

Вопросы ихтиологии, 2022, T. 62, № 2, стр. 180-187
Биологическая характеристика полиперы симуширской Polypera simushirae (Liparidae) из тихоокеанских вод северных Курильских островов в осенний период
Ю. Н. Полтев *
Сахалинский филиал Всероссийского научно-исследовательского института
рыбного хозяйства и океанографии − СахНИРО
Южно-Сахалинск, Россия
* E-mail: y.poltev@sakhniro.ru
Поступила в редакцию 04.03.2021
После доработки 27.04.2021
Принята к публикации 27.04.2021
- EDN: LNQKXK
- DOI: 10.31857/S0042875222010131
Аннотация
Рассмотрены размерно-половой состав, питание и плодовитость симуширской полиперы Polypera simushirae в тихоокеанских водах северных Курильских о-вов. В августе–ноябре 1997 г. уловы были представлены особями длиной 41–71 (57.0 ± 0.7) см и массой 1200–8800 (4035 ± 171) г. Самки доминировали над самцами в соотношении 8.4 : 1.0. Основу питания полиперы составляют рыбы (97.7% массы пищи), преимущественно Liparidae (85.3%), в том числе представители рода Careproctus (32.1%) и обыкновенный элассодискус Elassodiscus tremebundus (29.0%). Индивидуальная абсолютная плодовитость варьировала в пределах 36683−82992 (57367 ± 1558) икринок.
Симуширская полипера Polypera simushirae (далее полипера) – широкобореальный приазиатский вид; эндемик тихоокеанских вод центральной части Курильской гряды, где он обитает преимущественно на элиторальных глубинах (0–833 м), нередко встречается в пределах верхней батиали (Орлов, Питрук, 1996; Каталог …, 2000; Парин и др., 2014). Вид описан по особям, выловленным на литорали о-ва Симушир (Gilbert, Burke, 1912). Долгие годы сведения о его поимках относились к водам этого острова (Burke, 1930; Солдатов, Линдберг, 1930; Таранец, 1937; Шмидт, 1950; Matsubara, 1955; Линдберг, Красюкова, 1987). В дальнейшем полипера была поймана у северного побережья о-ва Хоккайдо (Ueno, 1971), а также у побережья о-ва Парамушир, вплоть до Первого Курильского пролива (Орлов, Бирюков, 2003), и даже у Юго-Восточной Камчатки (Орлов, 2010). В 1992–2002 гг. в рамках проведения научно-исследовательских работ по малоизученным рыбам материкового склона дальневосточных морей была получена информация по питанию (Орлов, Питрук, 1996; Токранов, 2000а, 2017, 2020), размерно-возрастному составу (Токранов, 2000б) и пространственному распределению (Токранов, 2000в; Орлов, Бирюков, 2003; Orlov, 2003) полиперы в водах восточного побережья северных Курильских о-вов.
В настоящей работе представлены данные, дополняющие имеющуюся информацию по полипере, а также первые сведения по её плодовитости.
МАТЕРИАЛ И МЕТОДИКА
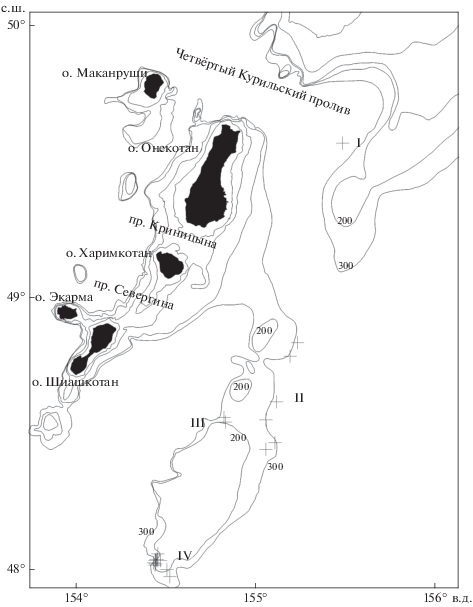
Пространственное распределение полиперы рассматривается по данным 22 донных тралений, в которых она присутствовала (рис. 1). Траления проводили в сентябре–октябре 1997 г. у восточного побережья северных Курильских о-вов Онекотан–Шиашкотан. Вертикальное и горизонтальное раскрытия трала составляли соответственно 6 и 18–25 м. Материалы по размерно-половому составу и питанию анализировали для всего района исследований и для четырёх пространственно разобщённых его участков: I – самый северный, находящийся напротив Четвёртого Курильского пролива (глубины 380–415 м, 1 траление, 1 особь); II – тянущийся вдоль островов (270–530 м, 4 траления, 6 экз.); III – на севере самого крупного подводного плато (180–240 м, 2 траления, 22 экз.); IV – на юге этого плато (170–303 м, 11 тралений, 46 экз.). Всего проанализировано 75 особей из 18 тралений.
Рис. 1.
Места поимок (+) симуширской полиперы Polypera simushirae в тихоокеанских водах северных Курильских о-вов в сентябре–октябре 1997 г.; I–IV – участки, (—) – изобаты 200 и 300 м.
У всех рыб измеряли длину по Смитту (FL). Содержимое желудков исследовали общепринятыми методами (Руководство …, 1961). В отдельных случаях массу пищевых объектов находили, исходя из их доли (в % на глаз) в пищевом комке с установленной массой, принимая, что полученные значения соответствуют долям по массе. Значение отдельных компонентов в питании полиперы оценивали по доле их массы (в % общей массы пищи). Яичники 39 самок (III−IV стадия зрелости) отбирали и фиксировали в 4%-ном растворе формальдегида. Камеральную обработку проводили весовым методом (Анохина, 1969). Освобождённые из формалина яичники выдерживали в течение нескольких часов в проточной воде, после чего обсушивали на фильтровальной бумаге. Массу яичников измеряли на электронных весах с точностью до 0.001 г. Индивидуальную абсолютную плодовитость (ИАП) определяли в пробах массой 1.882−5.459 г, взятых из средней части яичника; в каждой пробе измеряли диаметр 10 икринок.
Корреляцию показателей оценивали как очень сильную (>0.90), сильную (0.71–0.90), значительную (0.51–0.70), умеренную (0.30–0.50) и слабую (<0.30) (Лакин, 1973).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В районе исследований полипера встречалась в уловах донного трала на участке 47°58′–49°32′ с.ш. и 154°26′–155°30′ в.д. В пределах этих координат данный вид отмечали и другие авторы (Орлов, Питрук, 1996; Токранов, 2000в; Orlov, 2003). В августе–октябре 1997 г. численность полиперы была существенно ниже (до 17 экз/ч траления), чем в мае–декабре 1995–1997 гг. (до 323 экз/ч) (Токранов, 2000в) и в 1992–2000 гг. (до 100 экз/ч) (Orlov, 2003). В 1993–1999 гг. средняя численность полиперы в уловах составляла 1.2 экз/ч траления (Орлов, 1998; Orlov, 2005). Пространственная разобщённость тралений с уловами полиперы позволяет сравнить полученные данные на разных участках. На I участке улов полиперы составил 0.2 экз/ч траления, на II – 0.1–7.0 (в среднем 2.7) экз., на III – 11–14 (12.5) экз., на IV – 1–17 (7.8) экз. Эти данные соответствуют представлениям, согласно которым полипера наиболее часто встречается в районе подводного плато в южной части северного звена подводного хребта Витязя (Орлов, Питрук, 1996).
Полипера в уловах была представлена особями FL 41–71 (57.0 ± 0.7) см и массой 1200–8800 (4035 ± 171) г. Основу её численности составили размерные группы FL 51–55 (23.9% для самок и 24.0% оба пола), 56–60 (28.4 и 26.7%) и 61–65 (32.8 и 32.0%) см. Размерные ряды самок и в целом обоих полов совпадают (рис. 2). В сентябре–декабре 1995–1997 гг. длина полиперы в траловых уловах составляла 39–67 см, масса – 900–7200 г, доминировали особи 45–65 см (87%) и 1000–5000 г (88%) (Токранов, 2000б).
Рис. 2.
Размерный состав (FL) симуширской полиперы Polypera simushirae в уловах в тихоокеанских водах северных Курильских о-вов: (- ⚪ -) – самки, 67 экз.; (− ⬜ −) – самцы, 8 экз.; (−◇−) – оба пола, 75 экз.
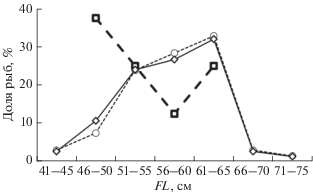
В наших уловах самки значительно преобладали по численности над самцами (8.4 : 1.0) и были крупнее их: FL 57.4 ± 0.7 (41.0–71.0) против 53.0 ± 2.5 (45.5–64.5) см, масса 4172 ± 173 (1200–8800) против 2888 ± 574 (1300–5200) г. Тогда как по обобщённым данным за сентябрь–октябрь 1993 и апрель–май 1995 гг. (Орлов, Питрук, 1996), наоборот, доминировали самцы (1.7 : 1.0); при этом средние значения их длины и массы (59.18 см и 5086 г) были больше, чем у самок (53.21 см и 3420 г). Отличия наших данных от полученных ранее, возможно, обусловлены тем, что в 1997 г. облавливаемые участки не совпадали с местами преимущественного обитания самцов, в том числе и более крупных.
При сравнении размерного и полового состава уловов полиперы на разных участках прослеживается тенденция увеличения в юго-западном направлении как средних значений длины и массы особей, так и доли самок (табл. 1).
Таблица 1.
Размерно-половой состав уловов симуширской полиперы Polypera simushirae на разных участках тихоокеанских вод северных Курильских о-вов
| Участок | Число рыб, экз. | Длина (FL), см | Масса, г | Доля рыб, % | |||
|---|---|---|---|---|---|---|---|
| min–max | M | min–max | M | Самки | Самцы | ||
| I | 1 | 50.5 | 1500 | 0 | 100 | ||
| II | 6 | 41–59 | 52.4 | 1200–4500 | 2992 | 83.3 | 16.7 |
| III | 22 | 46–65 | 57.1 | 1300–5800 | 3900 | 90.9 | 9.1 |
| IV | 46 | 45–71 | 57.6 | 1400–8800 | 4291 | 91.3 | 8.7 |
Полипера по типу питания является хищником-засадчиком, преимущественно ихтиофагом (Орлов, Питрук, 1996; Токранов, 2000а, 2017, 2020). Наши данные с этим согласуются – на долю рыб приходится 97.7% массы пищи (табл. 2). Основу рыбной пищи составляют липаровые (Liparidae) (85.3%) – преимущественно представители рода Careproctus (32.1%) и обыкновенный элассодискус Elassodiscus tremebundus (29.0%). Токранов (2000а) в сентябре–октябре 1996 г. получил близкие к нашим значения липаровых в питании полиперы (84.0%). В этой связи данный вид можно более точно охарактеризовать как преимущественного липарофага. В исследованном районе самым значимым объектом питания полиперы является элассодискус. Данный вид как наиболее важный кормовой объект полиперы отмечен и в 1993–1995 гг. – 16.51% по частоте встречаемости (Орлов, Питрук, 1996), а в 1996 г. – 64.8% массы пищи (Токранов, 2000а).
Таблица 2.
Состав пищи симуширской полиперы Polypera simushirae на разных участках тихоокеанских вод северных Курильских о-вов, % массы
| Компонент пищи и другие показатели | Участок | Весь район | |||
|---|---|---|---|---|---|
| I | II | III | IV | ||
| Amphypoda | 0.56 | 0.01 | |||
| Isopoda | 0.05 | 0.03 | |||
| Decapoda | 10.00 | 1.76 | 0.05 | 2.25 | 1.68 |
| Caridea, неопределённые остатки | 1.76 | 0.05 | 0.75 | 0.58 | |
| Pandalus sp. | 10.00 | 0.05 | |||
| Lebbeus groenlandica | 1.50 | 1.05 | |||
| Teuthida (Berreteutis magister) | 0.51 | 0.36 | |||
| Pisces | 90.00 | 96.01 | 99.74 | 96.94 | 97.64 |
| Cottidae: | 8.33 | 1.38 | 4.59 | 3.81 | |
| Hemilepidotus gilberti | 1.59 | 1.11 | |||
| Icelus sp. | 0.17 | 0.12 | |||
| I. canaliculatus | 0.17 | 0.12 | |||
| Triglops scepticus | 2.32 | 1.62 | |||
| неопределённые остатки | 8.33 | 1.38 | 0.34 | 0.84 | |
| Psychrolutidae (Malacocottus zonurus) | 1.61 | 1.13 | |||
| Liparidae: | 83.98 | 92.38 | 83.17 | 85.85 | |
| Elassodiscus tremebundus | 52.78 | 54.01 | 18.49 | 29.16 | |
| неопределённые остатки | 31.20 | 30.71 | 21.59 | 24.37 | |
| Careproctus: | 7.66 | 43.09 | 32.32 | ||
| C. furcellus | 10.66 | 7.47 | |||
| C. rastrinus | 2.03 | 1.43 | |||
| C. zachirus | 1.27 | 0.89 | |||
| C. cypsellurus | 9.15 | 6.42 | |||
| C. rozeofuscus | 6.71 | 1.84 | |||
| C. macrocephalus | 8.92 | 6.25 | |||
| неопределённые остатки | 0.95 | 11.06 | 8.02 | ||
| Blennidae | 3.03 | 0.83 | |||
| Неопределённые остатки рыб | 90.00 | 3.70 | 2.95 | 7.57 | 6.02 |
| Икра: | 1.67 | 0.21 | 0.25 | 0.28 | |
| Liparidae | 0.11 | 0.08 | |||
| неопределённая | 1.67 | 0.21 | 0.14 | 0.20 | |
| Число желудков, шт. | 1 | 6 | 22 | 46 | 75 |
| Число тралений | 1 | 4 | 2 | 11 | 18 |
На всех рассматриваемых участках в желудках полиперы доминируют рыбы (90.00–99.73% массы пищи), а на участках II–IV – липаровые (83.17–92.38%) (табл. 2). На участке II идентифицированные до вида липаровые представлены элассодискусом (52.78%). На участке III помимо элассодискуса (54.01%) отмечены также карепрокты (7.66%). На участке IV доля элассодискуса существенно ниже (18.49%) за счёт возросшей значимости карепроктов (43.09%), которые представлены пятью видами.
Исходя из формы и окраски тела, свойственных для ведущих малоподвижный образ жизни на дне или вблизи него рыб, Орлов и Питрук (1996) полагают, что полипера подстерегает свои жертвы в укрытиях или маскируясь под цвет грунта. Данные по составу пищи особей на участке III позволяют предположить, что полипера может вести и активный поиск жертв: на этом участке основу её питания составил элассодискус, который отсутствовал в траловых уловах. Такое несоответствие можно объяснить кормовыми миграциями особей от занимаемых ими территорий к местам скоплений элассодискуса и обратно.
В табл. 3 представлены виды с относительно высокой численностью, встречающиеся в уловах с полиперой. Отмеченные в питании полиперы мягкий бычок Malacocottus zonurus и большеглазый триглопс Triglops scepticus являются наиболее многочисленными видами (соответственно 104–361 и 47–437 экз/ч траления); их длина (11–29 и 16–28 см) позволяет стать жертвами полиперы, однако в её питании они имеют ничтожное значение (1.13 и 1.61% массы). При численности командорского кальмара Berretheutis magister 108–244 экз/ч траления его доля ещё ниже (0.36% массы пищи). Совершенно отсутствовал в желудках полиперы наиболее многочисленный (1139–9623 экз/ч) на склонах плато в рассматриваемый период северный однопёрый терпуг Pleurogrammus monopterygius. Не был отмечен в питании полиперы данный вид и в сентябре–октябре 1996 г. (Токранов, 2000а), но в 1993–1995 гг., наряду с элассодискусом и карепроктами, имел наибольшее значение в питании полиперы (Орлов, Питрук, 1996).
Таблица 3.
Видовой состав уловов с симуширской полиперой Polypera simushirae на разных участках тихоокеанских вод северных Курильских о-вов
| Вид | Участок | |||
|---|---|---|---|---|
| I | II | III | IV | |
| Berretheutis magister | 244 | 134 | 132 | 108 |
| Bathyraja maculata | 11 | 33 | 25 | – |
| B. interrupta | 11 | – | – | – |
| Albatrossia pectoralis | – | 12 | – | – |
| Theragra chalcogramma | 708 | 398 | 105 | 19 |
| Sebastes alutus | – | 763 | 393 | 624 |
| S. borealis | – | 35 | * | – |
| Sebastolobus macrocir | 19 | 35 | – | * |
| Pleurogrammus monopterygius | – | – | 1139 | 9623 |
| Hemilepidotus gilberti | – | 37 | 38 | 142 |
| Triglops scepticus | – | 47 | 205 | 437 |
| Dasycottus setiger | 35 | – | – | – |
| Malacocottus zonurus | 104 | 361 | 242 | 292 |
| Sarritor frenatus | * | 14 | – | – |
| Careproctus cypsellurus | – | 87 | – | – |
| C. furcellus | 12 | 34 | – | * |
| C. rastrinus | * | – | – | 10 |
| C. roseofuscus | 16 | – | 11 | * |
| C. macrocephalus | – | 23 | * | * |
| Elassodiscus tremebundus | – | 572 | – | 36 |
| Paraliparis grandis | 14 | * | – | – |
| Polipera simushirae | * | * | 13 | * |
| Bothrocarina microcephala | 13 | – | – | – |
| Lycodes albalinneatus | 21 | * | – | – |
| L. brunneofasciatus | 11 | * | – | * |
| Bothrocara brunnea | 11 | – | – | – |
| Zaprora selena | – | – | * | 19 |
| Atherestes evermanni | * | 13 | * | – |
| Всего видов | 25 | 45 | 17 | 32 |
| Число тралений | 1 | 4 | 2 | 11 |
Собранный на участке IV материал позволяет сравнить питание разных размерных групп полиперы (табл. 4). Основу питания особей FL 46–50 см составляют рогатковые (Cottidae) (43.0% массы пищи) и липаровые (32.3%) виды рыб, а также десятиногие ракообразные (Decapoda) (16.8%). По мере роста полиперы потребление липаровых возрастает, а вклад рогатковых и десятиногих ракообразных становится несущественным. Доля элассодискуса варьирует в пределах от 11.2 (у особей FL 56–60 см) до 35.8% (FL 51–55 см). Для особей FL 56–60 и 61–65 см карепрокты становятся самыми значимыми объектами питания (соответственно 48.6 и 49.2%). У рыб FL 61–65 см питание характеризуется наибольшим видовым разнообразием карепроктов.
Таблица 4.
Состав пищи симуширской полиперы Polypera simushirae разных размерных групп в тихоокеанских вод северных Курильских о-вов (участок IV), % массы
| Компонент пищи | Размерная группа (FL), см | |||
|---|---|---|---|---|
| 46–50 | 51–55 | 56–60 | 61–65 | |
| Isopoda | 0.13 | 0.02 | ||
| Decapoda: | 16.82 | 7.72 | 0.66 | 0.40 |
| Caridea, неопределённые остатки | 5.15 | 3.53 | 0.11 | |
| Lebbeus groenlandica | 11.67 | 4.19 | 0.66 | 0.29 |
| Teuthida (Berreteutis magister) | 5.07 | |||
| Pisces: | 83.18 | 86.10 | 99.21 | 99.30 |
| Cottidae: | 43.01 | 8.41 | 1.84 | |
| Hemilepidotus gilberti | 10.31 | 1.84 | ||
| Icelus sp. | 1.68 | |||
| I. canaliculatus | 1.71 | |||
| Triglops scepticus | 31.34 | 2.50 | ||
| неопределённые остатки | 1.36 | 2.52 | ||
| Psychrolutidae (Malacocottus zonurus) | 1.36 | |||
| Liparidae: | 32.29 | 73.33 | 88.69 | 90.83 |
| Elassodiscus tremebundus | 20.62 | 35.81 | 11.20 | 21.47 |
| неопределённые остатки | 11.67 | 16.71 | 28.92 | 20.17 |
| Careproctus: | 20.81 | 48.57 | 49.19 | |
| C. furcellus | 28.00 | 6.85 | ||
| C. zachirus | 2.57 | |||
| C. cypsellurus | 18.52 | |||
| C. macrocephalus | 11.43 | 7.92 | ||
| неопределённые остатки | 20.81 | 9.14 | 13.33 | |
| Неопределенные остатки рыб | 6.52 | 4.36 | 10.52 | 6.63 |
| Икра: | 1.11 | 0.28 | ||
| Liparidae | 1.11 | |||
| неопределённая | 0.28 | |||
| Число желудков, шт. | 6 | 9 | 11 | 16 |
Все исследованные самки полиперы (FL 41–71 см) были половозрелыми. Длина самок, яичники которых находились на III−IV стадии зрелости, варьирует в пределах 46.0–65.5 (57.41 ± 0.77) см, масса – 1800–6000 (4124 ± 180) г; масса их яичников – 102.6−562.8 (309 ± 19 г). Корреляция между длиной и массой тела (r = 0.90), как и между длиной и массой яичников (0.73), оценивается как сильная. Зависимости описываются соответственно экспоненциальным (y = 140.98e0.0581x, R2 = 0.8181) и степенным (y = 2E − 06x4.6564, R2 = = 0.5952) уравнениями (рис. 3а, 3б).
Рис. 3.
Зависимость массы тела (а), массы яичников (б), абсолютной индивидуальной плодовитости (АИА) (в) и диаметра ооцитов (г) от длины тела (FL) симуширской полиперы Polypera simushirae в тихоокеанских водах северных Курильских о-вов.
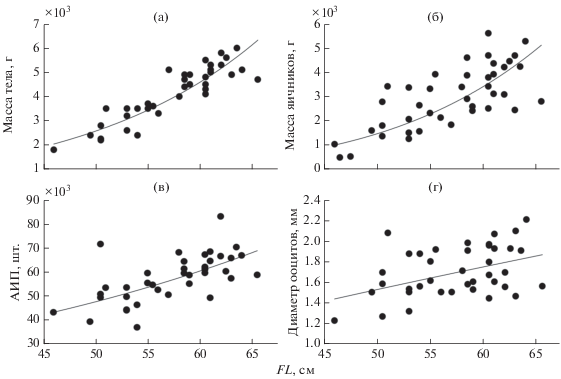
ИАП полиперы в среднем составляет 57367 ± 1558 икринок и варьирует от 36683 шт. у самки FL 54 см и массой 3500 г до 82992 шт. у самки FL 62 см и массой 5300 г (рис. 3в). Корреляция между длиной и ИАП значительная (0.64). Зависимость описывается экспоненциальным уравнением y = = 14224e0.024x (R2 = 0.4302).
Средний диаметр ооцитов равен 1.70 ± 0.039 мм; минимальное значение (1.23 мм) отмечено у самки FL 46 см и 1800 г, максимальное (2.2 мм) – FL 64 см и 5100 г (рис. 3в). Между длиной тела и диаметром икринок корреляция умеренная (r = = 0.416). Зависимость описывается степенным уравнением y = 0.0855x0.7365 (R2 = 0.1922).
Список литературы
Анохина Л.Е. 1969. Закономерности изменения плодовитости рыб. М.: Наука, 295 с.
Каталог позвоночных Камчатки и сопредельных морских акваторий. 2000. Петропавловск-Камчатский: Камчат. печат. двор, 165 с.
Лакин Г.Ф. 1973. Биометрия. М.: Высш. шк., 343 с.
Линдберг Г.У., Красюкова З.В. 1987. Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Ч. 5. Л.: Наука, 526 с.
Орлов А.М. 1998. Демерсальная ихтиофауна тихоокеанских вод северных Курильских островов и юго-восточной Камчатки // Биология моря. Т. 24. № 3. С. 146−160.
Орлов А.М. 2010. Количественное распределение демерсального нектона тихоокеанских вод северных Курильских островов и юго-восточной Камчатки. М.: Изд-во ВНИРО, 335 с.
Орлов А.М., Бирюков И.А. 2003. Новые данные по распространению симуширской полиперы Polypera simushirae (Liparidae, Scorpaeniformes) у Курильских островов // Бюл. МОИП. Отд. биол. Т. 108. Вып. 5. С. 78−80.
Орлов А.М., Питрук Д.Л. 1996. Материалы по питанию симуширской полиперы Polypera simushirae (Liparidae) и замечания о её распространении в районе северных Курильских островов // Вопр. ихтиологии. Т. 36. № 6. С. 821–826.
Парин Н.В., Евсеенко С.А., Васильева Е.Д. 2014. Рыбы морей России: аннотированный каталог. М.: Т-во науч. изд. КМК, 733 с.
Руководство по изучению питания рыб в естественных условиях. 1961. М.: Изд-во АН СССР, 263 с.
Солдатов В.К., Линдберг Г.У. 1930. Обзор рыб дальневосточных морей // Изв. ТИНРО. Т. 5. 563 с.
Таранец А.Я. 1937. Краткий определитель рыб советского Дальнего Востока и прилежащих вод // Там же. Т. 11. 200 с.
Токранов А.М. 2000а. Питание липаровых рыб (Liparidae) в тихоокеанских водах юго-восточной Камчатки и северных Курильских островов // Вопр. ихтиологии. Т. 40. № 4. С. 530–536.
Токранов А.М. 2000б. Размерно-возрастной состав липаровых рыб (Liparidae) в тихоокеанских водах юго-восточной Камчатки и северных Курильских островов // Там же. Т. 40. № 3. С. 347–352.
Токранов А.М. 2000в. Видовой состав и пространственное распределение липаровых рыб (Liparidae) в тихоокеанских водах юго-восточной Камчатки и северных Курильских островов // Там же. Т. 40. № 2. С. 176–186.
Токранов А.М. 2017. Пищевая специализация липаровых рыб (Liparidae, Pisces) в тихоокеанских водах северных Курильских островов и юго-восточной Камчатки // Матер. Всерос. науч. конф. “Современные проблемы экологии и эволюции”. Ульяновск: Изд-во УлГПУ. С. 366–371.
Токранов А.М. 2020. Трофические группировки донных и придонных рыб различных семейств отряда Scorpaeniformes в прикамчатских водах // Матер. VIII междунар. Балт. мор. форума. Т. 3. Водные биоресурсы, аквакультура и экология водоемов. Калининград: Изд-во КГТУ. С. 107–117.
Шмидт П.Ю. 1950. Рыбы Охотского моря. М.; Л.: Изд-во АН СССР, 370 с.
Burke C.V. 1930. Revision of the fishes of the family Liparidae // Bull. US Nat. Mus. № 150. P. 1–204.
Gilbert C.H., Burke C.V. 1912. New Cyclogasterid fishes from Japan // Proc. US Nat. Mus. V. 42. P. 351–380.
Matsubara K. 1955. Fish morphology and hierarchy. V. I−III. Tokyo: Ishizaki Shoten, 1605 p.
Orlov A.M. 2003. Impact of eddies on spatial distributions of groundfishes along waters off the northern Kuril Islands, and southeastern Kamchatka (north Pacific Ocean) // Ind. J. Mar. Sci. V. 32. № 2. P. 95−113.
Orlov A.M. 2005. Bottom trawl-caught fishes and some features of their vertical distribution in the Pacific waters off the north Kuril Islands and south-east Kamchatka, 1993−1999 // Aqua J. Ichthyol. Aquat. Biol. V. 9. № 4. P. 139−160.
Ueno T. 1971. List of marine fishes from the waters of Hokkaido and its adjacent regions // Sci. Rept. Hokkaido Fish. Exp. St. V. 13. P. 61–102.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии