

Вопросы ихтиологии, 2021, T. 61, № 4, стр. 455-463
Вектор разделения пищевых ресурсов в изменчивости особей генерализованной формы крупных африканских усачей комплекса Barbus (=Labeobarbus) intermedius в озёрах Тана, Лангано и Аваса, Эфиопия
1 Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
2 Институт биологии внутренних вод РАН – ИБВВ РАН
пос. Борок, Ярославская область, Россия
* E-mail: adissa@mail.ru
Поступила в редакцию 11.06.2020
После доработки 17.11.2020
Принята к публикации 26.11.2020
Аннотация
Анализ диверсификации усачей генерализованной формы комплекса Barbus (=Labeobarbus) intermedius в мономорфных популяциях озёр Лангано и Аваса и в полиморфной популяции озера Тана позволяет выявить параметры изменчивости, направленной на разделение пищевых ресурсов. Большое сходство выявленных параметров в трёх изучаемых водоемах показывает, что в мономорфных популяциях структура изменчивости, направленной на разделение пищевых ресурсов, практически совпадает с таковой в полиморфной популяции. Вводится понятие “вектора разделения пищевых ресурсов”. В полиморфной популяции этот вектор вносит основной вклад в диверсификацию особей, в мономорфных популяциях его значение второстепенно. Сделан вывод о том, что разный эволюционный потенциал полиморфной и мономорфных популяций определяется удельным весом диверсификации, обусловленной вектором разделения пищевых ресурсов. Предлагаемый подход полагается перспективным в анализе изменчивости особей не только изучаемой, но и других систематических групп.
В системе фенетического разнообразия крупных африканских усачей комплекса Barbus intermedius (sensu Banister, 1973) генерализованными именуются особи без выраженных признаков специализации питания (Nagelkerke et al., 1994; Mina et al., 1996a, 1996b; Лёвин, 2003; Голубцов, 2010; Levin et al., 2019). Такие особи есть в каждом водоёме, где обитают представители комплекса, тогда как практически все формы, специализированные в отношении питания, известны лишь из тех локальностей, где они были описаны. Неизменность присутствия в любой популяции ареала даёт основания рассматривать генерализованных особей как близких (или тождественных) особям предковой формы, в процессе своей диверсификации давшей начало всему многообразию комплекса B. intermedius. В пользу гипотезы о такой близости свидетельствуют результаты многочисленных исследований (Nagelkerke et al., 1994; Mina et al., 1996a, 1996b; Sibbing et al., 1998; Sibbing, Nagelkerke, 2000; Levin et al., 2019), в частности, анализ изменчивости морфологических признаков генерализованных особей разных популяций в сравнении с изменчивостью особей специализированных морфотипов и аллопатрических видов усачей (Мироновский, 2020). Как оказалось, векторы диверсификации генерализованной формы, обитающей в оз. Тана, весьма сходны как с одноранговыми векторами симпатричных ей танских морфотипов, так и с соответствующими векторами аллопатрических видов крупных африканских усачей. Высокая степень этого сходства даёт основания полагать, что диверсификация особей генерализованной формы из оз. Тана, диверсификация танских морфотипов и диверсификация аллопатрических видов – это последовательные этапы одного процесса, и диверсификация морфотипов и видов идёт в направлениях, заданных диверсификацией особей генерализованной формы. Это, очевидно, является веским свидетельством в пользу гипотезы о близости генерализованной формы к предковой. Вместе с тем сравнение пар одноранговых векторов диверсификации мономорфных популяций генерализованных особей озёр Лангано и Аваса выявило их сходство между собой и существенные отличия от соответствующих векторов генерализованных особей в оз. Тана. Это даёт основания полагать, что в разных локальностях диверсификация усачей генерализованной формы может идти по-разному.
Цель настоящего исследования – поиск ответа на вопрос: в чём заключается та разница диверсификации генерализованных особей в полиморфной популяции оз. Тана и в мономорфных популяциях озёр Аваса и Лангано, которая обусловливает различия их эволюционного потенциала?
МАТЕРИАЛ И МЕТОДИКА
Материал для исследования собран в трёх озёрах Эфиопии: Тана в 1992–2010 гг., Аваса в 1997−2016 гг. и Лангано в 2007−2011 гг. (рис. 1). Рыб отлавливали жаберными и накидными сетями, брали из уловов местных рыбаков. В оз. Тана обитает полиморфная популяция крупных африканских усачей; генерализированная форма (далее GF11) этой популяции описана как морфотип intermedius22 (Nagelkerke et al., 1994; Мина и др., 2016). В сравнении с разнообразием форм усачей в оз. Тана популяции озёр Аваса и Лангано можно рассматривать как практически мономорфные. Слабо выраженный полиморфизм по пропорциям черепа, обнаруженный у особей этих озёр, даёт основания предполагать лишь едва наметившуюся дивергенцию на две формы − эврифагов и ихтиофагов − и не сопряжён с различиями по внешним признакам (Мина и др., 2016). Численность выборки GF оз. Тана составила 119 особей стандартной длиной (SL) 17.0–28.7 см, оз. Аваса – 49 экз. SL 17.9–28.3 см, Лангано − 73 экз. SL 12.4–34.7 см.
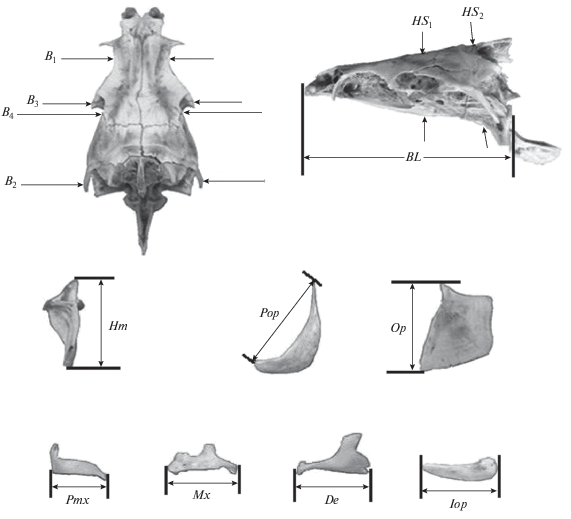
У каждой особи сделано по 14 промеров черепа (рис. 2). Как показало изучение изменчивости крупных африканских усачей, данный набор признаков, обладая высокой разрешающей способностью и хорошей воспроизводимостью результатов измерений как одним, так и разными операторами, позволяет достаточно уверенно оценивать фенетические отношения большинства морфотипов (Mina et al., 1993, 1996а, 1996b, 1998; Мироновский, 2006).
Рис. 2.
Схема промеров черепа рыб комплекса Barbus intermedius: BL − базальная длина черепа; B1, B2, B3 – расстояние между внешними краями соответственно frontalia, pterotica и sphenotica; B4 – ширина черепа на уровне соединения frontale и pteroticum, HS1 и HS2 – высота черепа на уровне соответственно изгиба parasphenoideum и заднего края parasphenoideum, Hm – высота hyomandibulare, Pop − длина praeoperculum, Op – высота передней части operculum; Pmx, Mx, De и Iop – длина соответственно praemaxillare, maxillare, dentale и interoperculum.
Статистическая обработка данных выполнена средствами пакетов NTSYS 2.02k (Rohlf, 1998) и Статистика 6. В расчётах использованы индексы − отношения абсолютных значений промеров к базальной длине черепа (BL). Далее при упоминании того или иного признака имеется в виду его индекс, а не сам промер. Значения индексов преобразовывали в натуральные логарифмы для нормализации распределений.
В анализе главных компонент (АГК) собственные векторы считали по корреляционной матрице; длину вектора принимали равной 1. В отличие от работ, в которых АГК используется для уменьшения числа переменных с последующим анализом распределения точек-объектов в двух- или трёхмерном пространстве первых ГК, настоящее исследование основано на анализе сходства и различия собственных векторов (СВ) корреляционных матриц, характеризующих главные направления дисперсии объектов рассматриваемых множеств (Андреев, Решетников, 1978; Андреев, 1980; Дгебуадзе и др., 2008; Мироновский, 2020). Главные направления дисперсии рассматриваются как основные направления изменчивости (диверсификации, дивергенции) усачей в изучаемых водоёмах. Для оценки сходства нагрузок признаков на собственные векторы использовали коэффициент ранговой корреляции Спирмена (rS), достоверность rS оценивали средствами пакета Статистика 6. Как показано в предыдущем исследовании (Мироновский, 2020), упорядоченная конструктивная изменчивость (сигнал), обусловливающая дивергенцию морфотипов и видов крупных африканских усачей, сосредоточена в двух первых собственных векторах – СВ1 и СВ2; далее (в СВ3, СВ4, СВ5 …) преобладает шум, т. е. изменчивость стохастическая, случайная (Kaiser, 1960). Исходя из этого, анализ в настоящем исследовании ограничен СВ1 и СВ2.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
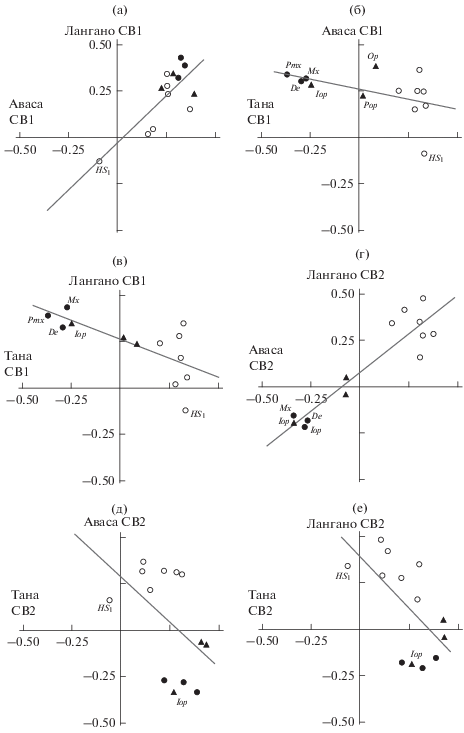
Чтобы сравнить дисперсию особей в изучаемых совокупностях, отложим на осях системы прямоугольных координат факторные нагрузки рассматриваемых переменных на СВ сравниваемых совокупностей. В гипотетическом случае, когда направления изменчивости в паре сопоставляемых совокупностей абсолютно одинаковы (т.е. диверсификационные процессы полностью совпадают), точки, соответствующие нагрузкам признаков, лягут на прямую, расположенную под углом 45° к осям координат, корреляция между значениями нагрузок будет равна 1. Точки на таком графике тем дальше расположены от начала координат, чем бóльшую нагрузку в данном направлении изменчивости имеет данный признак. У начала координат окажутся точки, соответствующие переменным с наименьшими нагрузками в данном направлении изменчивости. Графики, отражающие результаты однорангового сравнения двух первых СВ дисперсии особей в каждой из трёх анализируемых совокупностей усачей (таблица) приведены на рис. 3.
Рис. 3.
Сравнение одноранговых собственных векторов − СВ1 (а−в) и СВ2 (г−е) − дисперсии особей генерализованной формы (GF) комплекса Barbus intermedius из озёр Тана, Аваса и Лангано: а − Аваса−Лангано, rS = 0.472, р = 0.103; б – Тана−Аваса, rS = –0.566, р = 0.043; в − Тана−Лангано, rS = –0.664, р = 0.013; г – Аваса−Лангано, rS = 0.758, р = 0.003; д – Тана−Аваса, rS = –0.505, р = 0.078; е – Тана−Лангано, rS = –0.566, р = 0.044. Нагрузки параметров: (⚫) − челюстной дуги: Pmx, Mx, De; (▲) − жаберной крышки: Pop, Op, Iop; (⚪) − нейрокраниума: B1−B4, HS1, HS2 и Hm. (Чтобы не загромождать рисунки, параметры обозначены символами и надписями только в той мере, в какой это необходимо для понимания изложения.)
Собственные векторы (СВ1, СВ2): собственные значения и нагрузки рассматриваемых признаков в популяциях генерализованных усачей комплекса Barbus intermedius озёр Тана, Аваса и Лангано
| Признак | Тана | Аваса | Лангано | |||
|---|---|---|---|---|---|---|
| СВ1 | СВ2 | СВ1 | СВ2 | СВ1 | СВ2 | |
| B3 | 0.300 | 0.113 | 0.255 | 0.377 | 0.282 | 0.292 |
| B2 | 0.203 | 0.292 | 0.257 | 0.308 | 0.240 | 0.163 |
| B1 | 0.320 | 0.213 | 0.250 | 0.323 | 0.349 | 0.280 |
| B4 | 0.309 | 0.144 | 0.369 | 0.224 | 0.160 | 0.422 |
| HS1 | 0.332 | –0.063 | –0.094 | 0.167 | –0.126 | 0.347 |
| HS2 | 0.340 | 0.107 | 0.179 | 0.321 | 0.049 | 0.495 |
| Hm | 0.283 | 0.306 | 0.153 | 0.305 | 0.004 | 0.354 |
| Pmx | –0.369 | 0.317 | 0.344 | –0.276 | 0.392 | –0.211 |
| Pop | 0.011 | 0.422 | 0.231 | –0.068 | 0.266 | 0.055 |
| Op | 0.086 | 0.429 | 0.390 | –0.071 | 0.237 | –0.042 |
| Iop | –0.246 | 0.264 | 0.288 | –0.335 | 0.345 | –0.188 |
| Mx | –0.271 | 0.385 | 0.322 | –0.338 | 0.437 | –0.155 |
| De | –0.296 | 0.213 | 0.308 | –0.270 | 0.326 | –0.178 |
| Собственные значения | 3.700 | 2.580 | 4.490 | 2.370 | 4.150 | 2.160 |
| Объяснённая дисперсия % | 28.5 | 19.9 | 34.5 | 18.3 | 31.9 | 16.6 |
Примечание. B1, B2, B3 – расстояние между внешними краями соответственно frontalia, pterotica и sphenotica; B4 – ширина черепа на уровне соединения frontale и pteroticum; HS1, HS2 – высота черепа на уровне изгиба parasphenoideum и заднего края parasphenoideum, Hm – высота hyomandibulare; Pmx, Pop − длина praemaxillare и praeoperculum, Op – высота передней части operculum; Iop, Mx, De – длина соответственно interoperculum, maxillare и dentale.
В распределении нагрузок признаков на СВ1 общим для мономорфных популяций озёр Аваса и Лангано является знаковая (“−” и “+”) оппозиция HS1 и 12 других параметров: нагрузки HS1, почти совпадая по величине в обеих выборках, расположены в третьей четверти координатной плоскости, нагрузки других 12 параметров – в первой (рис. 3а). В популяции оз. Тана противостояния “HS1 против всех” нет, но есть другая оппозиция знаков: нагрузки параметров морфологии челюстей (De, Pmx и Mx) и одного из параметров жаберной крышки (Iop) расположены в третьей четверти координатной плоскости (“−”), а нагрузки других девяти признаков – в первой (“+”) (рис. 3б, 3в). Два параметра жаберной крышки (Pop и Op) находятся у начала координат, из чего следует, что их вес в дисперсии по СВ1 сравнительно невелик и основной вклад в изменчивость в этом направлении в популяции оз. Тана вносит противостояние групп De, Pmx, Mx, Iop (“−”) и B1−B4, HS1, HS2, Hm (“+”). Разница оппозиций знаков и состава противостоящих групп делает распределение нагрузок параметров на СВ1 в полиморфной и мономорфных популяциях несходным, а значимость корреляции между ними свидетельствует о закономерности этого несходства.
О сходстве дисперсии генерализованных усачей в популяциях озёр Аваса и Лангано по СВ2 свидетельствует высокая статистически значимая корреляция между соответствующими СВ2 (рис. 3г). Первое, что обращает на себя внимание при анализе соответствующих распределений, это отчётливое противостояние двух групп признаков, разных по знаку, но примерно одинаковых по абсолютной величине нагрузок на вектор. Первая группа – это De, Pmx, Mx и Iop (“−”), вторая – B1−B4, HS1, HS2 и Hm (“+”). Высокие значения нагрузок свидетельствуют о том, что основной вклад в дисперсию особей, определяемую СВ2, вносят именно эти противостоящие одна другой группы. Два признака жаберной крышки (Pop и Op) ни в одну из групп явно не входят, они расположены у начала координат, их нагрузки на векторы близки к 0, т. е. их вклад в дисперсию объектов изучаемых совокупностей в данном направлении изменчивости сравнительно невелик.
Как видим, распределение нагрузок признаков на СВ2 в мономорфных популяциях (рис. 3г) отчётливо напоминает распределение нагрузок признаков на СВ1 в полиморфной популяции оз. Тана (рис. 3б, 3в). Тогда как в распределении нагрузок признаков на СВ2 в выборке из оз. Тана (рис. 3д, 3е) мы видим оппозицию HS1 и 12 других параметров, наблюдавшуюся ранее (рис. 3а). В силу этого анализ данных рис. 3 даёт очевидные основания полагать, что направление диверсификации особей, которое в мономорфных популяциях соответствует СВ1 корреляционной матрицы (рис. 3а), в популяции полиморфной соответствует СВ2 (рис. 3д, 3е). И наоборот: направление диверсификации особей, соответствующее СВ1 корреляционной матрицы в полиморфной популяции (рис. 3б, 3в), соответствует СВ2 в популяциях мономорфных (рис. 3г).
Более наглядно это предположение иллюстрирует рис. 4, где отношения мономорфных и полиморфной популяций анализируются путём сравнения векторов не одного, а разных рангов. В сравнении СВ1 полиморфной популяции оз. Тана с СВ2 каждой из мономорфных популяций (рис. 4а, 4б) нагрузки признаков весьма сходны как между собой, так и с нагрузками признаков в сравнении СВ2 двух мономорфных популяций (рис 3г). Корреляция между СВ в трёх означенных сравнениях высокая и статистически значимая, что подтверждает закономерность их сходства. Сходны и графики, отражающие распределения нагрузок в сравнении СВ2 полиморфной популяции с СВ1 мономорфных популяций (рис. 4в, 4г), а также с графиком распределения нагрузок в сравнении СВ1 мономорфных популяций между собой (рис. 3а). В этих трёх сравнениях корреляция между СВ статистически не значима, что не позволяет рассматривать подобие векторов как достоверную закономерность. Вместе с тем разительное сходство паттернов распределения в трёх означенных сопоставлениях едва ли можно рассматривать как дело слепого случая. На каждом из трёх графиков (рис. 3а, 4в, 4г) признаки распределены только в первой и в третьей координатных плоскостях, в первой четверти координат сосредоточены 12 из 13 рассматриваемых параметров, в третьей четверти мы неизменно видим символ HS1 (высота осевого черепа на уровне изгиба parasphenoideum). Очевидно, разделение 13 признаков в соотношении 12 : 1 так, чтобы в трёх независимых наблюдениях (в популяциях трёх озёр) параметром, противостоящим прочим по знаку нагрузки на вектор, оказывался один и тот же параметр HS1, свидетельствует если не о закономерности, то об отчётливо выраженной тенденции, безусловно заслуживающей внимания и дальнейшего изучения.
Рис. 4.
Сравнение разноранговых собственных векторов (СВ1−СВ2) дисперсии особей GF комплекса Barbus intermedius: а – оз. Тана СВ1−оз. Аваса СВ2, rS = 0.786, р = 0.001; б – оз. Тана СВ1−оз. Лангано СВ2, rS = 0.907, р = 0.001; в – оз. Тана СВ2−оз. Аваса СВ1, rS = 0.428, р = 0.143; г – оз. Тана СВ2−оз. Лангано СВ1, rS = 0.379, р = 0.201; обозначения см. на рис. 3.
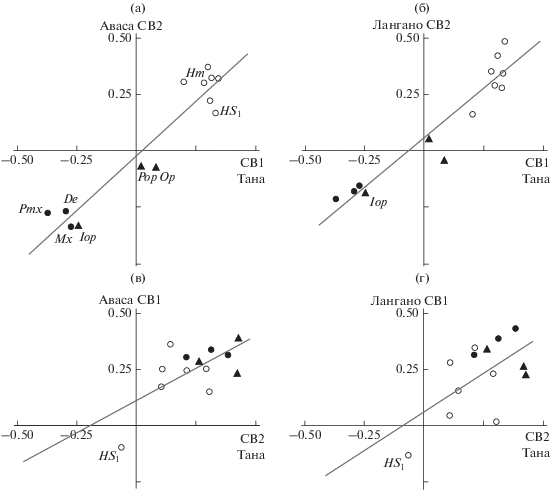
Структура распределений признаков на рис. 3г, 4а и 4б почти совпадает со структурой корреляционных плеяд, выявленных при оценке корреляций этих же 13 признаков в процессе диверсификации усачей комплекса B. intermedius (Мироновский, 2020). По краям распределений разнесены группа из De, Pmx, Mx, Iop и группа из B1−B4, HS1, HS2,Hm; по абсолютной величине нагрузки признаков двух данных групп на векторы примерно равны. Почти на равном расстоянии от этих групп у начала координат расположена пара Pop и Op. Подобно этому, на дендрограммах корреляций, выявивших плеяды, один кластер составили De, Pmx, Mx и Iop, второй – B1−B4, HS1, HS2,Hm (Мироновский, 2020). Признаки Pop и Oр, формально входя в кластер с B1−B4, HS1, HS2,Hm, занимают в нём крайнее положение, а анализ матриц корреляций, по которым стоились дендрограммы, показывает, что эта пара признаков примерно равноудалена от обеих групп и попадание в один из кластеров определили вторые знаки после запятой. Сделан вывод о том, что оппозиция выявленных плеяд, выражающаяся в значимой отрицательной корреляции между ними, присуща диверсификации особей в процессе разделения пищевых ресурсов (Мироновский, 2020). Аналогичная оппозиция по знаку нагрузок на СВ (“+” или “−”), высокое сходство паттерна распределений и тесные статистически значимые корреляции на рис. 3г, 4а и 4б позволяют рассматривать сопоставляемые СВ как разные реализации вектора разделения пищевых ресурсов в разных популяциях генерализованных особей. Именно разные реализации одного и того же вектора, общего для особей трёх изучаемых популяций, а не разные векторы в полиморфной и в мономорфных популяциях. В пользу такого предположения свидетельствует и то, что при сравнении СВ полиморфной и мономорфных популяций (рис. 4а, 4б) rS выше, чем при сравнении СВ мономорфных популяций между собой (рис. 3г).
Подобие СВ разного ранга, выявленное в настоящем исследовании, позволяет развить одно из положений предыдущего исследования изменчивости изучаемой группы рыб (Мироновский, 2020). Результаты однорангового сравнения СВ1 и СВ2 трактовались тогда как свидетельство существования четырёх главных векторов диверсификации крупных африканских усачей. Как представлялось, два из них определяют диверсификацию генерализованных усачей в оз. Тана. Другие два – диверсификацию усачей мономорфных популяций в озёрах Аваса и Лангано. Результаты настоящего исследования дают основания полагать, что главных направлений изменчивости не четыре, а два. Первое определяет отчётливо выраженная оппозиция двух групп признаков; основа одной из групп – это параметры челюстной дуги (рис. 3г, 4а, 4б). Это предполагает прямую связь диверсификации с разделением пищевых ресурсов (далее это направление изменчивости будет обозначено как рпр-вектор, т.е. вектор разделения пищевых ресурсов). Значение оппозиции “HS1 против 12 прочих признаков” неочевидно, неочевидны и факторы, определяющие второе направление изменчивости (поэтому далее второй вектор диверсификации будем именовать х-вектором). Обратившись к рис. 3а, 4в и 4г, можно убедиться, что нагрузки параметров челюстной дуги высоки и здесь, давая основания полагать фактор питания весомым и во втором направлении изменчивости, хотя здесь этот фактор явно не единственный и, видимо, не главный.
В случае двух (а не четырёх) главных векторов изменчивости разница диверсификации генерализованных усачей разных совокупностей определяется тем, какой из них в данной совокупности соответствует СВ1, а какой – СВ2. В популяции оз. Тана позицию СВ1 занимает рпр-вектор (рис. 4а, 4б), тогда как позиция СВ2 – это позиция x-вектора (рис. 4в, 4г). В популяциях озёр Лангано и Аваса наоборот: позицию СВ1 занимает x-вектор (рис. 3а, 4в, 4г), а СВ2 – это позиция рпр-вектора (рис. 3г, 4а, 4б). Таким образом собственный вектор любого из двух рассматриваемых рангов (СВ1 и СВ2) в одних случаях может соответствовать диверсификации, направленной на разделение пищевых ресурсов (рпр-вектор), в других случаях – диверсификации под влиянием x-вектора неясной на данном этапе исследований направленности. Обратившись к таблице, можно убедиться, что в оз. Тана, где рпр-вектор занимает позицию СВ1, на его долю приходится 28.5% общей дисперсии, в озёрах Аваса и Лангано, где позиция рпр-вектора – это СВ2, доля определяемой им изменчивости равна соответственно 18.3 и 16.6%. Как показывает настоящее исследование, именно эта разница долей изменчивости, направленной на разделение пищевых ресурсов, видимо, и определяет разницу интенсивности трофической радиации генерализованных особей в этих водоёмах.
Интересно, что на рисунках, где сравниваются векторы одной и той же пары популяций – Лангано и Аваса (рис. 3а, 3г), Тана и Аваса (рис. 4а, 4в), Тана и Лангано (рис. 4б, 4г), корреляции между парами рпр-векторов (рис. 3г, 4а, 4б) высокие и статистически значимые, тогда как корреляции между парами x-векторов (рис. 3а, 4в, 4г) недостоверны. Из этого следует, что в межпопуляционных сравнениях сходство диверсификации особей в процессе разделения пищевых ресурсов больше, чем сходство их диверсификации по сумме факторов неочевидной направленности (или наоборот: межпопуляционные различия в диверсификации в процессе разделения пищевых ресурсов ниже межпопуляционных различий диверсификации по факторам неочевидной направленности). Другими словами, диверсификация особей в отношении разделения пищевых ресурсов в полиморфной и в мономорфных популяциях генерализованных усачей идёт сходным образом и эволюционный потенциал популяции определяется рангом (СВ1 или СВ2) рпр-вектора, определяющего этот процесс, т.е. долей изменчивости, направленной на разделение пищевых ресурсов. В пользу предположения о том, что сходство диверсификации особей в процессе разделения пищевых ресурсов не зависит от типа сравниваемых популяций, свидетельствует и то, что на рис. 4а и 4б, где сравниваются рпр-векторы мономорфных и полиморфной популяций, корреляции выше, чем на рис. 3г, где между собой сравниваются рпр-векторы двух мономорфных популяций.
В заключение отметим, что результаты настоящего исследования хорошо соответствуют представлениям Блэкита (1968) о предпочтительных направлениях и биологически значимых векторах изменчивости.
ВЫВОДЫ
1. Упорядоченная конструктивная изменчивость генерализованных усачей комплекса B. intermedius определяется двумя векторами, общими для мономорфных популяций озёр Аваса и Лангано и полиморфной популяции оз. Тана.
2. Один из выявленных векторов соответствует диверсификации особей в процессе разделения пищевых ресурсов.
3. Роль второго вектора изменчивости не очевидна и требует дальнейшего изучения.
4. В диверсификацию генерализованных особей в оз. Тана основной вклад вносит вектор разделения пищевых ресурсов.
5. В диверсификации генерализованных особей в популяциях озёр Лангано и Аваса вклад вектора разделения пищевых ресурсов второстепенен.
6. Интенсивность формообразования, т.е. эволюционный потенциал популяций трёх изучаемых озёр, определяется той долей изменчивости, какую вектор разделения пищевых ресурсов вносит в общую изменчивость генерализованных особей в каждом исследованном водоёме.
Список литературы
Андреев В.Л., Решетников Ю.С. 1978. Анализ фенотипической изменчивости географически удалённых популяций одного вида // Математические методы в экологии и географии. Владивосток: Изд-во ДВНЦ АН СССР. С. 98−110.
Блэкит Р.Э. 1968. Морфометрический анализ // Теоретическая и математическая биология. М.: Мир. С. 247–273.
Голубцов А.С. 2010. “Пучки видов” рыб в реках и озерах: симпатрическая дивергенция в фаунистически обеднённых рыбных сообществах как особый модус эволюции // Актуальные проблемы современной ихтиологии. К 100-летию Г.В. Никольского. М.: Т-во науч. изд. КМК. С. 96–123.
Дгебуадзе Ю.Ю., Мина М.В., Мироновский А.Н. 2008. К оценке фенетических отношений алтайских османов (Oreoleuciscus, Cyprinidae) из трех озер Монголии по признакам черепа // Вопр. ихтиологии. Т. 48. №. 3. С. 315−323.
Лёвин Б.А. 2003. Структура локальных популяционных систем у рыб рода Barbus // Матер. ІІ Міжнар. наук. конф. “Біорізноманіття та роль зооценозу в природних і антропогенних екосистемах”. Днепропетровск: Изд-во ДНУ. С. 53−55.
Мина М.В., Мироновский А.Н., Дгебуадзе Ю.Ю. 2016. Полиморфизм по пропорциям черепа у крупных африканских усачей Barbus intermedius sensu Banister, 1973 (Cyprinidae) из озер Аваса и Лангано (Рифтовая долина, Эфиопия) // Вопр. ихтиологии. Т. 56. № 4. С. 403−409. https://doi.org/10.7868/S0042875216040081
Мироновский А.Н. 2006. Факторы, обуславливающие сопоставимость данных, полученных путем оценки пластических признаков рыб // Там же. Т. 46. № 2. С. 240−251.
Мироновский А.Н. 2020. Крупные африканские усачи: векторы диверсификации особей генерализованной формы как основа морфологического разнообразия комплекса Barbus intermedius в оз. Тана, Эфиопия // Там же. Т. 60. № 3. С. 283−294. https://doi.org/10.31857/S0042875220030121
Banister K.E. 1973. A revision of the large Barbus (Pisces, Cyprinidae) of East and Central Africa. Pt. II // Bull. Brit. Mus. Nat. Hist. Zool. V. 26. P. 1–148.
Kaiser H.F. 1960. The application of electronic computers to factor analysis // Educ. Psychol. Measmt. V. 20. № 1. P. 141−151.
Levin B.A., Casal-López M., Simonov E. et al. 2019. Adaptive radiation of barbs of the genus Labeobarbus (Cyprinidae) in the East African river // Freshwater Biol. V. 64. № 10. P. 1721−1736.
https://doi.org/10.1111/fwb.13364
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. 1993. Alometria et divergence entre les barbeaux du lac Tana (Ethiopie) // Cahiers d’Ethologie. V. 13. P. 219–222.
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. 1996a. Morphometry of barbel of Lake Tana, Ethiopia: multivariate ontogenetic channels // Folia Zool. V. 45. Suppl. 1. P. 109–116.
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. 1996b. Lake Tana large barbs: phenetics, growth and diversification // J. Fish Biol. V. 48. P. 383–404.
Mina M.V., Mironovsky A.N., Golubtsov A.S., Dgebuadze Yu.Yu. 1998. ‘Barbus’ intermedius species flock in Lake Tana (Ethiopia). II. Morphological diversity of “large barbs” from Lake Tana and neighbouring areas: homoplasies or synapomorphies? // Ital. J. Zool. V. 65. Suppl. P. 9–14.
Nagelkerke L.A.J., Sibbing F.A., van den Boogaart J.G.M. et al. 1994. The barbs (Barbus spp.) of Lake Tana: a forgotten species flock? // Environ. Biol. Fish. V. 39. P. 1–21.
Rohlf F.J. 1998. NTSYS-pc: numerical taxonomy and multivariate analysis system. Version 2.0. N.Y.: Exeter Software, 31 p.
Sibbing F.A., Nagelkerke L.A.J. 2000. Resource partitioning by Lake Tana barbs predicted from fish morphometrics and prey characteristics // Rev. Fish Biol. Fish. V. 10. № 4. P. 393–437.
Sibbing F.A., Nagelkerke L.A.J., Stet R.J.M., Osse J.W.M. 1998. Speciation of endemic Lake Tana barbs (Cyprinidae, Ethiopia) driven by trophic resource partitioning; a molecular and ecomorphological approach // Aquat. Ecol. V. 32. P. 217–227.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии