

Вопросы ихтиологии, 2022, T. 62, № 3, стр. 251-271
Таксономия и филогенетические отношения карповых рыб рода Hemiculter (Cyprinidae, Xenocypridinae): востробрюшки видовой группы Hemiculter lucidus
Е. Д. Васильева 1, *, В. П. Васильев 2, И. Л. Мирошниченко 3, С. В. Шедько 3
1 Зоологический музей Московского государственного университета
Москва, Россия
2 Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
3 Федеральный научный центр биоразнообразия наземной биоты Восточной Азии
Дальневосточного отделения РАН – ФНЦ биоразнообразия ДВО РАН
Владивосток, Россия
* E-mail: vas_katerina@mail.ru
Поступила в редакцию 11.03.2021
После доработки 04.10.2021
Принята к публикации 04.10.2021
- EDN: ZALZJQ
- DOI: 10.31857/S0042875222030237
Аннотация
На основе сравнительного морфологического анализа многотычинковые востробрюшки разделены на четыре группы, различающиеся по числу ветвистых лучей в анальном плавнике, относительной длине головы, высоте тела, длине колючки спинного плавника, окраске брюшины: ханкайская востробрюшка Hemiculter lucidus, буирнурская востробрюшка H. varpachovskii; востробрюшки из бассейна р. Янцзы, пригодным названием для которых, как показано, является H. clupeoides, и востробрюшки из бассейна нижнего Амура, описанные как самостоятельный вид, статус которого подтверждается его отличиями от H. lucidus и H. clupeoides по составу гаплотипов митохондриального гена цитохрома b. Обсуждаются филогенетические отношения видов многотычинковых востробрюшек, рассматриваются некоторые проблемы синонимии, приведён ключ для диагностики видов.
Настоящая работа продолжает изучение карпообразных рыб (Cypriniformes) рода Hemiculter Bleeker, 1860, который, согласно современной классификации, входит в состав подсемейства Xenocypridinae Günther, 1868 семейства карповых (Cyprinidae) (Tan, Armbruster, 2018), рассматриваемого также в статусе самостоятельного семейства Xenocyprididae (Fricke et al., 2021; Froese, Pauly, 2021). Принятая до сих пор структура рода и номенклатура его представителей (Fricke et al., 2021; Froese, Pauly, 2021) находятся в противоречии с результатами современного филогеографического анализа нативных популяций (Chen et al., 2017; Wang et al., 2021; Васильева и др., 2022; настоящая работа). Данное исследование посвящено группе так называемых многотычинковых востробрюшек, объединённых ранее (Васильева, Козлова, 1988) в один вид H. lucidus (Dybowski, 1872), который в работах Берга (1909, 1949) и Никольского (1947, 1956) ошибочно (Васильева, Козлова, 1988) относился в ранге подвида к виду H. leucisculus (Basilewsky, 1855). В отличие от востробрюшек видовой группы H. leucisculus, чьи филогенетические отношения и таксономия рассмотрены нами ранее (Васильева и др., 2022), многотычинковые востробрюшки видовой группы H. lucidus характеризуются относительно большим числом густо сидящих жаберных тычинок на 1-й жаберной дуге (18–30, в среднем 22–24), наличием обычно не менее 12 ветвистых лучей в анальном плавнике; началом спинного плавника, расположенным, как правило, ближе к концу рыла, чем к началу хвостового плавника; и не резко изгибающейся вниз у грудного плавника боковой линией, в которой 40–53 (в среднем 42–46) прободённых чешуй (Васильева, Козлова, 1988; Васильева, 2004). В настоящее время в этой группе признаются валидными три вида: ханкайская востробрюшка H. lucidus в пресных водах России и Китая, H. bleekeri Warpachowski, 1887 в водоёмах Китая и буирнурская востробрюшка H. varpachovskii Nikolski, 1903 в бассейне верхнего течения Амура на территории Монголии, России и Китая (Fricke et al., 2021; Froese, Pauly, 2021); ареалы всех этих видов нуждаются в уточнении.
Цель настоящей работы – оценить морфологическую изменчивость и дивергенцию многотычинковых востробрюшек из разных бассейнов; выяснить филогенетические связи локальных групп популяций, таксономические отношения и номенклатуру обнаруженных филетических линий; выявить диагностические морфологические признаки всех таксонов, определить их нативные ареалы и разработать идентификационные ключи.
МАТЕРИАЛ И МЕТОДИКА
Морфологические исследования проводили на материалах из коллекции Зоологического музея МГУ (ЗММУ); в общей сложности было изучено 405 экз. Исходно все пробы ЗММУ, собранные в период до 1960 г., в соответствии с ошибочными представлениями Берга (1949) и Никольского (1947, 1956) о таксономии и номенклатуре востробрюшек были идентифицированы сборщиками или сотрудниками ЗММУ как H. leucisculus (включая рыб из озёр Буйр-Нуур и Далайнор) или его подвид H. leucisculus lucidus (оз. Ханка). Все эти пробы в соответствии с проведённой ранее ревизией рода (Васильева, Козлова, 1988) и результатами настоящего исследования переопределены и вместе с нашими современными сборами отнесены к четырём разным видам, таксономический статус которых дан ниже в интерпретации авторов настоящей работы.
H. lucidus, бассейн оз. Ханка – 78 экз., стандартная длина (SL) 55.0–172.5 мм и 4 экз. молоди SL < 50 мм: Р-7550 − 3 экз., Астраханка, 24.06.1949 г., коллектор В.Д. Лебедев; Р-7557 − 1 экз., 11.07.1948 г.; Р-7906 − 7 экз., сопка Лузанова, 19.07.1949 г.; Р-8343 − 14 экз., 1949 г.; Р-20064 − 1 экз., каналы рисовых чеков у дер. Владимиро-Петровка, 20.09.1996 г., коллектор В.П. Васильев; Р-20066 − 1 экз., Камень-Рыболов, база ТИНРО, 11.09.1996 г., коллектор В.П. Васильев; Р-20105 − 43 экз., Камень-Рыболов, база ТИНРО, 15.09.1996 г., коллектор В.П. Васильев; Р-21966 − 12 экз., Камень-Рыболов, база ТИНРО, 25−26.09.2007 г., коллекторы В.П. Васильев, Е.Д. Васильева, С.В. Шедько (включают ваучеры на ДНК- анализ).
H. varpachovskii, оз. Буйр-Нуур, Монголия – 82 экз. SL 85.5–129.0 мм: Р-8041 − 3 экз., лето 1948 г.; Р-8069 – 10 экз., июль–август 1948 г.; Р-8375 – 16 экз., август 1948 г.; Р-8606 − 16 экз., в плохом состоянии, 1948 г., коллектор А. Дашдорж (на этикетках здесь и далее указано: “coll. Анударин”); Р-8608 − 7 экз., лето 1948 г., коллектор А. Дашдорж; Р-8683 − 3 экз., коллектор А. Дашдорж; Р-13018 − 27 экз., коллектор А. Дашдорж.
Бассейн оз. Далайнор, Монголия – 106 экз. SL 77.0–119.0 мм. Сборы А.А. Световидовой: Р-8674 − 8 экз., Далайнор, 17.06.1957 г.; Р-8675 − 9 экз., р. Керулен, 17.06.1957 г.; Р-8699 − 1 экз., Далайнор, мутная протока, 13.06.1957 г.; Р-8700 − 21 экз., р. Уршун (= Оршун-гол), 07.06.1957 г.; Р-8701 − 20 экз., р. Уршун, 28.06.1957 г.; Р-8702 − 3 экз., р. Уршун, 09.06.1957 г.; Р-8728 − 4 экз., Далайнор, лето 1957 г.; Р-8729 − 29 экз., Далайнор, лето 1957 г.; Р-8730 − 10 экз., Далайнор, 16.06.1957 г.; Р-22031 − 1 экз., р. Керулен, 47°01′ с.ш. 109°08′ в.д., 28.08.2007 г., коллектор Ю.В. Слынько.
H. clupeoides Nichols, 1925, Китай – 44 экз. SL 71.0–118.0 мм: Р-8638 – 1 экз., получен от Института гидробиологии Китайской АН; Р-9436 – 35 экз., р. Янцзы (“collected from G-Chang”), 28.04.1958 г.; Р-22294 – 8 экз., р. Янцзы, Цзинчжоу (Jingzhou), провинция Хубэй (Hubei), 31.10.2009 г., коллекторы Е.Д. Васильева, В.П. Васильев.
Hemiculter sp., описан в статусе нового вида в настоящей работе, бассейн нижнего Амура – 89 экз. Бассейн оз. Болонь – 46 экз. SL 82.0–146.0 мм: Р-6166 – 1 экз., м. Нергуль, 01.08.1946 г., коллектор Г.В. Никольский; Р-6172 – 3 экз., м. Серебряный, 24.07.1946 г., коллектор Г.В. Никольский; Р-6174 – 2 экз., о. Туф, 06.07.1946 г., коллектор М.Н. Лишев; Р-6234 – 7 экз., июнь 1946 г., коллектор С.Г. Соин; Р-6506 – 10 экз., 20.06.1947 г., коллектор Т.К. Сысоева; Р-6545 – 2 экз., 28.05.1947 г.; Р-6600 – 3 экз., м. Серебряный, 10.07.1947 г., коллектор Т.К. Сысоева; Р-6859 – 1 экз., 28.06.1947 г., коллектор Т.К. Сысоева; Р-6860 – 4 экз., о. Туф, 05.07.1947 г., коллектор Т.К. Сысоева; Р-6867 – 6 экз., 28.06.1947 г., коллектор Т.К. Сысоева; Р-6880 – 3 экз., 01.08.1946 г., коллектор Т.К. Сысоева; Р-8432 – 3 экз., м. Серебряный, 03.08.1946 г.; Р-17128 – 1 экз., выделен из пробы Р-6391 с 4 экз. H. leucisculus, протока Сий, 30.07.1947 г., коллектор Г.В. Никольский.
Р. Амур в районе Елабуги – 12 экз. SL 99.0–142.0 мм: Р-6405 – 1 экз., 19.06.1947 г., коллектор Г.В. Никольский; Р-7502 – 2 экз., 24.07.1949 г., коллектор Т.К. Сысоева; Р-7536 – 2 экз., 12.07.1948 г., коллектор А.А. Световидова; Р-7884 – 1 экз., 30.07.1949 г.; Р-7885 – 3 экз., 12.06.1949 г.; Р-7956 – 1 экз., 01.08.1949 г.; Р-8574 – 2 экз., оз. Медовое, 20.06.1948 г., коллектор С.Г. Соин.
Нижний Амур на разных участках – 29 экз. SL 61.0–115.0 мм: Р-6082 – 7 экз., оз. Большое Кизи, 26.07.1946 г., коллектор М.Н. Лишев; Р-6098 – 3 экз., Амур выше дер. Монгольское, 23.08.1946 г., коллектор Г.В. Никольский; Р-6144 – 1 экз., оз. Удыль, 29.07.1946 г., коллектор М.Н. Лишев; Р-6479 – 9 экз., Амур у Сикача-Алян, 12.06.1947 г., коллектор Г.В. Никольский; Р-7129 – 2 экз., Амур у Джелинды, устье Б. Неверо, 25.07.1948 г., коллектор В.Д. Спановская; Р-7134 – 6 экз., Амур, 7 км выше Ольгино, 07.08.1948 г., коллектор В.Д. Спановская; Р-21562 – 1 экз., Амур у Безымянного (выше Хабаровска), 07.08.2004 г., коллектор Е.Д. Васильева.
Р. Сунгари (бассейн Амура) у Харбина (Китай): Р-3513 – 2 экз. SL 75.5 и 82.0 мм, 07.07.1932 г., коллектор А.С. Лукашкин.
У всех особей измеряли стандартную длину тела (SL) и анализировали признаки, использовавшиеся при дифференциации и описании разных номинальных видов рода Hemiculter и близких таксонов (Bănărescu, 1968; Васильева, Козлова, 1988; Luo, Chen, 1998; Tan, Armbruster, 2018), прежде всего, характеристики, обнаруживающие наибольшую межпопуляционную изменчивость в пределах группы H. lucidus и использовавшиеся китайскими авторами для выделения подвидов в бассейне Амура (Цзин-цзян, Бе-лу, 1959; Васильева, Козлова, 1989а): относительная длина головы (с), наибольшая высота тела перед спинным плавником (Н), высота спинного плавника (hD), длина последнего неветвистого луча (колючки) спинного плавника (Hsp), число ветвистых лучей в анальном плавнике (А) (два последних сближенных луча считали за один луч), число жаберных тычинок на 1-й жаберной дуге (sp.br.), чешуй в боковой линии (ll). В таблицу не включены данные по молоди востробрюшек из оз. Ханка. При анализе дифференциации рыб из разных бассейнов и составлении характеристик таксонов использовали также полученные ранее результаты исследования изменчивости широкого набора морфометрических характеристик у востробрюшек видовой группы H. lucidus на внутри- и межпопуляционном уровне (Васильева, Козлова, 1989а). У отдельных особей из музейных коллекций изучены также окраска брюшины, строение плавательного пузыря, формула глоточных зубов и относительная длина кожного подключичного выроста (flepl − flesh slip at the pectoral fin base) (Dai, Yang, 2003. Р. 78).
Статистический анализ выполнен с применением программного пакета MS Excel; величину различий между сравниваемыми выборками по признакам с выявленными достоверными различиями (tst) оценивали на основе значений коэффициента различий CD (Майр и др., 1956). При оценке нативных ареалов видов использовали материалы музейной коллекции, результаты собственных полевых работ авторов настоящей статьи и молекулярно-генетических исследований, материалов из Генбанка, а также информацию из публикаций, цитируемых в работе.
Полноразмерные последовательности митохондриального гена цитохрома b (cyt b) были секвенированы у 55 образцов многотычинковых востробрюшек (депонированы в Генбанк под номерами доступа MW508412–MW508458). В дополнение к обработанному ранее (Васильева и др., 2022) материалу от востробрюшек из р. Янцзы, провинция Хубэй, (проба ЗММУ Р-22294, n = 8) были использованы особи H. lucidus из оз. Ханка (n = 20, включая экземпляры из пробы ЗММУ Р-21966), а также описанного в настоящей работе самостоятельного вида рода Hemiculter, собранные в 2006–2008 гг. из р. Сунгари у г. Харбин (n = 8) и р. Амур у г. Хабаровск (n = 19). Кроме собственных материалов, в работе использованы данные по последовательности гена cyt b KF760461, полученной от востробрюшки из Южной Кореи, определённой (Kim et al., 2014) как H. leucisculus, но причисленной в Генбанке к H. eigenmanni (Jordan et Metz, 1913).
Выделение ДНК, ПЦР-амплификация, секвенирование ДНК и филогенетический анализ проведены так же, как описано ранее (Васильева и др., 2022). Отличие состояло лишь в том, что из-за сравнительно низкого уровня дивергенции гена cyt b внутри многотычинковых востробрюшек (максимальные дистанции порядка 2.5% различающихся позиций) нуклеотидную последовательность этого гена не подразделяли на блоки и матрицу данных анализировали при использовании одной оптимальной модели нуклеотидных замещений (TN + F + G4), подобранной в программе IQ-TREE v.1.6.12 (Nguyen et al., 2015).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Морфологическая изменчивость и дивергенция многотычинковых востробрюшек группы Hemiculter lucidus
Сравнительный морфологический анализ изменчивости многотычинковых востробрюшек из локальных популяций показал, что особи из разных бассейнов достаточно чётко различаются по совокупности следующих характеристик: числу ветвистых лучей в анальном плавнике, относительной длине головы, высоте тела и длине последнего неветвистого луча (колючки) спинного плавника, окраске брюшины.
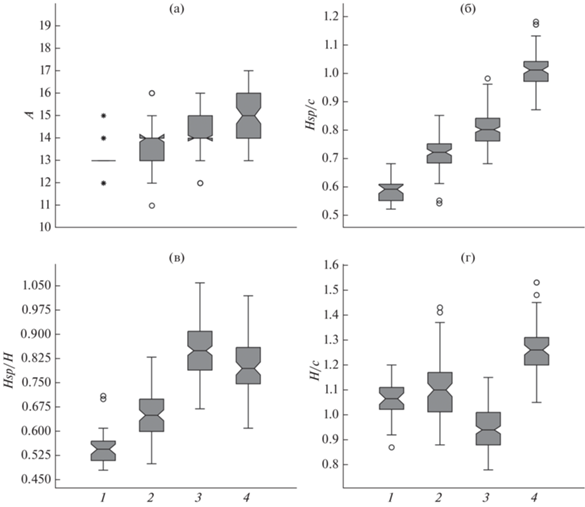
Востробрюшки бассейна оз. Ханка обнаруживают наиболее высокий уровень морфологической дивергенции от популяций других водных бассейнов. У них, как правило, в анальном плавнике не менее 14 ветвистых лучей (рис. 1): среди изученных 78 экз. только у одного было 13 лучей в плавнике, преобладали особи с 15–16 лучами (56.4%). По данным китайских авторов (Цзин-цзян, Бе-лу, 1959), у востробрюшек из Ханки редко бывает 13 ветвистых лучей (2/28), обычно их 14–15 (17/28), встречаются и особи с 19 лучами (1/28). Большое число лучей в анальном плавнике – от 14 до 18 − указывают и другие авторы (Warpachowski, 1887; Варпаховский, Герценштейн, 1887; Линдберг, Таранец, 1929; Берг, 1949; Никольский, 1956; Bănărescu, 1968; Luo, Chen, 1998). Голова у рыб из Ханки относительно небольшая (таблица), укладывается 4.2–5.4 раза в SL (обычно 5 и более раз), как указывается также в ключах Луо и Чена (Luo, Chen, 1998). По данным Цзин-цзян и Бе-лу (1959), длина головы у этих востробрюшек варьирует от 17 до 20% SL, по данным других авторов – содержится от 5.3 до 5.5 раза в SL (Warpachowski, 1887; Варпаховский, Герценштейн, 1887; Берг, 1949).
Рис. 1.
Диаграммы размаха основных морфологических признаков, использованных для диагностики многотычинковых востробрюшек рода Hemiculter из разных бассейнов: а – число ветвистых лучей в анальном плавнике (А), б – отношение длины колючки спинного плавника к длине головы (Hsp/с), в – отношение длины колючки спинного плавника к наибольшей высоте тела у начала спинного плавника (Hsp/H), г – отношение наибольшей высоты тела к длине головы (H/с). Выборки: 1 – р. Янцзы, 2 – бассейн нижнего Амура, 3 – бассейн верхнего Амура, 4 – система оз. Ханка; (I) – пределы варьирования показателя (от 25 до 75% данных заключены в залитую серым фигуру), (─) – медиана; (⚪), (*) – выбросы.
Некоторые морфометрические характеристики многотычинковых востробрюшек (Hemiculter) из разных водных бассейнов
| Признак | Янцзы | Нижний Амур | Верхний Амур | Ханка |
|---|---|---|---|---|
| n | 37 | 88 | 170 | 78 |
| SL, мм | $\frac{{71{\kern 1pt} --{\kern 1pt} 118}}{{88.1}}$ | $\frac{{56{\kern 1pt} --{\kern 1pt} 146}}{{99.4}}$ | $\frac{{77{\kern 1pt} --{\kern 1pt} 129}}{{98.6}}$ | $\frac{{55{\kern 1pt} --{\kern 1pt} 172.5}}{{119.0}}$ |
| A | $\frac{{12{\kern 1pt} --{\kern 1pt} 15}}{{13.1 \pm {\text{ }}0.13}}$ | $\frac{{11{\kern 1pt} --{\kern 1pt} 16}}{{13.7 \pm 0.12}}$ | $\frac{{12{\kern 1pt} --{\kern 1pt} 16}}{{14.1 \pm 0.07}}$ | $\frac{{13{\kern 1pt} --{\kern 1pt} 17}}{{15.4 \pm 0.12}}$ |
| Hsp/H | $\frac{{0.48{\kern 1pt} --{\kern 1pt} 0.71}}{{0.55 \pm 0.009}}$ | $\frac{{0.50{\kern 1pt} --{\kern 1pt} 0.83}}{{0.66 \pm 0.008}}$ | $\frac{{0.67{\kern 1pt} --{\kern 1pt} 1.06}}{{0.85 \pm 0.006}}$ | $\frac{{0.61{\kern 1pt} --{\kern 1pt} 1.02}}{{0.80 \pm 0.009}}$ |
| Hsp/c | $\frac{{0.52{\kern 1pt} --{\kern 1pt} 0.68}}{{0.59 \pm 0.007}}$ | $\frac{{0.54{\kern 1pt} --{\kern 1pt} 0.85}}{{0.71 \pm 0.006}}$ | $\frac{{0.68{\kern 1pt} --{\kern 1pt} 0.96}}{{0.80 \pm 0.004}}$ | $\frac{{0.71{\kern 1pt} --{\kern 1pt} 1.18}}{{1.00 \pm 0.008}}$ |
| H/c | $\frac{{0.87{\kern 1pt} --{\kern 1pt} 1.20}}{{1.07 \pm 0.012}}$ | $\frac{{0.88{\kern 1pt} --{\kern 1pt} 1.43}}{{1.10 \pm 0.013}}$ | $\frac{{0.78{\kern 1pt} --{\kern 1pt} 1.15}}{{1.00 \pm 0.053}}$ | $\frac{{1.05{\kern 1pt} --{\kern 1pt} 1.53}}{{1.26 \pm 0.010}}$ |
| *По данным: Васильева, Козлова, 1988, 1989а | ||||
| n | 35 | 88 | 98 | 19 |
| ll | $\frac{{40{\kern 1pt} --{\kern 1pt} 47}}{{43.0 \pm 0.41}}$ | $\frac{{40{\kern 1pt} --{\kern 1pt} 45}}{{40.7{\kern 1pt} --{\kern 1pt} 43.9}}$ | $\frac{{40{\kern 1pt} --{\kern 1pt} 49}}{{42.1{\kern 1pt} --{\kern 1pt} 44.3}}$ | $\frac{{40{\kern 1pt} --{\kern 1pt} 49}}{{44.4 \pm 0.48}}$ |
| В % SL | ||||
| aD | $\frac{{48.4{\kern 1pt} --{\kern 1pt} 52.2}}{{50.2 \pm 0.16}}$ | $\frac{{46.0{\kern 1pt} --{\kern 1pt} 54.2}}{{49.6{\kern 1pt} --{\kern 1pt} 51.9}}$ | $\frac{{46.6{\kern 1pt} --{\kern 1pt} 54.4}}{{49.3{\kern 1pt} --{\kern 1pt} 50.8}}$ | $\frac{{47.8{\kern 1pt} --{\kern 1pt} 53.6}}{{50.3 \pm 0.29}}$ |
| aV | $\frac{{46.0{\kern 1pt} --{\kern 1pt} 50.7}}{{48.3 \pm 0.20}}$ | $\frac{{42.5{\kern 1pt} --{\kern 1pt} 55.7}}{{48.0{\kern 1pt} --{\kern 1pt} 50.3}}$ | $\frac{{44.9{\kern 1pt} --{\kern 1pt} 52.8}}{{47.7{\kern 1pt} --{\kern 1pt} 48.5}}$ | $\frac{{44.4{\kern 1pt} --{\kern 1pt} 47.3}}{{45.9 \pm 0.21}}$ |
| lP | $\frac{{19.0{\kern 1pt} --{\kern 1pt} 22.7}}{{20.4 \pm 0.12}}$ | $\frac{{17.8{\kern 1pt} --{\kern 1pt} 27.2}}{{21.0{\kern 1pt} --{\kern 1pt} 25.4}}$ | $\frac{{21.4{\kern 1pt} --{\kern 1pt} 26.4}}{{23.5{\kern 1pt} --{\kern 1pt} 24.2}}$ | $\frac{{22.2{\kern 1pt} --{\kern 1pt} 24.6}}{{23.4 \pm 0.15}}$ |
| lV | $\frac{{13.4{\kern 1pt} --{\kern 1pt} 16.0}}{{14.7 \pm 0.10}}$ | $\frac{{12.9{\kern 1pt} --{\kern 1pt} 19.0}}{{15.2{\kern 1pt} --{\kern 1pt} 16.9}}$ | $\frac{{14.4{\kern 1pt} --{\kern 1pt} 19.38}}{{16.4{\kern 1pt} --{\kern 1pt} 17.1}}$ | $\frac{{14.7{\kern 1pt} --{\kern 1pt} 18.6}}{{16.6 \pm 0.27}}$ |
| lA | $\frac{{12.5{\kern 1pt} --{\kern 1pt} 16.3}}{{14.6 \pm 0.15}}$ | $\frac{{11.6{\kern 1pt} --{\kern 1pt} 20.5}}{{14.2{\kern 1pt} --{\kern 1pt} 15.6}}$ | $\frac{{12.0{\kern 1pt} --{\kern 1pt} 17.8}}{{14.8{\kern 1pt} --{\kern 1pt} 15.4}}$ | $\frac{{15.4{\kern 1pt} --{\kern 1pt} 18.7}}{{17.2 \pm 0.20}}$ |
| hD | $\frac{{15.5{\kern 1pt} --{\kern 1pt} 18.3}}{{16.8 \pm 0.12}}$ | $\frac{{15.5{\kern 1pt} --{\kern 1pt} 23.6}}{{17.7{\kern 1pt} --{\kern 1pt} 20.8}}$ | $\frac{{17.4{\kern 1pt} --{\kern 1pt} 23.9}}{{20.3{\kern 1pt} --{\kern 1pt} 21.5}}$ | $\frac{{19.7{\kern 1pt} --{\kern 1pt} 27.1}}{{22.5 \pm 0.47}}$ |
| Hsp | $\frac{{12.2{\kern 1pt} --{\kern 1pt} 16.1}}{{14.4 \pm 0.15}}$ | $\frac{{13.7{\kern 1pt} --{\kern 1pt} 20.8}}{{15.2{\kern 1pt} --{\kern 1pt} 17.6}}$ | $\frac{{16.3{\kern 1pt} --{\kern 1pt} 22.7}}{{19.2{\kern 1pt} --{\kern 1pt} 20.4}}$ | $\frac{{18.5{\kern 1pt} --{\kern 1pt} 23.6}}{{20.5 \pm 0.38}}$ |
| H | $\frac{{20.7{\kern 1pt} --{\kern 1pt} 29.4}}{{25.4 \pm 0.27}}$ | $\frac{{19.0{\kern 1pt} --{\kern 1pt} 30.9}}{{22.5{\kern 1pt} --{\kern 1pt} 26.6}}$ | $\frac{{19.8{\kern 1pt} --{\kern 1pt} 28.0}}{{23.1{\kern 1pt} --{\kern 1pt} 23.3}}$ | $\frac{{25.3{\kern 1pt} --{\kern 1pt} 30.4}}{{27.2 \pm 0.37}}$ |
| c | $\frac{{21.5{\kern 1pt} --{\kern 1pt} 24.7}}{{23.3 \pm 0.13}}$ | $\frac{{20.4{\kern 1pt} - {\kern 1pt} 27.4}}{{22.0{\kern 1pt} --{\kern 1pt} 24.5}}$ | $\frac{{21.3{\kern 1pt} --{\kern 1pt} 27.1}}{{23.3{\kern 1pt} --{\kern 1pt} 24.6}}$ | $\frac{{18.6{\kern 1pt} --{\kern 1pt} 23.8}}{{21.0 \pm 0.34}}$ |
Примечание. n – число особей; над чертой – диапазон изменчивости, под чертой − среднее значение и его ошибка (*по бассейнам нижнего и верхнего Амура – изменчивость средних значений в разных выборках); SL – стандартная длина тела, А – число ветвистых лучей в анальном плавнике; aD, aV – антедорсальное и антевентральное расстояния; lP, lV – длина грудного и брюшного плавника соответственно, lA – длина основания анального плавника, hD – высота спинного плавника, Hsp – длина колючки спинного плавника, H – наибольшая высота тела у начала спинного плавника, c – длина головы, ll – число чешуй боковой линии.
Наибольшая высота тела у изученных нами востробрюшек из Ханки всегда больше длины головы (таблица, рис. 1г), что отмечается для них и в ключах китайских авторов (Luo, Chen, 1998); содержится 3.3–4.0 раза в SL, 3.8–4.0 − по данным других авторов (Warpachowski, 1887; Варпаховский, Герценштейн, 1887; Берг, 1949). Длина колючки спинного плавника обычно составляет 75–85% Н, как правило, превышает 85% с, а обычно равна или больше длины головы (рис. 1б); по данным Берга (1949), длина колючки больше или равна длине головы, по ключам китайских авторов (Цзин-цзян, Бе-лу, 1959; Luo, Chen, 1998) − обычно больше длины головы. Брюшина у всех изученных особей из пробы Р-21966 (12 экз.) чёрная, у 3 экз. из пробы Р-8343 – тёмно-коричневая.
Востробрюшки из бассейна р. Янцзы в наибольшей степени отличаются по морфологическим характеристикам от особей из бассейна Ханки (здесь и далее рассматриваются только признаки, по которым выявлены статистически достоверные различия). У изученных нами рыб в анальном плавнике 12–15 ветвистых лучей, обычно не более 13 (27/37); 12–13 лучей для особей из бассейна Янцзы приводят и другие авторы (Цзин-цзян, Бе-лу, 1959). Соответственно и длина основания анального плавника у востробрюшек из Янцзы существенно меньше, чем у ханкайских (таблица): уровень различий превышает формально подвидовой уровень – CD = 1.48. Ещё более низкие средние значения lA приводят для востробрюшек из Янцзы китайские авторы (Li et al., 2020) – 13.9 ± 0.9% SL. Длина головы у изученных особей из Янцзы больше, чем у востробрюшек из Ханки, укладывается 4.1–4.7 раза в SL. Выявленные различия невелики (CD = 1.02) и в определённой степени могут быть обусловлены меньшими размерами востробрюшек из Янцзы., хотя сходные средние размеры головы – 21.2 ± 1.0% SL – приводят и китайские авторы (Li et al., 2020).
Наибольшая высота тела у востробрюшек из Янцзы обычно чуть превышает длину головы (таблица, рис. 1г), что отмечается для них и в ключах китайских авторов (Luo, Chen, 1998); содержится 3.4–4.8 раза в SL; однако китайские авторы (Li et al., 2020) приводят меньшую среднюю величину признака, чем получили мы – 20.2 ± 1.4% SL. Длина колючки спинного плавника обычно < 60% с и < 55% Н. По индексу Hsp/c наблюдается хиатус между распределениями значений у востробрюшек Янцзы и Ханки, CD = 3.70. По индексу Hsp/H (рис. 1в) величина различий превышает формально подвидовой уровень: CD = 1.57. В отличие от изученных востробрюшек из Ханки у 3 экз. из Янцзы из пробы Р-22294 и у особи из Китая из пробы Р-8638 брюшина светлая.
Помимо отмеченных выше характеристик востробрюшки из р. Янцзы достоверно отличаются от изученной выборки из Ханки (Васильева, Козлова, 1989а) меньшей высотой спинного плавника (наблюдается хиатус между распределениями значений признака, CD = 2.07), меньшей длиной грудного плавника (CD = 2.20), а также меньшей длиной брюшного плавника (таблица). Китайские авторы (Li et al., 2020) приводят для этих характеристик следующие средние значения − соответственно 18.1 ± 1.2, 19.3 ± 0.8 и 14.7 ± 0.9% SL.
Востробрюшки из разных популяций бассейна нижнего Амура демонстрируют наибольшее морфологическое сходство с таковыми бассейна Янцзы со сдвигом средних значений в направлении диапазона изменчивости востробрюшек из Ханки (таблица). В анальном плавнике у них от 11 до 16 ветвистых лучей (рис. 1а), чаще 13 (28/88) или 14 (30/88). Сходные данные приводятся для популяций Амура с территории Китая (Цзин-цзян, Бе-лу, 1959): 12–16 лучей, чаще 13 (38/126) или 14 (56/126); по данным Никольского (1947), у востробрюшек из разных водоёмов бассейна нижнего Амура среднее число лучей 13.4–14.1. Длина основания анального плавника варьирует в более широком диапазоне, чем у рыб из Янцзы и Ханки, но средние значения в разных амурских выборках ближе к средней величине признака в выборке из Янцзы (таблица), хотя их отличия от средних значений у востробрюшек Ханки невелики (CD не достигает формально подвидового уровня 1.28). Длина головы нижнеамурских востробрюшек укладывается 3.7−4.9 раза в SL.
Наибольшая высота тела обычно больше длины головы (таблица, рис. 1г); в разных локальных популяциях значение соотношения H/c в среднем варьирует от 0.92 до 1.16; содержится 3.2−5.3 раза в SL. По средней величине индекса H/c амурские востробрюшки сходны с особями из Янцзы и достоверно отличаются от ханкайских, но различия эти невелики (таблица). Длина колючки спинного плавника обычно не превышает 75% с и составляет от 60 до 70% Н (таблица, рис. 1в). По средним значениям индексов Hsp/H и Hsp/c востробрюшки нижнего Амура достоверно отличаются и от рыб из Янцзы, и от рыб из Ханки, однако значительные различия наблюдаются только по индексу Hsp/c между амурскими и ханкайскими востробрюшками: CD = 2.32. В то же время средние значения длины колючки в разных амурских популяциях варьируют в пределах 15.2−17.6% SL, и в случае выборки из оз. Кабар с Hsp 17.3 ± 0.34% SL отличия от востробрюшек из Янцзы достигают величины CD = 1.29. В свою очередь распределение значений Hsp в выборке амурских востробрюшек из оз. Большое Кизи – 13.8–16.5 (в среднем 15.2)% SL (n = 7) – не перекрывается с диапазоном изменчивости ханкайских востробрюшек.
По высоте спинного плавника изученные популяции нижнего Амура занимают промежуточное положение между востробрюшками Янцзы и Ханки. Все амурские выборки достоверно отличаются от выборки из Янцзы, и в ряде случаев эти отличия достигают значительной величины: так, при сравнении с популяцией оз. Кабар с hD 20.4 ± ± 0.29% SL CD = 1.93, а с выборкой из Амура у пос. Елабуга с hD 20.3 ± 0.30% SL CD = 1.38. Аналогично достоверны различия всех амурских выборок с ханкайскими востробрюшками, и в ряде случаев они значительны: с выборкой из Амура с hD 17.7 ± 0.20% SL CD = 1.60, с выборкой из оз. Болонь с hD 18.6 ± 0.16 CD = 1.37, а распределение значений признака в выборке из оз. Большое Кизи hD 17.2–19.4% SL не перекрывается с диапазоном изменчивости ханкайских востробрюшек (таблица).
Диапазон изменчивости относительной длины грудного и брюшного плавников у нижнеамурских востробрюшек превышает суммарную изменчивость востробрюшек Янцзы и Ханки; наименьшие средние выборочные значения амурских выборок близки к средним значениям признаков в выборке из Янцзы, а наибольшие – к средним востробрюшек Ханки (таблица).
Популяции системы р. Аргунь (верхнее течение Амура) – озёра Буйр-Нуур (=Хулун-Буйр) и Далайнор, реки Уршун и Керулен – по большинству признаков более сходны с востробрюшками из Ханки, чем с особями из бассейнов нижнего Амура и р. Янцзы. В анальном плавнике у них, как правило, не менее 13 ветвистых лучей (12 лучей обнаружено только у 6 из 170 экз.), чаще 14 (70/170) или 15 (51/170) лучей. Сходные данные приводят для буирнурской востробрюшки китайские авторы (Цзин-цзян, Бе-лу, 1959): 12–15 лучей, чаще 13 (12/40), 14 (10/40) или 15 (15/40); в описании H. varpachovskii, составленном по 8 экз., приводится 15 лучей (Никольский, 1903). Средние значения длины основания анального плавника в разных выборках сходны со средними значениями этого признака у востробрюшек из бассейна нижнего Амура (таблица). Длина головы изученных востробрюшек из бассейна верхнего Амура укладывается 3.7–4.7 раза в SL, что согласуется с характеристикой буирнурских востробрюшек в диагностическом ключе Луо и Чена (Luo, Chen, 1998) (<5.0 раза в SL) и с описанием Никольского (1903) – 4½−4⅔. Средние значения длины головы в разных популяциях сходны с таковыми востробрюшек из Янцзы и нижнего Амура (таблица). По данным Цзин-цзян и Бе-лу (1959), длина головы у этих востробрюшек варьирует от 21 до 25% SL, в среднем 22.7%.
Наибольшая высота тела обычно не превышает длину головы (таблица, рис. 1г), содержится 3.6–5.1 раза в SL (4–4½ – по описанию Никольского (1903)); по ключам китайских авторов (Цзин-цзян, Бе-лу, 1959), высота тела буирнурских востробрюшек не превышает 22% SL. По средней величине индекса H/c верхнеамурские востробрюшки сходны с особями из Янцзы и из бассейна нижнего Амура и достоверно отличаются от ханкайских, но различия эти невелики. Длина колючки спинного плавника обычно больше 75%, но меньше 90% с (короче головы: sensu Luo, Chen, 1998) и, как правило, превышает 80% Н (рис. 1б, 1в). По индексу Hsp/c отличия востробрюшек бассейна верхнего Амура достигают значительной величины как от востробрюшек Янцзы (CD = 2.18, распределения значений признака в выборках практически не перекрываются), так и от ханкайских востробрюшек (CD = 1.56). По индексу Hsp/H значительные различия наблюдаются между востробрюшками верхнего Амура и Янцзы: CD = 2.21. Средние значения Hsp (в % SL) у верхнеамурских востробрюшек сходны с ханкайскими; с распределением значений признака у особей из Янцзы наблюдается хиатус, и различия со всеми верхнеамурскими выборками достигают значительной величины: CD = 2.26 с выборкой из оз. Буйр-Нуур с Hsp 19.2 ± 0.18% SL, CD = 3.15 с выборкой из р. Уршун с Hsp 19.2 ± ± 0.13% SL, CD = 2.68 с выборкой из оз. Далайнор с Hsp 20.4 ± 0.26% SL. Значительной величины достигают различия по индексу Hsp между верхнеамурскими и нижнеамурскими выборками с низкими средними значениями индекса: при сравнении выборки из нижнего Амура с Hsp 15.8 ± 0.25% SL с выборкой из оз. Буйр-Нуур CD = 1.40, с выборкой из р. Уршун − 1.85, с выборкой из оз. Далайнор − 1.80. В то же время отличия от нижнеамурских выборок с большими средними значениями Hsp (например, с выборкой из оз. Кабар с Hsp 17.3 ± 0.34% SL) достоверны, но невелики − CD не достигает уровня 1.28. У двух вскрытых экземпляров из пробы Р-8069 брюшина тёмная.
По средним значениям высоты спинного, длины грудного и брюшного плавников востробрюшки из водоёмов бассейна верхнего Амура наиболее сходны с ханкайскими востробрюшками (таблица).
Таксономическая структура и проблемы номенклатуры многотычинковых востробрюшек
Дифференцируемые по морфологическим признакам четыре группы многотычинковых востробрюшек населяют разные водные бассейны, различия между ними по отдельным характеристикам превышают формально подвидовой уровень CD = 1.28, а по другим – наблюдается хиатус между распределениями значений признака, что соответствует видовому морфологическому критерию. В любом случае по совокупности морфологических характеристик, использовавшихся ранее в определительных ключах, востробрюшки из разных групп чётко дифференцируются друг от друга (см. ниже разработанный в настоящем исследовании диагностический ключ) и принимаются нами в статусе самостоятельных таксонов видового ранга. Востробрюшки из бассейна Ханки и оз. Буйр-Нуур ранее были описаны в качестве самостоятельных видов, которые признаются и в настоящее время: соответственно H. lucidus и H. varpachovskii (Fricke et al., 2021; Froese, Pauly, 2021). В то же время номенклатура двух других видов – востробрюшек бассейна р. Янцзы и востробрюшек нижнего течения Амура − нуждается в специальном анализе.
Востробрюшки р. Янцзы. В современных сводках по ихтиофауне для р. Янцзы указываются два вида востробрюшек с sp.br. > 20: H. tchangi Fang, 1942 и уже упоминавшийся H. bleekeri (Luo, Chen, 1998; Fu et al., 2003; He, 2010). В предыдущей нашей работе (Васильева и др., 2022) подтверждён вывод Бэнэреску (Bănărescu, 1968) о том, что H. tchangi был описан по гибридной особи, и поэтому название tchangi не может употребляться как валидное для родительских видов этого гибрида (Международный кодекс зоологической номенклатуры, 2000. Ст. 23.8). Валидным названием для вида рода Hemiculter, участвовавшего в гибридизации, является предложенное позже (Yih, Wu, 1964) – nigromarginis. Эндемичный вид притоков верхнего течения Янцзы H. nigromarginis принадлежит филогенетической кладе малотычинковых востробрюшек (Chen et al., 2017; Васильева и др., 2022) и, как и эти виды, характеризуется малым числом ветвистых лучей в анальном плавнике (11–12) и началом спинного плавника, расположенным, как правило, ближе к началу хвостового плавника, чем к концу рыла. От других видов собственно малотычинковых востробрюшек он отличается бóльшим числом жаберных тычинок (25–28), а также относительно бóльшим числом чешуй в боковой линии (49–53) (Bănărescu, 1968).
Вид H. bleekeri был описан Варпаховским (Warpachowski, 1887) на основе описания Бликером (Bleeker, 1863−1864, 1871) двух особей из Китая (предположительно из р. Янцзы), отнесённых им к виду H. leucisculus. Описание Варпаховского повторяет следующие важные характеристики из публикаций Бликера: D II 7–8, А III 11–13, P I 13–14, V II 8, ll 40−42; Н укладывается 4⅔–4¾ раза в SL, с – 4⅓–4½ раза в SL; начало спинного плавника ближе к основанию хвостового плавника, чем к концу рыла. Однако в нём отсутствуют приведённая Бликером формула трёхрядных глоточных зубов (2.4.5/5.4.2) и то, что у изученных Бликером рыб 2-й неветвистый луч спинного плавника короче головы (Bleeker, 1871). При этом Варпаховский приводит в диагнозе, что у H. bleekeri брюхо перед брюшными плавниками уплощённое, а за ними – с очень тупым килем: “der Bauch ist vor den Ventralen abgeflacht, hinter denselben sehr stumpf gekielt” (Warpachowski, 1887. Р. 702). Тогда как Бликер не упомянул в описании наличие/отсутствие киля на брюхе, а в сравнительных замечаниях указал, что у рода Hemiculter (т.е. у изученных им рыб, которых он относил к данному роду) в отличие от рода Culter Basilewsky, 1855 брюшко тупое даже позади брюшных плавников: “par le ventre qui est obtus même en arrière des ventrales” (Bleeker, 1871. Р. 78). При этом Варпаховский в характеристике рода Hemiculter указал, что у этих рыб брюхо в основном с килем: “Bauch meist gekielt” (Warpachowski, 1887. Р. 693), а в ключах видов для H. bleekeri отметил отсутствие киля (“nicht gekielt” − Р. 694).
Отсутствие киля у рыб, изученных Бликером (Bleeker, 1863−1864, 1871), обсуждали в последующем некоторые авторы. Так, Петерс (Peters, 1880, цит. по: Warpachowski, 1887) полагал, что Бликер упустил из виду киль из-за малых размеров исследованных им рыб. Варпаховский (Warpachowski, 1887), однако, считал это очень сомнительным, поскольку в более поздней работе, после опубликования работ Кнера и Гюнтера (Kner, 1867; Günther, 1868 – цит. по: Warpachowski, 1887), Бликер (Bleeker, 1871) (который, кстати, цитировал упомянутых авторов) должен был бы сообщить о своей ошибке. В связи с этими высказываниями следует заметить, во-первых, что в описании Бликера использовались особи общей длиной тела (TL) 136 и 143 мм (Bleeker, 1871), которые крупнее изученных нами рыб из пробы Р-22294 TL 101–129 мм, у которых хорошо заметен киль от грудных плавников до анального отверстия. Во-вторых, как цитируется выше, Бликер (Bleeker, 1871) отмечал отсутствие киля у видов рода Hemiculter как отличие от рыб рода Culter. Берг (1949), который ошибочно относил многотычинковых востробрюшек к виду H. leucisculus, по-видимому, руководствуясь описанием Варпаховского (Warpachowski, 1887), включал H. bleekeri в род Pseudohemiculter Nichols et Pope, 1927, у которого киль развит только в задней части, за брюшными плавниками. В своей ревизии рода Hemiculter Бэнэреску (Bănărescu, 1968) отмечал отсутствие упоминания киля в описаниях Бликера (Bleeker, 1863−1864, 1871), но счёл возможным принять вид H. bleekeri на том основании, что указанное Бликером наличие “42 чешуй” (Bănărescu, 1968. Р. 526) встречается только у этого вида.
К сожалению, типовые экземпляры H. bleekeri отсутствуют (Bănărescu, 1968; Fricke et al., 2021), зато имеется изображение одной из рыб, на основе которых этот вид был описан (Bleeker, 1871. Table 2 . Fig. 1). Эта особь (рис. 2) заметно отличается от изученных нами востробрюшек из р. Янцзы (рис. 3a, 3б) сдвинутым в заднюю часть тела спинным плавником, начало которого в соответствии с описанием Бликера ближе к основанию хвостового плавника (у изученных нами рыб, как у всех многотычинковых востробрюшек, оно ближе к концу рыла), большей длиной головы, которая укладывается не более 4.5 раза (у востробрюшек из коллекции ЗММУ − 4.1–4.7 раза) в SL и более длинными грудными плавниками, почти достающими основания брюшных плавников (далеко не доходят до основания брюшных плавников у многотычинковых востробрюшек из Янцзы: lP/P−V в среднем 79.1%). У изображённой на рисунке рыбы из работы Бликера также более длинная колючка спинного плавника − >70% Н, тогда как у изученных нами рыб − <55%.

Рис. 3.
Hemiculter clupeoides из р. Янцзы, ЗММУ Р-22294: а – мелкий экземпляр SL 81.5 мм, б – более крупный экземпляр SL 103.0 мм, в – вскрытый экземпляр SL 106.0 мм со светлой брюшиной, г – правая глоточная кость, глоточные зубы 2.4.5.

Если не рассматривать отсутствие киля на брюхе, то идентификация Бликером (Bleeker, 1863−1864, 1871) изученных им рыб из Китая как H. leucisculus определённо правильнее отнесения их к многотычинковым востробрюшкам, которых в настоящее время в Китае определяют как H. bleekeri. Действительно, у широко распространённого от бассейна Амура до устья р. Меконг H. leucisculus начало спинного плавника ближе к началу хвостового плавника, чем к концу рыла, а в резко изгибающейся вниз у грудного плавника боковой линии 40–59 (в среднем 46–51) прободённых чешуй, обычно не более 12 ветвистых лучей в анальном плавнике и длинные грудные плавники: lP/P–V 76.6–107.5 (в среднем 90.7 ± 0.68)% (Васильева, Козлова, 1988; Васильева, 2004; Ва- сильева и др., 2022). Однако киль на брюхе у H. leucisculus, как и у всех видов рода Hemiculter в современном понимании, хорошо развит как за брюшными плавниками, так и впереди них.
Проведённый нами сравнительный анализ собственных и литературных данных (Nichols, 1943; Берг, 1949; Chen et al., 1998; Kim, Park, 2002; Dai, Yang, 2003; Kottelat, 2006; Froese, Pauly, 2021) показал, что среди известных пресноводных рыб Китая и пограничных с ним стран карповые рыбы, лишённые киля на брюхе, но обладающие другими признаками из описания H. leucisculus Бликера (Bleeker, 1871), отсутствуют. Нет таких видов и среди рыб, у которых киль развит только между анальным отверстием и брюшными плавниками, к которым относятся представители родов Chanodichthys Bleeker, 1860 и Pseudohemiculter. В то же время родовое название Hemiculter впервые появилось в составленных Бликером (Bleeker, 1860) диагностических ключах для карпообразных рыб. Следуя этим ключам, можно утверждать, что по морфологическим особенностям роды Hemiculter и Chanodichthys очень сходны, их объединяет наличие трёхрядных глоточных зубов и колючки в спинном плавнике. Дифференцирует их Бликер по следующему набору признаков: у рода Chanodichthys тело не удлинённое, рыло заострено, чешуя умеренных размеров, боковая линия слабо изогнутая, а у рода Hemiculter тело удлинённое, рыло короткое, чешуя средних размеров или мелкая, боковая линия сильно изогнутая. В связи с этим мы склонны принять, что замечание Бликера (Bleeker, 1871) по поводу киля у изученных им рыб следует оценивать как сравнительное по отношению к видам с более выраженным острым килем. По другим признакам описания они соответствуют современным представлениям о роде Hemiculter c круто изогнутой боковой линией (“la ligne latérale fortement et subetement courbée” − Bleeker, 1871. Р. 78), D II−III (6)7−8 с гладкой колючкой, A III 10–19, ll 40–59 и трёхрядными глоточными зубами.
Однако, в отличие от многих других авторов (Цзин-цзян, Бе-лу, 1959; Yih, Wu, 1964; Bănărescu, 1968; Luo, Chen, 1998; Fu et al., 2003; Jiang et al., 2008; He, 2010; Li et al., 2020; Fricke et al., 2021; Froese, Pauly, 2021), в соответствии с результатами представленного выше анализа мы считаем H. bleekeri не самостоятельным видом или подвидом, а младшим синонимом H. leucisculus.
Среди названий, пригодных для многотычинковых востробрюшек из р. Янцзы, ранее других был описан H. clupeoides Nichols, 1925. Описание основано на 1 экз. из оз. Тунгтинг (Tungting Lake) (=Дунтинху – Dongting Hu) в провинции Хунань (Hunan), относящегося к бассейну Янцзы: в озеро впадает ряд притоков Янцзы, с основным течением Янцзы оно соединено каналами. Приведённые в описании формулы плавников, глоточных зубов, форма боковой линии, ll ~ 55, наличие на брюхе киля, простирающегося вперёд за брюшные плавники, а также сравнительные замечания по отношению к другим видам рода Hemiculter (Nichols, 1925, 1943) не позволяют сомневаться в родовой принадлежности данной рыбы. При этом типовой экземпляр, безусловно, не конспецифичен H. leucisculus, на что ошибочно указывают ряд авторов (Yih, Wu, 1964; Kottelat, 2006, 2013; Fricke et al., 2021). Согласно описанию, у него 14 ветвистых лучей в анальном плавнике, а судя по рисунку (Nichols, 1943. Fig. 61 . Pl. 4. Fig. 4), спинной плавник начинается значительно ближе к концу рыла, чем к основанию хвостового плавника (aD ~ 48.4% SL), тогда как у H. leucisculus начало спинного плавника сильно сдвинуто в заднюю часть тела и обычно не более 12 ветвистых лучей в анальном плавнике. Следует отметить, что Бэнэреску (Bănărescu, 1968) на основе описания и сравнительного анализа голотипа считал H. clupeoides синонимом H. bleekeri.

По основным характеристикам, дифференцирующим виды в группе многотычинковых востробрюшек, типовой экземпляр H. clupeoides полностью укладывается в основные характеристики изученных нами востробрюшек из р. Янцзы. Длина головы у него содержится 4.6 раза в SL, наибольшая высота тела превышает длину головы и содержится 4.3 раза в SL, длина колючки спинного плавника, судя по рисунку (Nichols, 1943), заметно меньше длины головы (~0.55) и <0.55 высоты тела. В этой связи мы относим востробрюшек р. Янцзы к виду H. clupeoides, валидность которого подтверждается результатами морфологического анализа.
Востробрюшки бассейна нижнего Амура. К числу номинальных видов, описанных из водоёмов с территорий севернее бассейна Янцзы и включаемых разными авторами (Цзин-цзян, Бе-лу, 1959; Bănărescu, 1968; Luo, Chen, 1998) в состав многотычинковых востробрюшек как синонимов H. bleekeri, относятся Toxabramis argentifer Abbott, 1901, Parapelecus elongatus Mori, 1927 и H. shibatae Mori, 1933. T. argentifer рассматривается в настоящее время в статусе самостоятельного вида рода Toxabramis (Fricke et al., 2021), что однозначно подтверждается его описанием (Abbott, 1901). Здесь следует отметить, что вид описан на основе 1 экз. SL 130 мм – голотипа USNM 49545 (=CAS-SU 6299) (Abbott, 1901, Fricke et al., 2021), для которого приведено наличие двурядных глоточных зубов 5.3/3.4 и зазубренного последнего неветвистого луча в спинном плавнике “second spine rather stout, weakly serrated” (Abbott, 1901. Р. 485). Изученные Бэнэреску (Bănărescu, 1968) две востробрюшки из коллекции Стэнфордского университета (SU 6300) с трёхрядными глоточными зубами в описании Эбботта не указаны и не могут считаться паратипами указанного таксона: исходно допущенная Бэнэреску ошибка привела к неправильному заключению.
P. elongatus был описан по 1 экз. TL 107 мм из р. Хан (Hun), притока р. Ляохэ (Liao River), впадающей в Ляодунский залив в Южной Маньчжурии. Этот экземпляр, современное нахождение которого неизвестно, характеризовался следующими основными признаками: D II 7, А II 13, ll 48, брюшко заострено от уровня грудных плавников до анального отверстия, начало спинного плавника существенно ближе к основанию хвостового плавника, чем к концу рыла (на расстояние равное дистанции от конца рыла до середины глаза), грудные плавники длинные, заострённые достигают ¾ P−V (Mori, 1927). На основе этих характеристик его следует считать конспецифичным H. leucisculus и, соответственно, P. elongatus должен быть включён в синонимы этого вида.
H. shibatae был описан по из 1 экз. из Тси-наня (Tsi-nan, в настоящее время Jinan – Цзинань, центр провинции Шаньдун), водоёмы которого принадлежат бассейну р. Хуанхэ и относятся к западному побережью Жёлтого моря. В описании (Mori, 1933) указано, что у этого вида D II 7, A II 13, ll 41, глоточные зубы трёхрядные 5.4.2 / 2.4.5; длина головы содержится 4.5 раза в SL, высота тела – 4.3 раза в SL; глаза небольшие – 1.3 раза в длине рыла; на брюхе киль от основания грудных плавников до анального отверстия; начало спинного плавника на равном расстоянии от конца рыла и от основания хвостового плавника, чуть позади основания брюшных плавников; 1-я колючка спинного плавника примерно (“nearly”: Mori, 1933. Р. 166) такой же длины, как и 2-я; брюшина чёрная. Сведения о числе жаберных тычинок в описании отсутствуют. Отмечается, что новый вид очень близок к H. bleekeri из Янцзы и Шань-си, но отличается заметно более высоким телом, меньшим размером глаз, более короткими грудными плавниками и более сдвинутым вперёд спинным плавником.
Место нахождения типового экземпляра H. shibatae, принадлежность которого роду Hemiculter не вызывает сомнений, неизвестно. На основе описания и приведенного рисунка (рис. 4) определить видовой статус экземпляра не представляется возможным. На малотычинковых востробрюшек, которые, согласно генетическим данным (Chen et al., 2017; Васильева и др., 2022), представлены в типовом месте обитания H. shibatae широко распространённым видом H. leucisculus, он похож наличием небольших глаз (у многотычинковых востробрюшек горизонтальный диаметр глаза обычно превышает длину рыла, у H. leucisculus заметно короче рыла) и положением начала спинного плавника по отношению к брюшным плавникам (у многотычинковых видов начало спинного плавника обычно на уровне основания брюшных плавников и aV/aD обычно >95%, тогда как у H. leucisculus обычно <92%). В то же время у малотычинковых востробрюшек начало спинного плавника, как правило, ближе к основанию хвостового плавника, чем к концу рыла, и aD превышает 51.5% SL, а у многотычинковых − начало спинного плавника обычно ближе к концу рыла, но aD варьирует от 46.0 до 54.4% SL (Васильева, Козлова, 1988, 1989а, 1989б; Васильева и др., 2022, настоящая работа). Таким образом, по последнему признаку тип H. shibatae сходен с многотычинковыми видами. Однако среди многотычинковых востробрюшек особи с чёрной брюшиной обнаружены нами только у H. lucidus, но у этого вида, по нашим данным, в анальном плавнике, как правило, не менее 14 ветвистых лучей, длина головы содержится обычно 5 и более раз в SL, а высота тела – не более 4 раз в SL. Среди вскрытых малотычинковых востробрюшек особи с чёрной брюшиной не выявлены: у описанного нами нового вида из бассейна Жемчужной реки (Васильева и др., 2022) брюшина светлая, как и у трёх особей H. leucisculus из низовьев Амура (пробы ЗММУ Р-7795, Р-6155), коричневая брюшина − у единственного экземпляра пробы Р-9188 из Вьетнама. В связи с изложенным мы считаем, что до проведения генетических и морфологических исследований востробрюшек из типового места обитания H. shibatae название shibatae следует считать nomen dubium. Вполне возможно, что в этом районе обитает самостоятельный вид, но также можно полагать и гибридное происхождение типового экземпляра.
Многотычинковые востробрюшки бассейна нижнего Амура в настоящей работе описаны в качестве нового для науки вида. Описание приводится ниже вместе с диагнозами (составлены на основе собственных данных и данных цитированной при синонимии литературы), основной синонимией и сведениями по распространению остальных видов группы многотычинковых востробрюшек. В диагнозы не включены характеристики, общие для всех видов группы lucidus: начало спинного плавника, как правило, ближе к концу рыла, чем к основанию хвостового плавника, aD обычно < 50% SL; плавательный пузырь двухкамерный, вторая камера удлинённая, с узкой полосой соединительной ткани, образующей несколько витков спирали; задний конец второй камеры заострён, с коротким отростком, икра пелагическая.
Hemiculter lucidus (Dybowski, 1872) – ханкайская востробрюшка
(рис. 5)
Рис. 5.
Ханкайская востробрюшка Hemiculter lucidus SL 150.0 мм из оз. Ханка, ЗММУ Р-8343: внешний вид (а) и брюшная часть со вскрытой тёмной брюшиной (б).

Culter lucidus Dybowski, 1872. P. 214 (Chankasee – оз. Ханка).
Culter lucidus: Дыбовский, 1877. C. 14.
Hemiculter lucidus: Варпаховский, Герценштейн, 1887. C. 46; Warpachowski, 1887. P. 704; Васильева, Козлова, 1988. C. 894 (partim); Васильева, Козлова, 1989а. C. 22 (partim); Богуцкая, Насека, 1996. C. 26 (partim); Насека, 1998. C. 76 (partim); Богуцкая, Насека, 2004. C. 58 (partim); Васильева, 2004. C. 151 (partim); Bogutskaya et al., 2008. P. 319.
Hemiculter leucisculus lucidus: Берг, 1909. C. 150; Никольский, 1947. C. 775, 776; Берг, 1949. C. 809; Никольский, 1956. C. 301.
Hemiculter bleekeri lucidus: Цзин-цзян, Бе-лу, 1959. C. 166; Yih, Wu, 1964. Р. 88.
Hemiculter lucidus lucidus: Bănărescu, 1968. Р. 526; Luo, Chen, 1998. Р. 169.
D II−III 7, А III (13) 14–17, как правило, не менее 14 ветвистых лучей, V II 7–8, ll 40–52, sp.br. 20–29; глоточные зубы трёхрядные, 5.4.2/2.4.4, 5.4.1/1.3.4, 4.4.2/2.4.4; киль на брюхе хорошо выражен, в передней части доходит до горла; длина колючки спинного плавника, как правило, >0.85 c, обычно почти равна или более длины головы; голова относительно короткая, обычно укладывается 5 и более раз в SL; тело высокое, высота тела всегда больше длины головы и обычно заметно больше длины колючки спинного плавника (длина последней чаще составляет 0.68–0.92 Н); брюшина тёмная; кожный подключичный вырост короткий, его длина равна или чуть превышает диаметр зрачка и составляет от 36.4 до 62.7% горизонтального диаметра глаза (о). Распространён в бассейне оз. Ханка, где симпатричен с видом малотычинковых востробрюшек H. leucisculus.
Hemiculter varpachovskii Nikolski, 1903 – буирнурская востробрюшка
(рис. 6)
Рис. 6.
Буирнурская востробрюшка Hemiculter varpachovskii SL 121.0 мм из оз. Буйр-Нуур (Монголия), ЗММУ Р-8069: внешний вид (а) и вид со стороны вскрытой тёмной брюшины (б).

Hemiculter varpachovskii Nikolski. 1903. P. 359 (Lac Buir-Nor – оз. Буйр-Нуур).
Hemiculter leucisculus (non Basilewsky, 1855): Берг, 1909. C. 146 (partim); Nichols, 1943. Р. 134 (partim); Никольский, 1947. C. 775 (partim); Берг, 1949. C. 808 (partim); Богуцкая, Насека, 1996. C. 25 (partim); Насека, 1998. C. 75 (partim); Kottelat, 2006. Р. 35 (partim); Ocock et al., 2006. Р. 37.
Hemiculter varpachowskii (sic): Никольский, 1947. C. 774.
Hemiculter varpachovskii: Даши-доржи, 1955. C. 571, 574; Kottelat, 2006. Р. 36; Ocock et al., 2006. Р. 13; Bogutskaya et al., 2008. Р. 320.
Hemiculter leucisculus warpachowskii (sic): Никольский, 1956. C. 298; Баасанжав и др., 1983. C. 194; Luo, Chen, 1998. Р. 169.
Hemiculter bleekeri warpachowskii (sic): Цзин-цзян, Бе-лу, 1959. C. 166.
Hemiculter lucidus warpachowskii (sic): Bănărescu, 1968. Р. 527.
Hemiculter lucidus (non Dybowski, 1872): Васильева, Козлова, 1988. C. 894 (partim); Васильева, Козлова, 1989а. C. 22 (partim); Богуцкая, Насека, 1996. C. 26 (partim); Насека, 1998. C. 76 (partim); Васильева, 2004. C. 151 (partim).
D II−III (6) 7 (8), A III (12) 13–16, чаще 14 или 15 ветвистых лучей, V II 7–9, ll 40–55, sp.br. 20–28, глоточные зубы 4–5.3–4.1–2/2.4.4–5; киль на брюхе хорошо выражен, доходит до горла; длина колючки спинного плавника всегда меньше длины головы (обычно больше 0.75, но менее 0.9); длина голова содержится менее 5 раз в SL; высота тела обычно не превышает длину головы, но обычно заметно больше длины колючки спинного плавника, длина которой составляет от 0.67 до 1.06 Н; брюшина тёмная; кожный подключичный вырост короткий, его длина заметно меньше величины зрачка и составляет 44–51% о.
Распространён в бассейне р. Аргунь, верхнего течения Амура: озёра Буйр-Нуур, Далайнор, реки Халхин-гол, Уршун и Керулен. Ошибочное отнесение многими авторами востробрюшек этого региона к малотычинковому виду H. leucisculus (именно так были исходно определены все пробы ЗММУ, кроме Р-22031) послужило основанием для утверждений о том, что в оз. Буйр-Нуур обитают два вида востробрюшек (Богуцкая, Насека, 1996; Насека, 1998; Kottelat, 2006; Ocock et al., 2006). Из этого региона мы изучили в общей сложности 17 проб, включающих 188 экз., и все они относились к одному виду, тогда как в пробах из других бассейнов, где виды группы H. lucidus симпатричны с видами малотычинковых востробрюшек, всегда встречались особи двух видов. Это позволяет полагать, что виды из группы малотычинковых востробрюшек в бассейне верхнего течения Амура не встречаются, что соответствует сделанному ранее заключению об их более южном происхождении (Васильева, Козлова, 1988).
Hemiculter clupeoides Nichols, 1925 – китайская востробрюшка
(рис. 3)
Hemicultur clupeoides (sic) Nichols, 1925. P. 7 (Tungting Lake, Hunan).
Hemiculter clupeoides: Nichols, 1943. P. 135; Bănărescu, 1968. P. 525, 526 (partim).
Hemiculter leucisculus (non Basilewski, 1855): Nichols, 1943. Р. 134 (partim); Берг, 1949. C. 808 (partim); Kim, Park, 2002. Р. 192 (partim); Kottelat, 2006. P. 35 (partim).
Hemiculter bleekeri bleekeri (non Warpachowski, 1888): Yih, Wu, 1964. Р. 87; Jiang et al., 2008. Р. 300.
Hemiculter bleekeri (non Warpachowski, 1888): Цзин-цзян, Бе-лу, 1959. C. 163 (partim); Bănărescu, 1968. Р. 525 (partim); Luo, Chen, 1998. Р. 167; Fu et al., 2003. Р. 1669; He, 2010. Р. 9; Li et al., 2020. Р. 1.
Hemiculter jabouilei (sic) (non Chevey, 1936): Bănărescu, 1968. Р. 525, 526 (partim).
Hemiculter lucidus (non Dybowski, 1872): Васильева, Козлова, 1988. C. 894 (partim); Васильева, Козлова, 1989а. C. 22 (partim); Богуцкая, Насека, 1996. C. 26 (partim); Насека, 1998. C. 76 (partim).
D II–III 7, A III 12–14 (15), обычно не более 13 ветвистых лучей, V II 7–9, ll 40–51, sp.br. 20–30, глоточные зубы (4)5.(3)4.(1)2/(1)2.4.4(5); киль на брюхе тянется от анального отверстия до горла, в самой передней части иногда плохо выражен; длина колючки спинного плавника <0.7 с (обычно <0.6); длина головы содержится < 5 раз в SL; высота тела соизмерима с длиной головы (0.87–1.20) и обычно в 1.5–2.0 раза превышает длину колючки спинного плавника, как правило, не более 0.55 Н; брюшина светлая; кожный подключичный вырост маленький, его длина заметно меньше диаметра зрачка и не достигает 40.0 (26.4–37.3)% о.
Распространён в бассейне Янцзы. Единственный изученный нами гаплотип гена cyt b особи из Кореи принадлежит к одной филетической линии с востробрюшками Янцзы (см. ниже), поэтому многотычинковых востробрюшек Кореи (sp.br. 26–32, ll 45–49), которых Ким и Парк (Kim, Park, 2002) определяют как H. leucisculus, возможно, следует также относить к виду H. clupeoides. Конспецифичность популяций из других речных бассейнов Китая нуждается в подтверждении, в частности, популяций из речных систем провинции Шаньдун (Shandong) (основные речные системы Хуанхэ и Вэйхэ), сходство которых с ихтиофауной Янцзы отмечалось ранее (Yang et al., 1994) и откуда был описан H. shibatae. Очень сомнительно присутствие H. clupeoides в водах Вьетнама, где он отмечен Орси (Orsi, 1974). В притоках верхнего течении Янцзы в районе Сычуаня H. clupeoides встречается симпатрично с эндемичным видом, для которого, согласно нашим исследованиям (Васильева и др., 2022), валидным пригодным названием является H. nigromarginis Yih et Wu, 1964, а на большей части реки обитает совместно с H. leucisculus (Fu et al., 2003; He, 2010) – другим видом филетической группы малотычинковых востробрюшек, широко распространённым в водах Китая и в бассейне Амура.
В число синонимов H. bleekeri (non Warpachowski, 1888) Бэнэреску (Bănărescu, 1968) включал вид из бассейна вулканического оз. Куанг-Чоу-Ван (Kouang-Tchéou-Wan), Южный Китай, провинция Гуандун – H. jabouillei Chevey, 1935 – на основе его описания (местонахождение голотипа неизвестно). Согласно этому описанию (Chevey, 1935), вид характеризуется D II 7, А II 14, ll 44; киль по всему брюху (“abdomen entirèment caréné” – Chevey, 1935. P. 431); длина головы намного больше высоты тела и содержится 3.56 раза в SL, высота тела – 5.66 раза в SL. Судя по рисунку (Chevey, 1935. Fig. 3), начало спинного плавника позади основания брюшных плавников и существенно ближе к основанию хвостового плавника, чем к концу рыла; длина колючки спинного плавника ~ 0.6 с и ~ 0.8 Н. Коттла (Kottelat, 2013) включает H. jabouillei в синонимы Pseudohemiculter hainanensis (Boulenger, 1900), что является ошибкой, поскольку Шёве (Chevey, 1935) различал собственно Hemiculter и Pseudohemiculter и подчёркивал, что описанный им вид относится к первой группе видов, поскольку у него киль начинается от грудных плавников, а не от брюшных, как у видов группы Pseudohemiculter. От всех многотычинковых востробрюшек H. jabouillei отличается положением спинного плавника и не соответствует диагнозу H. clupeoides по относительной длине головы, высоте тела и длине колючки спинного плавника. Не согласуются характеристики H. jabouillei и с диагнозом вида малотычинковых востробрюшек, описанного нами из водоёмов провинции Гуандун, у которого A III 11–13, ll 49–56, наибольшая высота тела укладывается 4.0–4.7 раза в SL, длина головы – 3.4–4.4, обычно 4 раза в SL (Васильева и др., 2022). Поэтому очевидно, что jabouillei следует считать сомнительным названием.
Hemiculter nikolskyi Vasil’eva, Vasil’ev et Shedko sp. nova – востробрюшка Никольского
(рис. 7–9)

Рис. 8.
Паратипы Hemiculter nikolskyi sp. nova, ЗММУ Р-7502, р. Амур у Елабуги: а –плавательный пузырь особи SL 142 мм, б – то же особи SL 113 мм; в – правая глоточная кость, глоточные зубы 2.3.4.

Рис. 9.
Hemiculter nikolskyi sp. nova SL 98 мм из р. Амур в районе Елабуги, ЗММУ Р-6405, вскрытый экземпляр с тёмной брюшиной.

Hemiculter leucisculus (non Basilewsky, 1855): Берг, 1909. C. 146 (partim); Никольский, 1947. C. 773 (partim); Берг, 1949. C. 808 (partim); Kim, Park, 2002. Р. 192 (partim).
Hemiculter leucisculus leucisculus (non Basilewsky, 1855): Никольский, 1956. C. 290.
Hemiculter bleekeri (non Warpachowski, 1888): Цзин-цзян, Бе-лу, 1959. C. 163 (partim); Bănărescu, 1968. Р. 525 (partim).
Hemiculter lucidus (non Dybowski, 1872): Васильева, Козлова, 1988. C. 894 (partim); Васильева, Козлова, 1989а. C. 22 (partim); Богуцкая, Насека, 1996. C. 26 (partim); Насека, 1998. C. 76 (partim); Богуцкая, Насека, 2004. C. 58 (partim); Васильева, 2004. C. 151 (partim).
Hemiculter cf. lucidus: Bogutskaya et al., 2008. Р. 320.
Материал. Голотип Р-21562 − TL 128 мм, SL 102 мм, Амур у Безымянного (выше Хабаровска), 07.08.2004 г., коллектор Е.Д. Васильева. Паратипы Р-7502 − 2 экз. TL 174 и 142 мм, SL 142 и 113 мм, Амур у Елабуги, 24.07.1949 г., коллектор Т.К. Сысоева. Дополнительный материал представлен в разделе “Материал и методика” как Hemiculter sp.
Диагноз. D II (III) (6) 7 (8), А III (10, 11) 12–16 (17), обычно 13–14 ветвистых лучей, P I (10) 11–14, V II 8–9, ll 40–52, sp.br. 20–30, глоточные зубы 4–5.3–4.1–2/1–2.3–4.4–5; брюхо с хорошо выраженным килем от горла до анального отверстия; длина колючки спинного плавника всегда меньше длины головы, обычно не превышает 0.75 с; голова содержится < 5 раз в SL; высота тела обычно больше длины головы (в разных локальных популяциях Н/с в среднем 0.92–1.16) и всегда заметно больше длины колючки спинного плавника, длина которой составляет от 0.50 до 0.83 Н; кожный подключичный вырост маленький, его длина менее или равна диаметру зрачка и не превышает 50.0 (29.0–50.0)% о; брюшина тёмная.
Описание голотипа. D II 7, A III 12, P I 14, V II 8, ll ~ 49 (чешуя сохранилась не полностью), sp.br. 29; брюхо заострено, киль выражен от анального отверстия до конца межжаберного промежутка; спинной плавник начинается ближе к концу рыла, чем к основаниям средних лучей хвостового плавника, aD составляет 49.9% SL; голова небольшая, 22.8% SL; рот верхний; жаберные тычинки короткие, густые; глаз большой, горизонтальный диаметр глаза содержится ~ 4 раза в с; тело невысокое, H 21.8% SL, высота тела составляет 0.95 с; боковая линия не очень круто опускается вниз, достигает самой низкой точки не доходя до конца грудных плавников, и вновь поднимается к средней линии тела за анальным плавником, делая резкий изгиб вверх; грудные плавники удлинённые – 20.8% SL, длина брюшных плавников 14.7% SL; последний неветвистый луч спинного плавника колючий с короткой мягкой частью на самой вершине, длина колючки составляет 0.72 с и 0.76 H; хвостовой плавник глубоко вырезанный. Кожный подключичный вырост маленький, плохо заметный, его длина менее диаметра зрачка и 29.0% о. Фиксированный изначально в 4%-ном растворе формальдегида и переведённый в 75%-ный этанол голотип имеет коричневато-жёлтую окраску брюха, спина и верхняя часть головы коричневые; от верхнего края жаберной крышки вдоль всего тела проходит продольная тёмно-коричневая полоса, отделяющая коричневую спину от более светлой нижней части, спинной и хвостовой плавники сероватые, с заметными тёмными крапинками вдоль лучей; нижние плавники светлые; глаз черноватый, с чёрной окантовкой по нижнему краю (рис. 7).
Краткая характеристика паратипов. D II 7, A III 13−14, P I 14, V II 8, ll 45−49, sp.br. 27, глоточные зубы 4.3.1−2/2.3.4 (рис. 8в); спинной плавник начинается ближе к концу рыла, киль доходит вперёд до оснований грудных плавников, длина колючки 0.57–0.61 Н и 0.66–0.82 с, Н 1.16–1.33 с; кожный подключичный вырост маленький, плохо заметный, его длина менее диаметра зрачка и составляет 31.9% о у более крупного экземпляра и 46.4% − у более мелкого. Брюшина у крупного экземпляра тёмно-коричневая, плавательный пузырь двухкамерный, типичной формы: вторая камера удлинённая, с узкой полосой соединительной ткани, образующей несколько витков спирали; задний конец второй камеры заострён, с коротким отростком (рис. 8а). У особи меньших размеров брюшина светло-коричневая, плавательный пузырь атипичной формы, с изогнутой второй камерой (рис. 8б). Пигментация у паратипов не сохранилась, всё тело и плавники светло-коричневые.
Изменчивость морфологических признаков. Ранее был проведён подробный анализ межпопуляционной изменчивости морфометрических характеристик у многотычинковых востробрюшек бассейна нижнего Амура на основе попарного сравнения выборок из оз. Болонь, русла Амура в районе пос. Новогеоргиевка, Комсомольска-на-Амуре, Елабуги и из оз. Кабар (Васильева, Козлова, 1989а). Достоверные различия выявлены по всем пластическим характеристикам, кроме относительной высоты спинной колючки, часть из них обусловлена размерной изменчивостью признаков и разнокачественностью выборок по длине тела особей. Из меристических признаков достоверные межпопуляционные различия выявлены лишь по числу чешуй в боковой линии и ветвистых лучей в грудном плавнике, число последних варьирует от 10 до 14, а средние выборочные значения – от 11.4 до 12.4. Однако эти различия невелики (СD варьирует от 0.20 до 1.12). Не было выявлено направленного изменения морфометрических характеристик у популяций, обитающих в разных гидрологических условиях (озёрных и речных), а также отмеченной Никольским (1947, 1956) тенденции уменьшения числа чешуй, лучей в анальном плавнике и относительной длины основания анального плавника и длины хвостового стебля в направлении вниз по течению. Это позволило дать суммарную характеристику пластических признаков нижнеамурских востробрюшек (Васильева, Козлова, 1989а). На основании этих данных было отмечено, что нижнеамурские востробрюшки достоверно отличаются от выборок трёх других видов по ряду пропорций тела, представленных в таблице по данным из цитируемой публикации, однако в большинстве случаев выявленные различия невелики. Признаки, оказавшиеся пригодными для диагностики видов, рассмотрены выше (в начале раздела “Результаты и обсуждение”).
Прижизненная окраска: спина зеленовато-серая, бока и брюхо серебристые; спинной плавник зеленовато-серый, парные, анальный и хвостовой плавники – желтовато-серые (Никольский, 1956; наши данные).
Этимология. Вид назван в честь известного ихтиолога Г.В. Никольского, организатора и активного участника Амурских экспедиций, сборы которых послужили основой для выполнения настоящего исследования.
Распространение. Согласно настоящим исследованиям и данным Никольского (1956), вид распространён в Сунгари и бассейне нижнего и среднего течения Амура от п. Свербеево до с. Даерги, наиболее обычен от с. Джалинда до оз. Кизи (Большое Кизи). По всему ареалу H. nikolskyi распространён симпатрично с малотычинковыми востробрюшками H. leucisculus, но он гораздо более многочислен, о чём свидетельствует как видовой состав рода Hemiculter из нижнего Амура в коллекции ЗММУ, так и сведения Никольского (1947) о соотношении этих видов в уловах: в озерке у Даерги в улове на 160 экз. H. nikolskyi приходился только 1 экз. H. leucisculus, а в улове в оз. Петропавловском их соотношение было 186 : 7. По-видимому, относительная численность видов обусловлена особенностями их размножения: H. nikolskyi вымётывает пелагическую икру, и его нерест не ограничен наличием нерестового субстрата, в отличие от H. leucisculus, у которого икра приклеивается к субстрату. Возможно также, что последний вид держится в водоёмах преимущественно на течении и избегает озёрных биотопов, где чаще отлавливали востробрюшек.
Филогенетические отношения многотычинковых востробрюшек группы H. lucidus
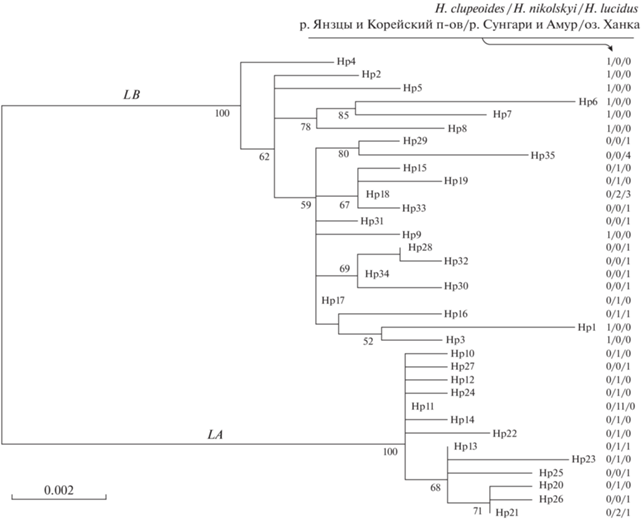
По результатам секвенирования полноразмерных последовательностей гена cyt b у 55 многотычинковых востробрюшек (MW367253–MW367260, MW508412–MW508458) было выявлено 34 различных варианта гаплотипов. Последовательность cyt b, полученная от востробрюшки из Южной Кореи (KF760461), оказалась ещё одним уникальным вариантом. Построенное методом максимального правдоподобия филогенетическое дерево для этих 35 уникальных гаплотипов приведено на рис. 10. Выявленные гаплотипы сформировали две клады – LA и LB, включающие соответственно 13 и 22 гаплотипа. Средние дистанции TN (Tamura, Nei, 1993) выражены в числе нуклеотидных замен на позицию; дистанции, рассчитанные с применением программы SENDBS (Nei, Jin, 1989), внутри клад оказались равными − соответственно 0.0027 ± 0.00071 и 0.0052 ± 0.00086, а между кладами – 0.0179 ± 0.0037 (без коррекции) и 0.0139 ± ± 0.0036 (нетто – т.е., исключая изменчивость внутри клад; дистанция рассчитана согласно уравнению 10.21 Нэя (Nei, 1987)). Средние дистанции TN между гаплотипами из клад LA и LB многотычинковых востробрюшек оказались намного меньше, чем дистанции (общие и нетто, рассчитаные нами) между кладами A, B и C малотычинковых востробрюшек из работы Чена с соавторами (Chen et al., 2017): в 2.5–2.7 раза при сравнении с дистанциями между кладами B и C, в 3.4–3.8 раза – между кладами A и B, в 3.8–4.3 раза – между кладами A и C.
Рис. 10.
Оптимальное ML-дерево (ln L = –2118.5252, модель нуклеотидных замещений TN + F + G4), которое построено для 35 гаплотипов гена cyt b (Hp1–Hp35), выявленных у многотычинковых востробрюшек Hemiculter spp. из трёх районов: р. Янцзы и Корейского п-ова, рек Сунгари и Амур, оз. Ханка. Справа разделённые косой чертой цифры – встречаемость гаплотипов в выборках востробрюшек из трёх указанных районов. Цифры у основания клад – оценки бутстреп-поддержки (в % от 2000 циклов). Масштаб длины ветвей указан слева внизу в числе замен на сайт.
Основываясь на калибровке хода молекулярных часов, проведённой для гена cyt b у карповых рыб (0.011 замен/сайт/млн лет – Smith et al., 2002), и применяя её к нашей оценке нетто-дистанции (0.0139 ± 0.0036), эволюционный возраст клад LA и LB можно принять равным 1.26 ± 0.33 млн лет.
У большинства образцов нового вида многотычинковых востробрюшек H. nikolskyi присутствовали гаплотипы клады LA (21 из 27, или 78.8%). У образцов ханкайской востробрюшки H. lucidus, напротив, преобладали гаплотипы клады LB (15 из 20, или 75.0%). Различия H. nikolskyi и H. lucidus по соотношению гаплотипов из клад LA и LB достоверны, согласно точному критерию Фишера (р = 0.0004). У востробрюшек из р. Янцзы (H. clupeoides) и экземпляра с Корейского п-ова были найдены только гаплотипы из клады LB. Учитывая эту картину, а также то, что гаплотипы, занимающие базальное положение в кладе LB (Hp2, 4–8), принадлежат образцам из р. Янцзы, можно предложить следующую филогенетическую гипотезу. Общий предок многотычинковых востробрюшек разделился на две группировки – бассейн Амура и водоёмы, расположенные к югу от указанного бассейна. По прошествии некоторого времени южная линия многотычинковых востробрюшек вселилась в бассейн Амура вновь, принеся с собой гаплотипы из клады LB. Со временем она здесь исчезла, сохранившись лишь в виде линии ханкайской востробрюшки. Репродуктивная изоляция между южной и аборигенной для бассейна Амура линиями многотычинковых востробрюшек оказалась неполной, что и привело к частичному смешению их митохондриальных генофондов.
Нетто-дистанция между выборкой многотычинковых востробрюшек из оз. Ханка и выборкой из р. Янцзы и Корейского п-ова равна 0.0006 ± 0.0004 замен на позицию (расчёт проводился только по гаплотипам из клады LB). Тогда время, прошедшее с момента вторичного вселения южной линии в бассейн Амура, можно определить в 54 ± 33 тыс. лет. С целью сравнения по данным из предыдущей работы (Васильева и др., 2022) была рассчитана нетто-дистанция между выборкой H. leucisculus из бассейна Амура (n = 21) и объединённой выборкой востробрюшек этого вида из Пекина и Корейского п-ова (n = 14). Она оказалась равной 0.0007 ± 0.0007 замен на позицию и близкой к оценке, полученной для вышеописанного аналогичного случая с многотычинковыми востробрюшками. То есть вселение в бассейн Амура H. leucisculus и предка H. lucidus произошло, по всей видимости, в одно и то же время.
Хотя генетические дистанции по гену cyt b между многотычинковыми востробрюшками невысоки (в сравнении с малотычинковыми востробрюшками), отличия H. nikolskyi от H. lucidus (а также и от H. clupeoides) по составу гаплотипов подтверждают его репродуктивную изоляцию и, следовательно, самостоятельный видовой статус.
Отметим, что, несмотря на существенную морфологическую дивергенцию, ханкайская востробрюшка, как показывают наши данные, является молодым видом. К сожалению, вплоть до настоящего момента H. varpachovskii генетическими методами не исследовали. Водоёмы, в которых обитает буирнурская востробрюшка, расположены в Восточно-Монгольской степной ландшафтно-климатической области, характеризующейся обилием небольших бессточных, солоноводных и периодически пересыхающих озёр; оз. Буйр-Нуур – самое крупное из немногочисленных пресноводных озёр. Поздний плейстоцен и голоцен на этой территории отличались значительными колебаниями увлажнённости и стока; уровни озёр, близкие к современным, датируются средним голоценом (приблизительно 5000−4500 лет назад) (Дорофеюк, 2008). Небольшой возраст озёр позволяет полагать, что H. varpachovskii − тоже молодая филетическая линия, сформировавшаяся по тому же сценарию, что и линия ханкайской востробрюшки: наличие особой ниши (крупных озёр) запустило процесс дифференциации. Следует подчеркнуть, что для более полного понимания эволюционной истории многотычинковых востробрюшек совершенно необходим анализ популяций как из бассейна верхнего Амура, так и из южной части ареала этой группы видов.
В заключение мы считаем необходимым дать ключ для диагностики многотычинковых видов востробрюшек.
КЛЮЧ ДЛЯ ИДЕНТИФИКАЦИИ ВИДОВ МНОГОТЫЧИНКОВЫХ ВОСТРОБРЮШЕК ГРУППЫ HEMICULTER LUCIDUS
1 (2) Длина головы обычно 5 и более (4.2–5.4) раз в длине тела; длина колючки спинного плавника обычно равна длине головы; в анальном плавнике обыкновенно не менее 14 ветвистых лучей; бассейн оз. Ханка …….….…. Hemiculter lucidus
2 (1) Длина головы менее 5 раз в длине тела; длина колючки спинного плавника меньше длины головы; в анальном плавнике 12–16 ветвистых лучей …..……..….......................................………….. 3
3 (4) В анальном плавнике обычно не более 13 ветвистых лучей; длина колючки спинного плавника обычно не превышает 55% высоты тела; брюшина светлая ….………….. Hemiculter clupeoides
4 (3) В анальном плавнике обычно не менее 13 ветвистых лучей; длина колючки спинного плавника обычно более 60% высоты тела; брюшина тёмная …..….…………..…….........................… 5
5 (6) Длина колючки спинного плавника обычно более 80% высоты тела, в среднем 19.2–20.4% SL; высота тела обычно меньше длины головы (в разных локальных популяциях H/c в среднем 0.96–1.00); бассейн верхнего Амура (озёра Буйр-Нуур, Далайнур) ……………Hemiculter varpachovskii
6 (5) Длина колючки спинного плавника обычно меньше 70% высоты тела, в среднем 15.2–17.6% SL; высота тела обычно больше длины головы (в разных локальных популяциях H/c в среднем 0.92–1.16); бассейн нижнего Амура ...........… Hemiculter nikolskyi
Список литературы
Баасанжав Г., Дгебуадзе Ю.Ю., Демин А.Н. и др. 1983. Обзор видов ихтиофауны МНР // Рыбы Монгольской Народной Республики. М.: Наука. С. 102–224.
Берг Л.С. 1909. Рыбы бассейна Амура // Зап. Императ. АН. Т. 24. № 9. 270 с.
Берг Л.С. 1949. Рыбы пресных вод СССР и сопредельных стран. Ч. 2. М.; Л.: Изд-во АН СССР. С. 469–926.
Богуцкая Н.Г., Насека А.М. 1996. Круглоротые и рыбы бассейна озера Ханка (система реки Амур): Аннотированный список видов с комментариями по их таксономии и зоогеографии региона. СПб.: Изд-во ГосНИОРХ. 89 с.
Богуцкая Н.Г., Насека А.М. 2004. Каталог бесчелюстных и рыб пресных и солоноватых вод России с номенклатурными и таксономическими комментариями. М.: Т-во науч. изд. КМК, 389 с.
Варпаховский Н., Герценштейн С. 1887. Заметки по ихтиологии бассейна р. Амура и прилежащих стран // Тр. СПб. о-ва естествоиспыт. Отд. зоология. Т. 18. № 7. С. 1–58.
Васильева Е.Д. 2004. Популярный атлас-определитель. Рыбы. М.: Дрофа, 400 с.
Васильева Е.Д., Козлова M.C. 1988. О таксономии востробрюшек рода Hemiculter (Cyprinidae) Советского Союза // Вопр. ихтиологии. Т. 28. Вып. 6. С. 883–895.
Васильева Е.Д., Козлова М.С. 1989а. Внутривидовая структура и изменчивость ханкайской востробрюшки Hemiculter lucidus (Dybowski) (Pisces, Cyprinidae) // Бюл. МОИП. Т. 94. Вып. 1. С. 22–36.
Васильева Е.Д., Козлова М.С. 1989б. К морфологической характеристике обыкновенной востробрюшки Hemiculter leucisculus (Basilewsky) (Pisces, Cyprinidae) // Там же. Т. 94. Вып. 3. С. 36–46.
Васильева Е.Д., Васильев В.П., Немкова Г.А., Шедько С.В. 2022. Филогенетические отношения и таксономия карповых рыб рода Hemiculter (Cyprinidae, Xenocypridinae): востробрюшки видовой группы Hemiculter leucisculus // Вопр. ихтиологии. Т. 62. № 1. С. 3–18.
Даши-доржи А. 1955. Материалы по ихтиофауне верховьев Селенги и Амура в пределах Монголии // Зоол. журн. Т. 34. Вып. 3. С. 570–577.
Дорофеюк Н.И. 2008. Реконструкция природных условий Внутренней Азии в позднеледниковье и голоцене (по материалам диатомового и палинологического анализов озерных осадков Монголии): Автореф. дис.… докт. биол. наук. М.: ИПЭЭ РАН, 49 с.
Дыбовский Б. 1877. Рыбы системы вод Амура // Изв. Сиб. отд. Императ. Рус. геогр. о-ва. Т. 8. № 1–2. С. 1–29.
Линдберг Г.У., Таранец А.Я. 1929. Список рыб Владивостокского государственного музея // Зап. Владивосток. отд. гос. Рус. геогр. о-ва. Т. 4. Вып. 21. С. 221–266.
Майр Э., Линсли Э., Юзингер Р. 1956. Методы и принципы зоологической систематики. М.: Иностр. лит-ра, 352 с.
Международный кодекс зоологической номенклатуры. 2000. СПб.: Изд-во ЗИН РАН, 221 с.
Насека А.М. 1998. Подсемейство Cultrinae // Аннотированный каталог круглоротых и рыб континентальных вод России. М.: Наука. С. 74–77.
Никольский А.М. 1903. Новые виды рыб из восточной Азии. Acanthogobio paltschevskii sp. n., Acanthogobio oxyrhynchus sp. n., Hemiculter varpachovskii sp. n., Hemicultrella soldatovi sp. n. (Cyprinidarum); et Ussuria leptocephala gen et sp. n. (Cobitidarum) // Ежегодник Зоомузея Императ. АН. Т. 8. С. 356–363.
Никольский Г.В. 1947. О роде Hemiculter (Pisces, Cyprinidae) в бассейне Амура // Докл. АН СССР. Т. 56. № 7. С. 773–776.
Никольский Г.В. 1956. Рыбы бассейна Амура. М.: Изд-во АН СССР, 551 с.
Цзин-цзян У., Бе-лу И. 1959. Род Hemiculter и его предварительные экологические исследования в бассейне Хэйлунцзяна // Acta Hydrobiol. Sinica. № 2. Р. 157–169.
Abbott J.F. 1901. List of fishes collected in the River Pei-Ho, at Tien-Tsin, China, by Noah Fields Drake, with descriptions of seven new species // Proc. US Nat. Mus. V. 23. № 1221. P. 483–491.
Bănărescu P. 1968. Revision of the genus Hemiculter (Pisces, Cyprinidae) // Travaux Mus. Natl. Hist. Nat. Grigore Antipa. V. 8. P. 523−529.
Bleeker P. 1860. Conspectus systematis Cyprinorum. Cyprini // Natuurkundig tijdschrift voor Nederlandsch Indië. V. 20. P. 421–439. Available: https://www.biodiversitylibrary.org/page/13420936#page/441/mode/1up.
Bleeker P. 1863–1864. Atlas ichthyologique des Indes Orientales Néêrlandaises, publié sous les auspices du Gouvernement colonial néêrlandais. T. III. Cyprins. V. 3. P. 1–150. Pls. 102–144.
Bleeker P. 1871. Mémoire sur les cyprinoïdes de Chine. Amsterdam: Chez C.G. van der post, 91 pp. Pls. 1–14. Available: https://books.google.ru/books?id=aG5hAAAAcAAJ&printsec=frontcover&hl=ru#v=onepage&q&f=false.
Bogutskaya N.G., Naseka A.M., Shedko S.V. et al. 2008. The fishes of the Amur River: updated check-list and zoogeography // Ichthyol. Explor. Freshw. V. 19. № 4. P. 301–366.
Chen W., Zhong Z., Dai W. et al. 2017. Phylogeographic structure, cryptic speciation and demographic history of the sharpbelly (Hemiculter leucisculus), a freshwater habitat generalist from southern China // BMC Evol. Biol. V. 17. № 1. P. 216. https://doi.org/10.1186/s12862-017-1058-0
Chen Y.-Y. (eds.). 1998. Fauna Sinica. Osteichthyes. Cypriniformes II. Beijing: Sci. Press, 531 p.
Chevey P. 1935. Le lac-cratère de Kouang-Tchéou-Wan (Chine du Sud) description de deux poissons nouveaux // Bull. Soc. Zool. Fr. V. 60. P. 427–433.
Dai Y.-G., Yang J.-X. 2003. Phylogeny and zoogeography of the cyprinid Hemicultrine Group (Cyprinidae: Cultrinae) // Zool. Stud. V. 42. № 1. P. 73−92.
Dybowski B. 1872. Zur Kenntniss der Fischfauna des Amurgebietes // Verh. Zool.-Bot. Ges. Wien. Bd. 22. P. 209–220.
Fricke R., Eschmeyer W.N., van der Laan R. (eds.). 2021. Eschmeyer’s catalog of fishes: genera, species, references (http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Version 03/2021).
Froese R., Pauly D. (eds.). 2021. FishBase. World Wide Web electronic publication. (www.fishbase.org. Version 06/2021).
Fu C., Wu J., Chen J. et al. 2003. Freshwater fish biodiversity in the Yangtze River basin of China: patterns, threats and conservation // Biodiv. Conserv. V. 12. P. 1649−1685. https://doi.org/10.1023/A:1023697714517
He Y. 2010. Structure of endemic fish assemblages in the Upper Yangtze River basin and population differentiation of an endangered endemic fish (Gobiocypris rarus): PhD Thesis. Toulouse: Univ. Toulouse; Wuhan: Inst. d’Hydrobiologie, 65 p.
Jiang R., Zhong J., Zhang D., Fu C. 2008. Species composition and diversity of fish larvae and juveniles in the surf zone of the Yangtze River estuary // Zool. Res. V. 29. № 3. P. 297–304. https://doi.org/10.3724/SP.J.1141.2008.03297
Kim I.-S., Park J.-Y. 2002. Freshwater fishes of Korea. Seoul: Kyo-Hak Publ. Co., 467 p.
Kim M.J., Lee J.-S., Song C.B. 2014. Molecular phylogenetic status of Korean Hemiculter species // Kor. J. Fish. Aquat. Sci. V. 47. № 1. P. 72–78. https://doi.org/10.5657/KFAS.2014.0072
Kottelat M. 2006. Fishes of Mongolia. A check-list of the fishes known to occur in Mongolia with comments on systematics and nomenclature. Washington: World Bank, 103 p.
Kottelat M. 2013. The fishes of the inland waters of Southeast Asia: a catalogue and core bibliography of the fishes known to occur in freshwaters, mangroves and estuaries // Raffles Bull. Zool. Suppl. № 27. P. 1–663.
Li W.J., Gao X., Liu H.Z., Cao W.X. 2020. Coexistence of two closely related cyprinid fishes (Hemiculter bleekeri and Hemiculter leucisculus) in the Upper Yangtze River, China // Diversity. V. 12. № 7. Article 284. https://doi.org/10.3390/d12070284
Luo Y., Chen Y. 1998. Cultrinae // Fauna Sinica. Osteichthyes. Cypriniformes II / Eds. Chen Y.-Y. et al. Beijing: Sci. Press. P. 112−207.
Mori T. 1927. On four new fresh water fishes from the River Liao, south Manchuria // J. Chosen Nat. Hist. Soc. № 5. P. 30–33.
Mori T. 1933. Second addition to the fish fauna of Tsi-nan, China, with descriptions of three new species // Jpn. J. Zool. V. 5. № 2. P. 165−169.
Nei M. 1987. Molecular Evolutionary Genetics. N.Y.: Columbia Univ. Press, 512 p.
Nei M., Jin L. 1989. Variances of the average numbers of nucleotide substitutions within and between populations // Mol. Biol. Evol. V. 6. № 3. P. 240–300. https://doi.org/10.1093/oxfordjournals.molbev.a040547
Nguyen L.-T., Schmidt H.A., von Haeseler A., Minh B.Q. 2015. Q-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies // Mol. Biol. Evol. V. 32. № 1. P. 268–274. https://doi.org/10.1093/molbev/msu300
Nichols J.T. 1925. Some Chinese fresh-water fishes // Amer. Mus. Novit. № 182. P. 1−8.
Nichols J.T. 1943. The fresh-water fishes of China. N. Y.: Amer. Mus. Nat. Hist., 322 p. https://doi.org/10.5962/bhl.title.12103.
Ocock J., Baasanjav G., Baillie J.E.M. (eds.). 2006. Mongolian Red List of fishes. Regional Red List Series. V. 3. London: Zool. Soc. London, 37 p.
Orsi J.J. 1974. A check list of the marine and freshwater fishes of Vietnam // Publ. Seto. Mar. Biol. Lab. V. 21. № 3–4. P. 153–177.
Smith G.R., Dowling T.E., Gobalet K. et al. 2002. Biogeography and rates of evolution of Great Basin fishes // Great Basin Aquatic Systems History. Contributions to the Earth Sciences. № 33. Washington: Smithsonian Inst. P. 175–234.
Tamura K., Nei M. 1993. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees // Mol. Biol. Evol. V. 10. № 1. P. 512–526. https://doi.org/10.1093/oxfordjournals.molbev.a040023
Tan M., Armbruster J.W. 2018. Phylogenetic classification of extant genera of fishes of the order Cypriniformes (Teleostei: Ostariophysi) // Zootaxa. V. 4476. № 1. P. 006–039. https://doi.org/10.11646/zootaxa.4476.1.4.
Wang L., Zhu L., Tang K. et al. 2021. Population genetic structure of sharpbelly Hemiculter leucisculus (Basilesky, 1855) and morphological diversification along climate gradients in China // Ecol. Evol. V. 11. № 11. P. 6798–6813. https://doi.org/10.1002/ece3.7528
Warpachowski N. 1887. Über die Gattung Hemiculter Bleek. und über eine neue Gattung Hemiculterella // Bull. Acad. Impér. Sci. Saint-Petersbourg. V. 32. P. 13–24.
Yang Q., Zhou C., Wang J. 1994. Composition features and geographical division of Cyprinidae fishes of Shandong Province // Zool. Res. V. 15. P. 68–73.
Yih P.-L., Wu C.-K. 1964. Abramidinae // The cyprinid fishes of China. V. 1 / Ed. Wu H.-W. Shanghai: Sci. Press. P. 63–120.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии