих оледенений сток Лены в Северный Ледовитый океан блокировался сползавшим с Верхоянского хребта ледником (по мнению сторонников гипотезы глобального оледенения Евразии, смыкавшимся со сплошным панарктическим ледниковым щитом (Гросвальд, 1999)). В результате на территории Восточной Сибири возникало гигантское ледниково-подпрудное Лено-Вилюйское палеоозеро (озеро Лена, Якутское “море”), которое имело сток в бассейн Енисея через Вилюй-Нижнюю Тунгуску и в самаровское и тазовское оледенения достигало границ Байкало-Станового нагорья (Еникеев, 2009, 2011). Благодаря этому озеру арктический голец смог попасть в удалённые от моря внутренние области Восточной Сибири, включая Забайкалье, которых из-за ограниченной дальности его речных миграций не смог бы достичь, распространяясь с северных побережий по рекам. Можно полагать, что в условиях холодного климата ледниковых эпох Лено-Вилюйское озеро и водоёмы его бассейна были широко заселены арктическим гольцом и явились центром возникновения сибирской филогенетической подгруппы вида. В межледниковья озеро исчезало вместе с рыбным населением, но гольцы могли переживать потепления в горных районах, как переживают в них современное.
Поскольку во время самаровского оледенения всё Северное Забайкалье было покрыто сплошным ледяным щитом (Еникеев, 2009, 2011), арктические гольцы, даже если они обитали там ранее, должны были погибнуть. Во время же трёх следующих оледенений на его территории оставались свободные ото льда водоёмы. Соответственно, предки современных забайкальских гольцов могли колонизировать горные верховья бассейна Лены из Лено-Вилюйского озера либо во время таяния самаровских ледников около 200 тыс. лет назад, либо на 20−70 тыс. лет позже в тазовское оледенение и с тех пор обитать там постоянно: во времена оледенений − в низко расположенных долинных озёрах, в межледниковья − в освободившихся ото льда высокогорных. Более поздняя колонизация во время муруктинского и сартанского оледенений маловероятна в связи с отдалённостью в эти эпохи южных границ Лено-Вилюйского озера. Таким образом, и в бассейн Байкала арктические гольцы могли попасть не ранее 200−130 тыс. лет назад.
Описаны существовавшие в Северном Забайкалье во время тазовского, муруктинского и сартанского оледенений крупные ледниково-подпрудные озёра, пригодные для обитания гольцов: огромное Витимское (Муйско-Витимское), занимавшее бóльшую часть бассейна верхнего Витима выше оронской депрессии (Krivonogov, Takahara, 2003; Еникеев, 2009; Margold et al., 2011, 2016, 2018) c площадью поверхности до 23.5 тыс. км2 и объёмом до ~3 тыс. км3 (Margold et al., 2011), меньшее Чарское (Музис, 1968; Еникеев, 2009, 2018; Margold, Jansson, 2011), а в тазовское и муруктинское время − также Олёкминское (Еникеев, 2009, 2011). Витимское и Олёкминское озёра имели сток в бассейн Амура, но неоднократно прорывались через ледниковые подпруды в бассейн Лены; последний прорыв Витимского озера во время таяния сартанских ледников (~14 тыс. лет назад) привёл к одному из самых крупных катастрофических сбросов пресных вод в мире (Margold et al., 2011, 2018). В бассейне Северного Байкала, покрывавшегося Баргузинским ледником, подобных палеоозёр, вероятно, не было, однако во время плейстоценовых ледниковых максимумов сам Байкал мог служить убежищем для гольцов. Существует гипотеза (Леви и др., 2015), что в начале казанцевского потепления в результате таяния ледников предшествовавшего ему тазовского оледенения верхнеангарская депрессия заполнялась водой и превращалась в залив Байкала (Восточносибирского озера), который вплотную приближался к Байкало-Ленскому водоразделу в районе долины Правой Мамы.
Находка арктического гольца и пестроногого подкаменщика в оз. Амут на водоразделе с бассейном Правой Мамы указывает на место, где можно предполагать их проникновение в бассейн Байкала из слабо врезанной долины этой реки даже без использования классического сценария перехвата верховий. Здесь на низких слабовыраженных водораздельных участках, изобилующих мелкими ручьями, старыми руслами, небольшими подболоченными озерцами, даже в настоящее время в многоводные годы могут создаться условия, способствующие миграции рыб между озером и Правой Мамой по сети временных водотоков. Ещё более вероятно, что такие условия возникали в прошлом при заполнении низких приводораздельных участков талыми ледниковыми водами. При высоком уровне вод озера мог даже возникать эпизодический сток из Амута в Правую Маму. Вероятно, спустившись по Чуро, вселенцы через основное русло Верхней Ангары распространялись по её бассейну: гольцы достигли оз. Светлинское и через Байкал − оз. Фролиха, а подкаменщики могли попасть в бассейн Кичеры через проточные соединения её низовий с низовьями Верхней Ангары как раз в районе устья Холодной.
Арктический голец в соседствующих с бассейном Байкала верховьях Лены распространён мозаично, этот вид населяет только горные озёра и имеет относительно крупные размеры, затрудняющие активную миграцию через мелкие водотоки. По этим причинам трудно ожидать, что он проникал в бассейн Байкала многократно в разных местах. Путь его распространения через долину Правой Мамы и оз. Амут на сегодняшний день представляется наиболее обоснованным геологическими, зоогеографическими и генетическими данными. В бассейне Мамы арктический голец отмечен в четырёх озёрах: в оз. Амудиса и двух озёрах в верховьях ручья Озёрный, относящихся к бассейну р. Большая Конкудера, а также в оз. Соли в бассейне Левой Мамы (Alekseyev et al., 2009), хотя в малообследованном бассейне Правой Мамы пока не найден.
Во всех популяциях арктического гольца из бассейна Байкала обнаружен широко распространённый предковый для сибирской подгруппы гаплотип контрольной области мтДНК SIB10, он же является единственным гаплотипом во всех популяциях бассейнов Мамы и Чаи. В озёрах Амут и Светлинское от него произошли по одному (отличающемуся на одну мутацию) уникальному гаплотипу – SIB32 и SIB11 (Alekseyev et al., 2009; Гордеева и др., 2018). Это свидетельствует о близком родстве байкальских популяций с верхнеленскими и конкретно с их вероятными предками, гольцами бассейна нижнего Витима (Мамы). Следует отметить, что в отличие от гольцов бассейнов как нижнего Витима, так и Байкала, гольцы бассейна верхнего Витима характеризуются уникальным генетическим (10 гаплотипов мтДНК) и высоким морфологическим (в частности, наличие крайне многотычинковых форм) разнообразием. Становление этого разнообразия, видимо, связано с их первоначальной диверсификацией в Витимском палеоозере с последующей эволюцией в изолированных популяциях высокогорных озёр.
Пестроногий подкаменщик как вид, гораздо шире распространённый в верховьях бассейна Лены, населяющий не только озёра, но и реки, встречающийся от равнинных водоёмов до самых истоков горных ручьёв и отличающийся малыми размерами, очевидно, имел больше возможностей для проникновения в бассейн Байкала, и путь распространения через Амут для него является не единственным. В частности, вполне вероятно его проникновение в р. Холодная из верховий р. Асектамур, относящейся к бассейну Чаи (система Лены).
Полученные данные расширяют представления о зоогеографии бассейна Байкала, об истории и путях проникновения в него аллохтонных видов рыб, а также о разнообразии арктических гольцов этого бассейна, которое до сих пор слабо исследовано и нуждается в дальнейшем изучении.
Список литературы
Алексеев С.С., Булдыгеров В.В., Пичугин М.Ю., Самусeнок В.П. 1999. Распространение арктического гольца Salvelinus alpinus (Salmonidae) в Забайкалье // Вопр. ихтиологии. Т. 39. № 1. С. 48−56.
Алексеев С.С., Пичугин М.Ю., Самусeнок В.П. 2000. Разнообразие гольцов Забайкалья по меристическим признакам, их положение в комплексе Salvelinus alpinus и проблема происхождения симпатрических форм // Там же. Т. 40. № 3. С. 293−311.
Андреев Р.С., Матвеев А.Н., Самусeнок В.П. и др. 2020. Первые находки пестроногого подкаменщика Cottus cf. poecilopus Heckel, 1837 в бассейне Байкала // Изв. ИркутГУ. Сер. биол. экология. Т. 31. С. 30−51.
Аров И.В., Мишарина Е.А. 2018. Современное состояние изученности и пути генезиса фауны коловраток (Rotifera) озера Байкал // Там же. Т. 25. С. 70−90. https://doi.org/10.26516/2073-3372.2018.25.70
Базаров Д.Б., Будаев Р.Ц., Дергаусова М.И. и др. 1981. Геоморфология северного Прибайкалья и Станового нагорья. М.: Наука, 202 с.
Вокин А.И. 2008. Экология хариусовых рыб (Thymallidae) горных водоемов Байкальской рифтовой зоны: Автореф. дис. … канд. биол. наук. Иркутск: ИГУ, 23 с.
Гордеева Н.В., Алексеев С.С., Кириллов А.Ф. и др. 2018. Распространение, состав и родственные отношения филогенетических групп арктического гольца Salvelinus alpinus (Salmoniformes, Salmonidae) в европейской части России и Сибири по данным анализа нуклеотидных последовательностей митохондриальной ДНК // Вопр. ихтиологии. Т. 58. № 6. С. 659−669. https://doi.org/10.1134/S0042875218050107
Гросвальд М.Г. 1999. Евразийские гидросферные катастрофы и оледенение Арктики. М.: Науч. мир, 120 с.
Еникеев Ф.И. 2009. Плейстоценовые оледенения восточного Забайкалья и юго-востока Средней Сибири // Геоморфология. Т. 2. С. 33−49. https://doi.org/.https://doi.org/10.15356/0435-4281-2009-2-33-49
Еникеев Ф.И. 2011. Россыпеобразование в условиях гляциального морфогенеза восточного Забайкалья: Автореф. дис. … докт. геол.-минерал. наук. Чита: ЗабайкалГУ, 41 с.
Еникеев Ф.И. 2018. Палеогеография сартанского оледенения горного обрамления Муйской впадины (северное Забайкалье) // Геосферные исследования. № 3. С. 58–70
Животовский Л.А. 1979. Показатель сходства популяций по полиморфным признакам // Журн. общ. биологии. Т. 40. № 4. С. 587–602.
Книжин И.Б., Вайс С. Дж., Богданов Б.Э., Копун Т. 2008. Новые данные о распространении верхнеленской формы хариуса (Thymallidae) в бассейне озера Байкал и ее таксономическом статусе // Вопр. ихтиологии. Т. 48. № 2. С. 166−172.
Кожов М.М. 1972. Очерки по байкаловедению. Иркутск: Изд-во ИГУ, 252 с.
Кононов Е.Е. 2016. О новых данных по проблеме Пра-Манзурского канала стока байкальских вод // Изв. СО РАЕН. Геология, поиски и разведка рудных месторождений. № 2 (55). С. 116–128.https://doi.org/10.21285/0301-108X-2016-55-2-116-128
Кузьмин М.И. 2001. Во льдах Байкала. Новосибирск: Гео, 140 с.
Ламакин В.В. 1968. Неотектоника Байкальской впадины. М.: Наука, 245 с.
Леви К.Г., Мирошниченко А.И., Козырева Е.А., Кадетова А.В. 2015. Модели эволюции озерных бассейнов Восточной Сибири в позднем плейстоцене и голоцене // Изв. ИркутГУ. Сер. геоархеология, этнология, антропология. Т. 11. С. 55−85.
Логачев Н.А., Ломоносова Т.К., Климанова В.М. 1964. Кайнозойские отложения Иркутского амфитеатра. М.: Наука, 195 с.
Матвеев А.Н., Самусeнок В.П. 2008. Происхождение (источники и пути проникновения) ихтиофауны озера Байкал // Матер. науч. конф. “Развитие жизни в процессе абиотических изменений на Земле”. Новосибирск: Изд-во СО РАН. С. 401−414.
Матвеев А.Н., Самусенок В.П., Тельпуховский А.Н. и др. 2005. Новый подвид сибирского хариуса Thymallus arcticus baiсalolenensis ssp. nova (Salmoniformes, Thymallidae) // Вестн. БурятГУ. Сер. биолог. Вып. 7. С. 69−82.
Матвеев А.Н., Самусенок В.П., Пронин Н.М. и др. 2006. Биология нового подвида сибирского хариуса Thymallus arcticus baiсalolenensis ssp. nova (Salmoniformes, Thymallidae) в бассейне озера Байкал // Там же. Вып. 8. С. 222−233.
Матвеев А.Н., Самусeнок В.П., Юрьев А.Л. и др. 2020. Биология рыб озера Фролиха (Северный Байкал, Восточная Сибирь) // Изв. ИркутГУ. Сер. биол. экология. Т. 32. С. 50−82.
Мац В.Д., Уфимцев Г.Ф., Мандельбаум М.М. и др. 2001. Кайнозой Байкальской рифтовой впадины: строение и геологическая история. Новосибирск: Изд-во СО РАН, 252 с.
Мац В.Д., Ефимова И.М., Кульчицкий А.А. 2010. Древние долины западного Прибайкалья (история формирования) // Геоморфология. Т. 2. С. 91−101. https://doi.org/10.15356/0435-4281-2010-2-91-101
Музис А.И. 1968. Среднечетвертичный озерный водоем Чарской впадины // Мезозойские и кайнозойские озера Сибири. М.: Наука. С. 125−138.
Мухомедияров Ф.Б. 1942. К биолого-систематической характеристике даватчана // Тр. Вост.-Сиб. ун-та. Т. 2. Вып. 3. С. 119−126.
Нагорья Прибайкалья и Забайкалья. 1974 / Под ред. Флоренсова Н.А. М.: Наука, 359 с.
Олейник А.Г., Скурихина Л.А., Брыков В.А. 2015. Филогения гольцов рода Salvelinus по данным анализа митохондриальной ДНК // Генетика. Т. 51. № 1. С. 63−77. https://doi.org/10.7868/S0016675815010099
Петров Е.А. 2003. Байкальская нерпа (эколого-эволюционные аспекты): Автореф. дис. … докт. биол. наук. Улан-Удэ: Востсибрыбцентр, 38 с.
Правдин И.Ф. 1966. Руководство по изучению рыб. М.: Пищ. пром-сть, 324 с.
Пронин Н.М., Матвеев А.Н., Самусенок В.П. и др. 2007. Рыбы озера Байкал и его бассейна. Улан-Удэ: Изд-во БурятНЦ СО РАН, 284 с.
Пудовкина Т.А., Ситникова Т.Я., Матвеев А.Н., Щербаков Д.Ю. 2014. Родственные связи байкальских полихет рода Manayunkia [Polychaeta: Sedentaria: Sabellidae] по данным анализа COI с анализом истории расселения // Экол. генетика. Т. XII. № 3. С. 32−42.
Раднаев Н.Д. 2007. Морфоэкологическая характеристика хариусов Thymallus из бассейна реки Баргузин // Вестн. БурятГУ. Сер. химия, биология, география. Вып. 3. С. 238−244.
Редкозубов И.Ю., Мовчан В.А. 1974. К изучению даватчана Salvelinus alpinus erythrinus (Georgi) оз. Фролиха // Вопр. ихтиологии. Т. 14. Вып. 2. С. 330−332.
Розенбаум Г.Э., Шполянская Н.А. 2000. Позднекайнозойская история криолитозоны Арктики и тенденции ее будущего развития. М.: Науч. мир, 100 с.
Савваитова К.А., Максимов В.А., Медведева Е.Д. 1977. Даватчан Salvelinus alpinus erythrinus (Georgi) // Вопр. ихтиологии. Т. 17. Вып. 2. С. 203−219.
Самусeнок В.П., Алексеев С.С., Матвеев А.Н. и др. 2006. Вторая в бассейне Байкала и самая высокогорная в России популяция арктического гольца Salvelinus alpinus complex (Salmoniformes, Salmonidae) // Там же. Т. 46. № 5. С. 616−629.
Ситникова Т.Я., Прозорова Л.А. 2008. Еще раз о происхождении байкальских эндемичных моллюсков семейства Baicaliidae Clessin 1880 (Gastropoda) // Матер. науч. конф. “Развитие жизни в процессе абиотических изменений на Земле”. Новосибирск: Изд-во СО РАН. С. 371–375.
Alekseyev S.S., Samusenok V.P., Matveev A.N., Pichugin M.Yu. 2002. Diversification, sympatric speciation, and trophic polymorphism of Arctic charr (Salvelinus alpinus complex) in Transbaikalia // Environ. Biol. Fish. V. 64. № 1−3. P. 97−114. https://doi.org/10.1007/978-94-017-1352-8_8
Alekseyev S.S., Bajno R., Gordeeva N.V. et al. 2009. Phylogeography and sympatric differentiation of the Arctic charr Salvelinus alpinus (L.) complex in Siberia as revealed by mtDNA sequence analysis // J. Fish Biol. V. 75. P. 368–392. https://doi.org/10.1111/j.1095?8649.2009.02331.x
Berg L.S. 1907. Die Cataphracti des Baikal-Sees. Wissentschaftliche Ergebnisse einer zoologischen Expedition nach dem Baikal-See. St. Petersburg; Berlin: S. n., II+75 s.
Brunner P.C., Douglas M.R., Osinov A. et al. 2001. Holarctic phylogeography of Arctic charr (Salvelinus alpinus L.) inferred from mitochondrial DNA sequences // Evolution. V. 55. P. 573−586. https://doi.org/10.1554/0014-3820(2001)055[0573:hpoacs]-2.0.co;2
Georgi J.G. 1775. Bemerkungen einer Reise im Russischen Reich im Jahre 1772. Bd. I. St. Petersburg: Gedruckt bei der Keiserl Academie der Wissenschaften, 506 s.
Krivonogov S.K., Takahara H. 2003. Late Pleistocene and Holocene environmental changes recorded in the terrestrial sediments and landforms of Eastern Siberia and North Mongolia // Proc. Int. Symp. “21-st Century COE Program”. V. 1 / Ed. Kamata N. Kanazawa: Kanazawa Univ. P. 30–36.
Lecaudey L.A., Schliewen U.K., Osinov A.G. et al. 2018. Inferring phylogenetic structure, hybridization and divergence times within Salmoninae (Teleostei: Salmonidae) using RAD-sequencing // Mol. Phylogen. Evol. V. 124. P. 82–99. https://doi.org/10.1016/j.ympev.2018.02.022
Margold M., Jansson K.N. 2011. Glacial geomorphology and glacial lakes of central Transbaikalia, Siberia, Russia // J. Maps. V. 7. P. 18−30. https://doi.org/10.4113/jom.2011.1132
Margold M., Jansson K.N., Stroeven A.P., Jansen J.D. 2011. Glacial Lake Vitim, a 3000-km3 outburst flood from Siberia to the Arctic Ocean // Quat. Res. V. 76. P. 393−396. https://doi.org/10.1016/j.yqres.2011.06.009
Margold M., Jansen J.D., Гуринов А.Л. и др. 2016. Плейстоценовые ледниково-подпрудные озера впадин северного Забайкалья и последствия их катастрофического спуска // Матер. Всерос. науч. конф. “Пути эволюционной географии”. Москва. С. 500−503.
Margold M., Jansen J.D., Codilean A.T. et al. 2018. Repeated megafloods from glacial Lake Vitim, Siberia, to the Arctic Ocean over the past 60,000 years // Quat. Sci. Rev. V. 187. P. 41−46. https://doi.org/10.1016/j.quascirev.2018.03.005
Mayr E. 1963. Animal species and evolution. Cambridge, USA: Harward Univ. Press, 797 p.
Weiss S., Knizhin I., Kirillov A., Froufe E. 2006. Phenotypic and genetic differentiation of two major phylogeographic lineages of arctic grayling Thymallus arcticus in the Lena River, and surrounding Arctic draining basins // Biol. J. Linn. Soc. V. 88. P. 511−525. https://doi.org/10.1111/j.1095-8312.2006.00621.x
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии