

Вопросы ихтиологии, 2021, T. 61, № 4, стр. 394-399
Молодь Amphelikturus dendriticus (Syngnathidae) и специфика онтогенетической изменчивости вида
1 Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
2 Институт океанологии РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 15.10.2020
После доработки 05.11.2020
Принята к публикации 05.11.2020
Аннотация
Описаны мальки Amphelikturus dendriticus SL 25 и 30 мм из Карибского моря. Выявлено, что на стадии “планктонного малька” у этого вида отсутствует пластинчатый шельф вентрального костного края орбиты, слабее развит медиальный рыльный гребень, лучше выражена шиповатость окончаний основных гребней на туловищных поясках и повсеместно присутствует меланофорная пигментация. Переход к дефинитивному строению осуществляется у данного вида по достижении SL примерно 30 мм, но некоторые ювенильные черты (отсутствие латеральных рыльных гребней, полный оперкулярный гребень) сохраняются и при более крупных размерах тела. Степень развития и ветвления мочек адаптивна и не всегда чётко коррелирует с ростом.
Игловые и морские коньки (Syngnathidae) включают рыб причудливого облика, преимущественно связанных с морской растительностью и населяющих прибрежные воды на малых глубинах, но встречающихся и в пелагиали открытого океана, и на глубинах до 400 м (Dawson, 1985). В отличие от большинства других костистых рыб молодь игловых уже на самых ранних стадиях развития мало отличается от взрослых рыб, что позволяет легко идентифицировать даже недавно вылупившихся мальков. Вместе с тем у молоди некоторых видов отмечены настолько специфические отличия от взрослых особей, что такие рыбы описывались в качестве самостоятельных видов (Dawson et al., 1979). Наиболее характерным отличием планктонных мальков игловых от взрослых рыб является шиповатость окончаний гребней на поясках, лучшая выраженность самих гребней и зачастую зазубренность их свободного края. Подобные различия описаны у представителей родов Corythoichthys Kaup, 1853 (Dawson, 1977), Oostethus Hubbs, 1929 (Dawson et al., 1979; Frias-Torres, 2006) и Syngnathus Linnaeus, 1758 (Dawson et al., 1979; Fritzsche, 1980). Для мальков Corythoichthys описано персистирование дорсальной и вентральной кожных складок в хвостовом отделе (Dawson, 1977); молодь Oostethus aculeatus (Kaup, 1856) выделяется исключительно крупным хвостовым плавником, из-за чего она была описана в качестве самостоятельного вида (Poll, 1953); у представителей родов, утрачивающих спинной и грудные плавники во взрослом состоянии, у молоди они вполне развиты (Dawson, Allen, 1978); наконец, для молоди некоторых видов рода Trachyrhamphus Kaup, 1853 характерно присутствие нитевидных выростов на теле (Dawson et al., 1979; Dawson, 1985). Однако в целом ранние стадии развития Syngnathidae изучены ещё очень слабо и характер онтогенетических преобразований для большинства родов не описан.
Одним из малоизученных родов является Amphelikturus Parr, 1930, включающий карликовых Syngnathidae (максимальная известная SL 81 мм), имеющих облик “иглоконьков” (в англоязычной литературе “pipehorses”), морфологически промежуточных между типичными игловыми и морскими коньками. Род был установлен Парром (Parr, 1930) для вида A. brachyrhynchus Parr, 1930 от Багамских о-вов, который оказался синонимом ранее описанного вида Siphostoma dendriticum Barbour, 1905 из вод Бермуд (Barbour, 1905). Этот вид распространен в Западной Атлантике от Нью-Брунсвика (Канада) до Пернамбуко (Бразилия), но редок в коллекциях: Доусон (Dawson, 1982a) сообщил всего о 32 экземплярах в американских музеях. Хотя в настоящее время Amphelikturus является монотипическим родом, Доусон (Dawson, 1982a, 1982b) указал на существование нескольких неописанных видов в Восточной Атлантике и Индо-Пацифике. От других цепкохвостых родов Amphelikturus отличается не соединяющимися дорсальными туловищными и хвостовыми гребнями, отсутствием костных пластинок на склере глазного яблока и развитых шипов на поясках, а также наличием вполне развитого С с 9–10 лучами (Dawson, 1982a, 1985). Хотя в своих последних работах Доусон (Dawson, 1984, 1985) рассматривал Amphelikturus наряду с Idiotropiscis Whitley, 1947 в качестве подродов рода Acentronura Kaup, 1853, в настоящее время им придается статус самостоятельных родов (Fricke et al., 2020), что принято и в данной работе. Сведения об онтогенетической изменчивости A. dendriticus ограничиваются замечаниями Доусона (Dawson, 1982a) о степени развития мочек на теле в зависимости от размера рыб.
В коллекции Института океанологии РАН (ИО РАН, Москва) имеется два ранее не описанных малька A. dendriticus из Карибского моря, пойманных в 14-м рейсе научно-исследовательского судна (НИС) “Академик Курчатов” и в 20-м рейсе НИС “Дмитрий Менделеев”. Их отождествление с A. dendriticus основано на присутствии у них цепкого хвостового конца тела с хорошо развитым С, содержащим девять лучей; наличии вентрального наклона головы, не соединяющихся дорсальных туловищных и хвостовых гребней, 14 + 37–40 поясков и положении начала D на последнем туловищном пояске (последние два признака отличают A. dendriticus от неописанного вида из Восточной Атлантики: Dawson, 1982b). Несмотря на близкие абсолютные размеры, между ними выявлен ряд заметных структурных различий, которые для данного вида ранее не были известны и впервые описываются в настоящем сообщении. Эти различия могут отражать переход стадии “планктонного малька” в дефинитивную форму в процессе метаморфоза, который, таким образом, осуществляется у A. dendriticus при значениях SL, близких к 30 мм.
МАТЕРИАЛ И МЕТОДИКА
Изученные экземпляры Amphelikturus dendriticus имеют следующие этикеточные данные: SL 25.0 мм, 10°21′–10°23′ с. ш. 78°50′ з. д., планктонная сеть (ПС), поверхность, время лова 20.30–21.30, НИС “Дмитрий Менделеев”, рейс 20, ст. 1625, проба 29, 19.02.1978 г.; SL ~ 30 мм (хвостовой конец тела свернут кольцом), 19°56′ с. ш. 68°14′ з. д., ПС, свет, поверхность, время лова 21.20–21.40, НИС “Академик Курчатов”, рейс 14, ст. 1181, проба 16, 01.02.1973 г. Методика изучения и терминология соответствует работам Доусона (Dawson, 1982a, 1985), англоязычные термины адаптированы к русскому языку. В тексте использованы следующие сокращения: D, A, P, С – соответственно спинной, анальный, грудные и хвостовой плавники; SL – стандартная длина.
РЕЗУЛЬТАТЫ
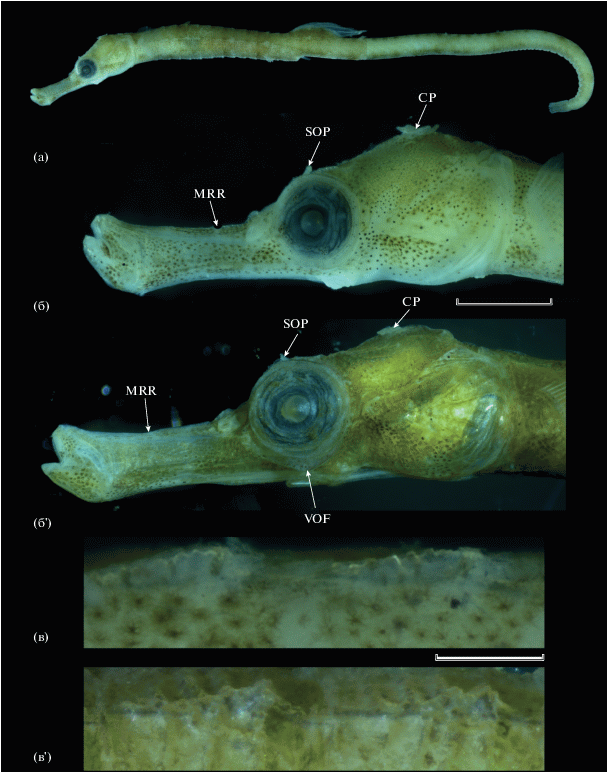
Малёк SL 25 мм. D 15, A 3, P 16, C 9. Пластические признаки приведены в таблице. Поясков 14 + 37 = 51, последние 17 хвостовых поясков образуют цепкий хвостовой конец тела (рисунок, а). Голова немного наклонена вниз относительно продольной оси тела, 5.3 раза в SL. Рыло 2.4 раза в длине головы, наиболее низкое у начала его медиального гребня, несколько увеличивается в высоту к вершине, максимальная высота рыла (на уровне нижнечелюстного сустава) 2.7 раза содержится в его длине и в 1.3 раза превышает высоту рыла на уровне начала медиального гребня (рисунок, б). Медиальный рыльный гребень начинается в 1.5 раза ближе к переднему краю глаза, чем к вершине рыла, образует лопастевидный выступ над ноздрями, латеральный – не выражен. Супраорбитальные гребни хорошо развиты, в том числе позади орбит. Вентральный костный край орбиты без пластинчатого расширения. Глазное яблоко с плоскими бородавочками и единичными короткими папиллами, без склеротизированных пластинок. Супрапреоперкулярный гребень отсутствует. Медиальный гребень головы образует нерезкий выступ. Operculum с изогнутым к дорсокаудальному углу кости гребнем, от которого вниз отходят многочисленные радиальные гребешки. Верхние туловищные гребни не соединяются с верхними хвостовыми; боковые туловищные гребни соединяются с нижними хвостовыми. Щиточки (scutella) развиты на туловищных и передних хвостовых поясках. Гребни туловищных поясков резкие, протяжённые (на всю длину пояска), с зазубренным свободным краем, оканчиваются отчётливыми шиповидными выступами (рисунок, в); на передних хвостовых поясках гребни и их выступы развиты гораздо слабее и постепенно сходят на нет в каудальном направлении. Основание D отчётливо поднято; начало D расположено на последнем (14-м) туловищном пояске, а окончание – на четвёртом хвостовом. Р закруглены, оканчиваются на уровне середины длины второго туловищного пояска. А и С хорошо развиты, дистальный конец С усечённый. На вершине медиального гребня головы сидит крупная (в расправленном состоянии равна по длине диаметру глаза) непарная ветвистая корональная мочка; над глазом имеется короткая простая супраорбитальная мочка. Короткие простые мочки имеются с каждой стороны у верхнего края жаберного отверстия. Парные короткие округлые ветвящиеся мочки имеются на дорсальной поверхности четвёртого и вентральной поверхности одиннадцатого туловищных поясков, пара коротких простых мочек – на вентральной поверхности восьмого туловищного пояска. Редко расположенные короткие простые парные мочки имеются на дорсальной или вентральной поверхностях отдельных хвостовых поясков.
Amphelikturus dendriticus, SL 25 (а–в) и ~30 мм (б', в'): а – общий вид; б, б' – голова сбоку; в, в' – верхние гребни пятого и шестого туловищных поясков, вид сбоку. CP – корональная мочка, MRR – положение начала медиального рыльного гребня, SOP – надглазничная мочка, VOF – пластинчатое расширение вентрального костного края орбиты. Масштаб, мм: б, б' – 1; в, в' – 0.25.
Морфометрические признаки изученных особей Amphelikturus dendriticus
| Признак | SL 25.0 мм | SL ~ 30 мм |
|---|---|---|
| В % SL | ||
| Длина головы (lc) | 19.0 | 20.8 |
| Длина рыла | 8.0 | 8.3 |
| Максимальная высота рыла | 3.0 | 3.3 |
| Диаметр глаза | 3.0 | 4.2 |
| Длина основания D | 8.0 | 8.3 |
| Длина Р | 4.0 | 4.2 |
| Длина С | 3.0 | 3.3 |
| Максимальная высота тела | 6.0 | 5.8 |
| Высота тела на уровне середины основания D | 4.0 | 5.0 |
| Предорсальное расстояние | 42.0 | 46.7 |
| Преанальное расстояние | 44.0 | 48.3 |
| В % lc | ||
| Длина рыла | 42.1 | 40.0 |
| Максимальная высота рыла | 15.8 | 16.0 |
| Диаметр глаза | 15.8 | 20.0 |
| Длина Р | 21.1 | 20.0 |
Окраска фиксированного экземпляра светлая, с густо и несколько неравномерно расположенными меланофорами разного размера; меланофорная пигментация рыла, вентральной половины жаберной крышки и вентральной стороны самых передних туловищных поясков сгущена сильнее; позади орбиты на жаберной крышке прослеживается чёткая светлая продольная полоса (рисунок, б). Лучи С густо пигментированы буроватыми меланофорами, остальные плавники прозрачные.
Малёк SL ~ 30 мм. D 16, A 2, P 14, C 9. Пластические признаки приведены в таблице. Поясков 14 + 40 = 54, около 20 последних хвостовых поясков образуют цепкий хвостовой конец тела. Рыло 2.5 раза в длине головы, почти равномерно высокое по всей длине, максимальная высота рыла (на уровне нижнечелюстного сустава) 2.5 раза содержится в его длине и в 1.3 раза превышает высоту рыла на уровне начала медиального гребня. Медиальный рыльный гребень начинается на середине длины рыла, с низким лопастевидным выступом над ноздрями; латеральный не выражен. Медиальный гребень головы более угловатый, чем у малька SL 25 мм. Вентральный костный край орбиты с широким боковым пластинчатым расширением (рисунок, б'). Гребни туловищных поясков менее резкие и не столь приподнятые, как у малька SL 25 мм, шипики по их свободному краю менее отчётливы (рисунок, в'). Расположение мочек на голове и теле соответствует описанному для малька SL 25 мм, но на голове мочки заметно короче, корональная мочка простая и содержится в диаметре глаза около 3.8 раза. Меланофорная пигментация на голове и теле у данного экземпляра практически не различима. По другим признакам он соответствует вышеприведённому описанию малька SL 25 мм.
ОБСУЖДЕНИЕ
Малёк SL 25 мм характеризуется чуть бóльшим числом лучей Р, чем было ранее отмечено для вида (16 против 12–15, обычно 14 (Dawson, 1982a)), других отличий по счётным признакам между изученными экземплярами и имеющимися описаниями вида нет. Вместе с тем два исследованных экземпляра при близких абсолютных размерах заметно отличаются друг от друга по степени развития мочек на голове, некоторых гребней на голове и гребней на туловищных поясках. Учитывая специфику межвидовой изменчивости в пределах других родов (Dawson, 1985), различия в строении гребней между исследованными особями вполне могли бы быть расценены как межвидовые. Наиболее существенным отличием представляется наличие у малька SL ~ 30 мм сильно развитого пластинчатого шельфа вентрального костного края орбиты, который совершенно отсутствует у малька SL 25 мм (рисунок, б, б'). На наличие такого шельфа у Amphelikturus ранее не обращали внимания, однако он показан на рисунке у Доусона (Dawson, 1982a. Fig. 4 ) и, по-видимому, является отличительной чертой A. dendriticus на дефинитивных стадиях онтогенеза. У малька SL 25 мм менее протяжённый медиальный рыльный гребень (около трети длины рыла, тогда как у малька SL ~ 30 мм – около половины его длины) (рисунок, б, б'), а гребни на туловищных поясках более высокие и протяжённые, сильнее зазубрены, и их шиповидные окончания развиты лучше (рисунок, в, в'). Вышеописанные признаки малька SL ~ 30 мм соответствуют характеристике A. dendriticus, приводимой Доусоном (Dawson, 1982a); единственным его отличием от этих данных является отсутствие латерального рыльного гребня, который, по данным Доусона (Dawson, 1982a), развит у A. dendriticus в виде тупых выступов. Оба изученных экземпляра характеризуются развитым оперкулярным гребнем, что, по данным Доусона (Dawson, 1982а), является ювенильной чертой (SL < 30 мм, у более крупных рыб он редуцируется); эта же особенность весьма характерна для многих видов рода Syngnathus (Fritzsche, 1980; Dawson, 1982a, 1985).
Доусон (Dawson, 1982a) приводит некоторые наблюдения в связи с изменчивостью в размере и строении мочек на голове и теле у A. dendriticus в зависимости от размеров тела. По его данным, у более крупных рыб мочки более многочисленные и сильнее ветвящиеся, а мочки головы ветвятся лишь при SL > 40 мм. Не отрицая подобную и, безусловно, ожидаемую закономерность, можно, однако, отметить, что у меньшего из изученных мною мальков мочки на голове неожиданно крупнее, чем у большего, причём корональная мочка значительно длиннее (равна диаметру глаза против почти вчетверо более короткой) и к тому же отчётливо ветвится. Таким образом, величина и степень ветвления мочек, вероятно, помимо онтогенетической имеют выраженную индивидуальную изменчивость, и корреляция этой изменчивости с размером рыб не всегда достаточно чёткая.
К диагнозу Amphelikturus, опубликованному Доусоном (Dawson, 1982a, 1985), следует добавить наличие парных супраорбитальных мочек, непарной корональной мочки и парных коротких округлых мочек очень характерной формы, мелко ветвящихся по свободному краю и расположенных на дорсальной поверхности четвёртого и вентральной поверхности одиннадцатого туловищных поясков. Судя по изученному материалу и данным, представленным Доусоном (Dawson, 1982a), положение этих мочек строго детерминировано, и они, вероятно, появляются уже на самых ранних стадиях онтогенеза. Расположение прочих мочек на голове и теле у A. dendriticus, по-видимому, подвержено возрастной и индивидуальной изменчивости.
Мальки SL 25 и ~30 мм заметно различаются между собой по пигментации – плотное расположение меланофоров, повсеместно развитое у меньшего экземпляра, практически исчезает у более крупного (более или менее сохраняясь лишь на челюстях и вентральной стороне головы и самых передних туловищных поясков). Из пластических признаков обращают на себя внимание заметно больший диаметр глаза, а также бóльшие высота тела на уровне середины основания D, предорсальное и преанальное расстояния у экземпляра SL ~ 30 мм (таблица).
Как же объяснить столь существенные различия между двумя исследованными мальками при их близких абсолютных размерах? Мне представляется правомочной следующая гипотеза. Малёк SL 25 мм представляет собой стадию “планктонного малька” (Dawson, Allen, 1978), которой свойственны менее развитый медиальный рыльный гребень, более “шипастые” окончания основных гребней на туловищных поясках и развитая меланофорная пигментация тела, тогда как характерный для вида пластинчатый шельф вентрального костного края орбиты ещё не выражен. Переход к дефинитивному строению гребней и утрата ювенильной пигментации происходит у A. dendriticus при достижении SL примерно 30 мм, но развитие латеральных рыльных выступов и редукция оперкулярного гребня происходят позднее, что и наблюдается у исследованного малька SL ~ 30 мм, структурно почти соответствующего взрослой рыбе. По данным Доусона (Dawson, 1982a), минимальный размер половозрелых самцов составляет SL 46.5 мм, при SL 33 мм оперкулярный гребень ещё полностью развит, но латеральные рыльные выступы уже присутствуют. С другой стороны, степень развития мочек не всегда хорошо коррелирует с ростом из-за вероятного адаптивного характера этих структур, связанных с камуфляжем и поддержкой тела в толще воды.
Список литературы
Barbour T. 1905. Notes on Bermudan fishes // Bull. Mus. Compar. Zool. V. 46. № 7. P. 109–134. Pls. 1–4.
Dawson C.E. 1977. Review of the pipefish genus Corythoichthys with description of three new species // Copeia. № 2. P. 295–338.
Dawson C.E. 1982a. Family Syngnathidae. The Pipefishes / Ed. Böhlke J.A. // Fishes of the Western North Atlantic. Sears Found. Mar. Res. Mem. 1. Pt. 8. New Haven: Yale Univer. P. 4–172.
Dawson C.E. 1982b. Descriptions of Cosmocampus retropinnis sp. n., Minyichthys sentus sp. n. and Amphelikturus sp. (Pisces, Syngnathidae) from the eastern Atlantic region // Zool. Scripta. V. 11 № 2. P. 135–140.
Dawson C.E. 1984. A new pipehorse (Syngnathidae) from Western Australia, with remarks on the subgenera of Acentronura // Jpn. J. Ichthyol. V. 31. № 2. P. 156–160.
Dawson C.E. 1985. Indo-Pacific Pipefishes (Red Sea to the Americas). Ocean Springs, Mississippi: Gulf Coast Res. Lab., vi + 230 p.
Dawson C.E., Allen G.R. 1978. Synopsis of the “finless” pipefish genera (Penetopteryx, Apterygocampus and Enchelyocampus, gen. nov.) // Rec. West Aust. Mus. V. 6. № 4. P. 391–411.
Dawson C.E., Yasuda F., Imai C. 1979. Elongate dermal appendages in species of Yozia (Syngnathidae) with remarks on Trachyrhamphus // Jpn. J. Ichthyol. V. 25. № 4. P. 244–250.
Frias-Torres S. 2006. Ch. 107. Syngnathidae: Seahorses and pipefishes / Ed. Richards W.J. // Early stages of Atlantic fishes: an identification guide for the Western Central North Atlantic. Boca Raton, Florida: CRC Press, Taylor and Francis Group. P. 1255–1260.
Fricke R., Eschmeyer W.N., van der Laan R. 2020. Catalog of Fishes: Genera, Species, References. (http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Version 01/2020)
Fritzsche R.A. 1980. Revision of the Eastern Pacific Syngnathidae (Pisces: Syngnathiformes), including both recent and fossil forms // Proc. Calif. Acad. Sci. V. 42. № 6. P. 181–227.
Parr A.E. 1930. Teleostean shore and shallow-water fishes from the Bahamas and Turks Island. (Scientific results of the third oceanographic expedition of the “Pawnee” 1927) // Bull. Bingham Oceanogr. Coll. Yale Univ. V. 3. Art. 4. P. 1–148.
Poll M. 1953. Poissons III. Téléostéens Malacoptérygiens // Résultats Scientifiques. Expédition Océanographique Belge dans les Eaux Côtières Africaines de l’Atlantique Sud (1948–1949). Bruxelles. V. 4. № 2. 258 p.+ 8 pls.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии