

Вопросы ихтиологии, 2021, T. 61, № 4, стр. 423-431
Особенности биологии налима Lota lota (Gadidae) нижнего течения Волги
В. С. Болдырев *
Волгоградский филиал Всероссийского научно-исследовательского института
рыбного хозяйства и океанографии – ВолгоградНИРО
Волгоград, Россия
* E-mail: neogobius@yahoo.com
Поступила в редакцию 02.06.2020
После доработки 11.08.2020
Принята к публикации 13.08.2020
Аннотация
Приведены данные по биологии краевой популяции налима Lota lota в нижнем течении Волги. В уловах отмечены особи в возрасте от 3 до 7 лет; сравнительно короткий возрастной ряд обусловлен гибелью налима в жаркие годы. Нерест проходит с середины января до середины февраля; скат личинок продолжается по апрель–начало мая. В питании доминируют черноморско-каспийская тюлька Clupeonella cultriventris и бычки (Gobiidae); среди последних наибольшее значение имеют бычок-кругляк Neogobius melanostomus, каспийский головач Ponticola gorlap и песочник N. fluviatilis. В настоящее время самым высоким темпом роста в водоёмах Северной и Восточной Европы характеризуются южные популяции налима Саратовского водохранилища и нижнего течения Волги. Это обусловлено, прежде всего, высокой численностью и доступностью кормовых объектов, основу которых составляют виды рыб, расширившие своё присутствие в Волжском бассейне в условиях зарегулированного стока.
Налим Lota lota широко распространён в пресных водах северных районов Европы, Азии и Америки. В отдельных водоёмах является массовым объектом промысла (Богданов, Копориков, 2011). В Волге этот холодолюбивый вид обычен в верхнем и среднем течении (Гримм, 1896; Исаев, Карпова, 1989; Герасимов и др., 2018б). По нижнему незарегулированному участку реки, где проходит южная граница его современного ареала в бассейне Каспийского моря (Казанчеев, 1981; Богуцкая и др., 2013), сведения о налиме имеются со второй половины XVIII в. (Медведев, 2013), но ограничиваются главным образом информацией о его редкой встречаемости (Коблицкая, 1964; Гольдентрахт, 1966; Чехова, Пономарёва, 1973; Вехов, Горский, 2010; Красная книга …, 2013; Литвинов, Подоляко, 2013).
Цель работы – характеристика размерно-возрастной структуры, размножения, питания, темпа роста и динамики численности налима нижнего течения Волги, а также анализ изменчивости роста этого вида в водоёмах Северной и Восточной Европы.
МАТЕРИАЛ И МЕТОДИКА
Материал по сетным уловам налима в нижнем бьефе Волжской ГЭС (ВГЭС) собирали в 2011−2013 и 2015−2018 гг. в рамках регулярных исследований Волгоградского отделения ГосНИОРХ (с 2019 г. Волгоградский филиал ВНИРО). Использованы также данные Волгоградского осетрового рыбоводного завода по отлову частика, органов рыбоохраны по изъятию браконьерских орудий лова на этом участке и опросные сведения рыбаков-любителей. Применяли лесковые сети с шагом ячеи 35−55 мм длиной 75 м и высотой 2.5 м; продолжительность постановок составляла 12−48 ч. Сети выставляли на участке с глубинами 15−22 м в 500-метровой зоне ниже водосливной плотины. При закрытом её положении в левобережной части акватории нижнего бьефа за пределами основного потока от агрегатов образуется зона локальных циркуляционных течений, их скорость обычно варьирует в пределах 0.1−0.3 м/с. В период, когда водосливная плотина открывалась, что происходит чаще со второй половины апреля по начало июня, сети убирали. За период наблюдений проанализированы уловы 1284 сетей и учтено 945 особей налима. Средние показатели уловов для характеристики их сезонной динамики рассчитывали только для месяцев, в которых число сетепостановок было не менее пяти.
У налима измеряли абсолютную длину (TL). Для сопоставления с литературными данными, в которых приводится стандартная длина (SL), использовали соотношение SL = 0.94ТL (Маркун, 1936; Махотин, 1960). Возраст рыб определяли по отолитам.
Сбор материала по питанию налима и обработку содержимого желудков проводили в декабре 2010–марте 2011 г. и октябре–декабре 2011 г. стандартными методами (Методическое пособие …, 1974). Целые организмы в пищевом комке идентифицировали, просчитывали и измеряли; непищевые объекты учитывали отдельно. Для оценки значения пищевых объектов рассчитывали долю массы компонента (% общей массы пищевого комка) и частоту встречаемости − долю желудков с данным компонентом (% общего число желудков с пищей). Всего исследовано питание 133 экз. налима, желудки девяти из которых были пустыми.
Ихтиопланктон собирали с конца апреля по июнь в 2008−2017 гг. на русловом участке реки в 65 км ниже гидроузла в районе пос. Светлый Яр с помощью икорных сетей, выставляемых в дневное время в трёх горизонтах (поверхность, толща, дно).
Материал по качественно-количественному составу бычков (Gobiidae), основных компонентов питания налима, собирали на 60-километровом участке Волги ниже ВГЭС в летне-осенние периоды 2010−2012 и 2016−2019 гг. мальковыми волокушами, изготовленными из безузловой дели (длина 6−30 м, ячея 4 мм). Облавливали прибрежные слабопроточные участки с глубинами до 2.5 м. Всего учтено 4897 экз. четырёх видов бычков. Названия рыб даны в соответствии с номенклатурными и таксономическими изменениями, принятыми в современных сводках (Kottelat, Freyhof, 2007; Froese, Pauly, 2019).
РЕЗУЛЬТАТЫ
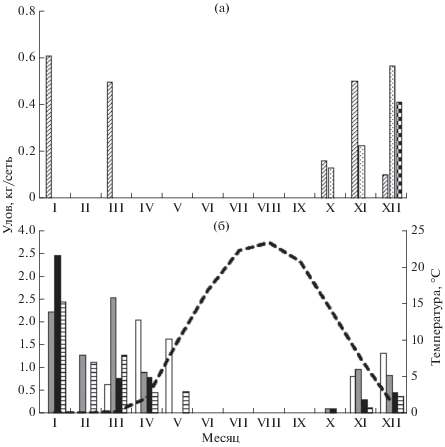
Динамика уловов. По данным многолетних наблюдений, сетные уловы налима на предплотинном участке очень неравномерны по сезонам (рис. 1). Летом и в начале осени в силу низкой активности он, если и встречается, то единично. Нагуливаться, судя по регулярным поимкам, начинает с октября, когда температура воды снижается до 11−13°С (рис. 1б). Максимальные уловы приходятся на зиму и весну. Уловы в 2011−2013 гг. (рис. 1а) были значительно ниже, чем в 2015−2018 гг. (рис. 1б).
Рис. 1.
Сезонная динамика уловов налима Lota lota и температуры воды в нижнем бьефе Волжской ГЭС: а − 2011−2013 гг., б − 2015−2018
гг.; (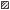 ) – 2011 г., (
) – 2011 г., (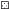 ) − 2012 г., (
) − 2012 г., (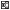 ) − 2013 г., (
) − 2013 г., ( ) − 2015 г., (
) − 2015 г., (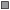 ) − 2016 г., (◼) − 2017 г., (
) − 2016 г., (◼) − 2017 г., ( ) − 2018 г., (− −) – температура воды.
) − 2018 г., (− −) – температура воды.
Размерно-возрастной состав. В уловах присутствовали особи пяти возрастных групп TL 380−700 мм и массой 500−2440 г. (табл. 1). Основу (84%) составляли трёх- и четырёхлетки; соотношение полов среди них примерно равное. В возрастной группе 4+ самцов было уже заметно больше (75%). Единичные в уловах особи в возрасте 5+…6+ представлены только самцами.
Таблица 1.
Длина (TL) и масса налима Lota lota разного возраста из нижнего бьефа Волжской ГЭС
| Возраст, лет | Число рыб, экз. | TL, мм | Масса, г | ||||
|---|---|---|---|---|---|---|---|
| Оба пола | Самцы | Самки | мin–max | M | min−max | M | |
| 2+−3 | 42 | 20 | 22 | 380−540 | 439.3 | 395−650 | 555.6 |
| 3+−4 | 65 | 34 | 31 | 400−650 | 484.2 | 490−1350 | 707.1 |
| 4+−5 | 17 | 13 | 4 | 460−640 | 536.2 | 610−1700 | 939.8 |
| 5+−6 | 2 | 2 | − | 605−617 | 611 | 1340−1600 | 1470 |
| 6+−7 | 1 | 1 | − | 700 | 2440 | ||
Размножение. В некоторых водоёмах Волжского бассейна налим начинает созревать уже в возрасте 1+ (Рыбы Рыбинского водохранилища …, 2015). В самой младшей возрастной группе в нашей выборке (2+) половозрелыми были 75% особей; среди рыб в возрасте 3+ их доля составляла уже 95%, в возрасте 4+…6+ − 100%.
Нерест налима на предплотинном участке проходит в январе−феврале и продолжается ~1 мес. В 2010 г. первые отнерестившиеся самки были отмечены в уловах 20 января, последние – 15 февраля, в 2017 г. − соответственно 17 января и 10 февраля.
В ходе наблюдений за скатом молоди рыб предличинки налима TL 4.0−4.5 мм были отмечены в уловах икорных сетей во всех облавливаемых горизонтах только дважды – 07.05.2008 и 13.05.2010 г. Температура воды к этому времени составляла ~10°С. В другие годы, вероятно, скат заканчивался раньше. Единичная поимка предличинки налима в низовьях дельты Волги была отмечена Коблицкой (1964) в 1957 г. также в начале мая при температуре воды 12°С. Из этого можно сделать вывод о том, что продолжительность эмбрионального периода развития налима в нижнем течении реки составляет ~2−3 мес.
Питание. Основу питания взрослого налима в нижнем бьефе ВГЭС составляют рыбы. В пищевых комках отмечено десять видов (табл. 2). Доминируют тюлька Clupeonella cultriventris и бычки, среди последних наибольшее значение имеет бычок-кругляк Neogobius melanostomus. Доля головача Ponticola gorlap и песочника N. fluviatilis несколько ниже. Составляющая цуцика Proterorhinus nasalis, гонца Babka gymnotrachelus и пуголовки Benthophilus durrelli незначительна. Максимальное число тюлек в пищевом комке налима составило 63 экз., бычков – 34 экз.
Таблица 2.
Состав пищи налима Lota lota из нижнего бьефа Волжской ГЭС
| Компонент пищи | Доля, % массы пищи | Частота встречаемости, % |
|---|---|---|
| Clupeonella cultriventris – черноморско-каспийская тюлька | 19.9 | 32.3 |
| Alburnus alburnus – уклейка | 0.2 | 0.8 |
| Perca fluviatilis – речной окунь | 3.7 | 1.6 |
| Sander lucioperca – обыкновенный судак | 0.1 | 0.8 |
| Neogobius melanostomus – бычок-кругляк | 16.1 | 30.6 |
| N. fluviatilis – бычок-песочник | 6.7 | 11.3 |
| Ponticola gorlap − каспийский бычок-головач | 10.2 | 9.7 |
| Babka gymnotrachelus − азово-черноморский бычок-гонец | 1.8 | 9.7 |
| Proterorhinus nasalis – бычок-цуцик | 0.5 | 4.0 |
| Benthophilus durrelli − донская пуголовка | 0.5 | 5.6 |
| Неопределённые до вида Gobiidae | 28.3 | 76.6 |
| Бокоплавы (Amphipoda) | 6.1 | 62.1 |
| Раки (Astacidae) | 1.7 | 11.3 |
| Насекомые (Insecta) | 0.1 | 2.4 |
| Створки дрейссены Dreissena sp. | 2.0 | 46.0 |
| Раковины брюхоногих моллюсков (Gastropoda) | 0.1 | 9.7 |
| Фрагменты растений (Embryophyta) | 0.8 | 14.5 |
| Песок, камни | 1.2 | 27.4 |
В рацион налима входят также часто отмечаемые бокоплавы (Amphipoda), реже − раки (Astacidae), личинки водных насекомых (стрекозы (Odonata) и ручейники (Hydropsychidae)). Раки в желудках представлены целыми особями (TL 31−46 мм) или их отдельными клешнями. Из непищевых объектов, видимо, случайно захватываемых при охоте, довольно часто отмечаются створки дрейссены Dreissena sp., раковины мелких брюхоногих моллюсков (Theodoxus, Lithoglyphus и Hippeutis), высшие растения (фрагменты рдеста и валлиснерии, кусочки древесины, веток и листьев деревьев), песок и галька. Из 34 окатанных камней, извлечённых из желудков, большая часть имела массу 0.02−0.30 г (77% общего числа), меньшая – 0.5−10.7 г. Самые крупные плоские гальки имели размер 25 × 20 × 5 мм, шаровидные – до 15−20 мм в диаметре. Масса пищевого комка варьировала в пределах 0.4−11.0% массы тела рыбы, в среднем 3.4%.
Обращает на себя внимание диспропорция в соотношении отдельных видов бычков в питании налима и в уловах мальковой волокуши. Так, доля песочника, самого массового (95%) представителя бычковых на прибрежных мелководьях Волги (кругляк 4%, цуцик 1%, головач < 1%)11, в питании налима составляла всего 10% общего числа съеденных бычков. В зимний период все эти виды откочёвывают с мелководий в глубоководные участки (Богуцкая и др., 2004), т.е. можно было ожидать их близкого соотношения на нагульных биотопах и в пищевом комке налима. Такое сильное расхождение объясняется, скорее, не избирательностью хищника, а разницей в биотопическом распределении песочника и кругляка (64%), являющегося основным объектом его питания. Первый широко заселяет открытые мелководья с песчаным грунтом, преобладающим типом русловых отложений в нижнем течении Волги, и поэтому доминирует в уловах волокуши. Кругляк же по сравнению с другими видами бычков в большей степени осваивает участки с каменистым субстратом, что во многом обусловлено наличием здесь его основного объекта питания − дрейссены. Нижний бьеф плотины, где отлавливали налима, как раз характеризуется наличием берегоукрепительных бетонных конструкций и большими площадями щебёночной отсыпки, что определяет высокую концентрацию кругляка на этом участке и его бóльшую долю в питании хищника. Вклад остальных четырёх видов бычков (головач, цуцик, гонец и пуголовка) в питание налима незначителен и варьирует в пределах 5−10% общего числа съеденных бычков (табл. 3). Среди бычков в питании налима преобладают сеголетки, что, видимо, определяется их доминированием в русловой части реки в местах нагула хищника. Наибольшую роль играют особи, длина которых составляет 4−11% TL налима; доля крупных жертв (SL > 60 мм) составляет всего ~10% общего числа съеденных рыб.
Таблица 3.
Размерный состав шести видов бычков в питании налима Lota lota
| Размерная группа (SL), мм | Число жертв, экз. | Всего | |||||
|---|---|---|---|---|---|---|---|
| кругляк | песочник | головач | гонец | цуцик | пуголовка | ||
| 10.0−15.0 | 1 | 1 | |||||
| 15.1−20.0 | 17 | 1 | 1 | 19 | |||
| 20.1−25.0 | 20 | 2 | 2 | 24 | |||
| 25.1−30.0 | 16 | 1 | 2 | 19 | |||
| 30.1−35.0 | 14 | 2 | 1 | 1 | 1 | 1 | 20 |
| 35.1−40.0 | 9 | 1 | 2 | 5 | 2 | 3 | 22 |
| 40.1−45.0 | 10 | 3 | 2 | 7 | 1 | 23 | |
| 45.1−50.0 | 9 | 2 | 1 | 2 | 1 | 2 | 17 |
| 50.1−55.0 | 9 | 2 | 1 | 1 | 1 | 14 | |
| 55.1−60.0 | 4 | 2 | 6 | ||||
| 60.1−65.0 | 3 | 1 | 1 | 5 | |||
| 65.1−70.0 | 2 | 2 | |||||
| 70.1−75.0 | 1 | 1 | 1 | 3 | |||
| 80.1−85.0 | 1 | 1 | 2 | ||||
| 95.1−100.0 | 1 | 1 | |||||
| 100.1−105.0 | 1 | 1 | 2 | ||||
| 110.1−115.0 | 1 | 1 | |||||
| 120.1−125.0 | 1 | 1 | 2 | 4 | |||
| Всего | 118 | 18 | 13 | 18 | 9 | 9 | 185 |
ОБСУЖДЕНИЕ
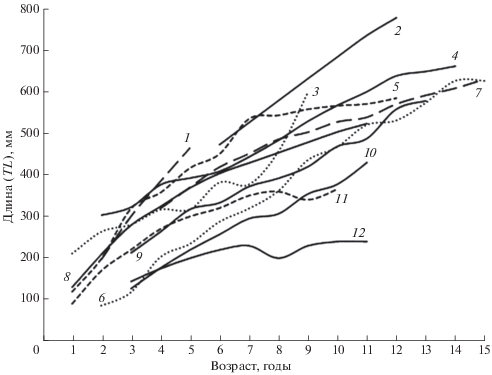
Представляет интерес рассмотрение географической изменчивости роста налима как одного из широкоареальных видов. Характер роста рыб в значительной степени обусловлен межорганизменными отношениями и влиянием среды обитания. Основными факторами, определяющими скорость роста, являются термический режим и обеспеченность пищей. Немаловажное значение могут иметь солёность, кислородный режим, плотность популяции самого вида и его конкурентов, размеры водоёма и количество пищевых ниш в нём для группировок разного размера и другие условия (Решетников и др., 1989, 2016; Дгебуадзе, 2001; Шатуновский и др., 2009). Особенностью биологии налима является то, что тёплое время года он проводит в состоянии оцепенения, а активизация нагула происходит при температуре воды ниже 11−13°С (Kottelat, Freyhof, 2007). Оптимумом роста у этого холодолюбивого вида является температурный диапазон 8−16°С (Голованов, 2014). Продолжительность нагула налима на юге ареала значительно меньше, чем в северных широтах. Тем не менее сравнение линейного роста этого вида из крупных озёр и водохранилищ Северной Европы и северо-запада России с водоёмами Волжского бассейна показывает, что в последних этот показатель выше, причём самыми быстро растущими являются южные популяции из Саратовского водохранилища и нижнего течения Волги (рис. 2, 3). По-видимому, это объясняется наиболее благоприятными кормовыми условиями.
Рис. 2.
Темп роста налима Lota lota в водоёмах Волжского бассейна: 1 – Саратовское водохранилище (Ермолин, 1984), 2 – нижнее течение Волги (наши данные), 3 − Куйбышевское водохранилище (Махотин, 1960), 4 – оз. Селигер (Никаноров, Никанорова, 1963), 5 – Горьковское водохранилище (Кожевников, 1965), 6 – оз. Плещеево (Стрельников, Пермитин, 1983), 7 – Воткинское водохранилище (Пушкин, 1988), 8 – Рыбинское водохранилище (Сергеев, 1959), 9 – р. Кама у г. Оханск (Маркун, 1936), 10 – р. Ока (Иванчев, Иванчева, 2010), 11 – р. Волга у г. Казань (Лукин, 1935).
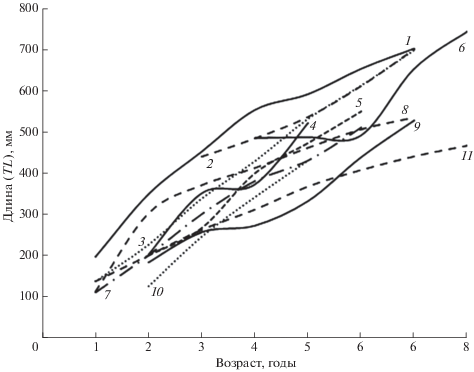
Рис. 3.
Темп роста налима Lota lota в водоёмах Северной Европы и северо-запада России: 1 – оз. Ильмень (Домрачев, Правдин, 1926), 2 – оз. Онежское (Харламов, Коваленко, 2019), 3 – оз. Кильписъярви, Финляндия (Лукин, 1935), 4 – оз. Выгозеро (Мовчан, 1988), 5 – оз. Врево (Тихомирова, Шумакова, 1979), 6 – оз. Сямозеро (Тихомирова, Шумакова, 1979), 7 – Топо-Пяозёрское водохранилище (Черепанова и др., 2019), 8 – оз. Rødenes, Норвегия (Vøllestad, 1992), 9 – оз. Ладожское (Федорова, 1979), 10 – оз. Керетьозеро (Тихомирова, Шумакова, 1979), 11 – оз. Эгдерен, Норвегия (Vøllestad, 1992), 12 – оз. Кильписъярви, Финляндия (Tolonen et al., 1999).
Основой питания взрослых особей налима обычно является рыба. Состав жертв зависит от характера их распределения и обилия в конкретной локальности (Сорокин, 1976; Герасимов и др., 2018а). Так как налим является донным хищником, наличие в водоёме мелких массовых демерсальных видов рыб определяет их доминирование в его питании. Обыкновенный ёрш Gymnocephalus cernua является основным объектом питания налима в большинстве водоёмов верхней и средней Волги (Маркун, 1936; Махотин, 1960; Кияшко, Половкова, 1983; Залозных, 1987), географически смежных с ними участках Дона (Фёдоров, 1958) и Днепра (Мовчан, 1988), а также в озёрах северо-запада России (Пиху, Пиху, 1974; Тихомирова, Шумакова, 1979; Архипцева, 1980) и Северной Европы (Vøllestad, 1992). Значимую долю составляют такие широко распространённые виды как окунь Perca fluviatilis, плотва Rutilus rutilus и пескари родов Gobio и Romanogobio (Иванова, 1963; Никаноров, Никанорова, 1963; Кожевников, 1965; Антонова, 1986; Залозных, 1987). В тех водоёмах, где высока численность таких мелких пелагических видов, как корюшки рода Osmerus и ряпушка Coregonus albula, они доминируют в питании налима (Домрачев, Правдин, 1926; Балагурова, 1966; Неличик, 1975; Фёдорова, 1979; Sandlund et al., 1985; Биоресурсы …, 2008).
Нарушение изоляции бассейна Волги и изменение её гидрорежима в результате зарегулирования в прошедшем столетии каскадом плотин повлекло за собой существенную качественно-количественную трансформацию рыбного населения реки и не в последнюю очередь вследствие расселения некоторых видов. Одним из примеров наиболее успешных инвазий рыб стали ряд донных понто-каспийских бычков и пелагическая тюлька. Они существенно расширили свои ареалы в бассейне Волги в северном направлении, натурализовавшись в ряде водохранилищ (Слынько, Терещенко, 2014). В тех водоёмах, где эти виды нарастили высокую численность (Karabanov et al., 2018), они стали составлять основу питания налима. Так, например, в Саратовском водохранилище уже в 1974−1977 гг. массовая доля тюльки, кругляка и пуголовки в питании этого хищника достигла 51% (Ермолин, 1984), в Куйбышевском в 2005−2008 гг. частота встречаемости пуголовки и кругляка – 38% (Семенов, 2009), в Рыбинском водохранилище в 2000−2015 гг. доля тюлька и кругляка составила 13−34% общего числа съеденных рыб (Рыбы Рыбинского водохранилища …, 2015). Наличие в исследованных пищеварительных трактах налима двух глубоководных видов бычков − гонца и пуголовки, вселенцев в Волжские водохранилища из Азово-Черноморского бассейна (Болдырев, 2002; Boldyrev, Bogutskaya, 2007), свидетельствует о дальнейшем расселении их в нижнее течение Волги. По нашим данным, в предплотинном участке ВГЭС доля бычковых и тюльки в питании налима достигает 84% массы пищи. В желудках нескольких налимов, добытых в 2005−2006 гг. в районе пос. Цаган-Аман, помимо бычков (песочника и кругляка) были отмечены плотва, окунь, судак Sander lucioperca, налим, рыба-игла Syngnathus abaster, а также остатки речных раков и ужа Natrix sp. (Красная книга …, 2013).
У налима наблюдается тенденция уменьшения с севера на юг размерно-весовых параметров и максимального возраста (рис. 2, 3), как это отмечено для некоторых других видов рыб (Дгебуадзе, 2001). В нижнем течении Волги масса самых крупных особей налима в районе с. Замьяны не превышает 0.9 кг (Коблицкая, 1964), у пос. Цаган-Аман – 2.1 кг (Красная книга …, 2013), на предплотинном участке ВГЭС в наших уловах – 2.4 кг, по опросным сведениям – до 3.5−4.0 кг. Причиной короткого возрастного ряда является, видимо, массовая гибель налима в жаркие годы, когда вода в июле−августе прогревается до критических для вида значений температуры. Подтверждением этому служит существенная разница в уловах налима в 2011−2013 и 2015−2018 гг. (рис. 1). Первому периоду, характеризовавшемуся относительно низкими уловами, предшествовал ряд жарких лет. Второму, когда уловы налима были значительно выше, – годы со сравнительно прохладным летом (рис. 4).
Рис. 4.
Средняя температура воздуха (г. Волгоград) (—) и воды в нижнем бьефе Волжской ГЭС
(- -) в самый жаркий месяц (июль или август) в периоды низких ( ) и высоких (
) и высоких ( ) уловов налима Lota lota.
) уловов налима Lota lota.
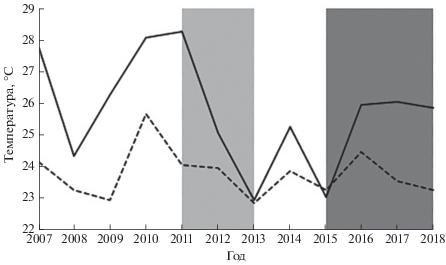
Определяющее значение температурного фактора на формирование численности налима в разных водохранилищах Волжского бассейна косвенно просматривается через изменение доли этого вида в общих уловах в зависимости от широтного расположения водоёмов. Так, в 1980−1987 гг. наибольшую долю в общем объёме добычи туводных видов рыб налим имел в самых северных водохранилищах: Рыбинском – 9.7% (при среднемноголетнем улове 269.6 т), Шекснинском (вместе с Белым озером) – 7.4% (7.4 т) и в находящемся с ними на одной широте Камском – 3.7% (9.2 т). В Горьковском водохранилище этот показатель составлял 0.7% (3.4 т), Куйбышевском – 0.4% (16.6 т), Саратовском – 0.5% (6.3 т) и в Волгоградском – 0.1% (4.1 т) (Исаева, Карпова, 1989). На незарегулированном участке Волги в границах Волгоградской области в 1965−1971 гг. доля налима в общих уловах (без проходных видов) составляла 0.3% при среднемноголетнем вылове 0.5 т (Чехова, Пономарева, 1973). Интенсивное потепление в 2000-е гг. рассматривается как основная причина снижения численности популяций налима в некоторых волжских водоёмах (Современное состояние …, 2004; Герасимов и др., 2018б).
Список литературы
Антонова Е.Л. 1986. Питание хищных рыб Воткинского водохранилища // Сб. науч. тр. ГосНИОРХ. Вып. 250. С. 41–45.
Архипцева Н.Т. 1980. Ихтиофауна и численность рыб озера Красного // Там же. Вып. 158. С. 94–99.
Балагурова М.В. 1966. Материалы по питанию налима Сямозера // Тр. Карел. отд. ГосНИОРХ. Т. 4. Вып. 2. С. 55–70.
Биоресурсы Онежского озера. 2008 / Под ред. Кухарева В.И., Лукина А.А. Петрозаводск: Изд-во КарНЦ РАН, 272 с.
Богданов В.Д., Копориков А.Р. 2011. Воспроизводство налима нижней Оби. Екатеринбург: Изд-во УрО РАН, 156 с.
Богуцкая Н.Г., Болдырев В.С., Насека А.М. 2004. Бычки Neogobiinae (Teleostei, Gobiidae) в экосистемах Евразии и Североамериканских великих озер // Биологические инвазии в водных и наземных экосистемах М.: Т-во науч. изд. КМК. С. 297–320.
Богуцкая Н.Г., Кияшко П.В., Насека А.М., Орлова М.И. 2013. Определитель рыб и беспозвоночных Каспийского моря. Т. 1. Рыбы и моллюски. СПб.: Т-во науч. изд. КМК, 543 с.
Болдырев В.С. 2002. Видовой состав бычков (Gobiidae) в Волгоградском и Цимлянском водохранилищах // Рыбохозяйственные исследования в бассейне Волго-Донского междуречья на современном этапе. СПб.: Изд-во ГосНИОРХ. С. 98–105.
Вехов Д.А., Горский К. 2010. Состав ихтиофауны водоемов северной части Волго-Ахтубинской поймы // Матер. науч.-практ. конф. “ООПТ Нижней Волги как важнейший механизм сохранения биоразнообразия: итоги, проблемы и перспективы”. Волгоград. С. 58–64.
Герасимов Ю.В., Иванова М.Н., Свирская А.Н. 2018а. Многолетние изменения роли местных и инвазийных видов рыб в питании хищных рыб Рыбинского водохранилища // Вопр. ихтиологии. Т. 58. № 5. С. 507–522 .https://doi.org/10.1134/S0042875218040045
Герасимов Ю.В., Малин М.И., Соломатин Ю.И. и др. 2018б. Распределение и структура рыбного населения в водохранилищах Волжского каскада в 1980-е и 2010-е гг. // Тр. ИБВВ РАН. Вып. 82 (85). С. 82–106. https://doi.org/10.24411/0320-3557-2018-1-0014.
Голованов В.К. 2014. Температурные критерии жизнедеятельности рыб. Прогноз, экологические риски и экспертная оценка // Матер. II Всерос. конф. “Современное состояние биоресурсов внутренних вод”. Т. 1. М.: Полиграф-Плюс. С. 162–170.
Гольдентрахт И.Н. 1966. Промыслово-биологические исследования в Волго-Ахтубинской пойме в 1959 г. // Тр. КаспНИРХ. Т. 22. С. 6–24.
Гримм О.А. 1896. Каспийско-волжское рыболовство. СПб.: Тип. В. Демакова, 153 с.
Дгебуадзе Ю.Ю. 2001. Экологические закономерности изменчивости роста рыб. М.: Наука, 275 с.
Домрачев П.Ф., Правдин И.Ф. 1926. Рыбы озера Ильменя и реки Волхова и их хозяйственное значение // Материалы исследований р. Волхова и его бассейна. Вып. 10. Ч. 1. Л.: Стр-во ВолховГЭСУ, 296 с.
Ермолин В.П. 1984. Экология питания рыб и пути повышения рыбопродуктивности Саратовского водохранилища: Автореф. дис. … канд. биол. наук. Л.: ГосНИОРХ. 24 с.
Залозных Д.В. 1987. Питание хищных рыб Чебоксарского водохранилища в первые годы его существования // Сб. науч. тр. ГосНИОРХ. Вып. 267. С. 118–128.
Иванова М.Н. 1963. О питании хищных рыб в первые четыре года существования Горьковского водохранилища // Тр. ИБВ АН СССР. Вып. 5 (8). С. 81–86.
Иванчев В.П., Иванчева Е.Ю. 2010. Круглоротые и рыбы Рязанской области и прилежащих территорий. Рязань: Голос губернии, 292 с.
Исаев А.И., Карпова Е.И. 1989. Рыбное хозяйство водохранилищ. Справочник. М.: Агропромиздат, 255 с.
Казанчеев Е.Н. 1981. Рыбы Каспийского моря. М.: Лёг. и пищ. пром-сть, 168 с.
Кияшко В.И., Половкова С.Н. 1983. Питание и пищевые взаимоотношения рыб оз. Плещеево // Тр. ИБВВ АН СССР. Вып. 51 (54). С. 112–125.
Коблицкая А.Ф. 1964. О нересте налима Lota lota L. в дельте Волги // Вопр. ихтиологии. Т. 4. Вып. 2(81). С. 392–393.
Кожевников Г.П. 1965. Формирование рыбных запасов Горьковского водохранилища в первые годы его существования // Изв. ГосНИОРХ. Т. 59. С. 43–97.
Красная книга Республики Калмыкия. 2013. Т. 1. Животные. Элиста: Джангар, 200 с.
Литвинов К.В., Подоляко С.А. 2013. Видовой состав и состояние ихтиофауны низовьев дельты Волги в 2006−2011 гг. в пределах Астраханского государственного природного заповедника. Приложение 2 // Определитель рыб и беспозвоночных Каспийского моря. Т. 1. Рыбы и моллюски. СПб.; М.: Т-во науч. изд. КМК. С. 516–525.
Лукин А.В. 1935. К биологии волжского налима (Lota lota (Linne)) // Тр. Татар. отд. ВНИОРХ. Вып. 2. С. 85–86.
Маркун М.И. 1936. К систематике и биологии налима р. Камы // Изв. БиолНИИ при ПГУ. Т. 10. Вып. 6. С. 211–238.
Махотин Ю.М. 1960. Рост, питание и плодовитость налима Куйбышевского водохранилища // Тр. Татар. отд. ГосНИОРХ. Вып. 9. С. 291–296.
Медведев В.Н. 2013. Рыболовный промысел населения Сарепты и Черноярского уезда Астраханской губернии // Этнограф. обозрение. № 5. С. 14–21.
Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях. 1974. М.: Наука, 254 с.
Мовчан Ю.В. 1988. Фауна Украины. Т. 8. Рыбы. Вып. 3. Киев: Наук. думка, 366 с.
Неличик В.А. 1975. Питание налима в Верхнетуломском водохранилище // Сб. науч. тр. ГосНИОРХ. № 14. С. 50–55.
Никаноров Ю.И., Никанорова Е.А. 1963 Рыбы озера Селигер и их биология // Тр. Осташков. отд. ГосНИИОРХ. Т. 1. С. 70–145.
Пиху Э.Х., Пиху Э.Р. 1974. Питание основных хищных рыб Псковско-Чудского водоема // Изв. ГосНИОРХ. Т. 83. С. 136–143.
Пушкин Ю.А. 1988. Ихтиофауна и рыбное хозяйство // Биология Воткинского водохранилища. Иркутск: Изд-во ИГУ. С. 118–143.
Решетников Ю.С., Мухачев И.С., Болотова Н.Л. и др. 1989. Пелядь Coregonus peled (Gmelin, 1788): систематика, морфология, экология, продуктивность. М.: Наука, 302 с.
Решетников Ю.С., Попова О.А., Кияшко В.И. и др. 2016. Обыкновенный ёрш Gymnocephalus cernuus (Linnaeus, 1758). Систематика, морфология, образ жизни и роль ерша в экосистемах. М.: Т-во науч. изд. КМК, 279 с.
Рыбы Рыбинского водохранилища: популяционная динамика и экология. 2015 / Под ред. Герасимова Ю.В. Ярославль: Филигрань, 418 с.
Семенов Д.Ю. 2009. Роль чужеродных видов в питании хищных рыб Куйбышевского водохранилища // Поволж. экол. журн. № 2. С. 148–157.
Сергеев Р.С. 1959. Материалы по биологии налима Рыбинского водохранилища // Тр. ИБВ АН СССР. Вып. 1 (4). С. 235–258.
Слынько Ю.В., Терещенко В.Г. 2014. Рыбы пресных вод Понто-Каспийского бассейна (разнообразие, фауногенез, динамика популяций, механизмы адаптаций). М.: Полиграф-Плюс, 328 с.
Современное состояние рыбного хозяйства на внутренних водоемах России. 2004 / Под ред. Иванова Д.И., Печникова А.С. СПб.: Изд-во ГосНИОРХ, 580 с.
Сорокин В.Н. 1976. Налим озера Байкал. Новосибирск: Наука, 144 с.
Стрельников А.С., Пермитин И.Е. 1983. Ихтиофауна оз. Плещеево и состояние рыболовства // Тр. ИБВВ АН СССР. Вып. 51 (54). С. 97–112.
Тихомирова Л.П., Шумакова Е.Н. 1979. Промысловое значение и биология налима озера Врево // Сб. науч. тр. ГосНИОРХ. Вып. 141. С. 55–58.
Федоров А.В. 1958. О рыбохозяйственном значении хищных рыб бассейна Верхнего Дона // Тр. ВоронежГУ. Т. 45. Вып. 1. С. 35–53.
Федорова Г.В. 1979. Налим Ладожского озера // Сб. науч. тр. ГосНИОРХ. Вып. 141. С. 50–54.
Харламов А.М., Коваленко В.Н. 2019. Рыбохозяйственная характеристика налима – Lota lota (L.) Онежского озера на современном этапе // Матер. II Междунар. конф. “Озера Евразии: проблемы и пути их решения”. Казань. С. 346–351.
Черепанова Н.С., Широков В.А., Коваленко В.Н. 2019. Характеристика роли налима (Lota lota L.) в рыбном промысле Топо-Пяозерского водохранилища (Карелия) // Матер. VII Междунар. Балт. мор. форума. Т. 3. С. 178–183.
Чехова М.А., Пономарева Э.Н. 1973. Рыбное хозяйство внутренних водоёмов Волгоградской и Ростовской областей. Статистический справочник // Тр. Волгоград. отд. ГосНИОРХ. Т. 7. 174 с.
Шатуновский М.И., Дгебуадзе Ю.Ю., Бобырев А.Е. и др. 2009. Некоторые закономерности изменчивости структуры и динамики популяций леща Abramis brama водоемов Восточной Европы // Вопр. ихтиологии. Т. 49. № 4. С. 495–507.
Boldyrev V.S., Bogutskaya N.G. 2007. Revision of the tadpole-gobies of the genus Benthophilus (Teleostei: Gobiidae) // Ichthyol. Explor. Freshwat. V. 18. № 1. P. 31–96.
Froese R., Pauly D. (eds.). 2019. FishBase. World Wide Web electronic publication. (www.fishbase.org. Version 12/2019)
Karabanov D.P., Pavlov D.D., Bazarov M.I. et al. 2018. Alien species of fish in the littoral of Volga and Kama reservoirs (Results of complex expeditions of IBIW RAS in 2005–2017) // Transact. IBIW RAS. Iss. 82 (85). P. 67–80. https://doi.org/0.24411/0320-3557-2018-1-0013.
Kottelat M., Freyhof J. 2007. Handbook of European freshwater fishes. Cornol; Berlin: Kottelat et Freyhof, 646 p.
Sandlund O.T., Klyve L., Næsje T.F. 1985. Growth, habitat and food of burbot Lota lota in Lake Mjøsa // Fauna. V. 38. P. 37–43.
Tolonen A., Kjellman J., Lappalainen J. 1999. Diet overlap between burbot (Lota lota (L.)) and whitesh (Coregonus lavaretus (L.)) in a subarctic lake // Ann. Zool. Fennici. V. 36. P. 205–214.
Vøllestad L.A. 1992. Age, growth and food of the Burbot Lota lota in two eutrophic lakes in southeast Norway // Fauna Norv. Ser. A. V. 13. P. 13–18.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии