

Вопросы ихтиологии, 2021, T. 61, № 5, стр. 499-523
К систематике звездочётовых рода Uranoscopus индо-тихоокеанских вод с описанием трёх новых видов (Uranoscopidae)
1 Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
2 Институт океанологии РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 09.02.2021
После доработки 05.04.2021
Принята к публикации 06.04.2021
Аннотация
Приведен обзор звездочётов рода Uranoscopus Индо-Тихоокеанского региона. На основании изучения оригинальных материалов П. Бликера установлено, что название U. oligolepis до сих пор неправильно применялось к другому виду, описываемому в настоящей работе как U. alius sp. nov. Типовая серия U. oligolepis оказалась представленной двумя видами. В качестве лектотипа выбран экземпляр из вод Японии, принадлежащий к виду, известному под названием U. chinensis, над которым название U. oligolepis имеет приоритет. Два сохранившихся паралектотипа U. oligolepis (от Явы и Суматры) оказались конспецифичными виду U. cognatus. Впервые охарактеризована онтогенетическая изменчивость этого вида. Из вод Индии описываются два новых вида – U. batrachops и U. oculocirrhis spp. nov. Предложено распределить известные виды рода на пять видовых групп; ещё три вида занимают обособленное положение. Впервые составлена таблица для определения видов рода Uranoscopus Индо-Пацифики.
Род Uranoscopus Linnaeus, 1758 в настоящее время включает 27 валидных видов (Fricke et al., 2021), населяющих тропические и умеренные воды Восточной Атлантики (включая Средиземное и Чёрное моря) и Индо-Пацифики на восток до Океании, с центром видового разнообразия в тропической Индо-Вест-Пацифике (Красное море, северная часть Индийского океана и воды Индо-Австралийского архипелага). Систематика рода весьма слабо разработана, из-за чего определение видов может вызывать серьёзные затруднения, до сих пор нередки и описания новых видов (Randall, Arnold, 2012; Fricke, 2018; Прокофьев, 2020). Обобщающие работы по индо-тихоокеанским видам отсутствуют, а по регионам они исследованы крайне неравномерно. Достаточно подробно изучены виды фауны Японии (Kishimoto, 1987; Yamada, 2002) и Красного моря (Brüss, 1986, 1987a, 1987b; Randall, Arnold, 2012). Обзор фауны Индо-Австралийского архипелага (De Beaufort, Chapman, 1951) является устаревшим, он содержит неполный перечень видов и непригодную для практического использования определительную таблицу, однако имеется сравнительно недавняя работа Кишимото (Kishimoto, 2001) для тропической Западной Пацифики, содержащая определительную таблицу, карты ареалов, сведения по прижизненной окраске и иллюстрации каждого вида. Составленный в этой работе ключ является весьма практичным, однако пониманию ряда видов препятствует отсутствие морфологических описаний (не предусмотренных форматом данной работы). Сведения по фауне австралийских вод очень неполны. В обзоре Мееса (Mees, 1960) для фауны Австралии указаны только широко распространённый индо-пацифический вид U. cognatus Cantor, 1850 и описанный от Квинсленда проблематичный U. terraereginae Ogilby, 1910, позднее признанный синонимом U. affinis Cuvier, 1829 (Kishimoto, 2001). Однако исходя из карт ареалов, приводимых Кишимото (Kishimoto, 2001), в водах Северной и/или Северо-Западной Австралии можно ожидать нахождения всех девяти видов, указываемых для тропической Западной Пацифики. Хуже всего изучены звездочёты северной и западной части Индийского океана за пределами Красного моря, даже перечни видов, известных из этого региона (Heemstra, 1984; Fricke et al., 2013; Fricke, 2018), противоречивы. По-видимому, только три из девяти обитающих здесь видов (U. affinis, U. cognatus и U. oligolepis sensu Kishimoto (2001)) являются общими с западной частью Тихого океана. Некоторые авторы (Fricke et al., 2013; Vilasri, 2013) предпочитают отделять западнотихоокеанские популяции U. affinis в отдельный вид U. tosae (Jordan et Hubbs, 1925); следуя Кишимото (Kishimoto, 2001), я считаю эти виды конспецифичными. Из шести остающихся видов один (U. dollfusi Brüss, 1987), очевидно, является красноморским экспатриантом, проникающим в Персидский и Оманский заливы, тогда как прочие (U. archionema Regan, 1921; U. crassiceps Alcock, 1890; U. filibarbis Cuvier, 1829; U. guttatus Cuvier, 1829 и U. marmoratus Cuvier, 1829) являются чисто индоокеанскими. Четыре последних из перечисленных видов обитают в Аравийском море и у южной оконечности Индостана, из них U. guttatus проникает на юг до Сейшельских о-вов (Fricke, 2018). U. archionema, синонимами которого являются U. haplostoma Regan, 1921 и, по-видимому, U. duvalii Bottard, 1889 (Fricke et al., 2021), встречается южнее, от Сейшельского плато (собственные данные), о-ва Реюньон (тип U. duvalii) и побережья Кении до Южной Африки (Heemstra, 1986; Fricke et al., 2013). Таким образом, имеется два центра видообразования звездочётов рода Uranoscopus в Индо-Пацифике: красноморско-индийский (обитающего южнее U. archionema тоже следует отнести к этому центру, так как морфологически он исключительно близок к U. crassiceps и, очевидно, является его дериватом; с другой стороны, сюда нельзя причислять атлантическо-средиземноморский вид U. scaber Linnaeus, 1758, проникающий в Красное море) и западнотихоокеанский (от Южной Японии до Северной Австралии). Часть эндемичных красноморско-индийских видов представляются родственными западнотихоокеанским (см. обсуждение видовых групп), а часть, по-видимому, имеет автохтонное происхождение.
Более 20 лет назад я запланировал детальное исследование морфологии и систематики рода Uranoscopus, которое, к сожалению, до сих пор осталось не реализованным из-за ограниченности и нерепрезентативности материала по группе в отечественных коллекциях, полного отсутствия какой-либо централизованной поддержки музейных поездок и занятости в других проектах. За истёкшее время ряд задач, поставленных в рамках данного проекта, был решён другими исследователями. Так, были детально охарактеризованы остеология и миология всех родов и (остеология) более чем половины видов семейства (Vilasri, 2013) и описана часть выявленных новых видов (Fricke, 2018; Vilasri et al., 2019). Поскольку в обозримом будущем мне не приходится ожидать завершения данного проекта в том объёме, в котором он был запланирован, я решил суммировать в настоящем сообщении ряд номенклатурных и таксономических решений по некоторым проблемным видам рода, описать три новых вида из Индийского океана и упорядочить разнообразие рода путем выделения надвидовых группировок. Также предлагается таблица для определения всех индо-пацифических видов рода, попыток составления которой до сих пор никто не предпринимал.
МАТЕРИАЛ И МЕТОДИКА
Этикеточные данные экземпляров новых и переописываемых видов приведены в соответствующих разделах видовых описаний. Этот материал хранится в коллекциях Института океанологии РАН, Москва − ИО РАН; Британского музея, Лондон, Великобритания (Natural History Museum, London − BMNH); Центра биоразнообразия, Лейден, Нидерланды (The Naturalis Biodiversity Center, Leiden − RMNH). Для сравнения изучены следующие материалы из коллекций Зоологического института РАН (ЗИН РАН), ИО РАН и Парижского музея естественной истории (Muséum national d’Histoire naturelle − MNHN): голотипы U. arafurensis (MNHN 2019-0258), U. filibarbis (MNHN A-3098), U. guttatus (MNHN A-3097), U. marmoratus (MNHN 0000-5254) и U. sulphureus (MNHN 0000-5152), 43 экз. U. affinis SL 23–160 мм, 9 экз. U. oligolepis (= U. chinensis) SL 30 и 97–180 мм, 5 экз. U. japonicus SL 140–220 мм и 35 экз. U. kishimotoi SL 40–130 мм из вод Японии, Китая, Вьетнама и Индонезии (ИО РАН, ЗИН РАН), 7 экз. U. archionema SL 90–240 мм из западной части Индийского океана (ИО РАН), 2 экз. U. albesca SL 52 и 92 мм (ЗИН № 41008, район Дакара) и 40 экз. U. scaber SL 90–210 мм из Чёрного моря и вод Западной Африки (ИО РАН). Кроме того, по фотографиям изучены голотипы Zalescopus tosae № 58835 и Z. satsumae № 58839 из коллекции Музея Фильда, Чикаго, США (Field Museum of Natural History, Chicago − FMNH), голотип U. turbisquamatus № 29458 из коллекции биологического факультета Университета Коти, Япония (Kochi University − BSKU) и два экземпляра “U. oligolepis” SL 40.5 и 45.0 мм от Новой Гвинеи (вероятно, мальки U. affinis), упоминаемые в работе (Fricke, 2018) и любезно присланные Р. Фрике (Лауда-Кёнигсхофен, Германия) (Ronald Fricke, Lauda-Königshofen, Germany). Сведения о других видах взяты из литературных источников (Brüss, 1986, 1987a, 1987b; Kishimoto, 1984, 2001; Randall, Arnold, 2012; Fricke et al., 2013; Smith, 2016; Fricke, 2018). Методика изучения и терминология соответствуют принятым ранее (Brüss, 1987a; Kishimoto, 1987, 2001). Внутренним краем supracleithrum называется сторона выступающей наружу части кости, обращённая медиально; расположенные на ней шипы именуются внутренними; шип, которым оканчивается наиболее выступающая назад часть supracleithrum, называется задним. Вслед за Кишимото (Kishimoto, 2001. Fig. 9), я называю шипы, расположенные по латеральному краю pelvis, базиптеригиальными, по медиальному – клейтральными (название, вероятно, объясняется их положением на вершине pelvis, помещающейся между плечевыми поясами). Если последний луч в вертикальных плавниках разделён до основания, он указан как 1½. Счётные признаки, различающиеся на разных сторонах тела одной и той же рыбы, разделены знаком “/”. В формуле C строчными римскими цифрами приводится число дополнительных (procurrent) лучей, прописными – основных неветвистых, арабскими – ветвистых. В тексте используются следующие сокращения: SL, TL – соответственно стандартная и общая длина; D, A, P, V, C – соответственно спинной, анальный, грудные, брюшные и хвостовой плавники; vert. – число позвонков (туловищных + хвостовых); loc. typ. – типовая территория; НИС, НПС – соответственно научно-исследовательское и научно-промысловое судно.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Таксономические описания
Uranoscopus oligolepis Bleeker, 1879
(рис. 1)
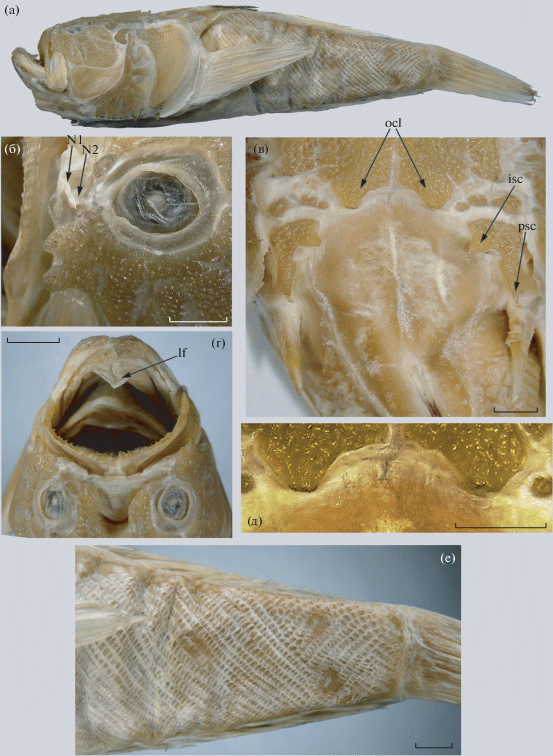
Рис. 1.
Uranoscopus oligolepis, лектотип RMNH 5941, SL 110 мм: а – общий вид, б – глаз и преорбитальная область, в – затылочная часть черепа и предорсальная область, г – прелингвальный вырост, д – затылочные доли, е – чешуя в задней части тела. lf – прелингвальный вырост; N1, N2 – передняя и задняя ноздри; isc, psc – внутренний и задний супраклейтральные шипы, ocl – затылочные доли. Масштаб, мм: б – 3; в, г – 4.5; д, е – 6.
Uranoscopus asper (non Temminck et Schlegel): Bleeker, 1853. P. 27.
Uranoscopus oligolepis: Bleeker, 1879. P. 55 (pars: Япония).
Uranoscopus chinensis: Guichenot in Sauvage, 1882. P. 169.
Uranoscopus flavipinnis: Kishimoto, 1987. P. 4. Fig. 2; Pietsch, Kishimoto, 1989. P. 748 (синонимия с U. chinensis).
Рис. 2.
Uranoscopus cognatus: RMNH 4884, SL 83 мм (а, в–ж, и, к) и RMNH 4843, SL 65 мм (б, 3), паралектотипы U. oligolepis: а, б – общий вид; в – глаза и межглазничная ямка, г – преорбитальные шипы, д – затылочная часть черепа и предорсальная область, е – выросты губ; ж, з – прелингвальный вырост, и – pelvis, к – затылочная комиссура. bp – базиптеригиальные шипы, pboc – задние ветви затылочной сейсмосенсорной комиссуры, so – супраокулярный усик, остальные обозначения см. на рис. 1. Масштаб, мм: в, е, ж – 4; г – 3.5; д, з – 5; и – 2; к – 2.5.

Uranoscopus chinensis auctorum.
Материал. Лектотип RMNH 5941 (выделен здесь), SL 110 мм (рис. 1а), Нагасаки, Япония.
Описание. D V + 13, A 12 ½, P 16, V i + 4, C v + I + 5 + 5 + I + iv; vert. 25 (10 + 15). Ширина головы в 1.15 раза больше её высоты. Покровные кости с грубым бугорчато-ямчатым (крышечные кости – с гребенчато-бугорчатым) орнаментом. Межглазничная ямка широкая (ширина в 1.9 раза меньше длины), параллельносторонняя, U-образная, с широко закруглённой вершиной, слегка заходящей за линию, соединяющую задние края орбит. Обе ноздри трубковидные. Два массивных преорбитальных шипа, из которых задний крупнее (рис. 1б). Супраорбитальные гребни плоские, надвисочные гребни едва намечены; париетальные гребни хорошо выражены, поверхность крыши черепа между ними вдавлена, бугорки не развиты; затылочные доли крупные, лопастевидные (рис. 1в, 1д). Верхняя губа с короткими, нижняя с более длинными простыми усиковидными выростами (рис. 1г). Пластинка maxillare с продольными гребнями. В челюстях острые конические зубы, на praemaxillaria близ симфиза расположенные в три неправильных ряда, назад переходящих в один; на dentalia соответственно в два, далее назад – в один ряд, задние зубы клыковидные. Зубы на сошнике образуют два раздельных поперечно вытянутых зубных пятна, около трёх неправильных рядов; на нёбных костях однорядные, но некоторые зубы выбиваются из ряда. Прелингвальный вырост в виде короткого треугольного лепестка, возможно, его дистальный конец, оборван (рис. 1г). На глазном яблоке (рис. 1б) и нижней челюсти нет усиковидного придатка. Нижний край praeoperculum с четырьмя шипами, suboperculum с одним направленным вниз шипом. Дорсальный край operculum слабо восходящий. Кожный край жаберной крышки практически ровный, лишь в своей верхней части волнистый. Supracleithrum оканчивается острым задним шипом и несёт крупный острый треугольный шип на внутреннем крае (рис. 1в). Клейтральный шип сравнительно короткий (4.4 раза в длине головы). У верхнего конца основания Р имеется кожная лопасть. Базиптеригиальных шипов одна пара. Верхнезадний край Р прямой, его седьмой–восьмой лучи наибольшие. Чешуя на боках тела образует ~38/42 косых рядов (с левой стороны чешуйные ряды инвертированы в задней трети тела (рис. 1е), с правой – несколько рядов непосредственно у основания С), которые доходят вперёд до заднего края плечевого пояса. Предорсальная область и участок между боковыми линями и основанием D голые, с единичными чешуйками под основанием колючей части D (рис. 1в). Затылочная комиссура с очень коротким срединным ответвлением (рис. 1д); поры восходящей части боковой линии открываются на очень коротких вентральных ответвлениях. Грудь, брюхо, основание Р и бока тела сразу позади основания Р голые; продольные кожные гребешки на брюхе не выражены. Экземпляр полностью депигментирован, сохранилась только чёрная окраска колючей части D.
Измерения суммированы в таблице.
Uranoscopus cognatus Cantor, 1849
Рис. 3.
Uranoscopus cognatus, голотип BMNH 1860.3.19.397a, верх головы (на врезке показана область supracleithrum) (а), и малёк ИО РАН № 3630, SL 22 мм (б–е): б, в – общий вид (б – сверху, в – сбоку); г – голова сбоку, д – межглазничная ямка, е – ошипление lacrimale и супраокулярный усик. pacr, tcr – соответственно париетальные и надвисочные гребни; остальные обозначения см. на рис. 1, 2. Масштаб, мм: а – 5; г, д – 2; е – 1.5.

Uranoscopus cognatus: Cantor, 1849. P. 1003.
Uranoscopus oligolepis: Bleeker, 1879. P. 55 (pars: Суматра, Ява, Амбоина).
Материал. Голотип BMNH 1860.3.19.397a, сухой гербарный образец, SL 98 мм (по сухому образцу), Пенанг. Паралектотипы U. oligolepis RMNH 4883, SL 65 мм, Ява; RMNH 4884, SL 83 мм, Суматра, Зондский пролив, коллектор П. Бликер. ИО РАН № 3630, SL 22 мм, 02°03′ с. ш. 105°37′ в. д., НИС “Витязь”, рейс 54, ст. 6227, трал Сигсби, глубина 73 м, 22.02.1973 г.
Диагноз. Вид рода Uranoscopus с двумя парами базиптеригиальных шипов, с умеренно широкой межглазничной ямкой (ширина в 2.5–3.4 раза меньше длины, увеличивается с ростом), с тремя задними ветвями затылочной комиссуры, 54–57 поперечными рядами чешуй, с усиковидным выростом на верхнем крае глазного яблока.
Описание11. D V + 14 (IV + 14 ½), A 13 ½, P 16, V i+ 4, C v + I + 5 + 5 + I + iv(v); vert. 25 (10 + 15). Голова и тело отчётливо сжатые с боков, ширина головы равна или в 1.15 раза меньше её высоты (рис. 2а, 2б, 3а). Глаза не выступают над верхним профилем черепа, но латерально виден лишь нижний край орбит (рис. 2а). Покровные кости головы не покрыты кожей, с грубым гребенчато-бугорчатым орнаментом. Межглазничная ямка слабо или умеренно сужена (ее ширина в 3.4 (3.1) раза меньше длины [у голотипа – примерно в 2.5 раза]), с широко закруглённой вершиной, слегка заходящей за линию, соединяющую задние края орбит [у голотипа немного не доходит до этой линии] (рис. 2в, 3а). У экземпляра SL 83 мм боковые стороны межглазничной ямки почти параллельны друг другу, тогда как у экземпляра SL 65 мм и у голотипа они отчётливо сходящиеся кзади. Обе ноздри трубковидные, передняя с ветвящимся придатком на вершине (рис. 2в). Преорбитальных шипов три, средний из них длинный и заострённый; первый и третий слабо развиты (рис. 2г) [у голотипа шипы выглядят более отчётливыми, но, возможно, это связано с тем, что образец сухой]. Супраорбитальные гребни простого строения, надвисочные и париетальные гребни хорошо выражены, поверхность крыши черепа между париетальными гребнями отчётливо желобовидно вдавлена; задние концы париетальных гребней образуют небольшие бугорки [у голотипа гребни на костях крыши черепа неотчётливые, дорсальная поверхность головы плоская]; затылочные доли слабо (SL 83 мм) (рис. 2д) или совсем не выражены. Нижняя губа несёт многочисленные выросты, в срединной трети раздвоенные у самых вершин или с короткими боковыми ответвлениями (у экземпляра SL 65 мм – только слегка раздвоённые); верхняя губа гладкая (рис. 2е). Верхний край maxillare с бугорковидным выступом примерно на середине длины; пластинка maxillare с резкими продольными гребнями. В челюстях острые конические зубы, в заднем отделе зубного ряда на dentale сильно увеличенные, клыковидные. На praemaxillaria зубы расположены в два раздельных ряда (у лектотипа непосредственно у симфиза – в три), зубы внутреннего ряда примерно в полтора раза крупнее наружных. На нижней челюсти в присимфизной части зубы в два, далее назад – в один ряд. Зубы на сошнике сидят в двух раздельных овальных зубных пятнах, как и на нёбных костях, мелкие. Прелингвальный вырост лентовидный, его боковые края фестончатые (рис. 2ж, 2з), будучи прижатым назад, немного не достигает (SL 83 мм) или слегка заходит за задний край межглазничной ямки. Верхний край глазного яблока с усиковидным придатком (рис. 2в); на нижней челюсти усика нет. Нижний край praeoperculum с четырьмя шипами, suboperculum с одним направленным вниз шипом. Дорсальный край operculum у экземпляра SL 83 мм слегка восходящий, верхнезадний угол кости закруглён; у экземпляра SL 65 мм – практически прямой, с отчётливо оттянутым верхнезадним углом; у голотипа – слабо наклонный к закруглённому верхнезаднему углу. Кожный край жаберной крышки с мелкими выростами, в нижней его трети постепенно сходящими на нет. Supracleithrum оканчивается коротким задним шипом, его внутренний край без развитых шипов (рис. 2д) [у голотипа – с двумя маленькими, но отчётливыми шипами (рис. 3а)]. Клейтральный шип очень длинный (2.4 (2.6) раза в длине головы), острый. У верхнего конца основания Р имеется кожная лопасть. Базиптеригиальных шипов две пары (рис. 2и), передняя пара длиннее задней; клейтральные шипы pelvis недоразвиты у обоих экземпляров [имеются у голотипа].
Морфометрические признаки рассматриваемых в статье видов рода Uranoscopus
| Признак | U. oligolepis, лектотип RMNH 5941 | U. cognatus | U. batrachops sp. nov., голотип ИО РАН № 3631 |
U. alius sp. nov., голотип ИО РАН № 3632 | U.oculocirrhis sp. nov., голотип ИО РАН № 3633 | ||
|---|---|---|---|---|---|---|---|
| RMNH 4884 | RMNH 4883 | ИО РАН № 3630 | |||||
| SL, мм | 110 | 83 | 65 | 22 | 82 | 115 | 108 |
| В % SL | |||||||
| Длина головы | 36.4 | 33.70 | 35.40 | 40.90 | 34.15 | 38.3 | 38.90 |
| Высота головы | 24.6 | 22.90 | 23.10 | 25.00 | 21.95 | 22.6 | 23.15 |
| Максимальная ширина головы | 28.2 | 19.90 | 23.10 | 29.55 | 21.95 | 27.8 | 29.60 |
| Максимальная высота тела | 27.3 | 23.50 | 23.10 | 22.70 | 26.80 | 23.5 | 25.00 |
| Минимальная высота тела | 11.8 | 12.05 | 11.50 | 11.40 | 11.60 | 10.9 | 11.10 |
| Длина хвостового стебля | 11.8 | 9.00 | 6.90 | 9.10 | 11.00 | 12.2 | 13.90 |
| Первое предорсальное расстояние | 36.4 | 36.15 | 35.40 | 43.20 | 39.00 | 40.9 | 44.40 |
| Второе предорсальное расстояние | 50.9 | 50.60 | 46.15 | 54.55 | 51.20 | 56.5 | 58.30 |
| Преанальное расстояние | 52.7 | 55.40 | 49.20 | 56.80 | 54.90 | 59.1 | 55.60 |
| Превентральное расстояние | 20.0 | 15.70 | 15.40 | 22.70 | 15.85 | 18.3 | 15.70 |
| Длина Р | 29.1 | 24.10 | 29.20 | 31.80 | 24.40 | 27.0 | 25.90 |
| Длина V | 24.6 | 16.90 | 18.50 | 20.50 | 15.85 | 23.5 | 22.20 |
| Длина С | 24.6 | 24.10 | 27.70 | 25.00 | 35.40 | 27.0 | 25.50 |
| Длина рыла | 3.6 | 2.40 | 3.10 | 3.40 | 3.70 | 4.8 | 4.20 |
| Горизонтальный диаметр глаза | 5.5 | 5.40 | 5.40 | 8.20 | 6.10 | 6.1 | 6.90 |
| Ширина костного межглазничного промежутка | 8.2 | 4.20 | 6.15 | 8.00 | 4.90 | 7.8 | 8.80 |
| Длина межглазничной ямки | 7.3 | 7.20 | 7.70 | 9.10 | 7.90 | 7.4 | 8.30 |
| Ширина межглазничной ямки на уровне середины глаза | 3.9 | 3.00 | 2.50 | 4.60 | 1.80 | 5.2 | 5.10 |
| Преорбитальная длина | 5.0 | 4.20 | 6.15 | 6.80 | 4.90 | 5.2 | 5.60 |
| Ширина промежутка между верхними краями жаберных отверстий | 22.7 | 19.30 | 20.00 | 20.50 | 18.30 | 20.0 | 18.50 |
| Расстояние между вершинами затылочных долей (при их отсутствии, задними концами париетальных гребней) | 8.2 | 6.00 | 5.40 | 9.10 | 6.10 | 5.2 | 5.60 |
| Длина клейтрального шипа | 8.2 | 13.90 | 13.85 | 11.40 | 14.60 | 9.6 | 9.70 |
| Длина прелингвального выроста | 3.2+ | 10.90 | 15.40 | 25.00 | 2.40 | 0 | 25.90 |
Колючая и мягкая части D разделены. P, прижатый к телу, заходит за основание третьего–четвертого луча А. Верхнезадний край Р прямой, лучи с шестого по восьмой наибольшие; длина лучей V прогрессивно увеличивается от первого луча к четвёртому, третий и пятый лучи примерно одинаковой длины. С усечённый. Тело покрыто циклоидной чешуёй, образующей на боках 57 (54) косых рядов, вперёд не доходящих до заднего края плечевого пояса [у голотипа чешуйный покров сохранился очень плохо и точное число рядов не может быть подсчитано, но их число, по-видимому, было меньше 60]. Предорсальная область и участок между боковыми линями и основанием D абсолютно голые, без чешуи (рис. 2д, 2к). Поры боковой линии на её восходящем участке открываются на коротких ответвлениях; затылочная комиссура с тремя задними ветвями (рис. 2к), достигающими назад уровня заднего шипа supracleithrum. Грудь, брюхо, основание Р и бока тела сразу позади основания Р голые. Брюхо с многочисленными тонкими продольными кожными гребешками.
Оба лейденских экземпляра депигментированы, сохранилась только чёрная окраска колючей части D. У голотипа окраска верха тела однотонно-коричневатая, низ светлее, нижняя часть Р и С жёлтая, колючая часть D чёрная, перепонка мягкой части D и А, по-видимому, слабо или совсем не пигментирована.
Измерения суммированы в таблице.
Малёк SL 22 мм (рис. 3б–3е). D V + 14, A 13 ½, P 16, V i + 4, C vi + I + 5 + 5 + I + iv. Ширина головы в 1.18 раза меньше её высоты; ширина межглазничной ямки вдвое меньше длины, её вершина не достигает линии, соединяющей задние края орбит, боковые стороны параллельны друг другу (рис. 3д). Передняя ноздря в виде длинной трубочки, несущей на вершине несколько бугорковидных выступов, будучи прижата вперёд, достигает переднего края верхней губы. Задняя ноздря в виде полутрубки, в полтора раза короче передней, с двумя короткими выростами на вершине. Скульптура покровных костей головы слабая, гребенчато-ячеистая, но гребни и бугорки на костях сильно развиты. Передний конец frontale образует отчётливый шипик у передневерхнего края орбиты; задневерхний костный край орбиты приподнят в виде валика; надвисочные гребни ребровидные, париетальные гребни высокие и оканчиваются треугольными выступами (рис. 3г). Все гребни головы несут тупые бугорки. Инфраорбитальные кости несут сильные тупые бугорки, имеющие вид шипиков (рис. 3е). Хорошо развит шипик в области головки hyomandibulare и у вершин нижнечелюстных гребней. Нижняя губа с множественными короткими выростами, на верхней имеется только один бугорковидный вырост на симфизе praemaxillaria. Прелингвальный вырост лентовидный, с фестончатыми боковыми краями, назад достигает заднего края черепа. Верхний край глазного яблока с крупным усиковидным придатком, длина которого составляет около половины диаметра глаза (рис. 3г–3е). Дорсальный край operculum с отчётливо оттянутым кверху верхнезадним углом. Клейтральные шипы pelvis и две пары базиптеригиальных шипов хорошо развиты. Р, прижатый к телу, достигает основания пятого луча А. С слабо закруглённый. Чешуя совершенно не развита. Затылочная комиссура с тремя очень короткими задними ветвями. В остальном соответствует описанным выше более крупным экземплярам, измерения приведены в таблице. По сравнению с более крупными экземплярами малёк выделяется более крупной головой, большей величиной глаза, гораздо более длинным прелингвальным выростом, большей величиной межглазничного промежутка и ямки, преорбитальной длиной и расстоянием между задними концами париетальных гребней, более задним положением D и несколько более длинными парными плавниками.
Пигментация представлена трудноразличимыми (видна только под бинокуляром) буроватыми хроматофорами на дорсолатеральной поверхности тела, множественными точечными чёрными меланофорами на верхней и боковых сторонах головы, в предорсальной области, в основании Р и в верхней половине боков туловища назад примерно до конца прижатого к телу Р и единичными – на боковых сторонах хвостового стебля. Плавники не окрашены (рис. 3б, 3в). Ноздри и прелингвальный вырост не пигментированы, супраокулярный усик буроватый.
Номенклатурные замечания
Хорошо известный в литературе вид U. cognatus характеризуется трубковидными ноздрями, из которых передняя несёт на вершине пучок филаментов, а у задней задняя стенка гораздо выше передней, и двумя парами базиптеригиальных отростков (Kishimoto, 2001)22; последний признак уникален в пределах семейства и справедливо рассматривается в качестве аутапоморфии (Vilasri, 2013). Строение ноздрей также специфично − как наличие дистальных филаментов у передней ноздри, так и форма задней, поскольку у других видов, имеющих трубковидную заднюю ноздрю (U. chinensis, U. kishimotoi), и передняя, и задняя её стенки примерно равной высоты (рис. 4). Два синтипа U. oligolepis от Суматры и Явы по этим и ряду других признаков полностью соответствуют виду U. cognatus. Третий синтип (от Японии) безусловно не конспецифичен остальным и определён мною, как U. chinensis. Первоописание Бликера (Bleeker, 1879) основано на четырёх экземплярах длиной (общей?) от 81 до 148 мм от Суматры, Явы, Амбоины и Японии. Амбонский экземпляр, по-видимому, утрачен, а три других хранятся в коллекции RMNH с пометкой “синтипы”. В Каталоге рыб (Fricke et al., 2021) типовой статус этих экземпляров указан под вопросом. Я считаю сомнения в типовой принадлежности данных рыб малообоснованными. Размеры экземпляров (83, 103 и 139 мм TL) не слишком отличаются от указанных в первоописании, а расхождения могут быть объяснены временем хранения и/или методикой измерений. Основным диагностическим признаком U. oligolepis признаётся малое число рядов чешуй на боках тела (38). Этот подсчёт совпадает со сделанным мною для синтипа RMNH 5941 из Нагасаки, однако столь низкое число рядов является несомненной аберрацией и обусловлено реверсией чешуй в задней части тела (рис. 1е). Это явление нередко у видов Uranoscopus, хотя в других известных мне случаях выражено в значительно меньшей степени. Таким образом, данный признак не имеет диагностической значимости, но он свидетельствует о том, что экземпляр RMNH 5941 входит в типовую серию и что основной диагностический признак вида у Бликера основан именно на нём. У двух других синтипов из вод Зондских островов число рядов чешуй заметно большее (52–57). В первоописании Бликера также указана окраска верхнебоковой поверхности тела как состоящая из нерегулярных розоватых или розово-жёлтых пятен, что соответствует таковой у U. chinensis. Прочие признаки, указанные в первоописании U. oligolepis, не имеют диагностической ценности. Учитывая, что экземпляр RMNH 5941 в наибольшей степени согласуется с первоописанием, я выделяю его в качестве лектотипа вида U. oligolepis, и это название оказывается старшим синонимом U. chinensis и валидным названием для данного вида (U. flavipinnis Kishimoto, 1987 = U. chinensis Guichenot, 1882 = U. oligolepis Bleeker, 1879). Паралектотипы RMNH 4883 и 4884 отнесены к виду U. cognatus.
Рис. 4.
Форма носовых трубочек (схематично) Uranoscopus cognatus (а) и U. oligolepis (б), обозначения см. рис. 1.

Следует отметить, что на конспецифичность U. asper sensu Bleeker (1853) виду U. flavipinnis (= U. chinensis) указал еще Кишимото (Kishimoto, 1987), однако работу Бликера 1853 г. с первоописанием U. oligolepis и отождествлением с этим видом его “U. asper” Кишимото не обсуждал. Позднее Кишимото (Kishimoto, 2001) применял название “U. oligolepis” для вида, называемого мною U. alius sp. nov. и не имеющего ничего общего с типовой серией U. oligolepis. В коллекции RMNH имеется рукописная этикетка Р. Брюсса (R. Brüss) с выделением экземпляра RMNH 5941 в качестве лектотипа U. oligolepis, однако это не было опубликовано.
Uranoscopus batrachops Prokofiev, sp. nov.
(рис. 5)
Рис. 5.
Uranoscopus batrachops sp. nov., голотип ИО РАН № 3631, SL 82 мм: а – общий вид, б – голова сверху, в – прелингвальный вырост, г – ноздри, д – глаз и преорбитальная область, е – pelvis, ж – верх головы дорсолатерально, з – затылочная комиссура. iof – межглазничная ямка, остальные обозначения см. на рис. 1–3. Масштаб, мм: а, б, г–е, з – 5; в – 2, ж – 3.

Материал. Голотип ИО РАН № 3631, SL 82 мм (рис. 5а), Индия, у устья р. Хугли, глубина 20 морских саженей (36.6 м), оттертрал, получено с экспериментального индийского траулера (Калькутта), НИС “Витязь”, рейс 33, проба 741, 23.02.1961 г.
Диагноз. Вид рода Uranoscopus с двумя парами базиптеригиальных шипов, с очень узкой межглазничной ямкой (ширина в 4.4 раза меньше длины при SL 82 мм), с шестью задними ветвями затылочной комиссуры, с около 65 поперечными рядами чешуй, без усиковидного выроста на глазном яблоке.
Описание. D IV + 14, A 14, P 17 (i + 15 + ii), V i + 4, C vi + I + 5 + 5 + I + v; vert. 25 (10 + 15).
Голова и тело заметно сжатые с боков, ширина головы равна её высоте. Глаза расположены дорсолатерально, верхний край глаз выступает над верхним профилем черепа. Покровные кости головы не покрыты кожей, с грубым бугорчатым и гребенчато-бугорчатым орнаментом. Межглазничная ямка узкая (ее ширина в 4.4 раза меньше длины), с широко закруглённой вершиной, немного не достигающей линии, соединяющей задние края орбит (рис. 5б). Передняя ноздря трубковидная, на вершине несёт пучок усиковидных придатков на узком основании; задняя открывается полутрубкой, у которой передний край заметно ниже заднего; высота трубочки передней ноздри без придатка в 1.5 раза больше таковой задней ноздри (рис. 5г). Преорбитальных шипов 3/4, антеромедиальный угол lacrimale справа выступает в виде маленького шипа; второй (справа третий) шип намного длиннее остальных, последующий за ним шип едва намечен (рис. 5д). Гребни и бугорки крыши черепа плохо выражены, хорошо заметны лишь париетальные гребни, которые в задней половине более отчётливы (рис. 5ж); поверхность крыши черепа плоская; затылочные доли не развиты. Нижняя губа несёт около 22 выростов, большей частью коротко разветвлённых на вершине (некоторые – с двумя боковыми веточками, самые крайние – бугорковидные); верхняя губа гладкая, лишь посередине симфиза имеется короткий выступ, по бокам которого сидят несколько коротких усиковидных выростов (рис. 5д). Верхний край maxillare с бугорковидным выступом на уровне нижнего края инфраорбитальных костей, пластинка – с тонкими продольными гребнями. В челюстях острые конические зубы, более крупные на dentalia, чем на praemaxillaria. На praemaxillaria непосредственно у симфиза зубы наиболее крупные, расположены в три нечётких ряда, далее назад зубы сидят в два чётко разделённых ряда, в наружном они крупнее и более расставлены. Нижнечелюстные зубы у симфиза в два ряда, далее назад однорядные. Зубы на сошнике мелкие, конические, образуют два разделённых, поперечно вытянутых зубных пятна, в которых они сидят в два ряда; на нёбных костях зубы более крупные, в один ряд, по четыре с каждой стороны. Прелингвальный вырост короткий, треугольный, с нитевидно оттянутой вершиной, конец которой, будучи прижатым кпереди, достигает края нижней губы (рис. 5в). Усиковидных придатков на глазном яблоке и на нижней челюсти нет (рис. 5д, 5ж). Нижний край praeoperculum с четырьмя расставленными шипами, suboperculum с одним направленным вниз шипом. Дорсальный край operculum прямой. Кожный край жаберной крышки с мелкими выростами, в нижней его трети сходящими на нет. Supracleithrum оканчивается коротким задним шипом и несёт два шипа на внутреннем крае (рис. 5ж), из которых передний короче заднего. Клейтральный шип очень длинный (2.3 раза в длине головы), острый. У верхнего конца основания Р имеется хорошо выраженная кожистая лопасть. Базиптеригиальных шипов две пары (рис. 5е), передняя пара длиннее задней.
Колючая и мягкая части D соприкасаются. Р, прижатый к телу, достигает вертикали, проходящей между основаниями второго и третьего лучей А. Верхнезадний край Р прямой, слабо выраженный за счёт того, что наибольшие лучи этого плавника − с пятого по седьмой, вершина плавника плавно закруглена. Длина лучей V прогрессивно увеличивается от первого луча к четвёртому, третий и пятый лучи равны и немного короче четвёртого. С усечённый. Тело покрыто мелкой циклоидной чешуёй, образующей на боках около 65 косых рядов, вперёд не доходящих до заднего края плечевого пояса. Предорсальная область и участок между боковыми линями и основанием D абсолютно голые, без чешуи. Поры боковой линии на её восходящем участке (до начала мягкой части D) открываются на хорошо развитых дорсальных и вентральных ответвлениях; затылочная комиссура с шестью длинными задними ветвями (рис. 5з). Грудь, брюхо, основание Р и бока тела сразу позади основания Р голые. Брюхо с многочисленными тонкими продольными кожными гребешками.
Окраска верхней части головы и тела коричневая, однотонная, книзу светлеющая; вентральная поверхность головы, грудь и брюхо чисто-белые. Перепонка колючей части D интенсивно-чёрная, лучи мягкой части D и дистальный конец С затемнены; перепонка и лучи Р с мелкими точечными меланофорами, значительно лучше развитыми в нижней половине плавника; V и А совершенно не пигментированы. Ротожаберная полость светлая, прелинвальный филамент с меланофорной пигментацией. Трубочки обеих ноздрей пигментированы не сильнее окружающей их поверхности рыла.
Измерения см. в таблице.
Этимология. Название (от греч. “batrachos” (лягушка) и “ops” (глаз)) отражает характерно выступающие в профиль глаза у нового вида.
Сравнительные замечания. Новый вид отличается от U. cognatus сильнее выступающими вверх глазами, более узкой межглазничной ямкой (1.8% против 2.5–3.0% SL при SL < 85 мм), отсутствием супраокулярного усика, шестью (против трёх) задними ветвями затылочной комиссуры, формой прелингвального выроста и более мелкой чешуёй (65 рядов против 52–57). В литературе эти два вида, по-видимому, не разделяли. Результаты изучения митогенома U. cognatus из вод Малайзии и Таиланда показали, что поддержка кластеризации выборок из разных районов низкая (Gan et al., 2019), что может быть связано со сборной природой материала. Голотип U. cognatus (BMNH 1860.3.19.397a) представляет собой сухой гербарный образец (левая сторона тела), на котором большинство вышеописанных отличий установить невозможно. Однако голотип U. cognatus имеет явственно более широкую межглазничную ямку (рис. 3а, 5б) и более крупную чешую (хотя точное число рядов чешуй подсчитать невозможно, их явно меньше 60). На этих основаниях название U. cognatus не может быть применено к описываемому виду.
Uranoscopus alius Prokofiev, sp. nov.
(рис. 6)
Рис. 6.
Uranoscopus alius sp. nov., голотип ИО РАН № 3632, SL 115 мм: а – общий вид, б – глаз и преорбитальная область, в – голова и предорсальная область сверху, г – голова дорсолатерально, д – крыша черепа и межглазничная ямка, е – выросты губ, ж – pelvis. amp – антеромедиальный выступ lacrimale, troc – поперечный заглазничный гребень, остальные обозначения см. на на рис. 1, 3, 5. Масштаб, мм: б, г–ж – 5; в – 10.

Uranoscopus oligolepis (non Bleeker, 1879): Kishimoto, 2001. P. 3523, 3528.
Материал. Голотип ИО РАН № 3632, SL 115 мм (рис. 6а), банка Сая-де-Малья, НПС “Профессор Месяцев”, рейс 12, трал 10.
Диагноз. Вид рода Uranoscopus с одной парой базиптеригиальных отростков, без чешуи на верхней стороне тела между ветвями боковой линии, с простой задней ноздрёй, с одним внутренним супраклейтральным шипом, четырьмя или пятью преоперкулярными шипами, слабыми, но отчётливыми поперечным постокулярным, париетальными и надвисочными гребнями крыши черепа, без затылочных долей, с 46 поперечными рядами чешуй на боках тела, с боковыми краями pelvis, расходящимися к вершине, без рисунка на дорсальной стороне тела (у выцветших коллекционных экземпляров точечные меланофоры концентрируются в нижних двух третях боков тела и в базальной трети Р).
Описание. D V + 13, A 13 ½, P 17 (i + 15 + ii), V i + 4, C v + I + 5 + 4 + I + iv; vert. 24 (10 + 14). Ширина головы в 1.2 раза больше её высоты. Покровные кости головы не покрыты кожей, с грубым бугорчатым и гребенчатым орнаментом. Межглазничная ямка широкая (её ширина только в 1.4 раза меньше длины), коническая, с широко закруглённой вершиной, не достигающей линии, соединяющей задние края орбит. Передняя ноздря трубковидная, задняя открывается овальным отверстием с немного приподнятыми краями. Преорбитальных шипов три, средний из них значительно длиннее остальных, первый шип отчётливый, третий – едва намечен; антеромедиальный край lacrimale перед орбитой образует валиковидный выступ, не продолжающийся в шип (рис. 6б). Гребни и бугорки крыши черепа низкие и слабые, поверхность крыши черепа плоская; затылочные доли не развиты, но задняя часть париетальных гребней приподнята, образуя невысокие выступы. Надвисочные гребни почти столь же отчётливы, как и париетальные. Супраорбитальный гребень не образует выступа у верхнезаднего края орбиты или позади него; эти гребни соседних сторон соединены поперечным посторбитальным гребнем (рис. 6в–6д). Нижняя губа несёт около 35 выростов, длинных, с простыми вершинами в средних двух третях, бугорковидных в боковых третях губы; верхняя губа несёт около 22 бугорковидных выростов, которые лишь непосредственно у симфиза praemaxillaria приобретают вид коротких усиков (рис. 6е). В челюстях острые конические зубы; на praemaxillaria они расположены в два разобщённых ряда, во внутреннем ряду зубы явственно крупнее. Нижнечелюстные зубы в один ряд, непосредственно у симфиза − в два. Зубы на сошнике в три ряда, образуют два разделённых, поперечно вытянутых зубных пятна; на нёбных костях – однорядные. Нет никаких следов прелингвального выроста. Усиковидных придатков на глазном яблоке и на нижней челюсти нет (рис. 6в–6д). Нижний край praeoperculum слева с четырьмя, справа с пятью шипами, suboperculum с одним направленным вниз шипом. Дорсальный край operculum восходящий от суставной головки к верхнезаднему углу. Кожный край жаберной крышки с мелкими выростами. Supracleithrum оканчивается острым шипом и несёт ещё один заострённый шип на внутреннем крае (рис. 6в). Клейтральный шип длинный и острый. У верхнего конца основания Р имеется небольшая кожистая лопасть. Базиптеригиальных шипов одна пара (рис. 6ж).
Колючая и мягкая части D снаружи разделены небольшим промежутком (около трети длины орбиты). Р, прижатый к телу, достигает вертикали основания четвёртого луча А. Верхнезадний край Р прямой, вершина плавника плавно закруглена, лучи с седьмого по девятый самые длинные. Длина лучей V прогрессивно увеличивается от первого луча к четвёртому, третий и пятый лучи равны и немного короче четвёртого. С усечённый. Тело покрыто циклоидной чешуёй, образующей на боках около 46 косых рядов на левой стороне (с правой стороны чешуйные ряды инвертированы в задней трети туловища), вперёд не доходящих до заднего края плечевого пояса. Предорсальная область и участок между боковыми линями и основанием D абсолютно голые, без чешуи (рис. 6в). Поры боковой линии на её восходящем участке (до начала мягкой части D) открываются на коротких дорсальных и вентральных ответвлениях; затылочная комиссура с тремя короткими задними ветвями, не достигающими линии, проведённой через вершины внутренних супраклейтральных шипов соседних сторон. Грудь, брюхо, основание Р и бока тела сразу позади основания Р голые. Брюхо с многочисленными тонкими продольными кожными гребешками.
Окраска. Голотип обесцвечен, лишь при увеличении на боках тела видна лишь очень мелкая тёмная меланофорная пигментация, меланофоры в нижней половине боков тела заметно более крупные, но редкие. Преорбитальная область и вершина нижней челюсти в разреженных чёрных точках. Плавники, кроме дистальной половины С и колючей части D, не пигментированы, но в основании Р имеются многочисленные, не различимые невооружённым глазом точечные меланофоры (в дистальных двух третях Р они совершенно отсутствуют). Дистальный конец С затемнён, лучи и перепонки четырёх передних колючих лучей D (кроме основания) интенсивно-чёрные.
Измерения см. в таблице.
Этимология. Название вида от латинского “alius” – другой, не тот, который нужен, в связи с тем, что ранее к нему применялось название U. oligolepis, в действительности принадлежащее иному, притом совершенно непохожему виду.
Сравнение. Новый вид очень похож на U. marmoratus Cuvier, 1829 (рис. 7), однако при непосредственном сравнении голотипов были выявлены следующие различия: у нового вида супраорбитальные гребни совершенно плоские, тогда как у U. marmoratus они образуют отчётливые бугорки позади глаз (рис. 7а); затылочные доли совершенно отсутствуют; задний край maxillare спрямлён, тогда как у U. marmoratus он отчётливо скошен к нижнезаднему углу (рис. 7а); выросты верхней губы бугорковидные или сосочковидные (усиковидные у U. marmoratus) (рис. 6е, 7б); pelvis расширен кпереди, а не наоборот (рис. 6ж, 7г); чешуйные ряды на боках не доходят до заднего края плечевого пояса и их число заметно меньшее (~46 против ~58). В настоящее время оба голотипа обесцвечены, однако на дорсальной поверхности голотипа U. marmoratus прослеживаются остатки тёмного рисунка, отграничивающего крупные белые пятна (рис. 7в). В первоописании даётся следующая характеристика прижизненной окраски голотипа U. marmoratus: “tout le dos est marbré de taches rondes, pâles, serrées et un peu mêlées, sur un fond brun” − вся спина пятнистая с круглыми, светлыми, плотно сидящими и слегка смешанными пятнами на коричневом фоне (Cuvier, 1829. P. 305). Кишимото описывает прижизненную окраску для своего “U. oligolepis” как «однотонно-оливково-коричневую» (Kishimoto, 2001. P. 3528). У голотипа U. alius sp. nov. никаких следов пигментации на дорсальной поверхности тела не прослеживается.
Рис. 7.
Uranoscopus marmoratus, голотип MNHN 0000-5254: а – голова сбоку, б – выросты губ и прелингвальный вырост, в – голова и предорсальная область сверху, г – pelvis. pot – заглазничный бугорок, остальные обозначения см. на на рис. 1. Масштаб, мм: а, б, г – 5; в – 2.5.

Uranoscopus oculocirrhis Prokofiev, sp. nov.
(рис. 8)
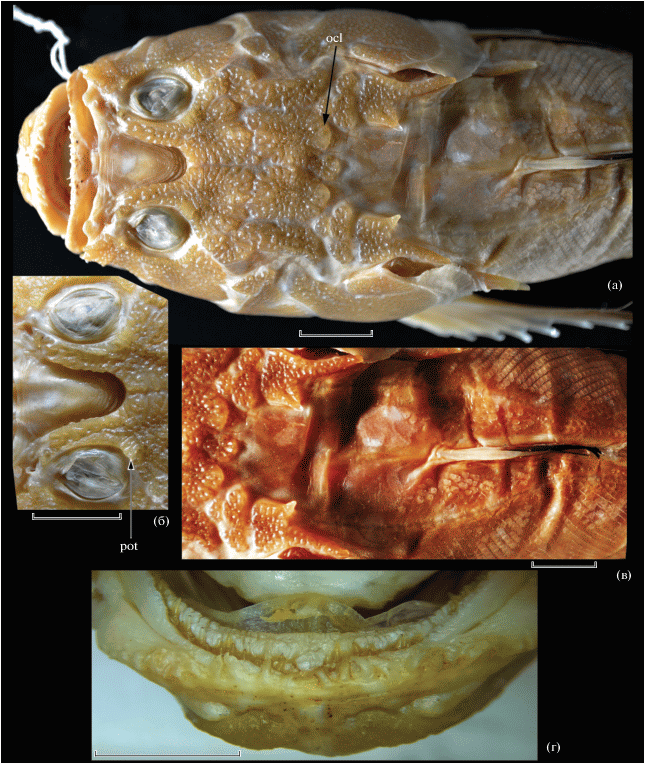
Рис. 8.
Uranoscopus oculocirrhis sp. nov., голотип ИО РАН № 3633, SL 108 мм: а – общий вид, б – затылочный край черепа и предорсальная область, в – супраокулярный усик, г – преорбитальная область, д – выросты губ и прелингвальный вырост. Обозначения см. на рис. 1, 2. Масштаб, мм: б – 10, в – 2.5; г, д – 5.

Рис. 9.
Uranoscopus guttatus, голотип MNHN A-3097: а – голова и предорсальная область сверху, б – область глаз и межглазья, в – затылочный край черепа и верх туловища (применён красный светофильтр для лучшей визуализации остатков пигментации), г – прелингвальный вырост. Обозначения см. на рис. 1, 7. Масштаб, мм: а–в – 10, г – 5.
Материал. Голотип ИО РАН № 3633, SL 108 мм (рис. 8а), 17°09′–17°07′ с. ш. 71°53′–71°51′ в. д., 118–133 м, НИС “Витязь”, рейс 33, станция 4863, проба 344, оттертрал, длина троса 500 м, время лова 11:55–12:30, 07.12.1960 г.
Диагноз. Вид рода Uranoscopus с одной парой базиптеригиальных отростков, с регулярными рядами чешуй между колючей частью D и боковой линией и небольшим участком чешуи в предорсальной области (рис. 8б), с простой задней ноздрей, с одним внутренним супраклейтральным шипом, четырьмя преоперкулярными шипами, без затылочных долей, с однорядными нижнечелюстными зубами, длинным лентовидным прелингвальным выростом и с усиковидным выростом на верхнем крае глазного яблока (рис. 8в).
Описание. D V + 12, A 12, P 18 (i + 16 + i), V i + 4, C vi + I + 5 + 4 + I + v; vert. 24 (10 + 14). Ширина головы в 1.3 раза больше её высоты. Покровные кости головы не покрыты кожей, с грубым бугорчатым и гребенчатым орнаментом. Межглазничная ямка конической формы (ширина в 1.6 раза меньше длины) с широко закруглённой вершиной, достигающей линии, соединяющей задние края орбит. Передняя ноздря трубковидная, задняя в виде продольно вытянутой щели с приподнятыми краями. Преорбитальных шипов три, средний из них значительно длиннее примерно равных и слабо обозначенных первого и третьего (рис. 8г); антеромедиальный край lacrimale образует маленький бугорок перед орбитой. Супраорбитальные гребни на frontalia образуют небольшие, но отчётливые бугорки кзади и кнутри от орбит, не соединенные поперечным гребнем; отграниченное вдавление в заглазничной части frontalia не выражено; надвисочные и париетальные гребни отчётливые, поверхность крыши черепа между ними слабо вогнута; затылочные доли не развиты. Нижняя губа несет 24 длинных ресницевидных выроста с простыми вершинами; верхняя губа гладкая (рис. 8д). В челюстях острые конические зубы; на praemaxillaria они расположены в два разобщённых ряда, во внутреннем ряду зубы заметно крупнее. Нижнечелюстные зубы однорядные. На сошнике острые конические зубы образуют два разделённых, поперечно вытянутых зубных пятна (до трёх рядов зубов в ширину); на нёбных костях такие же зубы в один ряд. Прелингвальный вырост (рис. 8д) длинный (1.5 раза в длине головы), лентовидный, в основании расширенный, в базальной трети своей длины с хорошо развитой по краям бахромой. Конец извлечённого из пасти и прижатого назад прелингвального выроста достигает заднего края нейрокрания. На глазном яблоке имеется усиковидный придаток, длина которого в 4.5 раза меньше диаметра глаза (рис. 8в). Нижний край praeoperculum с четырьмя шипами, suboperculum с одним направленным вниз шипом. Дорсальный край operculum восходящий от суставной головки к верхнезаднему углу. Кожный край жаберной крышки с мелкими выростами в верхней половине. Supracleithrum оканчивается острым задним шипом, верхний край которого зазубрен, и несёт один сильный шип на внутреннем крае (рис. 8б). Клейтральный шип длинный и острый. У верхнего конца основания Р имеется короткий кожный гребень. Базиптеригиальных шипов одна пара.
Колючая и мягкая части D снаружи соприкасаются, последний колючий луч отстоит от предшествующих, на равном расстоянии от предпоследнего и от первого луча мягкой части D. Р, прижатый к телу, достигает вертикали основания пятого луча А. Верхнезадний край Р прямой, вершина плавника плавно закруглена, девятый луч самый длинный. Длина лучей V прогрессивно увеличивается от первого луча к четвёртому, третий и пятый лучи незначительно короче четвёртого. Концы верхних лучей С обломаны, поэтому форму заднего края плавника с уверенностью установить нельзя, вероятно, он был усечённым или слабо закруглённым. Тело покрыто циклоидной чешуёй, образующей на боках 49 косых ряда, начинающихся от заднего края плечевого пояса. Предорсальная область в передней половине абсолютно голая, в задней покрыта округлой, врастающей в кожу чешуёй, образующей пятновидное скопление по срединно-спинной линии (рис. 8б). Поры боковой линии на её восходящем участке (до начала мягкой части D) открываются на хорошо развитых дорсальных и вентральных ответвлениях (самые передние из них наиболее длинные); затылочная комиссура с тремя длинными задними ветвями, оканчивающимися заметно позади линии, соединяющей вершины внутренних супраклейтральных шипов соседних сторон. Канал боковой линии под мягкой частью D идёт непосредственно под основаниями плавниковых лучей. Грудь, брюхо, основание Р и бока тела сразу позади основания Р голые. Брюхо с многочисленными тонкими продольными кожными гребешками.
Окраска. Голотип сильно выцвел, сохранилась лишь тёмная пигментация верхней части Р (пропадающая к основанию плавника) и дистальной части С. Перепонка между первым–четвёртым лучами колючей части D интенсивно-чёрная, основание первого луча белое. Нижняя половина боков тела и плавники с очень мелкой точечной меланофорной пигментацией, наиболее выраженной на мягкой части D и на А.
Измерения см. в таблице.
Этимология. Название вида от латинских “oculus” и “cirrhus” – глаз и усик, по характерному признаку нового вида – наличию надглазничного усика.
Сравнение. Новый вид наиболее сходен с U. guttatus Cuvier, 1829 (рис. 9), также описанным из вод Индии. Оба вида отличаются от прочих представителей рода, имеющих одну пару базиптеригиальных отростков и развитые супраклейтральные шипы, степенью развития чешуи на спине между боковыми линиями. Только у этих двух видов чешуя между боковой линией и колючей частью D образует регулярные ряды, тогда как в предорсальной области либо полностью отсутствует (U. guttatus), либо представлена нерегулярными чешуйками (U. oculocirrhis sp. nov.) (рис. 8б, 9а, 9в). У остальных видов чешуя либо хорошо развита между боковыми линиями, включая предорсальную область (хотя может и не доходить до заднего края черепа), либо совсем отсутствует или представлена единичными вросшими чешуйками. Как вариация, чешуя может быть в сходной степени развита только у U. kishimotoi, однако этот вид, имеющий три преоперкулярных шипа, трубковидную заднюю ноздрю и выемчатый верхнезадний край Р, ничего общего с описываемым не имеет. Помимо наличия чешуи в предорсальной области новый вид отличается от U. guttatus наличием супраокулярного усика, отсутствием затылочных долей, длинным прелингвальным выростом (у U. guttatus он не достигает верхней губы: рис. 9г), отсутствием белых пятен на дорсальной поверхности тела (у голотипа U. guttatus, хранящегося, по меньшей мере, с начала XIX века, они до сих пор прослеживаются: рис. 9в) и, возможно, меньшим числом поперечных рядов чешуй (49 против 55). От видов U. affinis-complex, у которых наблюдается изменчивость в степени развития чешуйного покрова в предорсальной области (но в её задней части чешуя всегда перекрывается в отличие от нового вида), U. oculocirrhis sp. nov. отличается наличием супраокулярного усика, отсутствием затылочных долей и однорядными нижнечелюстными зубами (против не менее двух рядов в передней половине челюсти). Кроме того, у красноморских видов данного комплекса (U. dahlakensis и U. marisrubri) короткий прелингвальный вырост присутствует только на ранних стадиях онтогенеза (Brüss, 1987a, 1987b).
Видовые группы в составе Uranoscopus
В составе рода Uranoscopus в настоящее время описано 43 номинальных вида, из которых 27 считаются валидными (Fricke et al., 2021). Очевидно, что при таком разнообразии внутри рода можно выделить отдельные совокупности видов, более близких друг другу, чем к другим подобным совокупностям. Выделение таких совокупностей (в данном случае они называются “группами видов”) позволяет упорядочить классификацию больших по объему родов и использовать более дробную систему в филогенетических и зоогеографических построениях. Путем ранжирования таксономически значимых признаков, использованных для составления приводимой ниже определительной таблицы, виды рода Uranoscopus могут быть объединены в следующие группы.
Группа видов “cognatus”. Характеризуется наличием двух пар базиптеригиальных отростков (очевидно, аутапоморфия), включает U. batrachops sp. nov. и U. cognatus Cantor, 1849. Ареалы этих видов не установлены, так как ранее они смешивались друг с другом (см. описания видов).
Группа видов “sulphureus”. Характеризуется специфическим строением supraoccipitale, выступающим назад в виде одной крупной лопасти (рис. 10а), и отсутствием преорбитальных и супраклейтральных шипов. По-видимому, монофилетическая линия, включающая три вида (U. fuscomaculatus, U. rosette и U. sulphureus) из Красного моря, вод Индо-Австралийского архипелага и Океании.
Рис. 10.
Uranoscopus spp.: а – U. sulphureus, голотип MNHN 0000-5152, голова и плечевой пояс сверху; б–е – рисунок дорсальной стороны туловища (б – U. affinis, ЗИН № 22846, Нагасаки; в, г – U. bicinctus, ИО РАН б/№, Нячанг; д – U. kishimotoi, ЗИН № 38218, Тонкинский зал.; е – U. japonicus, ЗИН № 22844, Нагасаки); ж – U. archionema, ИО РАН б/№, Сейшельские о-ва, голова и предорсальная область дорсолатерально. lac – lacrimale, scl – supracleithrum, soc – supraoccipitale. Масштаб, мм: а, б–г, е – 20; д, ж – 10.

Группа видов “affinis”. Чешуя на спине имеется как между боковыми линиями, так и перед началом D, в промежутке между боковой линией и основанием колючей части D образует правильные ряды. Ранее такие виды выделялись в отдельный род Zalescopus Jordan et Hubbs, 1925 (Jordan, Hubbs, 1925; Линдберг, Красюкова, 1969), сведённый Меесом (Mees, 1960) в синонимы Uranoscopus. Виды группы “affinis” распадаются на две совокупности (подгруппы). Одну из них составляют виды, имеющие выемку верхнезаднего края Р (U. albesca Regan, 1915, U. arafurensis Prokofiev, 2020, U. bauchotae Brüss, 1986, U. brunneus Fricke, 2018 и U. dollfusi Brüss, 1986). Объём этой подгруппы соответствует группе видов “albesca” в понимании Фрике (Fricke, 2018), за исключением U. kishimotoi Fricke, 2018, принадлежащего к группе видов “japonicus”. Среди этих видов резко выделяется U. brunneus, характеризующийся меньшим числом лучей А, бóльшим числом шипов praeoperculum, внутреннего края supracleithrum и поперечных рядов чешуй на боках тела, антеромедиальным выступом lacrimale, переходящим в отчётливый шип (Fricke, 2018. Fig. 2B), тогда как у других видов он лишь выступает в виде валика или бугорка перед глазом, а также наличием тёмного червевидного рисунка на дорсальной поверхности тела (у остальных видов она однотонная). Виды подгруппы “albesca” имеют дизъюнктивный ареал – один вид (U. albesca) обитает в водах Западной Африки, два вида (U. bauchotae и U. dollfusi) известны из Красного моря (U. dollfusi проникает в Оманский и Персидский зал.) и два (U. arafurensis и U. brunneus) – из восточной части Индо-Австралийского архипелага.
Вторую подгруппу составляют четыре вида, характеризующиеся прямым верхнезадним краем Р и хорошо развитой чешуёй в предорсальной области перед началом D. Различия между этими видами не вполне ясны. Красноморские эндемики (U. dahlakensis и U. marisrubri) легко могут быть отличены друг от друга по признакам, приводимым Брюссом (Brüss, 1986, 1987a, 1987b), наиболее существенные из которых суммированы в нижеприведенной определительной таблице, однако в надёжности этих признаков возникают сомнения при учёте изменчивости, наблюдаемой у широко распространенного индо-вест-пацифического вида U. affinis. Единственным отличием обоих красноморских видов от U. affinis, не подверженным изменчивости среди всех морфотипов последнего, является редукция у взрослых особей красноморских видов прелингвального выроста. Степень развития чешуи в предорсальной области, пропорции тела и детали скульптуры головы у U. affinis сильно варьируют, у отдельных ювенильных особей SL < 40 мм чешуя на спине между боковыми линиями вообще не прослеживается. У молоди U. affinis на спине различим густой сетчатый тёмный рисунок, иногда сохраняющийся и у взрослых рыб (ЗИН № 22846) (рис. 10б). Столь значительная изменчивость, не имеющая аналогов среди других видов рода, не позволяет исключить сборной природы вида U. affinis в его настоящем понимании. Типовой территорией (loc. typ.) U. affinis является Индийский океан (“mer des Indes” − Cuvier, 1829). Номинальные виды U. tosae (Jordan et Hubbs, 1925) (loc. typ.: зал. Тоса, Япония) и U. terraereginae Ogilby, 1910 (loc. typ.: Квинсленд, Австралия) рассматриваются качестве младших синонимов U. affinis (Kishimoto, 2001). Сюда же следует отнести номинальный вид U. satsumae (Jordan et Hubbs, 1925) (loc. typ.: Кагосима, Япония). Нужно отметить, что в последнее время некоторые авторы (Fricke et al., 2013; Vilasri, 2013; Fricke, 2018) употребляют название U. tosae в качестве валидного, не приводя для этого никаких оснований. У меня не получилось разделить индо- и западнотихоокеанские популяции, так как все варианты изменчивости были прослежены на материале из Южно-Китайского моря33. С другой стороны, нельзя исключить возможности перекрывания ареалов нескольких близких видов, в настоящее время объединяемых под названием U. affinis sensu Kishimoto (2001), в Западной Пацифике. Для решения этого вопроса необходимо изучение материала со всего ареала данного вида. В этой связи интересно отметить, что голотип U. tosae имеет предорсальную область, полностью покрытую чешуёй, тогда как у голотипа U. satsumae она не доходит до заднего края черепа.
Четвёртый вид комплекса, U. kaianus, характеризуется белопятнистым рисунком дорсальной поверхности тела, чем он напоминает виды следующих двух групп. Однако пятнышки у этого вида неправильной формы, большей частью представляют собой участки белого пигмента разнообразной конфигурации (Günther, 1880. Pl. XIX. Fig. A ). Кишимото (Kishimoto, 2001) указывает на то, что у части особей белопятнистый рисунок может заменяться на сетчатый и такие экземпляры трудноотличимы от U. affinis. По другим признакам U. kaianus очень близок к U. affinis (Kishimoto, 1984).
Группа видов “guttatus”. Эта группа представлена видами U. guttatus и U. oculocirrhis sp. nov., характеризующимися редукцией или полным отсутствием чешуи перед началом D при её нормальном развитии под основанием колючей части этого плавника. Окраска U. guttatus характеризуется наличием на дорсальной поверхности тела белых пятен на тёмном фоне, что сближает этот вид с центральными видами группы “japonicus” (прижизненная окраска U. oculocirrhis sp. nov. не известна, у фиксированного экземпляра какого-либо рисунка не прослеживается). Виды ограничены в распространении северной частью Индийского океана. Группа “guttatus” является морфологически промежуточной между группами “affinis” и “japonicus”, а если описанный выше белопятнистый тип окраски возник в эволюции однократно, её правильнее будет объединить с группой “japonicus”.
Группа видов “japonicus”. Характеризуется целиком бесчешуйным пространством между боковыми линиями или сохранением лишь единичных вросших чешуй в задней части участка между боковой линией и колючей частью D и включает большинство оставшихся индо-тихоокеанских (U. alius sp. nov., U. archionema Regan, 1921, U. bicinctus Temminck et Schlegel, 1843, U. crassiceps Alcock, 1890, U. japonicus Houttuyn, 1782, U. kishimotoi Fricke, 2018, U. oligolepis Bleeker, 1879 и U. marmoratus Cuvier, 1829) и два восточноатлантических вида (U. cadenati Poll, 1959 и U. polli Cadenat, 1951). Большинство составляющих группу видов имеют рисунок из крупных белых пятен на дорсальной стороне тела. Полностью однотонная окраска верха свойственна только U. alius sp. nov. У двух видов (U. bicinctus и U. kishimotoi) расчленяющий рисунок верха из тёмного и светлого компонентов не образует чётко оформленных белых пятен. U. bicinctus морфологически хорошо обособлен от остальных и по окраске не имеет аналогов в пределах рода. Этот вид характеризуется крупнопетлистым тёмным рисунком на светлом фоне в предорсальной области (рис. 10в). Однако у части крупных особей тёмный рисунок становится настолько сильно выраженным, что замещает собой фон, а остатки светлого фона преобразуются в округлые пятна (рис. 10г). У U. kishimotoi тёмный и светлый компоненты образуют червевидный рисунок, который сначала очень похож на рисунок соразмерной молоди U. affinis (SL < 40 мм), а с ростом преобразуется в сетчато-пятнистый, где светлый компонент начинает преобладать и приобретает вид нерегулярных белых пятен в узкой сети тёмного пигмента (рис. 10д). Преобразования рисунка, наблюдаемые у U. bicinctus и U. kishimotoi, можно рассматривать как промежуточный путь к формированию белопятнистого рисунка (рис. 10е, 10ж). Однако в онтогенезе белопятнистых видов этот этап выявить не удалось. Онтогенетическая изменчивость белопятнистого рисунка, прослеженная на материале по U. archionema и U. oligolepis, показывает, что у обоих видов молодь имеет более крупные пятна, чем взрослые рыбы, тогда как густота их распределения варьирует индивидуально. Означает ли это, что белопятнистый рисунок возникал в эволюции разными путями неоднократно или промежуточный этап попросту выпадает в онтогенезе указанных видов, неясно.
В целом следует отметить, что предложенное разделение видов рода Uranoscopus на группы отчасти формально. Монофилия групп “cognatus” и “sulphureus” сомнений не вызывает, однако насколько естественными являются группы видов “affinis”, “guttatus” и “japonicus” можно будет сказать только после нахождения дополнительных признаков, подтверждающих их монофилию. Редукция чешуйного покрова на спине между ветвями боковой линии могла возникать неоднократно, и количественное выражение степени редукции чешуи в предорсальной области варьирует если не в пределах вида (с учётом того, что U. affinis sensu Kishimoto (2001) может представлять собой комплекс близких видов), то в пределах группы точно. Особенности рисунка дорсальной стороны тела, возможно, связаны с филогенезом, но этот признак представляется слишком пластичным, чтобы реконструировать филогению только по нему. Однако в совокупности эти два признака позволяют достаточно уверенно распределить известные виды, поэтому для практических целей выделение групп “affinis”, “guttatus” и “japonicus” представляется обоснованным. Виды U. filibarbis Cuvier, 1829, U. turbisquamatus (Okamura et Kishimoto, 1993) и атлантический U. scaber Linnaeus, 1758 занимают обособленное положение и не могут быть отнесены к какой-либо из выделенных выше групп.
Единственной попыткой установить филогенетические отношения между видами рода Uranoscopus на основании, прежде всего, остеологических и миологических признаков является анализ Виласри (Vilasri, 2013). Однако он смог изучить только 12 видов рода (U. albesca, U. archionema, U. bicinctus, U. cognatus, U. crassiceps, U. filibarbis, U. japonicus, “U. oligolepis”44, U. polli, U. scaber, U. tosae и U. turbisquamatus), а из 30 признаков, на которых основана его кладограмма (Vilasri, 2013. Fig. 76 ), 24 являются гомоплазиями, мозаично распределёнными среди видов других родов семейства, либо трактуются как реверсии; три признака являются аутапоморфиями отдельных видов (U. archionema, U. cognatus, U. filibarbis) и ничего не говорят об их родстве, а два признака (положение верхнего края жаберной щели относительно supracleithrum (признак 99) и наличие/отсутствие прелингвального выроста (признак 112)), по моему мнению, не являются пригодными. Первый из этих признаков мозаично распределён в пределах семейства, а среди Uranoscopus узкие жаберные щели, не доходящие до supracleithrum, якобы характеризуют вид U. cognatus. Однако мне не удалось найти различий по этому признаку между исследованными экземплярами группы «cognatus» и другими видами рода. Наличие или отсутствие прелингвального выроста настолько мозаично распределено в пределах рода, что оба состояния признака, вполне вероятно, могут возникать в одной и той же филогенетической линии. Выделение двух главных филетических линий Uranoscopus, по Виласри (узел 16 на его кладограмме), основывается на двух гомоплазиях (одна из которых редуктивная) и одной реверсии, что вряд ли можно считать убедительным. На кладограмме (Vilasri, 2013. Fig. 76 ) виды U. japonicus и U. polli оказываются в одной кладе с U. albesca и U. tosae и уклоняющимися видами U. cognatus и U. turbisquamatus, противопоставляясь кладе, включающей другие виды группы “japonicus” (U. archionema, U. bicinctus, U. crassiceps, “U. oligolepis”?) и уклоняющийся вид U. filibarbis. При этом два чрезвычайно близких между собой и трудноразличимых вида, возможно, представляющие собой северную и южную форму одного вида (U. crassiceps и U. archionema), на кладограмме оказываются разделёнными совершенно несхожим видом U. filibarbis. Ни одно из промежуточных ветвлений древа Uranoscopus у Виласри не поддерживается аутапоморфиями. В связи с этим схема родственных отношений в пределах рода Uranoscopus, предложенная Виласри, представляется мне чрезвычайно дискуссионной.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ИНДО-ПАЦИФИЧЕСКИХ ВИДОВ РОДА URANOSCOPUS
1(4) Базиптеригиальных шипов две пары. Задняя ноздря в виде полутрубки, у которой задняя стенка заметно выше передней (рис. 4а) [спина между боковыми линиями абсолютно голая; чешуя ниже боковой линии образует косые ряды на всем протяжении] ………………………………………...2
2(3) Ширина межглазничной ямки в 4.4 раза меньше длины; косых рядов чешуй на боках тела около 65; надглазничный усик отсутствует; от затылочной комиссуры отходят шесть задних ветвей; прелингвальный вырост короткий, треугольный, с нитевидно оттянутой вершиной ….........................................…. U. batrachops sp. nov.
3(2) Ширина межглазничной ямки в 2.5–3.4 раза меньше длины; косых рядов чешуй на боках тела 54–57; надглазничный усик имеется; от затылочной комиссуры отходят три задние ветви; прелингвальный вырост лентовидный ….................................................………… U. cognatus
4(1) Базиптеригиальных шипов одна пара; если задняя ноздря трубковидная, то её передняя и задняя стенки равной высоты (рис. 4б)………….. 5
5(8) Чешуя на боках тела не образует косых рядов, а если они имеются, то только в передней половине тела, тогда чешуя вперёд не доходит до уровня вершины клейтрального шипа [спина между боковыми линиями голая; на внутреннем крае supracleithrum один острый шип] …………… 6
6(7) Чешуя не образует косых рядов на всём протяжении; субоперкулярный шип отсутствует; на нижней челюсти нет усика; тело максимально высокое у начала D, постепенно понижается к основанию С, его максимальная высота 3.4–4.8 раза в SL …… ……...............……………….U. turbisquamatus
7(6) Чешуя образует косые ряды в передней части тела, нарушающиеся перед вертикалью середины длины основания А и дорсолатерально; субоперкулярный шип хорошо развит; на симфизе нижней челюсти имеется усик; тело прогонистое, практически на всем протяжении одинаковой высоты (по облику напоминает звездочётов рода Xenocephalus), его максимальная высота 4.8 раза в SL .....................................................…. U. filibarbis
8(5) Чешуя на боках тела образует косые ряды на всём или почти на всём протяжении (лишь непосредственно близ основания С они иногда могут быть спутаны); вперёд чешуйные ряды доходят или не доходят до заднего края плечевого пояса, но всегда заходят за вертикаль вершины клейтрального шипа …………………..........…………. 9
9(14) Задний край крыши черепа (supraoccipitale) выступает назад как крупная, широко закруглённая лопасть, парные затылочные доли не развиты (рис. 10а); передненижний край lacrimale образует тупой выступ или вовсе без него; внутренний край supracleithrum имеет вид закруглённой лопасти или туповершинного выступа, задний конец supracleithrum без отчётливого шипа, самое большее в виде небольшого тупого выступа (рис. 10а); на голове и теле имеются чёткие тёмные пятна [спина между боковыми линиями голая; прелингвальный вырост лентовидный] ……….................……………. 10
10(11) Прелингвальный вырост без нитевидных филаментов в основании; тело более прогонистое, его максимальная высота около 4.5 раза в SL [пятна на боках тела не образуют кольцевидных структур] …………………………. U. fuscomaculatus
11(10) Прелингвальный вырост с многочисленными длинными нитевидными филаментами в основании; тело более высокое, его максимальная высота менее 4.1 раза в SL..………………….…. 12
12(13) Пятна на боках тела не образуют кольцевидных структур; на нижней челюсти только один ряд сильных клыковидных зубов; от восточной Индонезии до о-вов Самоа и Гавайских.……………………………................... U. sulphureus
13(12) Пятна на боках тела собраны в розетковидные кольца; близ симфиза нижней челюсти зубы в два ряда, далее назад – в один; Красное море....……………………………………………… U. rosette
14(9) Задний край крыши черепа более или менее прямой или образует парные затылочные лопасти (рис. 1в); передненижний край lacrimale с отчётливыми шипами; внутренний край supracleithrum с одним или несколькими шипами, задний конец – с острым шипом; на теле не бывает обособленных тёмных пятен ………………………. 15
15(30) Предорсальная область по средней линии полностью покрыта чешуёй хотя бы в задней половине (у молоди U. affinis при SL менее 40 мм может совсем отсутствовать (соразмерные экземпляры других видов мне не известны, а в литературе не описаны), тогда прелингвальный вырост лентовидный, с бахромой по краям на всём протяжении; преоперкулярных шипов более трёх, на спине нет округлых светлых пятен, затылочные доли хорошо развиты), между боковой линией и основанием колючей части D чешуя всегда имеется; надглазничный усик всегда отсутствует ……..….…..... 16
16(17) В А 12 лучей; косых рядов чешуй на теле 62; восемь шипов на praeoperculum и пять – на внутреннем крае supracleithrum; перед наибольшим преорбитальным шипом два хорошо сформированных шипа (Fricke, 2018. Fig. 2B); Р однотонно тёмно-коричневые [верхнезадний край Р с выемкой; верхнебоковая поверхность тела с тёмным червевидным рисунком] ….…….... U. brunneus
17(16) В А 13–14 лучей; косых рядов чешуй на теле 40–60 (если более 55, то верхнезадний край Р прямой); четыре–шесть шипов на praeoperculum и от одного до трёх – на внутреннем крае supracleithrum; перед наибольшим преорбитальным шипом только один отчётливый шиповидный выступ (Прокофьев, 2020. Рис. 2б); Р двухцветные или светлые (до светло-коричневых) …….......………. 18
18(23) Верхнезадний край Р с выемкой; затылочные доли слабо или совсем не выражены …..……… 19
19(20) Предорсальная область сплошь (до заднего края черепа) покрыта хорошо развитой чешуёй; Р двухцветные, их нижняя половина пигментирована гораздо сильнее верхней [колючая и мягкая части D разделены промежутком; прелингвальный вырост в 1.2 раза короче глаза, дистально расширен и на вершине усечён]; Арафурское море .……..U. arafurensis
20(19) Чешуя в передней половине предорсальной области редуцирована; Р однотонные, от прозрачных до светло-коричневых; Красное море и (U. dollfusi) Оманский и Персидский зал.…….. 21
21(22) Колючая и мягкая части D соединены; прелингвальный выступ широкий и короткий, соизмерим с диаметром глаза; ширина головы 30.5–33.5% SL ....................................….. U. dollfusi
22(21) Колючая и мягкая части D разделены; прелингвальный выступ длиннее, около трети длины головы; ширина головы 28.5–30.0% SL …...........................................................U. bauchotae
23(18) Верхнезадний край Р прямой; затылочные доли обычно хорошо развиты (уменьшаются с ростом; не описаны для U. kaianus) (U. affinis-complex: см. “Обсуждение”)…………………………. 24
24(27) Вне Красного моря; колючая и мягкая части D разобщены или соприкасаются; прелингвальный вырост длинный, лентовидный, с бахромой по краям, хорошо развит у всех возрастов .............. 25
25(26) Верх и бока при SL более 60 мм, как правило, коричневатые, однотонные, у молоди часто с тонким тёмным червевидным рисунком, редко присутствующим и у крупных рыб (SL 125 мм (рис. 10б)); широко распространён в Индо-Вест-Пацифике....……………………………………… U. affinis
26(25) Верхнебоковая поверхность тела в рассеянных белых пятнышках, изредка замещающихся тонким сетчатым рисунком (Kishimoto, 2001); Индо-Австралийский архипелаг ….............…U. kaianus
27(25) Красноморские эндемики; колючая и мягкая части D всегда разобщены; прелингвальный вырост выражен только у молоди ………….. 28
28(29) Чешуя на спине между боковыми линиями столь же крупная, как и ниже их, организована в ряды; верх тела однотонный …. U. dahlakensis
29(28) Чешуя на спине между боковыми линиями более мелкая, чем на боках тела и ниже их, не образует правильных рядов; верх тела с сетчатым рисунком ………. …..……………………….U. marisrubri
30(15) Чешуя перед началом D, как правило, полностью отсутствует; если участок чешуи всё же имеется, то присутствует надглазничный усик; чешуя между боковой линией и основанием колючей части D имеется или отсутствует ………... 31
31(32) Имеется участок чешуи перед началом D, чешуя между основанием колючей части D и боковой линией хорошо развита; верхний край глазного яблока с усиком; задняя ноздря простая; прелингвальный вырост длинный, лентовидный, с короткой бахромой в основании, достигает заднего края черепа …………….... U. oculocirrhis sp. nov.
32(31) Чешуя перед началом D полностью отсутствует; нет надглазничного усика (как вариация, зачаточный усик отмечен у молоди U. archionema); прелингвальный вырост обычно короткий или совсем отсутствует, а если длинный и лентовидный (у U. bicinctus и молоди U. oligolepis), то чешуя между основанием колючей части D и боковой линией редуцирована или отсутствует (у молоди U. oligolepis, кроме того, задняя ноздря трубковидная, а бахрома в основании прелингвального выроста часто с длинными выростами) .…...…….. 33
33(34) Чешуя между боковой линией и основанием колючей части D хорошо развита, столь же крупная, как и на боках тела, образует правильные ряды; надглазничный гребень образует у верхнезаднего края орбиты небольшой выступ (рис. 9б) [затылочные доли хорошо развиты; преоперкулярных шипов четыре или пять; задняя ноздря простая; прелингвальный вырост (рис. 9г) короткий, в основании широкий, дистально лентовидный, достигает только края нижней челюсти; прижизненная окраска верхней поверхности туловища представлена округлыми белыми пятнами на тёмном фоне] ....………………….. U. guttatus
34(33) Чешуя между боковой линией и основанием колючей части D полностью отсутствует или единичная, редуцированная, врастающая в кожу (наиболее развита у U. kishimotoi, отличающегося трубковидной задней ноздрей и тремя преоперкулярными шипами); надглазничный гребень не образует выступа у верхнезаднего края орбиты (кроме U. marmoratus; как вариация, может быть обнаружен у молоди U. archionema) ………………..35
35(36) На теле два крупных седловидных тёмных пятна под колючим и задней частью мягкого отделов D, сохраняющихся даже на старых и выцветших коллекционных экземплярах; предорсальная область с крупнопетлистым тёмным сетчатым рисунком; прелингвальный вырост длинный, лентовидный, непигментированный (при жизни ярко-оранжевый); задняя ноздря простая ...........… U. bicinctus
36(35) На теле никогда не бывает поперечных седловидных пятен, а на спине перед D – крупнопетлистого рисунка; если прелингвальный вырост длинный (у молоди U. oligolepis), то он пигментирован, а задняя ноздря трубковидная ….……….…….. 37
37(40) Три преоперкулярных шипа (как очень редкое исключение у U. japonicus отмечено четыре); верхнезадний край Р с выемкой [окраска верха тела никогда не бывает однотонной] ………….…………. 38
38(39) Задняя ноздря щелевидная; прелингвальный вырост у молоди в виде широкого пластинчатого придатка с бахромой по краям (Kishimoto, 1987. Figs. 3D, 3E), у взрослых – самое большее в виде маленького треугольного выступа; чешуя выше боковой линии совершенно отсутствует или единичная; окраска верха представлена округлыми белыми пятнами или нерегулярным белопятнисто-мраморным рисунком, тёмные участки достаточно широкие, чтобы рисунок воспринимался как светлый на тёмном фоне (рис. 10е); позвонков 26, изредка 27 …..………..……………. U. japonicus
39(38) Задняя ноздря трубковидная; прелингвальный вырост короткий или отсутствует у особей всех возрастов; чешуи выше боковой линии сравнительно многочисленные, но вросшие в кожу и различимые с трудом; в окраске верха белый цвет преобладает над тёмным, так что она воспринимается как тонкий червевидный тёмный рисунок на белом фоне (рис. 10д); позвонков 25 .…U. kishimotoi
40(37) Преоперкулярных шипов четыре или пять, редко шесть (как исключение, семь у U. crassiceps: Fricke et al., 2013); верхнезадний край Р прямой ................................……………….. 41
41(42) Задняя ноздря трубковидная; прелингвальный вырост у молоди длинный, заходит за глаз, с ростом значительно сокращается в длине, но сохраняет лентовидную форму (Kishimoto, 1987. Figs. 3A–3C) [верх тела в округлых или неправильной формы белых пятнах, иногда частично сливающихся или приобретающих розетковидную форму, промежутки между пятнами не превышают размера пятен] …........…… U. oligolepis
42(41) Задняя ноздря в виде простой поры; прелингвальный вырост, если лентовидный (у молоди U. archionema), то не достигает глаза, обычно зачаточный или отсутствует у особей всех возрастов …….……....……………………………….…… 43
43(44) Затылочные доли хорошо развиты, конические [надвисочные гребни отчётливо выражены; имеются развитые заглазничные бугорки (рис. 7а); верх тела в белых пятнах на тёмном фоне] ……..………………………….…………. U. marmoratus
44(43) Затылочные доли зачаточные или отсутствуют …………………….................................…….. 45
45(46) Надвисочные гребни низкие, но отчётливые; супраорбитальные гребни соседних сторон сзади соединяются поперечным посторбитальным гребнем (рис. 6в, 6г); нет бугорков или выступов у задневерхнего края орбиты; боковые стороны pelvis расходящиеся к вершине (рис. 6ж); прижизненная окраска верха однотонная (Kishimoto, 2001), у сильно выцветших фиксированных рыб меланофоры концентрируются в нижних двух третях боков тела, на Р присутствуют только в его базальной трети [диаметр глаза 6.1 раза укладывается в длине головы, межглазничный промежуток – 0.78 раза в диаметре глаза] …………...U. alius sp. nov.
46(45) Надвисочные и поперечный посторбитальный гребни не развиты (рис. 10ж); внутренний край infraorbitalia-3 и/или 4 нередко образует небольшой или едва намеченный лопастевидный выступ, но развитых заглазничных бугорков нет; боковые стороны pelvis параллельны друг другу; прижизненная окраска верха с крупными округлыми белыми пятнами, хорошо сохраняющимися у фиксированных рыб (рис. 10ж); у сильно выцветших экземпляров меланофоры равномерно распределены на боках тела, на Р присутствуют на всём его протяжении (у U. archionema; достоверно не установлено для U. crassiceps, но, судя по прижизненной окраске, у этого вида меланофоры должны концентрироваться в дорсальной половине плавника) .…......................................……… 47
47(48) Диаметр глаза 5.4–6.8 (как правило, более 5.5) раза укладывается в длине головы, ширина межглазничного промежутка – 0.59–0.75 раза в диаметре глаза; Р в дорсальной половине тёмный, в вентральной – светлый (при жизни – жёлтый); у берегов Индии и Аравийского п-ва …..…. U. crassiceps
48(47) Диаметр глаза 4.0–6.2 (как правило, менее 5.5) раза укладывается в длине головы, ширина межглазничного промежутка – 0.8–1.0 раза в диаметре глаза; Р равномерно тёмный со светлым дистальным краем; у берегов Восточной Африки и прилежащих островов и на подводных поднятиях .….....……. U. archionema
Список литературы
Линдберг Г.У., Красюкова З.В. 1969. Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Ч. 3. Teleostomi. XXIX. Perciformes. 1. Percoidei (XC. Сем. Serranidae – CXLIV. Сем. Champsodontidae). Л.: Наука, 479 с.
Прокофьев А.М. 2020. Новый вид звездочётов (Uranoscopidae) из Арафурского моря // Вопр. ихтиологии. Т. 60. № 5. С. 499–506. https://doi.org/10.31857/S0042875220050094
De Beaufort L.F., Chapman W.M. 1951. The fishes of the Indo-Australian Archipelago. IX. Percomorphi (concluded), Blennoidea. Leiden: A.J. Reprints Agency, i–xi + 484 p.
Bleeker P. 1853. Nalezingen op de ichthyologie van Japan // Verh. Bat. Gen. V. 25. P. 1–56.
Bleeker P. 1879. Révision des espèces insulindiennes du genre Uranoscopus L. // Versl. Akad. Amsterdam. Ser. 2. V. 13. [1878]. P. 47–59
Brüss R. 1986. Two new species of Uranoscopus Linnaeus, 1758, from the Red Sea: U. dollfusi n. sp. and U. bauchotae n. sp. // Bull. Mus. Natn. Hist. Nat. Ser. 4. V. 8. Sect. A. № 4. P. 955–967 .
Brüss R. 1987a. XIII. Uranoscopus marisrubri n. sp. aus dem zentralen und nördlichen Roten Meer (Pisces: Perciformes: Uranoscopidae) // Senckenberg. Biol. Bd. 68. Hf. 1/3. S. 38–48.
Brüss R. 1987b. Uranoscopus dahlakensis n. sp. und Neunachweis vom U. scaber Linnaeus 1758 im Roten Meer (Pisces: Perciformes: Uranoscopidae) // Ibid. Bd. 68. Hf. 1/3. S. 49–57.
Cantor T.E. 1849. Catalogue of Malayan fishes // J. Asiatic Soc. Bengal. V. 18. Pt. 2. P. i–xii + 983–1443.
Cuvier G. 1829. Le Règne Animal, distribué d’après son organisation, pour servir de base à l’histoire naturelle des animaux et d’introduction à l’anatomie comparée. V. 2. Paris: Chez Déterville, i–xv + 1–406 pp.
Fricke R. 2018. Two new species of stargazers of the genus Uranoscopus (Teleostei: Uranoscopidae) from the western Pacific Ocean // Zootaxa. V. 4476. № 1. P. 157–167.
Fricke R., Eschmeyer W.N., van der Laan R. 2021. Catalog of Fishes: Genera, Species, References. Available (http:// researcharchive.calacademy.org/research/ichthyology/catalog/
fishcatmain.asp. Version 01/2021).
Fricke R., Jawad L.A., Al-Kharusi L.H., Al-Mamry J.M. 2013. New record and redescription of Uranoscopus crassiceps Alcock, 1890 (Uranoscopidae) from Oman, Arabian Sea, northwestern Indian Ocean, based on adult specimens // Cybium. V. 37. № 3. P. 143–147.
Gan H.M., Syahadah M.Y.N.I., Vilasri V. et al. 2019. Four whole mitogenome sequences of yellowtail stargazers (Uranoscopus cognatus Cantor 1849) from East Peninsular Malaysia and West Coast of Thailand // Mitochondrial DNA. Part B. V. 4. № 1. P. 256–258. https://doi.org/10.1080/23802359.2018.1536473
Günther A. 1880. Report on the shore fishes procured during the voyage of H. M. S. Challenger in the years 1873–1876 // Zoology. V. 1. Pt. 6. P. 1–82.
Heemstra P.C. 1984. Uranoscopidae. Stargazers / Eds. Fischer W., Bianchi G. // FAO Spec. Ident. Sheets Fish. Purp. Western Indian Ocean. Fishing area 51. Rome: FAO, no pagination.
Heemstra P.C. 1986. Family no. 230: Uranoscopidae / Eds. Smith M.M., Heemstra P.C. // Smiths’ Sea Fishes. Berlin et al.: Springer-Verlag. P. 735–736.
Jordan D.S., Hubbs C.L. 1925. Record of fishes obtained by David Starr Jordan in Japan, 1922 // Mem. Carnegie Mus. V. 10. № 2. P. 93–346.
Kishimoto H. 1984. Redescription and lectotype designation of the stargazer, Uranoscopus kaianus Günther // Copeia. № 4. P. 1009−1011.
Kishimoto H. 1987. A new stargazer, Uranoscopus flavipinnis, from Japan and Taiwan with redescription and neotype designation of U. japonicus // Jpn. J. Ichthyol. V. 34. № 1. P. 1−14.
Kishimoto H. 2001. Uranoscopidae Stargazers / Eds. Carpenter K.E., Niem V. // FAO Spec. Ident. Guide Fish. Purp. The living marine resources of the Western Central Pacific. V. 6. Bony fishes. Part 4. (Labridae to Latimeriidae), estuarine crocodiles, sea turtles, sea snakes and marine mammals. Rome: FAO. P. 3519−3531.
Mees G.F. 1960. The Uranoscopidae of Western Australia (Pisces, Perciformes) // J. R. Soc. West. Aust. V. 43. Pt. 2. P. 46−58.
Pietsch T.W., Kishimoto H. 1989. Uranoscopus chinensis Guichenot in Sauvage, 1882, a senior synonym of Uranoscopus flavipinnis Kishimoto, 1987 (Teleostei: Uranoscopidae) // Copeia. № 3. P. 748−750.
Randall J.E., Arnold R.J. 2012. Uranoscopus rosette, a new species of stargazer (Uranoscopidae: Trachinoidei) from the Red Sea // Aqua Internat. J. Ichthyol. V. 18. № 4. P. 209−218.
Sauvage H.E. 1882. Description de quelques poissons de la collection du Muséum d’histoire naturelle // Bull. Soc. Philomath. Paris. Ser. 7. V. 6. P. 168−176.
Smith W.L. 2016. Uranoscopidae Stargazers / Eds. Carpenter K.E., De Angelis N. // FAO Spec. Ident. Guide Fish. Purp. The living marine resources of the Eastern Central Atlantic. V. 4. Part 2. Bony fishes (Perciformes to Tetraodontiformes) and sea turtles. Rome: FAO. P. 2786–2792.
Vilasri V. 2013. Comparative anatomy and phylogenetic systematics of the family Uranoscopidae (Actinopterygii: Perciformes) // Mem. Grad. Sci. Fish. Sci. Hokkaido Univ. V. 55. № 1/2. P. 1−106.
Vilasri V., Ho H.-C., Kawai T., Gomon M.F. 2019. A new stargazer, Ichthyscopus pollicaris (Perciformes: Uranoscopidae), from East Asia // Zootaxa. V. 4702. № 1. P. 49–59.
Yamada U. 2002. 303. Uranoscopidae stargazers / Ed. Nakabo T. // Fishes of Japan with pictorial keys to the species. English Edition. Tokyo: Tokai Univ. Press. V. 1. P. 1075–1076.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии