

Вопросы ихтиологии, 2022, T. 62, № 1, стр. 19-37
Морские петухи рода Pterygotrigla (Triglidae) залива Нячанг и сопредельных акваторий (Вьетнам, Южно-Китайское море)
1 Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
2 Институт океанологии РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 17.11.2020
После доработки 07.12.2020
Принята к публикации 07.12.2020
- EDN: NQZUTX
- DOI: 10.31857/S0042875222010143
Аннотация
Приведён обзор видов морских петухов рода Pterygotrigla, встреченных в заливе Нячанг и прилегающей акватории Южно-Китайского моря (юг Центрального Вьетнама). Документировано присутствие четырёх видов из трёх подродов: P. (Otohime) hemisticta, P. (O.) tagala, P. (Parapterygotrigla) multiocellata и P. (s. str.) ryukyuensis. P. multiocellata и P. tagala впервые указаны для фауны Вьетнама. Для каждого вида дано подробное иллюстрированное описание. Рассмотрены вопросы онтогенетической изменчивости ошипления головы и других признаков у представителей рода. Составлен ключ для определения видов Pterygotrigla Южно-Китайского моря.
Морские петухи рода Pterygotrigla Waite, 1899 характеризуются наличием костных щитков (расширений оснований дорсальных птеригиофоров) только под колючей частью спинного плавника, очень мелкой (>65 рядов над боковой линией) циклоидной чешуёй и наличием мелких тёмных пятен на теле (преимущественно в верхней половине) (Richards, Jones, 2002; Richards, Yato, 2012). Вместе с близкими родами Bovitrigla Fowler, 1938 и Uradia Kamohara, 1938 рассматриваемый род составляет трибу Pterygotriglini (Richards, Jones, 2002). В современной фауне известно 27 валидных видов, группируемых в три подрода, распространённых в Индо-Пацифике (Richards et al., 2003; Richards, Yato, 2012, 2014; Yato, Heemstra, 2019), из них семь было отмечено в Южно-Китайском море (Randall, Lim, 2000), но только два вида (P. hemisticta (Temminck et Schlegel, 1843) и P. ryukyuensis Matsubara et Hiyama, 1932) были указаны для фауны Вьетнама (Nguyen, 1999), где род Pterygotrigla до сих пор был отмечен только в Тонкинском заливе.
В ходе траловых съёмок и анализа траловых уловов местного промысла у побережья юга Центрального Вьетнама (заливы Нячанг и Ванфонг) в 2005–2013 гг. мною было собрано четыре вида рода Pterygotrigla, два из которых, хотя уже были указаны для Южно-Китайского моря, впервые отмечены в ихтиофауне страны. Для двух видов удалось собрать молодь, сохраняющую ювенильные черты в ошиплении головы. Ранние стадии развития Pterygotrigla изучены исключительно слабо и в литературе охарактеризованы только для представителей подрода Parapterygotrigla Matsubara, 1937 (Richards, Yato, 2014). Поэтому я изучил все доступные материалы по молоди Pterygotrigla для выяснения таксономической значимости различий в ювенильных чертах строения. Цель настоящей работы – описать виды птериготригл, встреченных в водах Вьетнама, и изучить особенности онтогенетической изменчивости признаков в пределах данного рода.
МАТЕРИАЛ И МЕТОДИКА
Весь исследованный материал хранится в коллекции Института океанологии (ИО РАН), Москва. Промеры выполнены согласно рекомендациям Ричардса с соавторами (Richards, Saksena, 1977; Richards et al., 2003); уклоняющиеся значения, наблюдаемые у отдельных экземпляров, приведены в скобках. “Соединённым Р” именуется часть плавника, образованная соединёнными перепонкой лучами. В плавниковых формулах соединённые и свободные лучи Р подсчитаны раздельно; последний, раздвоенный до основания луч вертикальных плавников указан как “1½”. Номенклатура шипов головы дана по работе Гомона и Каваи (Gomon, Kawai, 2018) со следующим дополнением: шипы на носовых костях (nasalia) называются “носовыми”. Направленные вперёд выросты lacrimale, обычно называемые в англоязычной литературе ростральными отростками (rostral processes), а также ростральными, преорбитальными или передними инфраорбитальными шипами, приводятся как “ростральные шипы”. Посттемпоральные шипы в литературе часто обозначаются как нухальные. Париетальные гребни и шипы у видов рода Pterygotrigla отсутствуют, поэтому в приводимых в работе описаниях о них не упоминается. В работе использованы следующие сокращения: SL – стандартная длина, lc – длина головы, ao – длина рыла, oo – горизонтальный диаметр костной орбиты, io – ширина костного межглазничного промежутка, lr – длина рострального шипа, so – суборбитальная высота, H – максимальная высота тела; lP, lPf, lV – соответственно длина наибольшего соединённого и первого свободного луча грудного плавника и длина брюшного плавника; D1, D2, A, P, V, С – соответственно первый (колючий) и второй (мягкий) спинные, анальный, грудные, брюшные и хвостовой плавники; LL – туловищный канал боковой линии и число чешуй в нём; sp. br. – число развитых жаберных тычинок в наружном ряду на первой жаберной дуге; n – число экземпляров; НИС, НПС – соответственно научно-исследовательское и научно-промысловое судно, ст. – океанографическая станция.
Для анализа межвидовых различий онтогенетической изменчивости дополнительно изучен материал по виду P. (s. str.) picta (Günther, 1880) из юго-восточной части Тихого океана из коллекции ИО РАН: 1 экз. SL 17 мм, 25°40′ ю.ш. 86°34′ з.д., 200–0 м, НПС “Профессор Месяцев”, рейс 13; 1 экз. SL 32.5 мм, хребет Сала-и-Гомес, НПС “Профессор Месяцев”, рейс 15, трал 65; 1 экз. SL 37.5 мм, 22°11′ ю.ш. 81°21′ з.д., 235 м, НПС “Одиссей”, рейс 2/81, трал 11(6); 1 экз. SL 39 мм, 25°03′–25°01′ ю.ш. 97°29′–97°35′ з.д., 290–600 м, НИС “Профессор Штокман”, рейс 18, ст. 1992; 1 экз. SL 55 мм, 25°38′–25°36′ ю.ш. 85°31′–85°33′ з.д., 250–260 м, НИС “Профессор Штокман”, рейс 18, ст. 1929; 3 экз. SL 79–105 мм, 25°34′–25°35′ ю.ш. 85°27′–85°30′ з.д., 240–245 м, НИС “Профессор Штокман”, рейс 18, ст. 1924; 11 экз. SL 59–203 мм, 25°07′–25°07′ ю.ш. 99°44′–99°41′ з.д., 330–350 м, НИС “Профессор Штокман”, рейс 18, ст. 1997; 1 экз. SL 350 мм, без этикетки, сборы АтлантНИРО в Юго-Восточной Пацифике, конец 1980-х гг.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Pterygotrigla (Otohime) hemisticta (Temminck et Schlegel, 1843)
Рис. 1.
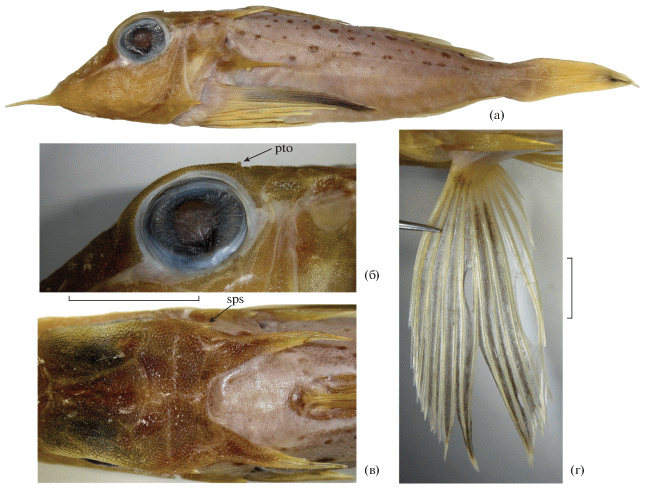
Pterygotrigla (Otohime) hemisticta: а – общий вид экз. SL 235 мм, б – ростральные шипы (левый – с дополнительным зубцом у вершины), в – ошипление предкрышки; г, д – варианты ошипления преоперкулярной лопасти, е – чешуйный покров боков на вертикали начала D2, ж – вариант пигментации боков тела, з – внутренняя сторона Р. OL – преоперкулярная лопасть, pLL – отверстие трубочки боковой линии, pos – верхний преоперкулярный шип, sLL – чешуя боковой линии. Масштаб, мм: б–д – 10, е – 2, ж – 25, з – 15, линейка общая для г и д.

Рис. 2.
Pterygotrigla (Otohime) hemisticta, малёк SL 62 мм: а – общий вид, б – верхний преоперкулярный шип, в – заглазничная часть крыши черепа дорсолатерально, г – D1, д – внутренняя сторона Р; ac.po – дополнительный шип в основании верхнего преоперкулярного шипа, pto – постокулярный шип, sps – сфенотикальный шип. Масштаб, мм: б, в, д – 4, г – 5.

Материал. Всего 4 экз. SL 62 и 225–295 мм, Вьетнам, Нячанг, рыбный рынок (Cang Ca Vinh Luong), май–июнь 2009 г.
Описание. D1 VII, D2 11½, A 11½–12½, P 12 + + 3, V I + 5; LL ~ 60–70; sp. br. 2–3 + 1 + 10–11, кроме того 0–1 плоский бугорок на epibranchiale 1 и три–пять – в нижней части cerato- и на hypobranchiale 1, у крупных рыб по одному бугорку имеется также в каждом промежутке между развитыми жаберными тычинками на ceratobranchiale 1.
Взрослые рыбы SL 225–295 мм (рис. 1а–1з). Ростральные шипы умеренной длины, узкие и плоские, острые, расходящиеся, прямые, мелко зазубрены по наружному краю, часто хотя бы с одной стороны несут дополнительный зубец у вершины с внутреннего края (рис. 1б). Носовые шипы отсутствуют. Praeoperculum с относительно коротким (соизмерим с диаметром зрачка) сильным шипом, являющимся окончанием подглазничной опоры, продолжающимся вперёд в виде очень слабого гребня, не доходящего до переднего края кости, с небольшим шипом под ним (под которым в свою очередь может быть ещё меньший шипик), отделённым выемкой (у наибольшего экземпляра очень пологой) от угловато закруглённой преоперкулярной лопасти, у верхнего и нижнего конца которой иногда развиты маленькие шипики (рис. 1в–1д). Посттемпоральные шипы массивные, треугольные, умеренной длины, их вершины оканчиваются узким острым шипиком на уровне переднего края – середины длины непарной предорсальной костной пластинки. Оперкулярный шип длинный, оканчивается на уровне основания пятого колючего луча D1, острый, мечевидный. Клейтральный шип хорошо выражен, расположен заметно ниже оперкулярного шипа, оканчивается на середине длины последнего. Межглазничный промежуток неглубоко вогнут, верхний край костной орбиты выступающий. Надглазничные гребни не выражены. Заглазничной борозды нет. Гребень sphenoticum не развит. Покровные кости головы и плечевого пояса утолщённые, с признаками пахиостоза, не покрыты кожей, несут очень мелкие бугорки, на одних участках организованные в чёткие продольные ряды, на других – с ячеистым орнаментом между ними.
Нижняя челюсть слегка короче верхней; maxillare расширено на заднем конце, оканчивается на вертикали середины глаза; на челюстях мелкие бугорковидные зубы полосками, озубление praemaxillaria видно снаружи при закрытом рте; сошник и нёбные кости без зубов. Язык короткий, массивный, треугольный, сзади широкий, а в передней части суженный, слабо обособлен. Жаберные тычинки уплощённые, наибольшие из них короче жаберных лепестков, несут крошечные шипики. Ложножабра крупная, из 45–51 элемента. Хорошо развита щель позади четвёртой жаберной дуги.
Перед основанием D1 один непарный костный щиток, под основанием D1 и в интердорсальном промежутке девять пар костных щитков, из которых только первые три (но у наибольшего экземпляра – также четвёртая) выступают из кожи; под основанием D2 щитки отсутствуют, как и в боковой линии. Выступающие наружу щитки несут ямчато-бугорчатый орнамент (но у наибольшего экземпляра бугорки построены в отчётливые ряды), лишены шипов. Четвёртая колючка D1 наиболее длинная и массивная; передний край первой колючки гладкий по всей длине. Первый луч D2 и до шести передних лучей А неветвистые (первый луч А в виде тонкой колючки); последний луч в этих плавниках раздвоен до основания. С глубоко вырезан, его лопасти равной длины, заострены. Р длинные, вершина соединённого Р достигает передней трети длины основания А (его четвёртого–пятого луча) у экз. SL 225 и 235 мм, только немного заходит за начало А у экз. SL 295 мм (до второго луча). Наибольший (верхний) свободный луч Р короче наибольших из соединённых лучей Р, достигает вертикали начала А или слегка заходит за неё (до уровня основания второго луча) у рыб SL 225 и 235 мм, не достигает начала А у наибольшего экземпляра. Начало V под нижним концом основания P, вершины заметно не достигают или почти достигают ануса. Колючка V короче ветвистых лучей, второй–третий, третий или четвёртый ветвистые лучи – наибольшие. Анус расположен у начала А.
Чешуя очень мелкая, округлая, циклоидная, с ровным свободным краем (рис. 1е), слабо налегающая, на груди, перед и под D1 выше LL и, если имеется на основании Р, вросшая в кожу и разобщённая. Чешуя покрывает грудь и брюхо, но отсутствует на вершине истмуса, между основаниями V и позади основания Р, на основании Р чешуя обычно отсутствует, если имеется (иногда только с одной стороны), то единичная. Предорсальная область не покрыта чешуёй. Чешуи боковой линии крупнее прочих чешуй на теле. Боковая линия практически прямая, идёт выше средней линии тела полого вниз, делая очень слабый изгиб у основания хвостового стебля, где она проходит по средней линии до основания лучей С. Трубочки боковой линии открываются назад и вниз.
Измерения, в % SL: lc 38.0–40.0, ao 15.8–16.9, oo 8.9–10.2, io 11.2–11.9, lr 3.4–5.1, so 8.4–8.5, H 22.6–25.4, lP 28.8–34.7, lPf 25.8–30.7, lV 20.8–24.4.
Окраска фиксированных экземпляров розовато-бежевая, вентральная сторона белая; кости головы, дорсальные щитки и колючки D1 желтоватые; верхнебоковая поверхность тела (выше боковой линии) в редко расположенных продолговатых чёрных пятнышках (иногда между ними могут присутствовать извилистые тонкие полоски и мелкие пестринки, формирующие подобие сетчатого рисунка: рис. 1ж); ряд таких пятнышек присутствует по нижнему краю орбиты и между латеральным краем черепа и жаберным отверстием (плохо выражены у самой крупной рыбы). На интеррадиальной перепонке в задней части D1 между четвёртой и шестой колючками (у двух из трёх рыб также на ограниченном участке перепонки перед четвёртой колючкой) имеется крупное, чётко отграниченное чёрное пятно; в остальном перепонка D1 не пигментирована. На D2 продольный медиальный ряд неярких тёмных пятен, расположенных только на плавниковых лучах. Внутренняя поверхность соединённого Р чёрно-серого цвета (лучи светлые), в базальной трети более интенсивно чёрная (лучи не выделяются), с диагональным рядом удлинённых белых пятен (рис. 1з), иногда с нечётко выраженным дополнительным рядом плохо оформленных светлых пятнышек на границе чёрной и чёрно-серой пигментации; перепонка между первым–третьим и десятым–двенадцатым соединёнными лучами белая; дистальный край соединённого Р со светлой каймой. Свободные лучи Р чисто белые. А, V и С не окрашены. Ротожаберная полость светлая. Прижизненная окраска не документирована.
Малёк SL 62 мм (рис. 2а–2д). Все шипы головы заметно более тонкие, узкие и длинные, чем у взрослых рыб, островершинные. Ростральные шипы длинные, их вершины расходятся заметно сильнее, чем у взрослых рыб. Praeoperculum с четырьмя шипами, самый нижний расположен в угловой части, три нижних небольшие, треугольные, равновеликие и равнорасставленные, ориентированы назад и (особенно второй и третий) немного вниз; самый верхний шип (являющийся окончанием подглазничной опоры) оканчивается на вертикали заднего края черепа, направлен вершиной назад, в основании несёт сильный дополнительный шип (рис. 2б), загнутый назад (с левой стороны ещё и отклонённый наружу). Посттемпоральные шипы очень длинные, достигают уровня переднего края второй субдорсальной костной пластинки. Оперкулярный шип длинный, слегка заходит за вертикаль основания третьей колючки D1; клейтральный шип оканчивается чуть позади уровня вершины оперкулярного шипа, его длина составляет около половины длины оперкулярного шипа без гребня (т.е. на отрезке от заднего края operculum до вершины шипа). Межглазничный промежуток глубоко вогнут. Надглазничные гребни хорошо развиты, оканчиваются сильным острым шипом с широким треугольным основанием, направленным назад. Заглазничной борозды нет. Гребень sphenoticum шельфовидный, оканчивается острым шипом (рис. 2в). Челюсти почти равной длины, зубы более тонкие, чем у взрослых рыб, щетинковидные; на сошнике поперечная, полулунно изогнутая полоска мелких зубов; нёбные кости без зубов. Развитые жаберные тычинки длинные и тонкие (длина наибольших из них немного превышает длину наибольших жаберных лепестков), вооружены мелкими шипиками. Ложножабра из 29 элементов.
Только непарный предорсальный и первые три из девяти пар субдорсальных щитков выступают из кожи; выступающие наружу щитки несут бугорки (непарный щиток) или тонкие продольные гребешки без шипов. Передний край первой колючки D1 очень мелко и слабо зазубрен только в базальной половине. Первые два луча D2 и передние пять лучей А неветвистые (пятый луч А с зачатком ветвления на вершине); последний луч в этих плавниках раздвоен до основания. С глубоко вырезан, его лопасти равной длины и слабо удлинены. Р очень длинные, вершина соединённого Р достигает середины длины последней трети основания А (до вертикали основания его девятого луча). Наибольший (верхний) свободный луч Р короче наибольших из соединённых лучей Р, немного не достигает вертикали середины длины основания А (его шестого луча). Конец V едва заходит за начало А, наиболее длинные лучи V со второго по четвёртый (ветвистые). Чешуя покрывает грудь и брюхо, но отсутствует на основании Р, между основаниями Р и V, между основаниями V и в предорсальной области; в передней половине тела, включая грудь и брюхо, сильно вросшая в кожу, в задней – слабо налегающая.
Измерения, в % SL: lc 46.8, ao 19.8, oo 14.5, io 12.9, lr 10.5, so 8.1, H 25.8, lP 40.3, lPf 35.5, lV 27.4.
Окраска принципиально не отличается от таковой у взрослых рыб, но на спинных плавниках меланофорная пигментация более интенсивная. Верхнебоковая поверхность тела в некрупных разноразмерных округлых и удлинённых тёмных пятнышках, в целом более малочисленных, чем у взрослых рыб, но также расположенных преимущественно выше LL и непосредственно на ней; последние, как и несколько пятнышек сразу под LL на уровне основания D1, несколько более крупные; между пятнами имеется меланофорный крап, не различимый невооружённым глазом; ряд тёмных пятнышек присутствует по нижнему краю орбиты и между латеральным краем черепа и жаберным отверстием. В задней части D1 имеется крупное, чётко отграниченное округлое чёрное пятно (рис. 2г), занимающее промежуток от задней половины третьей интеррадиальной мембраны до переднего края шестой колючки. Скопления меланофоров (плохо заметные невооружённым глазом) присутствуют также на первой и второй интеррадиальных мембранах, у дистального края третьей интеррадиальной мембраны и в основании перепонки между двумя последними колючками. На D2 продольный медиальный ряд довольно крупных черноватых пятен, расположенных только на плавниковых лучах. Внутренняя поверхность соединённого Р чёрно-серого цвета (лучи светлее перепонок между ними) с ярко-чёрным пятном в нижней части и диагональным рядом удлинённых белых пятен (рис. 2д); дистальный край соединённого Р со светлой каймой. Свободные лучи Р светлые, с меланофорным крапом, более густым в основаниях лучей.
Замечания. Изученные экземпляры полностью соответствуют признакам вида (Richards et al., 2003). Вид широко распространён в Западной Пацифике от Японии до Австралии и в индоокеанских водах Западной Австралии; указания на поимки в водах Индии (Richards, 1984) и Красного моря (Khalaf et al., 1996) относятся к другим видам (Richards et al., 2003). В водах Вьетнама он ранее указывался только для Тонкинского залива (Nguyen, 1999). В заливе Нячанг, по-видимому, более редок, чем другой встречающийся здесь вид подрода Otohime (P. tagala), что нельзя объяснять особенностями батиметрического распределения, поскольку для P. hemisticta указывается очень широкий диапазон глубин обитания (20–420 м) (Richards et al., 2003). Крупные экземпляры P. hemisticta (из моей коллекции) были пойманы близнецовыми тралами на глубине 100–300 м в присваловой части прибрежья провинции Кханьхоа.
Pterygotrigla (Otohime) tagala (Herre et Kauffman, 1952)
(рис. 3)
Рис. 3.
Pterygotrigla (Otohime) tagala SL 92 мм: а, б – общий вид (а – сбоку, б – снизу), в – внутренняя сторона Р. Масштаб: 5 мм.

Материал. Всего 52 экз.: 36 экз. SL 51–92 мм, Вьетнам, Нячанг, рыбный рынок Бэ (Cho Ca Be), 24.05–19.06.2005 г.; 14 экз. SL 62–89 мм, там же, апрель–июнь 2007 г.; 2 экз. SL 72 и 82 мм, залив Нячанг, 12°10′–12°07′ с.ш. 109°25′–109°26′ в.д., глубина 91–95 м, трал № 1, время лова 09:25–10:45, 31.05.2006 г.
Описание. D1 VII, D2 12½, A 12½, P 13 + 3, V I + 5; LL 55–67 (обычно ~ 60) (n = 20); sp. br. 1–2 + 1 + 8–10, кроме того, 0–1 бугорок на epibranchiale 1 и 4–6 на ceratobranchiale 1; элементов ложножабры 25–32 (n = 15).
Ростральные шипы длинные, узкие и плоские, острые, расходящиеся, слегка изогнуты и зазубрены по наружному краю (рис. 3а–3б). Носовые шипы отсутствуют. Praeoperculum с тремя шипами: самый нижний расположен в угловой части, небольшой, но всегда хорошо выраженный, острый, кроме наиболее крупных экземпляров, у которых притуплён, треугольный, направлен вершиной назад; второй шип мал, расположен в основании верхнего (третьего шипа), треугольный, направлен назад и вниз, у наиболее крупных рыб развит плохо; самый верхний шип (являющийся окончанием подглазничной опоры) узкий, наиболее длинный, направлен вершиной назад, без дополнительного шипа в основании. Посттемпоральные шипы очень длинные, достигают или почти достигают вертикали заднего края первой субдорсальной костной пластинки. Оперкулярный шип исключительно длинный, оканчивается на уровне середины основания D1, узкий и острый. Клейтральный шип маленький, субтреугольный, отходит ниже оперкулярного шипа. Межглазничный промежуток вогнут, верхний край костной орбиты выступающий. Надглазничные гребни плохо выражены, до полного отсутствия у наиболее крупных экземпляров; когда имеются, расходящиеся, оканчиваются крошечным шипом, не развитым у более крупных рыб. Заглазничной борозды нет. Гребень sphenoticum оканчивается маленьким шипом, не выраженным у наибольшего из имеющихся экземпляра (SL 92 мм). Покровные кости головы мелко гранулированы.
Нижняя челюсть немного короче верхней; maxillare несколько расширено в задней части, заходит за уровень переднего края глаза; на челюстях очень мелкие зубы полосками, озубление praemaxillaria видно снаружи при закрытом рте. На сошнике маленькое субтреугольное пятно очень мелких зубов; нёбные кости без зубов. Язык очень короткий, треугольный, в основании широкий, а в передней части суженный, едва обособлен. Развитые жаберные тычинки длинные и тонкие, вооружены очень мелкими, не частыми шипиками. Ложножабра крупная. Хорошо развита щель позади четвёртой жаберной дуги.
Перед основанием D1 один непарный костный щиток и девять пар щитков под основанием D1 и в интердорсальном промежутке; под основанием D2 и в боковой линии щитков нет. В интердорсальном промежутке расположены последние четыре пары щитков. Щитки скульптированы бугорками, без шипов. Третья колючка D1 самая длинная и толстая; по переднему краю первой колючки сидят очень мелкие шипики. Первый луч D2 и передние пять лучей А неветвистые (первый луч А очень жёсткий, у более крупных рыб – как тонкая колючка); последний луч D2 часто расщеплён, но не до самого основания, тогда как последний луч А обычно раздвоен до основания и выглядит как два луча. С глубоко вырезан, его лопасти равной длины или нижняя несколько короче верхней, заострённые. Р очень длинные, вершина соединённого Р достигает уровня середины длины основания А. Наибольший (верхний) свободный луч Р длинный, обычно немного короче наибольших из соединённых лучей Р, но у части рыб почти равен им по длине. Начало V под основанием P, вершины достигают ануса или немного заходят за него, иногда достигая начала А. Колючка V короче ветвистых лучей, наиболее длинные ветвистые лучи – второй и третий. Анус расположен у начала А.
Чешуя очень мелкая, округлая, циклоидная, с мелковолнистым или гладким свободным краем, в передней половине тела не налегающая и сильно вросшая в кожу, а в задней – лучше выраженная, не врастающая и налегающая. Чешуя покрывает грудь (вперёд до вершины истмуса), основание Р (иногда только частично) и брюхо, включая промежуток между основаниями V (рис. 3б), но отсутствует в предорсальной области и позади основания Р, непосредственно над ним и между основаниями Р и V (иногда голые участки позади основания Р и между основаниями Р и V соединяются друг с другом, но чаще разделены рядом чешуек, доходящих почти до нижнего края основания Р), у части особей может отсутствовать также между задней половиной оснований V и сразу за ними. У рыб с наименее развитым чешуйным покровом на вентральной стороне тела чешуйное пятно на груди может быть плохо различимо из-за врастания чешуй в кожу, оканчивается примерно у середины длины оснований V, отделено от чешуй брюха широким голым участком. Чешуи боковой линии несколько крупнее прочих чешуй на теле. Боковая линия делает очень слабый изгиб вверх над оперкулярным шипом, далее назад идёт выше средней линии тела полого вниз, вновь делая изгиб у основания хвостового стебля, где она проходит по средней линии до основания лучей С. Трубочки боковой линии открываются вниз или назад.
Измерения, в % SL (n = 20): lc 34.3–37.5, ao 17.1–19.4, oo 10.4–13.6, io 10.4–12.8(16.0), lr 7.9–9.7, so 8.3–11.0, H 25.7–27.3, lP (39.8)41.8–45.1(48.5), lPf 38.5–39.0, lV 26.2–31.4(34.9).
Окраска фиксированных рыб бело-розовая или розовато-бежевая, вентральная сторона чисто белая; кости головы и дорсальные щитки желтоватые; на спине разбросаны мелкие чёрные пятнышки, организованные в продольные ряды под основанием спинных плавников и вдоль боковой линии и отдельно присутствующие между этими рядами и группой под боковой линией в передней половине тела; иногда тёмные пятнышки присутствуют на дорсальной поверхности головы и рыла, по нижнему краю глаза, на щеке и на жаберной крышке. У некоторых рыб чёрные пятнышки на спине очень малочисленные. D1 прозрачный или имеются размытые участки тёмного пигмента на перепонках между первой и второй, второй и третьей и третьей и четвёртой колючками, а иногда и суббазально между второй и третьей, третьей и четвёртой, четвёртой и пятой колючками; интеррадиальная мембрана здесь, по крайней мере, затемнена, но никогда не бывает чётко оформленного тёмного пятна. На D2 продольный ряд черновато-серых пятен. Внутренняя поверхность соединённого Р чёрно-серого цвета (лучи светлее перепонок между ними) с ярко-чёрным пятном в нижней части, по краям которого расположены серии небольших округлых или продолговатых белых пятен (рис. 3в); дистальный край соединённого Р со светлой каймой, не выраженной у отдельных экземпляров. Суббазальная часть свободных лучей Р черновато-серая, остальная часть этих лучей светлая. V и С не окрашены. Ротовая полость светлая, жаберная полость с участками буроватого пигмента или большей частью желтовато-буроватого цвета. Прижизненная окраска сходна с описанной выше, но отличается более насыщенными красноватыми тонами.
Замечания. Вид считается эндемиком Южно-Китайского моря (Richards et al., 2003). Описан из вод Филиппин (Herre, Kauffman, 1952), позже обнаружен у китайского побережья Тонкинского залива, откуда был описан в качестве нового вида P. spinosa (Asano, Okamura, 1963), впоследствии синонимизированного с P. tagala (Eschmeyer, 1998; Richards et al., 2003). Для фауны Вьетнама отмечается впервые. Изученные экземпляры хорошо соответствуют характеристике этого вида (Richards et al., 2003), за исключением наличия у части особей голого участка между задними половинами и/или за основаниями V и тёмного пигмента на межлучевых перепонках D1. Учитывая, что разная степень выраженности вышеуказанных признаков отмечена у рыб, пойманных одним тралом, данные различия отражают не более чем индивидуальную изменчивость, принимая её во внимание, следует отметить, что различия между P. tagala и описанным от северо-западных берегов Австралии видом P. elicryste Richards, Yato et Last, 2003 нуждаются в уточнении. Основные различия между этими видами связаны с наличием или отсутствием тёмного пигмента на D1 (нет у P. tagala, но, как правило, имеется между первой–третьей и пятой–седьмой колючками D1 у P. elicryste) и чешуи между основаниями V (есть у P. tagala, но нет у P. elicryste) (Richards et al., 2003). В то же время отсутствие чёрных пятен на D1 у отдельных экземпляров P. elicryste и наличие чешуи между основаниями V у голотипа этого вида постулировалось уже в первоописании (Richards et al., 2003). Учитывая выявленную у вьетнамских рыб вариабельность указанных признаков, их диагностическая значимость представляется сомнительной. Однако рисунок белых пятен на внутренней поверхности соединённого P у P. elicryste (Richards et al., 2003. Fig. 7) несколько отличается от такового у P. tagala, включая описанные здесь экземпляры (рис. 3в). Вопрос об отношениях P. tagala и P. elicryste требует специального исследования. Если окажется, что эти номинальные виды действительно конспецифичны, ареал P. tagala будет значительно расширен (от Южно-Китайского моря до Северо-Западной Австралии).
По данным траловых съёмок в 2005–2009 гг., в заливе Нячанг P. tagala является обычным и многочисленным видом в глубоководной части залива в летний период, в уловах отмечен с глубины от 70 м. В зимний сезон при тралениях < 100 м не встречался. Ранее был известен в диапазоне глубин 95–119 м (Richards et al., 2003).
Pterygotrigla (Parapterygotrigla) multiocellata (Matsubara, 1937)
(рис. 4–6)
Рис. 4.
Pterygotrigla (Parapterygotrigla) multiocellata SL 220 мм: а – общий вид, б – носовые (na) и ростральные шипы (ac.ro – дополнительный шип в основании рострального шипа), в – постокулярный шип, г – сфенотикальный шип, д – предорсальная область (ads – непарная пластинка перед основанием D1), е – внутренняя сторона Р; ост. обозначения см. на рис. 2. Масштаб, мм: б, в, е – 20; г – 10, д – 15.


Рис. 6.
Pterygotrigla (Parapterygotrigla) multiocellata: а – голова SL 45 мм, вид сверху (pro – преорбитальный шип), б–г – ростральные и носовые шипы (б – SL 55 мм, вид сверху; в, г – соответственно SL 55 и 45 мм, вид сбоку); д – заглазничная часть крыши черепа (SL 55 мм, вид сбоку), е – верхний преоперкулярный шип (вверху – SL 55 мм, внизу – SL 45 мм), ж – SL 55 мм, предорсальная область; ост. обозначения см. на рис. 2, 4. Масштаб: 2 мм (а – 4 мм).
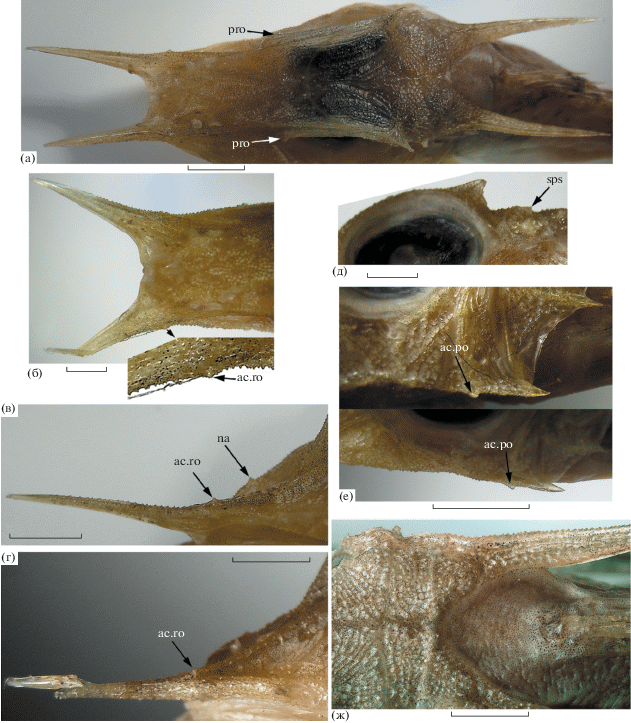
Материал. Всего 5 экз.: 2 экз. SL 45 и 55 мм, Вьетнам, Нячанг–Няфу (Cang Ca Vinh Luong), май–июнь 2009 г.; 3 экз. SL 207–220 мм, там же, близнецовые тралы, апрель–июнь 2012 г.
Описание. D1 VIII, D2 11½, A 12½, P 12 + 3, V I + 5; LL ~ 60–70; sp. br. 0–1 + 1 + 7–8, кроме того, один бугорок на epibranchiale 1 и пять – в нижней части ceratobranchiale 1.
Взрослые рыбы SL 207–220 мм (рис. 4). Ростральные шипы очень длинные, соизмеримы с суммарной величиной диаметра орбиты и длины посторбитальной части головы, прямые, плоские, на всем протяжении равномерно широкие и лишь у самой вершины резко сужены и приострены (у экз. SL 207 мм с левой стороны вершина широкая и оканчивается несколькими разноразмерными зубцами, возможно, это результат прижизненного повреждения на более ранней стадии онтогенеза). У латерального края рострального шипа в его основании имеется сильный дополнительный шип, направленный вперёд и вверх (рис. 4б). Носовые шипы (одна пара) хорошо развиты. Наружный гребень подглазничной опоры на инфраорбитальных костях не выражен, на praeoperculum – отчётливый, завершается хорошо развитым верхним преоперкулярным шипом, длина которого равна или немного превышает длину оперкулярного шипа. Вершина верхнего преоперкулярного шипа достигает заднего края бранхиостегальной мембраны. У экз. SL 212 мм под верхним преоперкулярным шипом хорошо развит небольшой (вдвое короче верхнего) островершинный дополнительный шип, у двух других экземпляров он редуцирован до крошечного выступа или совершенно не развит. Угловая часть praeoperculum образует плоский треугольный шип или угловатую лопасть, несущую несколько шипиков, отделена от верхнего преоперкулярного шипа пологой выемкой. Посттемпоральные шипы очень длинные, достигают переднего края – середины длины второй субдорсальной костной пластинки. Оперкулярный шип короткий, острый, оканчивается перед вертикалью заднего конца оперкулярной лопасти или на одной вертикали с ним. Клейтральный шип очень длинный, узкий и острый, отходит почти на уровне оперкулярного шипа, оканчивается на уровне основания четвёртой–пятой колючки D1. Межглазничный промежуток отчётливо вогнут, верхний край костной орбиты сильно приподнят. Надглазничные гребни не выражены, но у заднего конца приподнятого верхнего края костной орбиты развит маленький крепкий шипик (постокулярный), направленный вверх и назад (рис. 4в), за основанием которого прослеживается слабое вдавление. Гребень sphenoticum слабо намечен, но оканчивается маленьким острым шипиком (рис. 4г), едва прослеживаемым у наибольшего экземпляра, но отчётливым с обеих сторон у двух других. Покровные кости головы мелко гранулированы, в межглазничном промежутке и на ростральных шипах бугорки образуют продольные гребни, тогда как на крышечных и подглазничных костях формируют ячеистый орнамент. На ростральных шипах и колючках D1 наблюдаются начальные явления гиперостоза.
Челюсти почти равной длины; maxillare слабо расширено в задней части, оканчивается перед вертикалью переднего края глаза; на челюстях мелкие зубы полосками, озубление praemaxillaria полностью видно снаружи при закрытом рте. На сошнике маленькое треугольное пятно мелких зубов, нёбные кости без зубов. Язык короткий, массивный, широко закруглённый на вершине, слабо обособлен. Жаберные тычинки несут очень мелкие шипики по внутреннему краю. Ложножабра очень крупная, из 52–55 элементов. Хорошо развита щель позади четвёртой жаберной дуги.
Перед основанием D1 один небольшой непарный костный щиток полулунной формы (рис. 4д), ширина которого вдвое превышает ширину основания первой колючки D1; под основанием D1 восемь пар узких костных щитков и девятый маленький непарный треугольный щиток, расположенный в интердорсальном промежутке; все щитки полностью выступают из кожи, покрыты мелкими бугорками и/или рёбрышками, не несут шипов. Под основанием D2, как и в боковой линии, щитки отсутствуют. Третья колючка D1 наибольшая; основные две трети переднего края первой и второй колючки с зубчиками. Первые два луча D2 и передние четыре–шесть лучей А неветвистые (первый луч А очень жёсткий); последний луч в этих плавниках раздвоен до основания. С глубоко вырезан, его лопасти приострены, верхняя немного длиннее нижней. Р длинные, вершина соединённого Р достигает начала последней трети основания А (его восьмого–десятого луча). Наибольший (верхний) свободный луч Р значительно короче наибольших из соединённых лучей Р, достигает только начала А. Начало V под нижним концом основания P, вершины достигают начала А. Колючка V короче ветвистых лучей, третий ветвистый луч самый длинный (но незначительно длиннее второго и четвёртого). Анус расположен вблизи начала А.
Чешуя очень мелкая, округлая, циклоидная, с гладким свободным краем, в наиболее передней части туловища (до уровня шестой колючки D1) и под основаниями D1 и D2 выше LL не налегающая, частично вросшая в кожу, в остальных отделах налегающая. В предорсальной области (рис. 4д), под основанием D1 выше LL до уровня второй–четвёртой пары субдорсальных щитков, на горле, груди, на основании Р, между основаниями Р и V, между основаниями V и сразу позади них и на узкой полосе по среднебрюшной линии до ануса чешуя полностью отсутствует. Чешуи боковой линии крупнее прочих чешуй на теле. Боковая линия практически прямая, идёт полого вниз от верхнего края жаберного отверстия до начала хвостового стебля, где делает слабый изгиб и продолжается посередине хвостового стебля до основания С. Трубочки боковой линии открываются частью назад, частью назад и вниз (в задней половине LL в большинстве случаев только назад).
Измерения, в % SL: lc 45.3–46.9, ao 27.3–29.0, oo 8.7–9.1, io 8.6–9.2, lr 17.5–19.3, so 6.8–7.3, H 16.4–18.9, lP 37.3–39.2, lPf 21.4–22.2, lV 21.3–22.2.
Окраска фиксированных экземпляров розовато-бежевая, очень светлая, вентральная сторона чисто белая; покровные кости желтоватые; верхнебоковая поверхность тела (выше LL) в некрупных разноразмерных округлых тёмных пятнышках, ниже LL такие пятнышки единичны, непосредственно на LL они обычно частично сливаются в короткие продольные полоски. Мелкие округлые пятнышки присутствуют в межглазничном промежутке и на голом участке между вершиной praeoperculum и боковым краем черепа непосредственно за глазом. Перепонка D1 по дистальному краю черноватая, следы чёрного пигмента прослеживаются также по дистальному краю D2 и (более отчётливо) С (при увеличении на лучах D2 по всей их длине обнаруживается очень мелкий рассеянный меланофорный крап); в остальном непарные плавники не окрашены. Внутренняя поверхность перепонки между вторым–девятым соединёнными лучами Р черноватая, однотонная, без белых пятен; сами лучи светлые; перепонка между первым и вторым и между самыми нижними лучами Р чисто белая (рис. 4е); дистальный край соединённого Р со светлой каймой. Свободные лучи Р светлые; V не окрашены. Ротожаберная полость светлая. Прижизненная окраска не документирована.
Мальки SL 45 (рис. 5а, 6) и 55 мм (рис. 5б, 6). Все шипы заметно более тонкие и узкие, островершинные; ростральные шипы короче, а посттемпоральные, напротив, более длинные, чем у взрослых рыб (рис. 5, 6а). Ростральные шипы расходящиеся, в большей степени у малька SL 55 мм. Носовые шипы и дополнительный шип в основании рострального шипа присутствуют, но развиты очень слабо; бугорковидные, незначительно крупнее окружающих бугорков орнамента костей (у малька SL 55 мм носовые шипы едва заметны, но дополнительный шип развит столь же хорошо, как у малька SL 45 мм) (рис. 6б–6г). У переднего края орбиты в начале её утолщённого дорсального края имеется слабый, но вполне отчётливый выступ, который можно посчитать зачаточным преорбитальным шипом (лучше выражен у малька SL 55 мм). Постокулярные шипы крупные, треугольные, продолжаются вперёд в виде короткого слабого гребешка по утолщённому и приподнятому верхнему краю орбиты, вперёд достигающему вертикали заднего края зрачка. Гребень sphenoticum в виде короткого, но широкого треугольного или прямоугольного выступа, оканчивающегося коротким шипиком (рис. 6а, 6д). На пластинке praeoperculum в промежутке между верхним шипом (являющимся окончанием суборбитальной опоры) и угловой лопастью прослеживаются три слабо расходящихся гребешка, самый верхний из которых выступает за задний край пластинки в виде очень слабого шипа. В основании верхнего преоперкулярного шипа хорошо развит дополнительный шипик (рис. 6е). Посттемпоральные шипы у малька SL 55 мм достигают конца основания D1, а у малька SL 45 мм промежутка между первым и вторым лучами D2. В основании клейтрального шипа имеется широкий лопастевидный выступ. Элементов ложножабры 25–28. Пластинчатые расширения птеригиофоров в основании D1 полностью покрыты кожей, непарная предорсальная пластинка очень маленькая. На челюстях полоски мелких щетинковидных зубов, на сошнике поперечная полоска таких же зубов, нёбные кости без зубов. Вершина Р достигает пятого луча А, первый свободный луч Р оканчивается на одной вертикали с вершиной V, достигает ануса. Брюхо целиком голое у обоих мальков, но у малька SL 55 мм вросшая чешуя покрывает предорсальую область (рис. 6ж) (у малька SL 45 мм предорсальная область и участок под началом D1 выше LL голые, как и у взрослых рыб).
Измерения, в % SL11: lc 51.1 (47.3), ao 26.7 (26.4), oo 12.2 (10.0), io 12.2 (11.8), lr 12.7 (14.6), so 8.6 (9.1), H 13.3 (17.3), lP 31.1 (30.9), lPf 23.3 (22.7), lV 23.3 (22.7).
Окраска. Бока тела выше уровня клейтрального шипа и покровные кости верхней и боковой поверхности головы в рассеянном меланофорном крапе, местами на боках группирующемся в пятнышки; по ходу LL имеются немногочисленные, чётко оформленные продолговатые пятна. Перепонка D1 густо испещрена крупными черноватыми меланофорами, по дистальному краю сливающимися; D2 с субдистальной чёрной полоской; дистальный край С черноватый; перепонка между вторым и седьмым лучами соединённого Р с внутренней стороны чёрная; в остальном плавники не окрашены.
Замечания. Малёк SL 55 мм отличается от других исследованных рыб (включая малька SL 45 мм) наличием вросших в кожу чешуй в предорсальной области (рис. 6ж), а также (по сравнению с мальком SL 45 мм) более короткими ростральными и менее развитыми носовыми шипами, но более длинными посттемпоральными и заметно более расходящимися ростральными шипами. Различия в ошиплении головы без особых возражений можно связать с индивидуальной изменчивостью, однако присутствие вросших чешуй вызывает некоторые сомнения в идентификации этого малька. В пределах подрода Parapterygotrigla чешуя в предорсальной области отмечена у видов P. hoplites (Fowler, 1938) и P. megalops (Fowler, 1938), однако наличие дополнительного шипика в основании ростральных шипов у малька SL 55 мм исключает его отождествление с перечисленными видами. Последний признак среди всех известных триглид характерен только для P. multiocellata. Возможно, присутствие чешуй в предорсальной области у малька SL 55 мм является следствием случайной рекапитуляции этого признака, однако для окончательного суждения необходимы дополнительные материалы. В остальном исследованные рыбы хорошо соответствуют характеристике этого вида (Richards, Yato, 2014).
Вид P. multiocellata впервые отмечен для фауны Вьетнама. Он был описан из вод Японии, позже найден у о. Сайпан (Марианские о-ва), у западного и восточного побережья Австралии (Richards, Yato, 2014). В Южно-Китайском море ранее был известен из вод южной оконечности о. Тайвань (Shao et al., 2008).
Pterygotrigla (s. str.) ryukyuensis Kamohara, 1958
(рис. 7)
Материал. 1 экз. SL 126 мм, Вьетнам, Нячанг–Няфу (Cang Ca Vinh Luong), близнецовые тралы, май–июнь 2009 г.
Рис. 7.
Pterygotrigla (s. str.) ryukyuensis SL 126 мм: а – общий вид, б – постокулярный шип, в – заглазничная часть крыши черепа и посттемпоральные шипы (вид сверху) (линейка общая для б и в), г – внутренняя сторона Р; обозначения см. на рис. 2. Масштаб, мм: б, в – 15, г – 8.
Описание. D1 IX (последняя колючка очень короткая, неподвижно соединена с костной пластинкой и выглядит как шип в центре пластинки), D2 11½, A 12½, P 12 + 3, V I + 5; LL ~ 60; sp. br. 0 + 1 + 7, кроме того, два бугорка на epibranchiale 1 и пять – в нижней части ceratobranchiale 1; элементов ложножабры 50.
Ростральные шипы длинные, в основании широко треугольные, а к вершине узкие, плоские, острые, расходящиеся, очень слабо изогнутые, в основании мелкобугорчатые по наружному краю. Носовые шипы отсутствуют. Подглазничная опора маркирована слабым, но отчётливым гребнем, протягивающимся от основания рострального шипа до верхнего преоперкулярного шипа. Praeoperculum с крупным острым верхним шипом, длина которого (с гребнем) равняется таковой оперкулярного шипа; ниже его расположен маленький шип, отделённый пологой выемкой от слабого шиповидного выступа вершины преоперкулярной лопасти. Посттемпоральные шипы очень длинные, достигают середины длины второй субдорсальной костной пластинки. Оперкулярный шип короткий, оканчивается на вертикали заднего конца оперкулярной лопасти, острый. Клейтральный шип очень длинный, узкий и острый, отходит почти на уровне оперкулярного шипа, оканчивается на вертикали основания пятой колючки D1 (рис. 7а). Межглазничный промежуток отчётливо вогнут, верхний край костной орбиты сильно приподнят. Надглазничные гребни не выражены, но у заднего конца приподнятого верхнего края костной орбиты развит маленький крепкий шипик, направленный вверх и назад (рис. 7б), позади которого имеется маленькое углубление (но признаков заглазничной борозды нет). Гребень sphenoticum шельфовидный, оканчивается острым шипиком (рис. 7в). Покровные кости головы мелко гранулированы, бугорки на костях крыши черепа организованы в ряды, тогда как на оперкулярных и подглазничных костях формируют ячеистый орнамент.
Челюсти почти равной длины; maxillare слабо расширено в задней части, оканчивается перед вертикалью переднего края глаза; на челюстях мелкие зубы полосками, озубление praemaxillaria видно снаружи при закрытом рте. На сошнике поперечная полоска мелких зубов; нёбные кости без зубов. Язык короткий, широкий, массивный, слабо обособлен. Пять верхних развитых жаберных тычинок длинные и очень тонкие, далее последовательно укорачиваются и становятся более уплощёнными и расширенными в основаниях; по внутреннему краю несут мелкие шипики. Ложножабра очень крупная. Хорошо развита щель позади четвёртой жаберной дуги.
Перед основанием D1 один очень маленький непарный костный щиток, ширина которого не превышает ширины основания первой колючки D1; под основанием D1 и в интердорсальном промежутке девять пар костных щитков (очень узкие, последние две пары слиты, девятая расположена в интердорсальном промежутке), все полностью выступают из кожи, покрыты мелкими бугорками, не несут шипов. Под основанием D2, как и в боковой линии, щитки отсутствуют. Третья колючка D1 наиболее массивная и, по-видимому, была наиболее длинная (но у изученного экземпляра она была обломана ещё при жизни); передний край первой колючки в базальной половине со слабо развитыми зубчиками. Первый луч D2 и передние пять лучей А неветвистые (первый луч А отчётливо колючий); последний луч в этих плавниках раздвоен до основания. С глубоко вырезан, его лопасти приострены, верхняя чуть длиннее нижней. Р длинные, вершина соединённого Р достигает середины длины основания А (его шестого луча). Наибольший (верхний) свободный луч Р короче наибольших из соединённых лучей Р, немного заходит за начало А (до уровня основания его второго луча). Начало V под нижним концом основания P, вершины достигают ануса. Колючка V короче ветвистых лучей, второй ветвистый луч самый длинный (но едва длиннее первого и третьего). Анус расположен вблизи начала А.
Чешуя очень мелкая, округлая, циклоидная, с гладким свободным краем, в передней половине тела (до уровня четвёртого луча D2) выше LL, а также вблизи границы с голыми участками между основаниями Р и V вросшая в кожу, в остальных отделах налегающая. В предорсальной области, на горле, груди, на основании Р, между основаниями Р и V, между основаниями V и сразу позади них чешуя полностью отсутствует. Чешуи боковой линии крупнее прочих на теле. Боковая линия делает очень слабый изгиб книзу под основанием D1, идёт полого вниз от верхнего края жаберного отверстия до середины основания С. Трубочки боковой линии открываются назад и вниз.
Измерения, в % SL: lc 42.1, ao 20.6, oo 10.3, io 12.3, lr 10.7, so 9.9, H 22.2, lP 31.8, lPf 24.6, lV 22.2.
Окраскафиксированного экземпляра розовато-бежевая, очень светлая, вентральная сторона чисто белая; покровные кости желтоватые; верхнебоковая поверхность тела (выше уровня клейтрального шипа) в некрупных разноразмерных округлых тёмных пятнышках (большинство пятен – выше боковой линии); на голове пестрины отсутствуют. Перепонка D1 с разлитой точечной меланофорной пигментацией, не различимой невооружённым глазом; на D2 продольный медиальный ряд удлинённых черноватых пятен, расположенных только на плавниковых лучах. Внутренняя поверхность перепонки между первым–восьмым соединёнными лучами Р черноватая, однотонная, без белых пятен; сами лучи светлые; перепонка между последующими лучами чисто белая (рис. 7г); дистальный край соединённого Р со светлой каймой. Свободные лучи Р светлые, с единичным очень мелким меланофорным крапом, не различимым невооружённым глазом. А, V и С не окрашены. Ротожаберная полость светлая. Прижизненная окраска не документирована.
Замечания. Описанный экземпляр не отличается от ранее известных представителей вида (Richards, Yato, 2012). Распространён от Южной Японии через Южно-Китайское море и моря Индонезии до Северо-Западной Австралии. Во Вьетнаме ранее указывался только для Тонкинского залива (Nguyen, 1999). В заливе Нячанг, по-видимому, очень редок, что, возможно, объясняется относительной глубоководностью этого вида. Экземпляры, приведённые в работе (Richards, Yato, 2012), были пойманы на глубинах 205–330 м.
Онтогенетическая изменчивость видов рода Pterygotrigla
При сравнении малька P. hemisticta SL 62 мм со взрослыми рыбами SL 225–295 мм бросается в глаза ряд структурных различий в ошиплении головы, на которые ранее в литературе не обращали специального внимания. Все шипы на голове у малька гораздо ýже и тоньше, чем у взрослых рыб, хотя относительная длина большинства шипов (за исключением ростральных и посттемпоральных) у них соизмерима. Межглазничный промежуток у малька вогнут заметно сильнее (соответственно верхний край орбит сильнее приподнят), и, в отличие от взрослых рыб, имеются отчётливый супраорбитальный гребень, оканчивающийся хорошо развитым постокулярным шипом, и развитый гребень sphenoticum, оканчивающийся шипом. Посттемпоральные шипы у малька помимо того, что значительно длиннее, чем у взрослых рыб (достигают уровня второй пары субдорсальных пластинок, тогда как у взрослых не заходят за непарную предорсальную пластинку), отчётливо отклонены назад и вбок (прижаты к телу у взрослых рыб). Ошипление praeoperculum у малька выражено гораздо лучше, чем у взрослых, угловая часть praeoperculum образует отчётливый шип, в основании наиболее развитого (верхнего) шипа, являющегося окончанием суборбитальной опоры, имеется сильный дополнительный (accessory) шип; между ними в пологой выемке расположены два равнорасставленных треугольных шипа. У взрослых рыб никаких признаков дополнительного шипа в основании верхнего (основного) преоперкулярного шипа нет, в выемке между верхним шипом и угловатой преоперкулярной лопастью нижний шип утрачивается, а верхний оказывается сближенным с основным шипом (т.е. терминальным шипом суборбитальной опоры) (рис. 1в). Помимо различий в ошиплении для малька также характерны более длинные Р (до середины длины основания А, тогда как у взрослых – до конца передней трети его длины), более развитая диффузная пигментация межрадиальной перепонки колючей части D и более сгущённые пятнышки на теле.
Второй имеющийся в изученном материале вид подрода Otohime – P. tagala – представлен серией экземпляров SL 51–92 мм. Все они характеризуются дефинитивным ошиплением головы, вариации которого незначительны, при том что наименьшие исследованные экземпляры (SL 51 мм) заметно мельче вышеописанного малька P. hemisticta (SL 62 мм). Однако вид P. tagala имеет гораздо меньшие абсолютные размеры, чем P. hemisticta (наибольшие из известных рыб не превышают SL 98 мм (Richards et al., 2003), тогда как P. hemisticta достигает SL 300 мм (Ochiai, Yatou, 1985; Yamada, 2002)). Надо полагать, что дефинитивное ошипление головы формируется у P. tagala при гораздо меньших абсолютных размерах, чем у P. hemisticta, и экземпляры P. tagala SL > > 50 мм уже утрачивают ювенильные черты строения.
Подрод Parapterygotrigla представлен в изученном материале видом P. multiocellata. Мальки этого вида SL 45–55 мм по отношению ко взрослым рыбам SL 207–220 мм характеризуются теми же структурными различиями, что описаны выше для P. hemisticta, за исключением того, что при меньших абсолютных размерах шипы на praeoperculum у мальков P. multiocellata редуцированы в большей степени. Ростральные шипы у обоих мальков короче, чем у взрослых рыб, это, вероятно, видоспецифическая особенность, так как для молоди других видов подрода Parapterygotrigla описаны очень длинные ростральные шипы (Richards, Yato, 2014). Характерные для подрода носовые шипы, а также дополнительный шип в основании рострального шипа при SL 45 мм уже вполне сформированы, но ещё очень малы. По-видимому, у разных видов подрода Parapterygotrigla последовательность формирования шипов может сильно различаться. Так в опубликованной ранее работе (Richards, Yato, 2014. Figs. 8, 9 ) для личинки SL 10.2 мм из вод Северо-Западной Австралии с ещё не сформированными ростральными шипами изображены хорошо развитые носовой шип и дополнительный шип суборбитальной опоры, тогда как у личинки SL 14.5 мм из Тиморского моря они ещё не выражены. В отличие от других видов подрода у исследованных мальков P. multiocellata соединённый Р заметно короче, чем у взрослых рыб. Как и у малька P. hemisticta, у мальков P. multiocellata гораздо лучше, чем у взрослых рыб, выражена меланофорная пигментация на перепонке D1, но число пятнышек на теле меньше; в общих чертах пигментация молоди этого вида при SL 45 мм соответствует таковой у взрослых рыб.
Рис. 8.
Pterygotrigla (s. str.) picta, юго-восточная часть Тихого океана: а – голова (вид сбоку); б, г, е – общий вид; в, д, ж – верхний преоперкулярный шип (в, д – вентрально; ж – дорсолатерально); а–в – экз. SL 17 мм; г, д – экз. SL 37.5 мм; е, ж – экз. SL 55 мм. Обозначения см. на рис. 2. Масштаб: 1 мм (а – 2.5 мм).

К сожалению, я не располагаю экземплярами P. ryukyuensis мальковых стадий из вод Вьетнама, и в литературе они не описаны. Однако для сравнения была изучена возрастная серия вида P. picta из юго-восточной части Тихого океана (рис. 8), также принадлежащего к номинотипическому подроду, которая представлена особями SL 17–350 мм. Наименьший экземпляр в данной серии, имеющий SL 17 мм (рис. 8а–8в), по степени развития сопоставим с личинкой Pterygotrigla sp. SL 10.2 мм, изображённой в работе (Richards, Yato, 2014. Fig. 8). У него имеются короткие, но вполне развитые ростральный и оперкулярный шипы; зазубренные, очень длинные, слегка заходящие концами за начало D2 посттемпоральные шипы; клейтральный шип отсутствует; Р вееровидной формы, оканчивается на уровне передней трети А, свободных лучей в нём ещё нет. Межглазничный промежуток вогнут, а кроме того, имеется углубление заглазничной части крыши черепа перед основаниями посттемпоральных шипов. Верхний край орбит сильно приподнят и образует массивный треугольный шиповидный выступ, расположенный на уровне середины глаза. Обособленный гребень sphenoticum отсутствует, боковой край черепа образует сплошной утолщённый валик от заднего края орбиты до основания посттемпорального шипа. Praeoperculum с пятью шипами, мало различающимися по величине, в основании самого верхнего из них имеется массивный, направленный вбок дополнительный шип (соизмеримый по величине с другими) (рис. 8в). Пигментация этого малька не сохранилась. При SL 32–39 мм ростральные шипы остаются короткими, но посттемпоральные очень длинные, достигают середины длины основания D1; клейтральный шип сформирован (рис. 8г). Верхний преоперкулярный шип длинный, у его основания имеется шельфовидный дополнительный выступ (рис. 8д), но шипы заднего края praeoperculum в промежутке между верхним шипом и угловой лопастью этой кости уже полностью редуцированы (как вариация, непосредственно под верхним преоперкулярным шипом может присутствовать небольшой добавочный шип). В данной размерной группе отчётливо прослеживается редукция постокулярных шипов, которые у малька SL 32.5 мм представлены небольшими треугольными выступами, продолжающимися вперёд в виде короткого слабого гребня в задней части утолщённого приподнятого верхнего края орбиты. При SL 37.5 мм сохраняется только очень слабый гребень без шипа (рис. 8г), а при SL 39 мм и он уже утрачен. Малёк SL 55 мм по степени развития шипов уже вполне сопоставим со взрослыми рыбами (рис. 8е, 8ж), за исключением того, что ростральные шипы у него короче, а посттемпоральные – длиннее; в дальнейшем в онтогенезе наблюдается увеличение длины ростральных шипов и уменьшение посттемпоральных. У наиболее крупного из исследованных экземпляра (SL 350 мм) наблюдаются явления пахиостоза. У мальков SL 32–55 мм соединённый Р достигает середины длины основания А, свободные лучи Р полностью обособлены, самый верхний из них немного короче наибольших лучей соединённого Р. У рыб SL > 100 мм соединённые лучи Р достигают или почти достигают конца основания А, а свободные остаются короткими, самый длинный из них (верхний) не достигает середины длины основания А. Пигментация мальков вполне соответствует таковой у более крупных рыб, за исключением того, что разлитая тёмная меланофорная пигментация межлучевой перепонки D1 отчётливо выражена, по меньшей мере, до SL 55 мм (у взрослых рыб перепонка прозрачная).
Подводя итог, следует отметить, что общей чертой молоди Pterygotrigla является наличие у неё развитых постокулярных шипов, дополнительного шипа на подглазничной опоре в основании верхнего преоперкулярного шипа и в той или иной степени выраженное сохранение личиночного ошипления praeoperculum (но первая и последняя особенности быстро исчезают у молоди P. picta), а также более интенсивная меланофорная пигментация межлучевой перепонки D1. В дефинитивном состоянии все эти признаки утрачиваются. Для оценки степени метаморфоза, очевидно, наиболее подходят особенности ошипления головы, при этом следует иметь в виду, что у разных видов преобразование этого признака в онтогенезе идёт, по-видимому, с разной скоростью. Если у P. picta утрата постокулярных и добавочных преоперкулярных шипов наблюдается уже при SL 37–39 мм, то у P. hemisticta при SL 62 мм они хорошо развиты (максимальные размеры этих видов сходные). Онтогенетические преобразования могут быть видоспецифичными, например, у P. picta и P. multiocellata длина ростральных шипов и длина соединённого Р с ростом увеличивается, тогда как у P. hemisticta, по-видимому, наоборот. Однако для других видов подрода Parapterygotrigla на ранних стадиях отмечены более длинные ростральные шипы и соединённый Р, чем у взрослых рыб (Whitley, 1952; Richards, Yato, 2014), поэтому ни один из выявленных признаков нельзя считать специфичным на уровне подродов. Можно лишь отметить, что гребень sphenoticum и его шип, в изученном материале хорошо развитые у молоди P. hemisticta (подрод Otohime) и P. multiocellata (подрод Parapterygotrigla), совершенно отсутствуют у соразмерной молоди P. picta (номинотипический подрод), однако без сравнительных данных по другим видам связывать это различие с подродовой классификацией представляется преждевременным. Характерные для подрода Parapterygotrigla носовые шипы появляются уже на самых ранних стадиях онтогенеза (Richards, Yato, 2014. Fig. 8).
КЛЮЧ ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА PTERYGOTRIGLA ЮЖНО-КИТАЙСКОГО МОРЯ
1(2) Носовые шипы имеются (подрод Parapte-rygotrigla) ..………………………..............................…. 3
2(1) Носовые шипы отсутствуют ………….….… 9
3(4) В основании рострального шипа имеется дополнительный шип (рис. 4б) ……. P. (P.) multiocellata
(Япония, Тайвань, Вьетнам, Марианские о-ва, тропическая Западная и Восточная Австралия)
4(3) В основании рострального шипа нет дополнительного шипа ……………….…..................... 5
5(6) Нет шипов при основании лучей D2; длина рыла (без ростральных шипов) >26% длины головы ……………………….....……….. P. (P.) macrorhynchus
(Япония, Тайвань, Вьетнам, Филиппины, северное и западное побережье Австралии, Индия, Мальдивские о-ва, Мадагаскар, Восточная Африка (Мозамбик))
6(5) Ряд направленных вперёд шипов с каждой стороны у основания лучей D2; длина рыла (без ростральных шипов) < 20% длины головы ….….. 7
7(8) Непарная костная пластинка перед началом D1 имеется …….....................…. P. (P.) megalops
(Южная Япония, Филиппины, Марианские о-ва, Австралия (Квинсленд))
8(7) Перед началом D1 нет костной пластинки ..…………………….............................…. P. (P.) hoplites
(Филиппины, Северная и Западная Австралия)
9(10) Клейтральный шип значительно длиннее оперкулярного (подрод Pterygotrigla s. str.) …...………………………………..…. P. (s. str.) ryukyuensis
(От Южной Японии через Южно-Китайское море и моря Индонезии до Северо-Западной Австралии)
10(9) Клейтральный шип значительно короче оперкулярного (подрод Otohime) ….................... 11
11(12) В соединённой части Р 12 лучей; чешуйный покров на груди, если имеется, не заходит за интервентральную линию; D1 с чётко оформленным крупным чёрным пятном …….. P. (O.) hemisticta
(Западная часть Тихого океана от Японии до Австралии)
12(11) В соединённой части Р 13 лучей; чешуйный покров на груди всегда имеется и заходит за интервентральную линию (рис. 3б); D1 не пигментирован или с разлитой тёмной пигментацией …........................................... P. (O.) tagala
(Южно-Китайское море у берегов Китая, Вьетнама и Филиппин (о. Лусон))
Список литературы
Asano H., Okamura O. 1963. A new gurnard of the genus Pterygotrigla from Tonking Bay // Bull. Misaki Mar. Biol. Inst. № 4. P. 49–52.
Eschmeyer W.N. (Ed.). 1998. Catalog of Fishes. V. 1–3. San Francisco: Calif. Acad. Sci., 2905 p.
Gomon M.F., Kawai T. 2018. A review of Indonesia’s Indian Ocean species of Lepidotrigla gurnards (Teleostei: Scorpaeniformes: Triglidae) with descriptions of three new species from southern coastal waters // Raffles Bull. Zool. V. 66. P. 624–651.
Herre A.W.C.T., Kauffman D.E. 1952. New and little known Philippine triglids // Proc. Biol. Soc. Wash. V. 65. P. 27–30.
Khalaf M.A., Disi A.M., Krupp F. 1996. Four new records of fishes from the Red Sea // Fauna of Saudi Arabia. V. 15. P. 402–406.
Nguyen H.P. 1999. Checklist of marine fishes in Vietnam. V. 5. Hochiminh: Agricult. Publ. House, 305 p.
Ochiai A., Yatou T. 1985. Family Triglidae gurnards // The fishes of the Japanese Archipelago. Pt. 1 / Eds. Masuda H. et al. Tokyo: Tokai Univ. Press. P. 333–334.
Randall J.E., Lim K.K.P. 2000. A checklist of the fishes of the South China Sea // Raffles Bull. Zool. Suppl. № 8. P. 569–667.
Richards W.J. 1984. Triglidae // FAO Species Identification Sheets for Fishery Purposes. Western Indian Ocean, Fishing area 51. V. 5 / Eds. Fischer W., Bianchi G. Rome: FAO, pag. var.
Richards W.J., Jones D.L. 2002. Preliminary classification of the gurnards (Triglidae: Scorpaeniformes) // Mar. Freshwat. Res. V. 53. № 2. P. 274–282. https://doi.org/10.1071/MF01128
Richards W.J., Saksena V.P. 1977. Systematics of the gurnards, genus Lepidotrigla (Pisces, Triglidae) from the Indian Ocean // Bull. Mar. Sci. V. 27. № 2. P. 208–222.
Richards W.J., Yato T. 2012. The tropical species of the subgenus Pterygotrigla (Pisces: Triglidae: Pterygotrigla) with description of a new species // Zootaxa. V. 3484. № 1. P. 53–64. https://doi.org/10.11646/zootaxa.3484.1.3
Richards W.J., Yato T. 2014. Revision of the subgenus Parapterygotrigla (Pisces: Triglidae: Pterygotrigla) // Ibid. V. 3768. № 1. P. 23–42. https://doi.org/10.11646/zootaxa.3768.1.2
Richards W.J., Yato T., Last P.R. 2003. Revision of the gurnard fish subgenus Otohime (Triglidae, Pterygotrigla) // Smithiana Bull. V. 2. P. 1–18.
Shao K.-T., Ho H.-C., Lin P.-L. et al. 2008. A checklist of the fishes of southern Taiwan, northern South China Sea // Raffles Bull. Zool. Suppl. № 19. P. 233–271.
Whitley G.P. 1952. Some noteworthy fishes from eastern Australia // Proc. R. Soc. N.S.W., 1950–1951. P. 27–32.
Yamada U. 2002. Triglidae // Fishes of Japan with pictorial keys to the species. V. 1 / Ed. Nakabo T. Tokyo: Tokai Univ. Press. P. 604–609.
Yato T., Heemstra E. 2019. A new deepwater gurnard of the genus Pterygotrigla (Scorpaeniformes: Triglidae) from the southwestern Indian Ocean // Zootaxa. V. 4706. № 1. P. 189–200. https://doi.org/10.11646/zootaxa.4706.1.8
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии