

Вопросы ихтиологии, 2022, T. 62, № 3, стр. 313-322
Сезонное распределение и размерный состав многоиглого керчака Myoxocephalus polyacanthocephalus (Cottidae) в российских водах Японского моря
В. В. Панченко 1, *, А. Н. Вдовин 2
1 Национальный научный центр морской биологии Дальневосточного отделения РАН − ННЦМБ ДВО РАН
Владивосток, Россия
2 Тихоокеанский филиал Всероссийского научно-исследовательского института
рыбного хозяйства и океанографии – ТИНРО
Владивосток, Россия
* E-mail: vlad-panch@yandex.ru
Поступила в редакцию 30.03.2021
После доработки 07.04.2021
Принята к публикации 07.04.2021
- EDN: ZSVXOV
- DOI: 10.31857/S0042875222030146
Аннотация
В российских водах Японского моря многоиглый керчак Myoxocephalus polyacanthocephalus встречается на глубинах 6–640 м при температуре –1.2…+15.5°C. Его сезонные миграции носят преимущественно батиметрический характер. Широтные миграции явно прослеживаются только в северо-западной части Татарского пролива и в осенний период идут в направлении на юг, в весенний – на север. Зимой многоиглый керчак в массе избегает охлаждённую до отрицательных значений температуры верхнюю часть шельфа, предпочитая его нижнюю часть и верхний отдел материкового склона. Летом концентрируется в верхней и средней частях шельфа. Молодь более эвритермна, чем взрослые особи, и во все сезоны придерживается шельфовой зоны.
Многоиглый керчак Myoxocephalus polyacanthocephalus – наиболее крупный представитель семейства рогатковых (Cottidae), массовый элиторальный, широкобореальный вид, обитающий в открытых водах Тихого океана, в Беринговом, Охотском, и Японском морях, а также встречающийся в арктических водах (Линдберг, Красюкова, 1987; Amaoka et al., 1995; Новиков и др., 2002; Mecklenburg et al., 2002; Федоров и др., 2003; Соколовский и др., 2007; Парин и др., 2014). В настоящее время в наибольшей степени распределение этого вида изучено в северных частях ареала (Токранов, 1981, 1986; Борец, 1997; Гудков, Хованский, 2001; Орлов, 2010; Токранов, Орлов, 2013; Андронов, Датский, 2014; Матвеев, Терентьев, 2016). Распределение многоиглого керчака в Японском море исследовали лишь в отдельные сезоны на юге российских вод – в зал. Петра Великого (Панченко, 1999; Панченко, Пущина, 2004; Панченко, Зуенко, 2009) и отчасти в водах Северного Приморья (Калчугин, 1998). Однако ни в одной из известных нам работ пространственно-временнáя динамика размерного состава не рассмотрена.
Цель работы – охарактеризовать закономерности сезонного распределения многоиглого керчака на разных стадиях развития в акватории всей российской зоны Японского моря.
МАТЕРИАЛ И МЕТОДИКА
В работе использованы материалы донных траловых съёмок и контрольных тралений, выполненных ТИНРО в российских водах Японского моря в разные сезоны 1981–2017 гг. Проанализированы данные 10618 тралений на шельфе и материковом склоне на глубинах 2–935 м, из них 5542 сопровождали измерением придонной температуры воды.
Траления выполняли донными тралами с мягким грунтропом разных конструкций со скоростью 1.8–3.5 (в среднем 2.7) узла. Для получения сравнимых результатов уловы многоиглого керчака в каждом трале пересчитывали на плотность по формуле: P = B/S, где P – плотность (удельная численность), экз/км2; B – улов, экз.; S – площадь траления, км2; коэффициенты уловистости при пересчёте не вводили.
При анализе сезонного распределения учитывали деление на гидрологические сезоны по классификации Зуенко (1994): зимний период – январь–февраль, весенний – март–апрель, летний – июнь–сентябрь, осенний – ноябрь–декабрь; май – переходный месяц между весенним и летним сезонами, октябрь – между летним и осенним. Однако, по нашим данным, в декабре многоиглый керчак в массе уже смещён к местам зимовки, а в марте ещё не покидает их. Это вполне закономерно, так как в декабре деятельный слой Японского моря уже охлаждён до близких к зимним месяцам значений, а максимальное его охлаждение происходит, как правило, в феврале–марте (Лучин, 2007). В итоге мы приняли следующую хронологию сезонной ритмики распределения многоиглого керчака: зима – декабрь–март, весна – апрель–май, лето – июнь–сентябрь, осень – октябрь–ноябрь.
Анализ пространственного распределения выполнен с использованием программного пакета Surfer. Размерный состав многоиглого керчака охарактеризован по результатам промеров (абсолютная длина – TL) 13630 экз. Число тралений в разные сезоны по диапазонам глубин и число изученных особей приведены в таблице.
Число тралений (N) и изученных особей (n) многоиглого керчака Myoxocephalus polyacanthocephalus в разные сезоны 1981−2017 гг. в российских водах Японского моря
| Глубины, м | Зима | Весна | Лето | Осень | ||||
|---|---|---|---|---|---|---|---|---|
| N | n, экз. | N | n, экз. | N | n, экз. | N | n, экз. | |
| 2–5 | 55 | 18 | ||||||
| 6–10 | 2 | 4 | 352 | 2 | 68 | |||
| 11–20 | 9 | 43 | 67 | 709 | 87 | 136 | 13 | |
| 21–40 | 23 | 11 | 303 | 319 | 1423 | 949 | 189 | 187 |
| 41–60 | 74 | 37 | 235 | 467 | 909 | 1040 | 127 | 120 |
| 61–80 | 65 | 12 | 226 | 872 | 899 | 1626 | 125 | 463 |
| 81–100 | 62 | 29 | 193 | 518 | 368 | 283 | 76 | 214 |
| 101–150 | 115 | 47 | 312 | 803 | 367 | 169 | 107 | 282 |
| 151–200 | 75 | 98 | 209 | 798 | 117 | 22 | 89 | 335 |
| 201–250 | 64 | 218 | 244 | 1193 | 157 | 20 | 88 | 208 |
| 251–300 | 86 | 297 | 134 | 284 | 75 | 9 | 72 | 90 |
| 301–400 | 125 | 180 | 281 | 721 | 133 | 1 | 97 | 130 |
| 401–500 | 82 | 48 | 220 | 205 | 93 | 65 | 52 | |
| 501–700 | 60 | 21 | 230 | 74 | 83 | 64 | 9 | |
| 701–935 | 8 | 61 | 12 | |||||
| Всего | 850 | 998 | 2695 | 6321 | 5740 | 4208 | 1333 | 2103 |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В российских водах Японского моря многоиглый керчак отнесён к элиторально-сублиторальной группировке рыб, представители которой обитают в пределах всего шельфа, однако предпочитают его нижнюю часть и верхний отдел материкового склона, но встречаются и глубже. Летом для таких видов характерно перемещение в сторону меньших глубин, зимой – в сторону бо́льших (Соломатов, 2008).
Наиболее широко по акватории многоиглый керчак распределён в летний нагульный период за счёт большего освоения северных областей (рис. 1). В это время в водах Татарского пролива отмечены его значительные скопления. Для данного района характерно наличие широкой шельфовой зоны, в которую со стороны материкового склона многоиглый керчак смещается после зимовки. Широкий шельф имеется и в южной части района – в зал. Петра Великого, где летом также отмечены скопления многоиглого керчака, хотя и не столь значительные. В зал. Петра Великого многоиглый керчак летом был распределён по акватории более равномерно, чем в кутовой части Татарского пролива, где его относительно высокие уловы отмечали только с материковой стороны. В островной акватории скопления многоиглого керчака располагались южнее кутовой части пролива, в которой они вовсе не были отмечены. В центральном участке материковой акватории, в водах Северного Приморья, наиболее высокие уловы вида отмечали, как правило, в его верхней части.
Рис. 1.
Пространственное распределение многоиглого керчака Myoxocephalus polyacanthocephalus в российских водах Японского моря по сезонам: а – зима, б – весна, в – лето, г – осень.
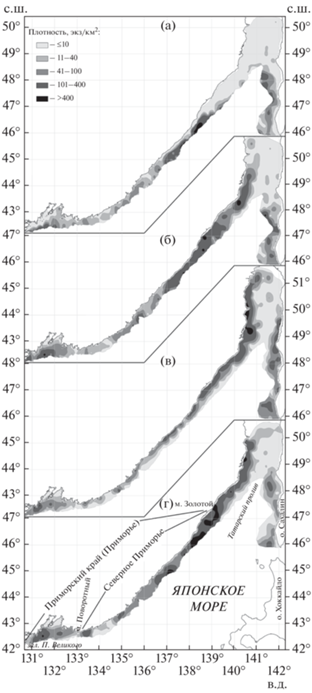
Во время осеннего похолодания наиболее заметное перераспределение скоплений многоиглого керчака отмечено в материковой акватории Татарского пролива (рис. 1г). Самые северные скопления у материкового побережья вообще покидали этот район. Судя по резкому возрастанию плотности южнее кутового района, происходило эшелонированное смещение всех рыб, нагуливавшихся на западном участке кутовой части. К декабрю эта миграция в основном завершалась. Вероятно, при смене сезонов повышается миграционная активность и агрегированность группировок многоиглого керчака.
Отход многоиглого керчака на зимовку из кутовой части Татарского пролива связан с его мелководностью. В холодный период у побережья о-ва Сахалин предпочитаемый район обитания многоиглого керчака существенно не изменился (рис. 1а), произошла лишь миграция рыб на зимовку в сторону глубоководной зоны. Ким Сен Ток (2001) ранее также отмечал, что многоиглый керчак у побережья о-ва Сахалин не относится к видам, смещающимся на зимовку в более северные или в более южные участки. Однако у материкового побережья произошла откочёвка гораздо далее на юг. В результате в прилегающей с севера к траверзу м. Золотой глубоководной акватории скопления многоиглого керчака не отмечены. Ранее уже было показано, что в Татарском проливе многоиглый керчак в зимовальный период с материковой стороны образует скопления в более низких широтах (ниже 48° с.ш.), чем со стороны островного побережья (Ким Сен Ток, 2001).
В весенний период проходил процесс смещения рыб к местам летнего нагула. В результате обратной миграции по направлению к кутовой части материкового побережья Татарского пролива в его акватории повышалась плотность многоиглого керчака в основном за счёт снижения его численности в прилегающих водах Северного Приморья (рис. 1б). В остальной акватории столь существенных межрайонных миграций не прослеживалось.
Особенности распределения рыб в различных участках акватории во многом обусловлены особенностями режима вод. По окончании тёплого периода года в северной части Татарского пролива формируется подповерхностный слой вод с пониженной температурой и солёностью, опускающийся за счёт зимней конвекции в придонные области и образующий холодный подстилающий слой (Зуенко, 2008). Наиболее ощутимо влияние этого фактора проявляется до 100-метровой изобаты, но затрагивает и более глубокие слои воды. Южнее м. Золотой влияние северного режима вод постепенно ослабевает и ниже 46° с.ш. для вод Приморья типичен менее суровый гидрологический режим, в особенности для расположенного на юге зал. Петра Великого. Акватория Татарского пролива у юго-западного побережья Сахалина в меньшей степени подвержена влиянию подповерхностного слоя вод, так как находится под воздействием тёплого Цусимского течения, проникающего вдоль Японских о-вов. Можно резюмировать, что выраженные широтные миграции многоиглого керчака прослеживаются в том участке материковой акватории, в котором формируется холодный подстилающий слой. Ниже, в зоне смешения вод, характерных для материковой части Татарского пролива и для вод Приморья, между м. Золотой и 46° с.ш. скопления рыб сохраняются постоянно. Летом на этом участке в связи с откочёвкой части рыб вдоль побережья на север их плотности менее значительны, чем в остальные сезоны.
Заметим, что у многоиглого керчака нерест повсеместно протекает в холодный период года (Токранов, 1984, 1986, 1988; Борец, 1997; Новиков и др., 2002; Панченко, Пущина, 2004; Датский, 2017). В российских водах Японского моря он, как и в северных районах, проходит, видимо, в нижней части шельфа. Судя по описанным сезонным миграциям, материковая акватория Татарского пролива выше м. Золотой не используется многоиглым керчаком в качестве нерестовых участков. Сезонные различия в пространственном распределении многоиглого керчака, описанные для тихоокеанских вод северных Курильских о-вов и Юго-Восточной Камчатки (Орлов, 2010; Токранов, Орлов, 2013), отчасти связываются указанными авторами с нерестовыми миграциями в холодный период года.
Общим для всех сезонов является стабильно низкая плотность многоиглого керчака в нижней области Северного Приморья, в районе 43° с.ш., а также снижение его плотности в центральной части зал. Петра Великого западнее 132° в.д. при наличии более плотных скоплений в прилегающих с запада и востока водах (рис. 1). В первом районе, между 42-й и 44-й параллелями, выражена динамика водных масс (вплоть до соприкосновения тёплых и холодных вод), в связи с чем ярко проявляется меандрирование фронтов (Яричин, Покудов, 1982; Никитин, Дьяков, 2016). В связи с особенностями гидрологии, в этом районе наблюдается разрыв в распределении многих видов рыб, и ихтиогеографическое районирование морских вод Приморья зачастую ориентировано на 43-ю параллель с.ш. (Дударев и др., 1998; Вдовин и др., 2004). Снижение плотности в центральной части залива Петра Великого, также отмечаемое для многих видов рыб, обусловлено прохождением здесь зоны раздела циркуляции вод (Зуенко, 2008). При сравнении восточной и западной акваторий залива заметно, что во все сезоны более значительные плотности многоиглого керчака отмечены в его западной части, хотя зал. Петра Великого считается южной границей его распространения (Линдберг, Красюкова, 1987; Новиков и др., 2002; Парин и др., 2014), что подразумевает снижение численности обитающих здесь рыб в направлении на юго-запад, к границе с Северной Кореей. Указанное выделение южной границы, видимо, произошло в результате отсутствия доступных достоверных сведений о составе рыбного населения в прилегающих северокорейских водах. Хотя зал. Петра Великого находится на стыке климатических и зоогеографических зон, испытывая влияние соседних акваторий, в целом его гидрологический режим типичен для вод Приморья (Зуенко, 1994, 2008). Здесь в период исследований мы отмечали многоиглого керчака на предпочитаемых им глубинах, в том числе и в проведённом наиболее близко к границе с Северной Кореей тралении в координатах 42°18′ с.ш. 130°45′ в.д. (МРТК “Янтарь”, 26.08.2010 г., глубина 60 м). Однако эта поимка многоиглого керчака не является самой южной, так как здесь на располагающемся ниже свале глубин отсутствуют участки, пригодные для работы используемыми нами орудиями лова. Наиболее южная поимка отмечена в координатах 42°16′ с.ш. 131°00′ в.д. (РКМРТ “Бухоро”, 06.04.2000 г., глубина 372 м). Здесь, в районе 131° в.д., располагались самые южные точки тралений. Учитывая уловы многоиглого керчака вплоть до границы с корейскими водами, можно с большой вероятностью предположить, что этот вид обитает и несколько далее на юг – в северокорейских водах вблизи их границы с российскими.
Сезонная изменчивость батиметрического распределения многоиглого керчака в отличие от широтного не имеет столь явных региональных различий (рис. 2). Наиболее сходным батиметрическое распределение было в зимний период, после прошедшего “стекания” большей части рыб с шельфа на материковый склон. Минимальная глубина поимок многоиглого керчака составила 25 м (рис. 2а). В это время в верхней части шельфа повсеместно преобладала отрицательная температура. Её среднее значение в целом на глубинах до 60 м было также отрицательным. Таким образом, многоиглый керчак зимой избегал выхоложенной верхней части шельфа. Глубже 80 м, где выхоложенные до минусовых значений участки встречались лишь эпизодически, его уловы стали возрастать, а наибольшие плотности отмечали ниже 150-метровой изобаты до 385 м при слабоположительной температуре в основном в пределах 0.6–1.2°C. Отмеченные плотности многоиглого керчака в нижней части шельфа отчасти можно связать с подходом рыб на нерестилища, а на материковом склоне, ниже глубин ~200 м, – с зимовальными скоплениями. В водах Камчатки зимовка основной массы этого вида проходит у нижней границы шельфа, находящейся под влиянием тёплой промежуточной водной массы с температурой >0°С (Борец, 1997). В основном в нижней части шельфа и в прилегающих участках материкового склона при положительной температуре (1.2–2.5°C) его наибольшие уловы в зимний период отмечены и в тихоокеанских водах северных Курильских о-вов и Юго-Восточной Камчатки (Токранов, Орлов, 2013).
Рис. 2.
Средняя плотность многоиглого керчака Myoxocephalus polyacanthocephalus (−◼−) и средняя температура (–$ \times |$–) в разных диапазонах глубин в российских водах Японского моря по сезонам: а – зима, б – весна, в – лето, г – осень.
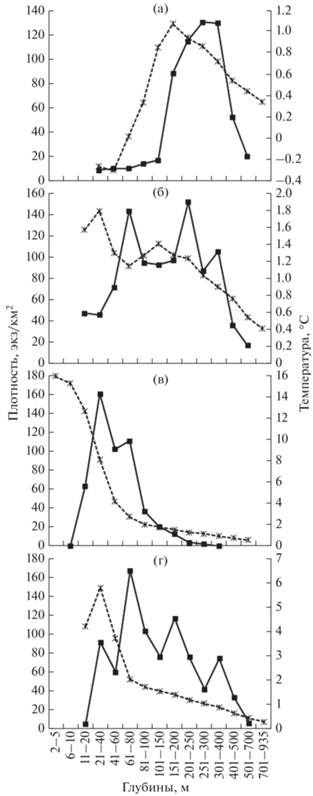
В зимние месяцы многоиглый керчак был отмечен до 628 м в температурном диапазоне ‒1.2…+4.1°C. Заметим, что в весенний период, в начале апреля, максимальная глубина поимки особи, явно не отошедшей ещё с мест зимовки, была несколько большей – 640 м. На сходных глубинах (630 м) многоиглый керчак обитает зимой и в тихоокеанских водах северных Курильских о-вов и Юго-Восточной Камчатки (Токранов, Орлов, 2013). По данным некоторых авторов, в исследуемом нами районе многоиглый керчак опускается до бо́льших глубин. По мнению Соколовского с соавторами (2007), в российских водах Японского моря максимальная глубина его обитания достигает 775 м, а по данным Ким Сен Тока (2001), исследовавшего япономорские воды о-ва Сахалин, он доходит зимой по материковому склону до 825 м. Мы тоже располагаем данными о нахождении многоиглого керчака в нескольких тралах, поднятых с глубин более 640 м. Из 149 проанализированных ловов, проведённых в пределах 641–935 м, в шести (на глубинах 645–715 м) присутствовали единичные особи этого вида. Однако, как показал проведённый нами анализ траловых данных для уточнения максимальных размеров и глубин обитания рыб российской зоны Японского моря (Панченко и др., 2016), поимки многоиглого керчака на глубинах свыше 640 м нельзя считать достоверными, так как каждому из этих ловов предшествовали работы на меньших глубинах, в которых присутствовали особи многоиглого керчака. Полностью очистить трал от гидробионтов до следующей его постановки в период съёмки удаётся далеко не всегда, поэтому остаётся вероятность нахождения особей из предыдущего улова.
Наиболее широко многоиглый керчак был распределён в период весенних миграций с мест зимовки к местам летнего нагула (рис. 2б) и в период обратных миграций в осенний период (рис. 2г). Соответственно, в начале весеннего сезона рыбы предпочитали ещё в основном близкие к зимним, относительно глубокие слои воды, а в конце сезона смещались мельче, на сходные с летом глубины. Осенью эти тенденции менялись на противоположные. Минимальные глубины обитания многоиглого керчака весной и осенью приближались к таковым в летний период, а максимальные – к зимним. Весной минимальная глубина его поимки составила 15 м, максимальная, как уже указывалось, – 640 м. Осенью первый показатель составил 12 м, второй приближался к 600 м.
Самый узкий батиметрический диапазон многоиглый керчак занимал летом. В это время он в наибольшей степени тяготел к мелководной зоне, встречаясь на глубинах 6–328 м при температуре 0.3–15.5°C, но концентрируясь в основном в шельфовой зоне (рис. 2в). Сходное распределение этого вида в летний период характерно и для вод Западной Камчатки. Здесь он в этот сезон обитает до 300-метровой изобаты, концентрируясь в основном на глубинах 20–100 м (Токранов, 1981; Борец, 1997; Матвеев, Терентьев, 2016). В российской зоне Японского моря по полученным нами усреднённым данным многоиглый керчак летом отдавал предпочтение диапазону 21–40 м, далее следовали глубины 41–60 и 61–80 м. Несмотря на сходство тенденций летнего батиметрического распределения многоиглого керчака на разных участках акватории Японского моря, имеются и различия. На большей части акватории Приморья, за исключением зал. Петра Великого и зоны смешения вод ниже м. Золотой, батиметрические предпочтения в целом соответствовали вышеуказанным. С продвижением далее на север, у восточного побережья Татарского пролива, прослеживалось некоторое увеличение плотностей на относительно небольших, близких к 20-метровой изобате глубинах. У побережья о-ва Сахалин также по величине плотности рыб лидировали три указанных диапазона, но на 61–80 м этот показатель был несколько выше, чем на 41–60 и 21–40 м. На юге района, в зал. Петра Великого, сдвиг в глубоководные слои более заметен: в отличие от остальной акватории до 40-метровой изобаты скопления многоиглого керчака здесь не отмечены.
В российских водах Японского моря среди керчаков, как и в целом среди рогатковых, по биомассе доминирует многоиглый керчак, за исключением зал. Петра Великого, где по этому показателю абсолютно лидирует достигающий близких размеров керчак-яок M. jaok (Калчугин и др., 2016). Суммарная доля этих двух видов керчаков, имеющих сходный спектр питания (Пущина и др., 2016), гораздо выше, чем в любом из остальных районов российской зоны Японского моря. Керчак-яок в зал. Петра Великого образует летом скопления в том числе и на предпочитаемых в остальной акватории многоиглым керчаком глубинах (Панченко, Зуенко, 2009). Возможно, указанные факторы в тёплый период года способствуют смещению многоиглого керчака в водах залива глубже, чем в других районах описываемой акватории.
В связи с выявленными батиметрическими предпочтениями вполне логично, что максимальные глубины распространения многоиглого керчака в летний период оказались наибольшими в зал. Петра Великого. Однако для залива оказались характерными и минимальные глубины обитания, но это может быть связано с недостатком данных по мелководной зоне остальных районов. Глубины до 5 м исследовали только в зал. Петра Великого, основную массу тралений в диапазоне 6–10 м (более 80%) также провели в этом районе.
Обращает на себя внимание более высокая в большинстве диапазонов плотность рыб в осенний и весенний периоды (рис. 2). Соответственно, при умножении показателей плотности на площадь учтённая численность рыб в периоды перемещения с мест зимовки к местам летнего нагула и обратно выше, чем в летний и зимний периоды. Площадь российских вод Японского моря на глубинах до 700 м составляет ~100 тыс. км2 (Калчугин и др., 2016). При умножении полученных нами значений плотности на приведённые в указанной работе площади по диапазонам глубин учтённая численность многоиглого керчака зимой составила 4.0, весной – 8.9, летом – 5.5, осенью – 8.8 млн экз. Наименьшее число учтённых рыб в зимний период отчасти связано с особенностями сбора материала. По этому сезону мы располагаем в основном сведениями по ловам 1980–1990-х гг., когда, по всей видимости, многоиглый керчак находился большей частью на спаде численности, поскольку в это время и в остальные сезоны его скопления отмечали гораздо реже, чем в более поздний период. Однако и летом учтённая численность оказалась гораздо ниже, чем весной и осенью. Можно заключить, что в период межсезонных миграций, когда идёт разрушение одних и формирование других структур вод, учитываемость многоиглого керчака лучшая, чем в стабильные в гидрологическом отношении зимние и летние сезоны. В тихоокеанских водах северных Курильских о-вов и Юго-Восточной Камчатки после увеличения в постзимовальный весенний период также отмечено снижение его уловов летом и последующее возрастание осенью (Орлов, 2010; Токранов, Орлов, 2013). Однако отмеченные здесь указанными исследователями низкие концентрации летом отчасти связаны с отсутствием у них сведений о распределении рыб на глубинах менее 76 м. Известно (Федоров, 2000), что у северных Курильских о-вов предпочитаемый диапазон обитания многоиглого керчака составляет 40–250 м. К наименьшим глубинам он здесь, как и в других районах, тяготеет в тёплый период года.
Следует отметить, что при обсуждении рассчитанной нами для различных сезонов численности многоиглого керчака в российских водах Японского моря речь идёт о его учитываемом количестве, которое, конечно же, ниже реального. Общеизвестно, что при съёмках постоянно происходит недоучёт всех размерных групп, что связано с особенностью работы используемых орудий лова. Для уменьшения погрешности учёта рыб при расчётах их запасов применяют коэффициенты уловистости, зачастую дифференцированные для разных размерных групп, так как уловистость средне- и крупноразмерных рыб выше, чем молоди, даже при использовании в кутце трала вставки дели с мелкоразмерной ячеёй. Очевидно, что особи меньшего размера недоучитываются в наибольшей степени. С уменьшением размеров уловистость молоди падает и поимки наиболее мелкоразмерных особей, в особенности сеголеток, могут быть только случайными. В период исследований мы отметили особей многоиглого керчака TL 6–81 см, в уловах обычно преобладали рыбы TL 36–50 см.
Тенденции распределения размерных групп многоиглого керчака в разных участках исследуемого района были сходными (рис. 3). Наиболее мелкоразмерные особи встречены в летний период (рис. 3в) в зал. Петра Великого. Вероятно, это вылупившиеся весной (Соколовский, Соколовская, 1997) и к лету перешедшие на донный образ жизни сеголетки. До достижения половой зрелости самцы многоиглого керчака дорастают до 35–40 см, самки – до 45–50 см (Панченко, Пущина, 2004). Судя по размерному составу, во все сезоны как неполовозрелые, так и взрослые рыбы обитали практически во всех диапазонах глубин, однако мелкоразмерная молодь до достижения определенных размеров предпочитала относительно небольшие глубины. Самые мелкоразмерные особи TL 5.2 и 6.5 см были отмечены летом на глубинах до 10 м. Глубже 21 м минимальные размеры рыб постепенно возрастали и особей столь малой длины не отмечалось. На глубине свыше 75 м не наблюдали уже рыб TL < 20 см. В отличие от молоди особи TL > 50 см, т.е. однозначно относящиеся к взрослым рыбам, избегали минимальных глубин обитания вида, начав встречаться только с глубины 19 м. Ниже 80-метровой изобаты прослеживалась тенденция уменьшения как минимальных, так и максимальных размеров, однако у молоди она была выражена в гораздо большей степени, за счёт чего средний размер рыб с возрастанием глубины увеличивался.
Рис. 3.
Размерный состав (абсолютная длина TL) многоиглого керчака Myoxocephalus polyacanthocephalus в уловах донного трала в российских водах Японского моря в разных диапазонах глубин по сезонам: а – зима, б – весна, в – лето, г – осень; (-) – среднее значение, (│) – пределы варьирования показателя.
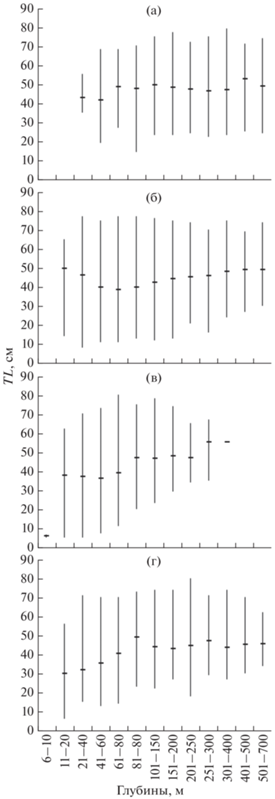
В осенний и весенний периоды сохранилась тенденция предпочтения мелкоразмерной молодью многоиглого керчака верхней и средней частей шельфа (рис. 3б, 3г). В зимний период наиболее мелкоразмерные особи также отмечались лишь в шельфовой зоне до 90-метровой изобаты (рис. 3а).
Обитая летом в относительно мелководной зоне, молодь многоиглого керчака использует более прогретые слои воды, чем взрослые рыбы. Зимовка же, протекая в основном в средней, а, возможно, и в верхней частях шельфа, проходит в водах с близкими к 0°C и отрицательными значениями температуры. Ранее для северо-восточной части Японского моря уже отмечалось, что молодь этого вида предпочитает зимовать на шельфе, не опускаясь на материковый склон (Ким Сен Ток, 2001). Для прикамчатских вод также указано, что в зимнее время при отрицательных температурах на глубинах менее 100 м встречается только молодь многоиглого керчака (Борец, 1997). На возрастание во все сезоны размеров этого вида с увеличением глубины обращено внимание и в тихоокеанских северокурильских водах (Токранов, Орлов, 2013). Мы при наиболее низких температурах (≤–1°C) отмечали только особей TL ≤ 32 см, т.е. принадлежавших к неполовозрелым рыбам. Особи, размер которых позволял однозначно отнести их к взрослым рыбам, начали встречаться лишь при температуре –0.2°C.
Можно заключить, что молодь многоиглого керчака, особенно на начальном этапе развития, является более эвритермной, чем взрослые особи. У многих рыб, совместно обитающих с многоиглым керчаком, с возрастом прослеживается стремление к бо́льшим глубинам с относительно стабильным температурным фоном (Вдовин, Зуенко, 1997). Проявляющееся с возрастом стремление холоднокровных животных в биотопы с относительно невысокой и стабильной температурой способствует замедлению и стабилизации в их организме обменных процессов, что приводит к увеличению продолжительности жизни (Бретт, Гроувс, 1983; Радзинская и др., 1987; Шмидт-Ниельсен, 1987; Вдовин, Четырбоцкий, 2018).
ЗАКЛЮЧЕНИЕ
Многоиглый керчак Myoxocephalus polyacanthocephalus в российских водах Японского моря встречается на глубинах 6–640 м при температуре –1.2…+15.5°C. В осенний период отходит на зимовку из северной части материковой акватории Татарского пролива, где широко распространён летом, в южном направлении, что обусловлено особенностями гидрологического режима. В связи с широтными миграциями плотности многоиглого керчака в прилегающей к м. Золотой акватории и ниже повышаются. В остальной части материковых вод, включая расположенный на юге зал. Петра Великого, а также у островного побережья о-ва Сахалин районы скоплений в течение года более стабильны.
Сезонные батиметрические различия проявляются в основном в предпочтениях диапазонов глубин в холодный и тёплый периоды. Зимой многоиглый керчак в массе избегает охлаждённую до отрицательных значений температуры верхнюю часть шельфа, предпочитая его нижнюю часть и верхний отдел материкового склона. Летом концентрируется в средней и верхней частях шельфа. В зал. Петра Великого в летний период скопления многоиглого керчака сдвинуты глубже, чем в остальных районах.
Молодь во все сезоны придерживается шельфовой зоны, являясь более эвритермной, чем взрослые особи.
Список литературы
Андронов П.Ю., Датский А.В. 2014. Закономерности пространственной изменчивости структуры донных сообществ рыб шельфа северо-западной части Берингова моря // Изв. ТИНРО. Т. 177. С. 40–76. https://doi.org/10.26428/1606-9919-2014-177-40-76
Борец Л.А. 1997. Донные ихтиоцены российского шельфа дальневосточных морей: состав, структура, элементы функционирования и промысловое значение. Владивосток: Изд-во ТИНРО-центр, 217 с.
Бретт Д.Р., Гроувс Д.Д. 1983. Физиологическая энергетика // Биоэнергетика и рост рыб. М.: Лег. и пищ. пром-сть. С. 203–274.
Вдовин А.Н., Зуенко Ю.И. 1997. Вертикальная зональность и экологические группировки рыб залива Петра Великого // Изв. ТИНРО. Т. 122. С. 152–176.
Вдовин А.Н., Четырбоцкий А.Н. 2018. Рост и стадийность онтогенеза южного однопёрого терпуга в водах Приморья (Японское море) // Тр. ВНИРО. Т. 170. С. 26–46. https://doi.org/10.36038/2307-3497-2018-170-26-46
Вдовин А.Н., Измятинский Д.В., Соломатов С.Ф. 2004. Основные результаты исследований ихтиофауны морского прибрежного комплекса Приморья // Изв. ТИНРО. Т. 138. С. 168–190.
Гудков П.К., Хованский И.Е. 2001. Бычки рода Myoxocephalus (Cottidae) из районов п-ова Кони (северная часть Охотского моря) // Состояние и перспективы рыбохозяйственных исследований в бассейне северной части Охотского моря. Сб. науч. тр. Вып. 1 / Под ред. Хованского Е.И. Магадан: Изд-во МагаданНИРО. С. 232–236.
Датский А.В. 2017. Особенности биологии массовых видов рыб в Олюторско-Наваринском районе и прилегающих водах Берингова моря. 4. Семейство рогатковые (Cottidae) // Вопр. ихтиологии. Т. 57. № 3. С. 251–263. https://doi.org/10.7868/S0042875217030031
Дударев В.А., Зуенко Ю.И., Ильинский Е.Н., Калчугин П.В. 1998. Новые данные о структуре сообществ донных и придонных рыб на шельфе и свале глубин Приморья // Изв. ТИНРО. Т. 123. С. 3–15.
Зуенко Ю.И. 1994. Типы термической стратификации вод на шельфе Приморья // Комплексные исследования морских гидробионтов и условий их обитания. Владивосток: Изд-во ТИНРО. С. 20–39.
Зуенко Ю.И. 2008. Промысловая океанография Японского моря. Владивосток: Изд-во ТИНРО-центр, 227 с.
Калчугин П.В. 1998. Распределение рогатковых (Cottidae) у северного Приморья // Изв. ТИНРО. Т. 123. С. 82–88.
Калчугин П.В., Бойко М.И., Соломатов С.Ф., Черниенко Э.П. 2016. Современное состояние ресурсов донных и придонных видов рыб в российских водах Японского моря // Там же. Т. 184. С. 54–69.
Ким Сен Ток. 2001. Зимние миграции шельфовых рыб в зону материкового склона юго-западного Сахалина // Вопр. ихтиологии. Т. 41. № 5. С. 593–604.
Линдберг Г.У., Красюкова З.В. 1987. Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Ч. 5. Л.: Наука, 526 с.
Лучин В.А. 2007. Сезонная изменчивость температуры воды в деятельном слое дальневосточных морей // Дальневосточные моря России. Кн. 1. Океанологические исследования / Под ред. Акуличева В.А. и др. М.: Наука. С. 232–252.
Матвеев А.А., Терентьев Д.А. 2016. Промысел, многолетняя динамика биомассы, распределение и размерный состав массовых видов рогатковых Cottidae у западного побережья Камчатки // Исследования водных биологических ресурсов Камчатки и северо-западной части Тихого океана. № 41. С. 17–42. https://doi.org/10.15853/2072-8212.2016.41.17-42
Никитин А.А., Дьяков Б.С. 2016. Океанологические исследования северо-западной части Японского моря по спутниковой информации и по данным океанографического разреза в зимние сезоны 2000–2015 гг. // Тр. ВНИРО. Т. 164. С. 86–99.
Новиков Н.П., Соколовский А.С., Соколовская Т.Г., Яковлев Ю.М. 2002. Рыбы Приморья. Владивосток: Изд-во Дальрыбвтуз, 552 с.
Орлов А.М. 2010. Количественное распределение демерсального нектона тихоокеанских вод северных Курильских островов и юго-восточной Камчатки. М.: Изд-во ВНИРО, 335 с.
Панченко В.В. 1999. Распределение керчаковых рыб рода Myoxocephalus в заливе Петра Великого Японского моря в летний период // Биология моря. Т. 25. № 2. С. 150–151.
Панченко В.В., Зуенко Ю.И. 2009. Распределение бычков семейства Cottidae в заливе Петра Великого Японского моря в летний период // Вопр. рыболовства. Т. 10. № 4 (40). С. 750–763.
Панченко В.В., Пущина О.И. 2004. Биологическая характеристика керчаковых рыб рода Myoxocephalus (Cottidae) зал. Петра Великого Японского моря // Изв. ТИНРО. Т. 138. С. 120–153.
Панченко В.В., Калчугин П.В., Соломатов С.Ф. 2016. Уточнение глубин обитания и максимальных размеров донных и придонных видов рыб в российских водах Японского моря // Вопр. ихтиологии. Т. 56. № 3. С. 264–283. https://doi.org/10.7868/S0042875216030152
Парин Н.В., Евсеенко С.А., Васильева Е.Д. 2014. Рыбы морей России: аннотированный каталог. М.: Т-во науч. изд. КМК, 733 с.
Пущина О.И., Соломатов С.Ф., Калчугин П.В., Будникова Л.Л. 2016. Питание и пищевые отношения массовых видов рогатковых (Cottidae, Pisces) зал. Петра Великого (Японское море) в летний период // Изв. ТИНРО. Т. 184. С. 186–203.
Радзинская Л.И., Никольская И.С., Чудакова И.В. 1987. Влияние аллатектомии на дыхание, вес и продолжительность жизни домового сверчка Achetadomestica L. // Онтогенез. Т. 18. № 3. С. 281–287.
Соколовский А.С., Соколовская Т.Г. 1997. К идентификации личинок керчаков (Myoxocephalus, Cottidae) залива Петра Великого (Японское море) // Вопр. ихтиологии. Т. 37. № 1. С. 54–61.
Соколовский А.С., Дударев В.А., Соколовская Т.Г., Соломатов С.Ф. 2007. Рыбы российских вод Японского моря: аннотированный и иллюстрированный каталог. Владивосток: Дальнаука, 200 с.
Соломатов С.Ф. 2008. Состав и многолетняя динамика донных ихтиоценов северного Приморья: Автореф. дис. … канд. биол. наук. Владивосток: ТИНРО-Центр, 24 с.
Токранов А.М. 1981. Распределение керчаковых (Cottidae, Pisces) на западнокамчатском шельфе в летний период // Зоол. журн. Т. 60. Вып. 2. С. 229–237.
Токранов А.М. 1984. О размножении многоиглого бычка Myoxocephalus polyacanthocephalus (Pallas) (Cottidae) в прикамчатских водах // Вопр. ихтиологии. Т. 24. Вып. 4. С. 601–608.
Токранов А.М. 1986. Керчаки и получешуйные бычки // Биологические ресурсы Тихого океана. М: Наука. С. 319–328.
Токранов А.М. 1988. Размножение массовых видов керчаковых рыб прикамчатских вод // Биология моря. № 4. С.28–32.
Токранов А.М., Орлов А.М. 2013. Особенности распределения, экология и динамика уловов многоиглого керчака Myoxocephalus polyacanthocephalus (Cottidae) в тихоокеанских водах северных Курильских островов и юго-восточной Камчатки // Биол. журн. Армении. Т. 4. № 65. С. 44–52.
Федоров В.В. 2000. Видовой состав, распределение и глубины обитания видов рыбообразных и рыб северных Курильских островов // Промыслово-биологические исследования рыб в тихоокеанских водах Курильских о-вов и прилежащих районах Охотского и Берингова морей в 1992−1998 гг. М.: Изд-во ВНИРО. С. 7−41.
Федоров В.В., Черешнев И.А., Назаркин М.В., Шестаков А.В., Волобуев В.В. 2003. Каталог морских и пресноводных рыб северной части Охотского моря. Владивосток: Дальнаука, 204 с.
Шмидт-Ниельсен К. 1987. Размеры животных: почему они так важны? М.: Мир, 259 с.
Яричин В.Г., Покудов В.В. 1982. Формирование структурных особенностей гидрофизических полей и течений в северной глубоководной части Японского моря // Тр. ДВНИГМИ. Вып. 96. С. 86–95.
Amaoka K., Nakaya K., Yabe M. 1995. The fishes of Northern Japan. Sapporo: Kita-Nihon Kaiyo Center Co. Ltd., 391 p.
Mecklenburg C.W., Mecklenburg T.A., Thorsteinson L.K. 2002. Fishes of Alaska. Bethesda: Amer. Fish. Soc., 1037 p.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии