

Вопросы ихтиологии, 2021, T. 61, № 4, стр. 464-470
Отдалённые последствия влияния медленных флуктуаций магнитного поля на эмбрионы плотвы Rutilus rutilus (Cyprinidae)
В. В. Крылов 1, *, Ю. В. Чеботарева 1, Ю. Г. Изюмов 1
1 Институт биологии внутренних вод РАН – ИБВВ РАН
пос. Борок, Ярославская область, Россия
* E-mail: kryloff@ibiw.yaroslavl.ru
Поступила в редакцию 10.07.2020
После доработки 30.07.2020
Принята к публикации 03.08.2020
Аннотация
Описаны отдалённые эффекты воздействия вариаций магнитного поля интенсивностью 100, 300 и 500 нТл и продолжительностью 6 ч на эмбрионы плотвы Rutilus rutilus. Экспозиция продолжалась со 2-го по 7-й или с 33-го по 38-й ч после оплодотворения. Воздействие исследуемых факторов привело впоследствии к изменениям размерно-массовых показателей и перераспределению позвонков между отделами позвоночника у сеголеток. Дисперсионный анализ выявил значимое влияние срока экспозиции эмбрионов и взаимодействия факторов интенсивности магнитных флуктуаций, а также разных сроков экспозиции на исследуемые показатели.
Все живые организмы на нашей планете существуют в условиях геомагнитного поля, которому свойственны незначительные периодические изменения – геомагнитные вариации (Акасофу, Чепмен, 1975). В частности, медленные изменения индукции на несколько десятков нанотесла с периодом 24 ч называют суточной геомагнитной вариацией (Yamazaki, Maute, 2017). Антропогенные и естественные изменения магнитного фона могут оказывать влияние на биологические объекты (Бинги, 2011), включая рыб (Крылов и др., 2013). Одним из наиболее заметных естественных нарушений привычного магнитного окружения являются геомагнитные бури различной интенсивности. Биологические эффекты имитации этого фактора в эксперименте (Таликина и др., 2013; Golovanova et al., 2017; Krylov et al., 2019a) подтверждают известную со второй половины прошлого века гипотезу о том, что геомагнитная буря может воздействовать на биологические объекты за счёт нарушения регулярной структуры суточной геомагнитной вариации, используемой в качестве внешнего водителя биологических циркадных ритмов (Bliss, Heppner, 1976; Brown, Chow, 1976; Welker et al., 1983; Крылов, 2017; Голованова и др., 2019).
В природе происходят различные по силе возмущения естественного магнитного фона, причём частота возникновения геомагнитных бурь обратно пропорциональна их интенсивности. Однако в годы максимумов солнечной активности число сильных геомагнитных бурь может быть существенным (Безродных и др., 2019). Проведённые ранее эксперименты, подразумевающие экспозицию организмов в имитации геомагнитных возмущений, не ставили задачи оценить зависимость проявления биологических эффектов от интенсивности сигнала, имитирующего геомагнитную бурю. Учитывая нелинейную зависимость биологических эффектов слабых магнитных полей от величины индукции (Леднев и др., 1996; Белова, Леднев, 2001), такие данные представляют существенный интерес.
Влияние на рыб магнитных полей, имитирующих естественные геомагнитные события, детально исследовано на примере плотвы Rutilus rutilus. В частности, установлено, что экспонирование эмбрионов плотвы в имитации геомагнитной бури в течение первых 24 ч развития оказывает стимулирующее действие на пролиферацию зародышевых клеток (Таликина и др., 2013). Оценка морфологических показателей у сеголеток, развившихся из экспонированных эмбрионов, показала значимое снижение размерно-массовых показателей. Кроме этого, после экспозиции эмбрионов в имитации геомагнитной бури у сеголеток наблюдали значимое перераспределение числа позвонков в отделах позвоночника и изменения в числе отверстий в каналах сейсмосенсорной системы на нижнечелюстных и предкрышечных костях черепа (Krylov et al., 2019a). Следует также отметить, что воздействие этого фактора на половые продукты плотвы до оплодотворения не сказывается на оцениваемых показателях (Изюмов и др., 2015). Однако в литературе отсутствуют сведения об эффектах влияния различных по интенсивности сигналов, имитирующих геомагнитные события, на эмбрионы R. rutilus.
Цель работы – изучить во время эмбриогенеза отдалённые последствия влияния различных по интенсивности сигналов, имитирующих геомагнитные события (ИГС), на размерные и морфометрические показатели сеголеток плотвы.
МАТЕРИАЛ И МЕТОДИКА
Объектом экспозиции в магнитных флуктуациях были эмбрионы плотвы. Производителей отловили в мае 2013 г. в Рыбинском водохранилище. 13 мая в 10:45 от четырёх самок и восьми самцов были получены половые продукты. Осеменение икры проводили сухим способом. Затем 2500–3000 оплодотворенных икринок размещали на дне семи кристаллизаторов с речной водой (высота 7.5 см, диаметр 23 см). Воду в кристаллизаторах ежедневно заменяли утром и вечером. Все манипуляции с тестируемыми объектами проводили одновременно. Температура воды в кристаллизаторах при проведении экспериментов составила в среднем 18°С.
Эмбрионы подвергались воздействию магнитных флуктуаций, имитирующих главную фазу и фазу восстановления умеренной, сильной и экстремальной геомагнитной бури в диапазоне 0–0.001 Гц. Продолжительность воздействия составила 6 ч. При этом в течение 3 ч магнитная индукция линейно отклонялась от базового уровня до максимального значения, в течение последующих 3 ч возвращалась к исходному состоянию (рисунок). Продолжительность применённого воздействия соответствует длительности главной фазы типичной геомагнитной бури, а также продолжительности наиболее выраженных изменений индукции поля вследствие суточной геомагнитной вариации. Размах сигналов, имитирующих умеренную, сильную и экстремальную бурю, составил ~100, 300 и 500 нТл соответственно.
Флуктуации магнитного поля в направлении горизонтальной D-компоненты в контроле (а)
и в вариантах опыта с имитацией геомагнитных событий (ИГС) разной интенсивности при
экспозиции эмбрионов плотвы Rutilus rutilus в течение 2−7-го (б−г) и 33−38-го (д−ж) ч п.о.: а − в контроле заметна регулярная
суточная вариация; б, д − ИГС 100 нТл; в, е − ИГС 300 нТл; г, ж − ИГС 500 нТл; (↕)
– момент осеменения икры, ( ) − применённые воздействия.
) − применённые воздействия.
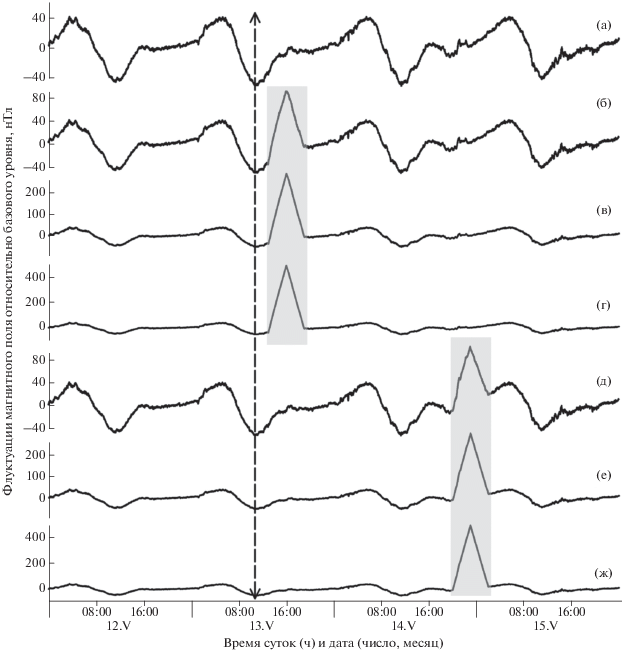
Генерация ИГС происходила в системе из трёх пар колец Гельмгольца в направлении x, y и z-компонент геомагнитного поля. Указанные выше сигналы поступали в систему через цифро-аналоговый преобразователь LTR-EU-8 (ЗАО “Л-кард”, Москва). Подробное описание экспериментальной установки, использованной в данной работе, опубликовано ранее (Крылов и др., 2011). В экспериментах исследовано шесть вариантов воздействия. Эмбрионы трёх опытных групп были экспонированы в ИГС с размахом амплитуды колебаний ~100, 300 и 500 нТл со 2-го по 7-й ч после оплодотворения включительно (2−7 ч п.о.), т. е. во время дробления (далее до гаструляции); пик ИГС пришёлся на 16:00 ч (рисунок, б, в, г). В трёх других группах эмбрионы подвергались тем же воздействиям, но в течение 33–38-го ч п.о., т. е. в начале органогенеза (далее после гаструляции); пик ИГС пришёлся на 23:00 ч (рисунок, д, е, ж). Контрольные эмбрионы находились в немодифицированном спокойном геомагнитном поле (рисунок, а).
Различия в выживаемости эмбрионов исследуемых вариантов были незначительны. Вылупление предличинок в контроле и опыте было синхронным. По 400 личинок из каждой группы после появления плавательного пузыря переместили в пруды прудового стационара ИБВВ РАН “Сунога” на 4 мес. К моменту вылова выживаемость плотвы в прудах составила 76−82%.
Отдалённые последствия действия ИГС на эмбрионы плотвы оценивали на основе анализа размерно-массовых и морфологических показателей 4-месячных сеголеток. Определяли длину до конца чешуйного покрова (SL) и массу тела. Подсчитывали число лучей в плавниках и число прободённых чешуй в боковой линии (ll). Скелет препарировали по методике Яковлева с соавторами (1981). Подсчитывали число позвонков в грудном (vert.а), переходном (vert.i.) и хвостовом (vert.c.) отделах, а также их сумму (vert.), включая Веберовы и преуральные. К позвонкам переходного отдела относили последние позвонки туловищного отдела, отличающиеся от типичных туловищных наличием развитых парапофизов, срастающихся с их телами, а от хвостовых – отсутствием нижнего остистого отростка. На очищенных от мягких тканей костях черепа подсчитывали число отверстий в каналах сейсмосенсорной системы на парных костях черепа – dentale, frontale, parietale и praeoperculum.
Полученные данные были протестированы на нормальность распределения (критерий Колмогорова−Смирнова или Шапиро−Уилка) и гомогенность (критерий Левена). Если условия были соблюдены (длина и масса тела, число прободённых чешуй в боковой линии, число позвонков), то значимость различий между средними значениями оценивали при помощи апостериорного критерия Даннета после проведения дисперсионного анализа. При ненормальном распределении данных использовали критерий Краскела−Уоллеса. Для определения влияния интенсивности ИГС и разных сроков экспозиции эмбрионов на изученные характеристики использовали двухфакторный дисперсионный анализ.
РЕЗУЛЬТАТЫ
Длина и масса сеголеток, подвергавшихся действию ИГС разной интенсивности до гаструляции (2−7 ч п.о.), были больше, чем у контрольных особей, на 1.5–16.6%, причём отличия вариантов ИГС 100 и 300 нТл от контроля были статистически значимы. Если же экспозиция эмбрионов в ИГС происходила после гаструляции (33−38 ч п.о.), то позднее наблюдали значимое снижение размерно-массовых показателей у молоди на 4.6–47.4% относительно контроля (табл. 1). Следует отметить, что наиболее существенное увеличение и снижение длины и массы у рыб отмечалось после воздействия ИГС с интенсивностью 100 нТл на эмбрионы соответственно до и после гаструляции.
Таблица 1.
Исследуемые показатели (M ± SE) сеголеток плотвы Rutilus rutilus, которые испытали на разных отрезках эмбриогенеза воздействие сигналов, имитирующих геомагнитные события (ИГС), разной интенсивности
| Признак | Контроль (96−97) |
Период воздействия, ч п.о./интенсивность ИГС, нТл (число рыб, экз.) | |||||
|---|---|---|---|---|---|---|---|
| 2−7 | 33−38 | ||||||
| 100 (94−96) |
300 (57−59) |
500 (61−64) |
100 (95−97) |
300 (81−98) |
500 (61−64) |
||
| SL, мм | 73.90 ± 0.42 | 78.99 ± 0.45* | 77.16 ± 0.58* | 75.00 ± 0.42 | 61.23 ± 0.30* | 67.94 ± 0.36* | 70.49 ± 0.32* |
| Масса, г | 7.70 ± 0.11 | 8.98 ± 0.15* | 8.71 ± 0.18* | 8.02 ± 0.14 | 4.05 ± 0.06* | 5.55 ± 0.09* | 6.08 ± 0.08* |
| ll | 43.24 ± 0.16 | 43.71 ± 0.17 | 42.47 ± 0.20* | 42.54 ± 0.19* | 42.66 ± 0.15 | 42.72 ± 0.18 | 43.83 ± 0.17 |
| P | 15.51 ± 0.06 | 15.53 ± 0.06 | 15.71 ± 0.08 | 15.55 ± 0.07 | 15.34 ± 0.06 | 15.43 ± 0.06 | 15.29 ± 0.07 |
| V | 8.01 ± 0.02 | 7.99 ± 0.03 | 8.08 ± 0.04 | 8.03 ± 0.03 | 7.96 ± 0.03 | 7.96 ± 0.02 | 7.96 ± 0.03 |
| D | 10.09 ± 0.04 | 10.11 ± 0.04 | 10.07 ± 0.05 | 10.03 ± 0.04 | 10.11 ± 0.04 | 10.18 ± 0.04 | 10.05 ± 0.04 |
| A | 10.27 ± 0.05 | 10.30 ± 0.05 | 10.08 ± 0.06 | 10.10 ± 0.05 | 10.13 ± 0.04 | 10.23 ± 0.05 | 10.18 ± 0.05 |
| vert. | 42.00 ± 0.13 | 42.82 ± 0.17* | 41.68 ± 0.16 | 41.23 ± 0.14* | 41.99 ± 0.15 | 41.91 ± 0.11 | 42.95 ± 0.15* |
| vert.a. | 16.52 ± 0.09 | 16.41 ± 0.09 | 16.03 ± 0.13* | 16.11 ± 0.10* | 16.55 ± 0.11 | 16.44 ± 0.09 | 16.43 ± 0.11 |
| vert.i. | 3.35 ± 0.06 | 3.70 ± 0.07* | 3.54 ± 0.10 | 3.23 ± 0.07 | 3.33 ± 0.07 | 3.23 ± 0.06 | 3.43 ± 0.07 |
| vert.c. | 15.13 ± 0.08 | 15.72 ± 0.09* | 15.10 ± 0.12 | 14.89 ± 0.09 | 15.12 ± 0.07 | 15.23 ± 0.07 | 16.09 ± 0.09* |
| Fr | 5.15 ± 0.05 | 5.01 ± 0.06 | 5.14 ± 0.08 | 4.88 ± 0.10 | 5.24 ± 0.06 | 5.11 ± 0.06 | 4.99 ± 0.08 |
| ParSo | 0.80 ± 0.05 | 0.76 ± 0.05 | 0.74 ± 0.06 | 0.78 ± 0.05 | 0.80 ± 0.04 | 0.63 ± 0.05 | 0.73 ± 0.05 |
| ParSt | 1.95 ± 0.05 | 1.74 ± 0.06 | 1.90 ± 0.07 | 1.95 ± 0.06 | 1.94 ± 0.06 | 2.00 ± 0.05 | 1.77 ± 0.07 |
| Dent | 3.30 ± 0.06 | 3.36 ± 0.06 | 3.32 ± 0.07 | 3.44 ± 0.08 | 3.62 ± 0.07 | 3.30 ± 0.07 | 3.42 ± 0.07 |
| Pr | 7.77 ± 0.08 | 8.09 ± 0.08 | 7.78 ± 0.10 | 8.37 ± 0.10* | 7.91 ± 0.10 | 7.78 ± 0.09 | 8.05 ± 0.09 |
Примечание. M ± SE – среднее значение и стандартная ошибка; SL – длина тела до конца чешуйного покрова, ll – число чешуй в боковой линии; P, V, D, A – число мягких лучей соответственно в грудном, брюшном, спинном и анальном плавниках; vert., vert.а., vert.i., vert.c. – число позвонков соответственно общее (включая Веберовы и преуральные), в грудном, переходном и хвостовом отделах; число отверстий каналов сейсмосенсорной системы в костях: Fr – frontale; ParSo, ParSt − parietale в субокулярном и субтемпоральном каналах; Dent – dentale, Pr – praeoperculum; * отличия от контроля значимы при p < 0.05 (однофакторный дисперсионный анализ, критерии Даннета и Краскела−Уоллеса).
Дисперсионный анализ выявил значимое влияние интенсивности ИГС, срока экспозиции эмбрионов и взаимодействия этих факторов на длину и массу сеголеток (табл. 2). Причём наибольшей силой обладает срок экспозиции эмбрионов, интенсивность же ИГС влияет на эти показатели в меньшей степени.
Таблица 2.
Результаты дисперсионного анализа значений длины (SL) и массы сеголеток плотвы Rutilus rutilus, испытавших на разных отрезках эмбриогенеза воздействие сигналов, имитирующих геомагнитные события (ИГС), разной интенсивности
| Фактор | Сумма квадратов | Степени свободы | F | p | η2 |
|---|---|---|---|---|---|
| SL | |||||
| Интенсивность ИГС [1 ] | 744.66 | 2 | 28.11 | <0.001 | 0.10 |
| Срок экспонирования [2 ] | 13 016.89 | 1 | 982.89 | <0.001 | 0.67 |
| Взаимодействие [1] × [2 ] | 3804.35 | 2 | 143.63 | <0.001 | 0.37 |
| Ошибка | 6449.58 | 487 | |||
| Масса | |||||
| Интенсивность ИГС [1 ] | 39.13 | 2 | 17.05 | <0.001 | 0.07 |
| Срок экспонирования [2 ] | 1319.27 | 1 | 1149.73 | <0.001 | 0.70 |
| Взаимодействие [1] × [2 ] | 188.78 | 2 | 82.26 | <0.001 | 0.25 |
| Ошибка | 558.81 | 487 | |||
Экспозиция зародышей в ИГС разной интенсивности до гаструляции (2−7 ч п.о.) привела к перераспределению позвонков между отделами позвоночника рыб. В вариантах 300 и 500 нТл это выразилось в значимом уменьшении относительно контроля vert.а., а в варианте 100 нТл регистрировали значимое увеличение vert.i. и vert.c. (табл. 1). Следствием этого перераспределения стали различия в общем числе позвонков: в варианте 100 нТл наблюдали значимое увеличение этого показателя, в варианте 500 нТл, напротив, снижение относительно контрольного значения. Экспонирование в ИГС эмбрионов после гаструляции (33−38 ч п.о.) не повлияло на позвонковые фенотипы, за исключением наиболее интенсивного воздействия. В варианте 500 нТл отмечено увеличение общего числа позвонков за счёт увеличения vert.c.
При этом дисперсионный анализ выявил значимое влияние срока экспозиции эмбрионов на vert.а. (F1, 487 = 10.9, p < 0.01, η2 = 2.2%) и взаимодействия факторов интенсивности ИГС и срока экспозиции на vert.i. (F2, 487 = 8.4, p < 0.001, η2 = = 3.3%) и vert.c. (F2, 487 = 53.3, p < 0.001, η2 = 18.0%). На общее число позвонков влияла интенсивность ИГС (F2, 487 = 8.6, p < 0.001, η2 = 3.4%) и взаимодействие интенсивности ИГС и срока экспозиции эмбрионов (F2, 487 = 36.0, p < 0.001, η2 = 12.9%).
Только воздействие ИГС на ранних этапах эмбриогенеза (2−7 ч п.о.) приводило впоследствии к значимому увеличению у сеголеток плотвы числа отверстий сейсмосенсорной системы в предкрышечных костях (вариант 500 нТл), а также к небольшому, но значимому снижению числа прободённых чешуй в боковой линии (варианты 300 и 500 нТл).
ОБСУЖДЕНИЕ
Влияние магнитных флуктуаций во время раннего развития на размерно-массовые показатели плотвы было описано ранее (Чеботарева и др., 2009; Голованова и др., 2013; Krylov et al., 2016, 2019a; Golovanova et al., 2017). Мы рассматриваем изменения этих показателей как отдалённый эффект магнитного воздействия, поскольку вероятность различий в кормовой базе между прудами минимальна. Отдельный анализ длины и массы сеголеток плотвы, выращенных в прудах биологического стационара “Сунога” в разные годы, не выявил зависимости между конкретными прудами и размерами рыб (Krylov et al., 2019b). Сложная цепь процессов, которая может лежать от воздействия магнитных флуктуаций на конкретную биологическую мишень во время эмбриогенеза до изменения размерно-массовых показателей сеголеток, не позволяет убедительно обсуждать механизмы этих эффектов. Однако мы можем предположить, что ИГС в раннем онтогенезе оказало влияние на пищеварительную систему плотвы. Известно, что активность пищеварительных ферментов у рыб напрямую зависит от магнитных полей (Кузьмина и др., 2015; Li et al., 2015). Изменения активности пищеварительных гликозидаз регистрировали у сеголеток плотвы, подвергавшихся воздействию магнитных флуктуаций во время эмбрионального развития (Голованова и др., 2013; Golovanova et al., 2017).
Примечательно обнаруженное влияние разных сроков экспозиции эмбрионов в ИГС на изменения размерно-массовых показателей. Подобная зависимость была показана в экспериментах с использованием имитации геомагнитной бури (Krylov et al., 2019a). В опытах с низкочастотным магнитным полем (500 Гц, 150 мкТл) только экспозиция эмбрионов в течение первых 27 ч п.о. приводила впоследствии к снижению длины и массы сеголеток. Магнитное воздействие во время более поздних отрезков эмбриогенеза не приводило к значимым отличиям от контроля (Чеботарева и др., 2009). Полученные в рамках настоящей работы результаты подтверждают предположение о том, разные этапы эмбриогенеза плотвы различаются по чувствительности к магнитным воздействиям. Наиболее существенное изменение длины и массы рыб после воздействия ИГС с интенсивностью 100 нТл на эмбрионы, вероятно, связано с тем, что в естественной среде магнитные события с таким размахом флуктуаций встречаются чаще, чем геомагнитные бури 300−500 нТл (Loewe, Prolss, 1997; Безродных и др., 2019).
Эффекты ИГС проявлялись также в изменении числа позвонков, их перераспределении между отделами позвоночника и в снижении числа прободённых чешуй в боковой линии. Позвонки формируются напротив миосепт, которые обособляются на 5–7-м зародышевых этапах (Ланге, Дмитриева, 1981; Павлов, 2007). Кальцификация тел позвонков в позвоночнике плотвы начинается на 1-м личиночном этапе (Ванюшина, 1990). В начале малькового периода дифференцируются чешуи боковой линии (Дислер, 1960). Перераспределение позвонков и изменение числа прободённых чешуй в боковой линии у экспериментальных рыб в ответ на действие ИГС в течение 2−7-го ч п.о. может говорить о том, что эти признаки модулируются магнитным воздействием довольно рано, до момента их чёткого анатомического определения.
В одном из экспериментальных вариантов было зарегистрировано увеличение числа отверстий сейсмосенсорной системы в предкрышечных костях. Этот эффект, на наш взгляд, также заслуживает внимания, поскольку ранее подобные изменения были обнаружены у сеголеток плотвы после экспозиции в низкочастотных магнитных полях и в имитации геомагнитной бури во время эмбриогенеза (Krylov et al., 2016, 2019a). Появление отверстий в каналах сейсмосенсорной системы зависит от темпов развития костей черепа. По нашим наблюдениям, у молоди плотвы SL 2.5−3.0 см в возрасте 2 мес. в зависимости от условий развития костные сейсмосенсорные каналы на костях черепа нередко отсутствуют, незамкнуты или развиты только частично. То есть их формирование к этому времени завершается не у всех особей. Изменение числа отверстий в каналах сейсмосенсорной системы в ответ на ИГС в раннем эмбриогенезе говорит о том, что магнитное воздействие может влиять на баланс между темпами закладки и развития периферической нервной системы и висцерального скелета рыб.
Дисперсионный анализ выявил заметное влияние срока экспозиции эмбрионов и взаимодействия факторов интенсивности ИГС, а также срока экспозиции на исследуемые показатели. Зависимость различных эффектов от того отрезка в раннем развитии, на который приходится воздействие, закономерно в связи с различиями, присущими организму на разных этапах эмбриогенеза. Обнаруженные существенные эффекты взаимодействия факторов интенсивности ИГС и срока экспозиции отражают сложную картину восприимчивости эмбрионов к воздействию исследуемых магнитных флуктуаций.
Полученные результаты подтверждают обнаруженную ранее чувствительность размерно-массовых показателей, числа позвонков в отделах позвоночника и числа отверстий сейсмосенсорной системы в костях черепа к различным магнитным воздействиям во время раннего развития. В частности, в других наших работах перечисленные показатели у сеголеток плотвы значимо отличались от контроля после экспозиции эмбрионов в переменных магнитных полях с интенсивностью 1.5 мкТл и частотой 72.5 или 500 Гц (Krylov et al., 2016), а также в имитации геомагнитной бури (Krylov et al., 2019a). Повторяемость результатов позволяет предложить данный комплекс эффектов, включающий модификации размерно-массовых показателей, изменение числа отверстий сейсмосенсорной системы в костях черепа и числа позвонков в отделах позвоночника, как индикатор различных магнитных воздействий на плотву во время раннего развития.
Список литературы
Акасофу С.И., Чепмен С. 1975. Солнечно-земная физика. Ч. 2. М.: Мир, 512 с.
Безродных И.П., Морозова Е.И., Петрукович А.А., Кожухов М.В. 2019. Динамика солнечной и геомагнитной активности. II. Периодические вариации солнечной и геомагнитной активности // Вопр. электромеханики. Тр. ВНИИЭМ. Т. 171. № 4. С. 24−38.
Белова Н.А., Леднев В.В. 2001. Влияние крайне слабых переменных магнитных полей на гравитропизм растений // Биофизика. Т. 46. № 1. С. 122−125.
Бинги В.Н. 2011. Принципы электромагнитной биофизики. М.: Физматлит, 592 с.
Ванюшина О.Г. 1990. Развитие осевого скелета у леща (Abramis brama L.) и плотвы (Rutilus rutilus L.) // Тр. ИБВВ РАН. Т. 59 (62). С. 4−9.
Голованова И.Л., Филиппов А.А., Крылов В.В. и др. 2013. Действие магнитного поля и меди на активность гидролитических ферментов у сеголеток плотвы Rutilus rutilus // Вопр. ихтиологии. Т. 53. № 2. С. 227−232.https://doi.org/10.7868/S0042875213020045
Голованова И.Л., Филиппов А.А., Чеботарева Ю.В. и др. 2019. Влияние смещения суточной геомагнитной вариации на активность пищеварительных гликозидаз молоди плотвы и их чувствительность к тяжелым металлам // Геофиз. процессы и биосфера. Т. 18. № 4. С. 49–55. https://doi.org/10.21455/GPB2019.4-5
Дислер Н.Н. 1960. Органы чувств системы боковой линии и их значение в поведении рыб. М.: Изд-во АН СССР, 312 с.
Изюмов Ю.Г., Таликина М.Г., Крылов В.В. 2015. Митоз бластомеров, вылупление, выживаемость и размерные характеристики предличинок плотвы Rutilus rutilus после действия главной фазы сильной магнитной бури на икру и спермии // Вопр. ихтиологии. Т. 55. № 1. С. 95−100. https://doi.org/10.7868/S0042875215010087
Крылов В.В. 2018. Биологические эффекты геомагнитной активности: наблюдения, эксперименты и возможные механизмы // Тр. ИБВВ РАН. Т. 84(87). С. 7−38. https://doi.org/10.24411/0320-3557-2018-10016
Крылов В.В., Зотов О.Д., Клайн Б.И. 2011. Устройство для генерации магнитных полей и компенсации локального низкочастотного магнитного поля: А.с. RU 108 640 U1 (Россия). № 2011119410/28 // Изобретения, полезные модели. 2011. № 26.
Крылов В.В., Изюмов Ю.Г., Извеков Е.И., Непомнящих В.А. 2013. Магнитные поля и поведение рыб // Журн. общ. биологии. Т. 74. № 5. С. 354–365.
Кузьмина В.В., Ушакова Н.В., Крылов В.В. 2015. Влияние магнитных полей на активность протеиназ и гликозидаз кишечника карася Carassius carassius // Изв. РАН. Сер. биол. № 1. С. 70–76. https://doi.org/10.7868/S0002332915010087
Ланге Н.О., Дмитриева Е.Н. 1981. Методика эколого-морфологических исследований развития молоди рыб // Исследование размножения и развития рыб. М.: Наука. С. 67−88.
Леднев В.В., Сребницкая Л.К., Ильясова Е.Н. и др. 1996. Магнитный параметрический резонанс в биосистемах: экспериментальная проверка предсказаний теории с использованием регенерирующих планарий Dugesia tigrina в качестве тест-системы // Биофизика. Т. 41. № 4. С. 815–825.
Павлов Д.А. 2007. Морфологическая изменчивость в раннем онтогенезе костистых рыб. М.: ГЕОС, 246 с.
Таликина М.Г., Крылов В.В., Изюмов Ю.Г., Чеботарева Ю.В. 2013. Влияние типичной магнитной бури на митоз зародышевых клеток и размерно-массовые показатели предличинок плотвы (Rutilus rutilus L.) // Биология внутр. вод. № 1. С. 56−60. https://doi.org/10.7868/S032096521203014X
Чеботарева Ю.В., Изюмов Ю.Г., Крылов В.В. 2009. Влияние переменного электромагнитного поля на раннее развитие плотвы Rutilus rutilus (L.) (Cyprinidae, Cypriniformes) // Вопр. ихтиологии. Т. 49. № 3. С. 422−428.
Яковлев В.Н., Изюмов Ю.Г., Касьянов А.Н. 1981. Фенетический метод исследований популяций карповых рыб // Биол. науки. № 2. С. 98−101.
Bliss V.L., Heppner F.H. 1976. Circadian activity rhythm influenced by near zero magnetic field // Nature. V. 261. P. 411–412. https://doi.org/10.1038/261411a0
Brown F.A., Chow C.S. 1976. Uniform daily rotation and biological rhythms and clocks in hamsters // Physiol. Zool. V. 49. № 3. P. 263–285.
Golovanova I.L., Filippov A.A., Chebotareva Y.V. et al. 2017. Delayed effect of geomagnetic storm simulation on size, mass and activity of digestive glycosidases in roach (Rutilus rutilus Linnaeus 1758) underyearlings // J. Appl. Ichthyol. V. 33. № 2. P. 291–299. https://doi.org/10.1111/jai.13290
Krylov V.V., Chebotareva Y.V., Izyumov Y.G. 2016. Delayed consequences of extremely low-frequency magnetic fields and the influence of adverse environmental conditions on roach Rutilus rutilus embryos // J. Fish Biol. V. 88. № 4. P. 1283–1300. https://doi.org/10.1111/jfb.12869
Krylov V.V., Chebotareva Y.V., Izyumov Y.G. 2019a. Delayed consequences of the influence of simulated geomagnetic storms on roach Rutilus rutilus embryos // Ibid. V. 95. № 6. P. 1422–1429. https://doi.org/10.1111/jfb.14150
Krylov V.V., Chebotareva Y.V., Izyumov Y.G. 2019b. The sizes of roach Rutilus rutilus L. underyearlings from different ponds at the “Sunoga” pond station from 2003 to 2015 // Trans. IBIW RAS. V. 87 (90). P. 7–11. https://doi.org/10.24411/0320-3557-2019-10014
Li Y., Ru B., Miao W. et al. 2015. Effects of extremely low frequency alternating-current magnetic fields on the growth performance and digestive enzyme activity of tilapia Oreochromis niloticus // Environ. Biol. Fish. V. 98. P. 337–343. https://doi.org/10.1007/s10641-014-0263-6
Loewe C.A., Prolss G.W. 1997. Classification and mean behavior of magnetic storms // J. Geophys. Res. Space Phys. V. 102. P. 14209–14213. https://doi.org/10.1029/96JA04020
Welker H.A., Semm P., Willig R.P. et al. 1983. Effects of an artificial magnetic field on serotonin-nacetyltransferase activity and melatonin content of the rat pineal gland // Exp. Brain Res. V. 50. P. 426–432. https://doi.org/10.1007/BF00239209
Yamazaki Y., Maute A. 2017. Sq and EEJ – a review on the daily variation of the geomagnetic field caused by ionospheric dynamo currents // Space Sci. Rev. V. 206. P. 299–405. https://doi.org/10.1007/s11214-016-0282-z
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии