

Вопросы ихтиологии, 2022, T. 62, № 4, стр. 450-464
Питание рыб подводных поднятий Китового хребта (Юго-Восточная Атлантика). 2. Низкотелый берикс Beryx splendens (Berycidae)
А. В. Гущин *
Институт океанологии РАН – ИО РАН
Москва, Россия
* E-mail: Poseidon-47@rambler.ru
Поступила в редакцию 15.12.2021
После доработки 03.02.2022
Принята к публикации 09.02.2022
- EDN: OSJEVG
- DOI: 10.31857/S0042875222040130
Аннотация
Исследованы особенности питания низкотелого берикса Beryx splendens подводных поднятий Китового хребта. Подтверждён широкий спектр пищевых организмов берикса, состоящий из мезо- и макропланктона звукорассеивающих слоёв мезопелагиали. Бентосные организмы в пище берикса не обнаружены. Спектр питания берикса различается по блокам подводных поднятий Китового хребта в связи с разницей в составе доступной кормовой фауны и условий среды блоков поднятий, размеров берикса, сезонов и времени лова. Берикс обладает выраженной суточной динамикой питания – интенсивнее всего питается вечером и в начале ночи, когда организмы звукорассеивающих слоёв мигрируют в верхние горизонты воды. В это время отмечена максимальная накормленность и минимальная переваренность пищи. Берикс выбирает для питания организмы размером 0.9–130.0 (в среднем 36.1) мм. По мере увеличения длины тела берикс переходит на питание более крупными жертвами, при этом соотношение размеров жертв и размеров берикса мало изменяется и составляет 9.7–11.5%.
Подводные горы Китового хребта простираются от Африканского материка с северо-востока на юго-запад в виде трёх блоков подводных поднятий, имеющих геологические и океанологические различия и создающих район повышенной биологической продуктивности (Ильин, 1976; Дубравин, 2001, 2013; Ломакин, 2014). Особенностью ряда вершин подводных поднятий является высокая динамическая активность придонных вод, вследствие чего на вершинах отсутствуют рыхлые осадки (Ломакин, 2014), в результате здесь доминируют прикреплённые формы организмов (Федоров, Карамышев, 1991). Видовой состав сообщества рыб Китового хребта хорошо изучен (Пахоруков, 1980, 2003; Пахоруков и др., 2014), но вопросы питания и пищевых взаимоотношений рыб ранее не были затронуты исследованиями за исключением сведений по питанию четырёх видов обитающих здесь рыб (Дудочкин, Котляр, 1989; Гущин, 2021).
Берикс – массовый промысловый вид рыб, встречающийся в Атлантическом океане от зал. Мэн до Патагонского шельфа и от Юго-Западной Европы и Канарских островов до Южной Африки. Обычен на поднятиях подводных хребтов, в том числе на подводных поднятиях Китового хребта Атлантического океана. Представляемая работа по изучению питания берикса служит продолжением серии статей, цель которых – изучить питание массовых видов рыб подводных поднятий Китового хребта. Данное сообщение расширяет представления о питании берикса Китового хребта за счёт большего объёма материала, собранного на пяти подводных поднятиях трёх блоков подводных гор Китового хребта.
МАТЕРИАЛ И МЕТОДИКА
Материал по питанию низкотелого берикса Beryx splendens собран из уловов в рейсах рыболовного траулера морозильного типа Атлантик “Зунд” в октябре 1988 г. и научно-исследовательского судна “Садко” в декабре–январе 1989–1990 гг. Работы проводили на трёх блоках поднятий Китового хребта: Северном – поднятие банка Северная (20°46′ ю.ш. 08°43′ в.д.), Центральном – поднятие банка Щедрая (25°39′ ю.ш. 06°12′ в.д.) и подводная гора Вальдивия (26°07′ ю.ш. 06°20′ в.д.), Южном – банки Бета (31°47′ ю.ш. 02°11′ в.д.) и Альфа-2 (32°50′ ю.ш. 02°35′ в.д.). Большинство тралений выполняли донным тралом “Хек-4М” и пелагическим канатным тралом специального конструкторского бюро промышленного рыболовства по скоплениям рыб, фиксируемых эхолотом. В дневное время (периоды суток по местному времени, ч: утро – 04–09, день – 09–19, вечер – 19–22, ночь – 22–04) траления проводили донным тралом на плоских вершинах подводных поднятий. Пелагический трал использовали в вечернее и ночное время для тралений в пелагиали над поднятиями. Несколько тралений, выполненных донным тралом без показаний эхолота ночью и пелагическим тралом днём, промысловых уловов не дали. Время тралений составляло 25–30 мин. Начало траления отмечали после выхода трала на трассу траления.
У рыб, взятых для биологического анализа методом случайного отбора, определяли абсолютную длину (TL) и массу тела, пол, стадию зрелости половых продуктов по 6-балльной шкале, степень переваренности пищи и ожирения внутренностей по 5-балльной шкале (Инструкция …, 1977). Для камерального анализа питания берикса также методом случайного отбора извлекали желудки и целиком фиксировали их 6%-ным раствором формальдегида.
Содержимое желудков анализировали по стандартной методике (Методическое пособие …, 1974). Общую массу пищевого комка и отдельных компонентов пищи взвешивали с точностью до 10 мг. Все пищевые организмы идентифицировали до возможного таксона, подсчитывали и, если позволяло состояние переваренности, определяли их индивидуальную массу и размеры. Всего проанализировано 390 желудков, из которых 288 содержали пищу.
Для характеристики накормленности рыб применяли индекс наполнения желудков (ИНЖ, ‰), который рассчитывали как отношение общей массы пищи к общей массе рыб в пробе, включая непитающихся рыб. На основании индивидуальных ИНЖ, % рассчитывали средний ИНЖ, % для пробы. Значение отдельных пищевых компонентов оценивали их долей по массе (% общей массы пищевого комка). Частоту их встречаемости (ЧВ, %) определяли как отношение числа нахождений данного компонента пищи к общему числу всех компонентов пищи.
Индекс длины жертвы (ИДЖ, %) рассчитывали как отношение длины жертвы к длине хищника. Степень сходства питания берикса, отловленного на разных поднятиях, определяли с применением индекса Чекановского–Сёренсена (Песенко, 1982): Ics = 2c/(2c + a + b), где a – число компонентов пищи берикса со станций поднятия A; b – число компонентов пищи берикса со станций поднятия B; c – число компонентов пищи, общих для обоих поднятий.
РЕЗУЛЬТАТЫ
Северный блок подводных поднятий
Лов берикса проводили донным тралом в вечернее время по скоплению рыб на дне и пелагическим тралом в ночное и утреннее время по скоплению в пелагиали. Все скопления рыб фиксировали эхолотом. В уловах отмечены берикс (80% по массе), рувета драгоценная Ruvettus pretiosus (15%) и хвостатая рыба-сабля Lepidopus caudatus (5%), единично встречались акулы семейства Etmopteridae. Длина берикса составляла 19–51 см, масса – 130–1350 г. Большинство рыб были неполовозрелыми, среди самок преобладали особи с гонадами II (84.9%) и III (15.1%) стадий зрелости, самцов – II (77.4%) и III (22.6%) стадий.
Максимальное значение ИНЖ берикса (98‱) отмечено в вечернее время в придонном слое на вершине поднятия. В пелагиали его накормленность снижалась, и к утру ИНЖ составлял 30‱. Степень переваренности пищи берикса была 2.4–2.6 балла, ожирение внутренностей – 1.7–1.9 балла (табл. 1).
Таблица 1.
Питание берикса Beryx splendens на подводном поднятии Северного блока (банка Северная)
| Компонент пищи и другие показатели |
Дата (время начала и глубина траления) | Всего | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 13.01.1990 г. (19:20, 215 м)* |
14.01.1990 г. (22:50, 170 м) |
15.01.1990 г. (02:50, 190 м) |
15.01.1990 г. (04:30, 200 м) |
|||||||
| М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | |
| Teuthida | 2.0 | 20.8 | 4.3 | 2.0 | 2.0 | 7.7 | 2.5 | 2.5 | ||
| Кальмары (всего) | 2.0 | 4.3 | 2.0 | 2.5 | ||||||
| Crustacea | 0.4 | 4.0 | 4.7 | 26.9 | 22.3 | 55.0 | 2.3 | 18.9 | ||
| Copepoda | 0.1 | 4.0 | + | 1.1 | ||||||
| Decapoda | 7.1 | 28.0 | 2.0 | 7.4 | ||||||
| Sergestidae | 0.4 | 4.2 | 1.5 | 4.0 | 4.6 | 3.8 | 1.3 | 3.2 | ||
| Systellaspis sp. | 0.3 | 4.0 | 0.1 | 1.1 | ||||||
| Acanthephyra acanthitelsonis | 16.8 | 25.0 | 8.5 | 6.3 | ||||||
| Funchalia woodwardi | 2.0 | 4.2 | 3.1 | 8.0 | 10.1 | 3.8 | 3.3 | 4.2 | ||
| Oplophorus novaezeelandiae | 11.3 | 41.7 | 18.6 | 40.0 | 29.9 | 38.5 | 15.2 | 31.6 | ||
| Robustosergia robusta | 0.9 | 4.0 | 0.2 | 1.1 | ||||||
| Plesionika richardi | 1.5 | 4.0 | 0.4 | 1.1 | ||||||
| Hyperiidae | 0.4 | 5.0 | + | 1.1 | ||||||
| Phronima sp. | + | 4.0 | + | 1.1 | ||||||
| Vibilia sp. | 0.1 | 8.3 | 0.5 | 8.0 | 0.4 | 15.4 | 0.2 | 8.4 | ||
| Platyscelus ovoides | 0.2 | 8.3 | 0.5 | 8.0 | 1.6 | 7.7 | 16.2 | 35.0 | 1.7 | 13.7 |
| Ракообразные (всего) | 30.8 | 34.5 | 51.3 | 38.9 | 35.2 | |||||
| Appendicularia | 0.9 | 3.8 | 0.1 | 1.1 | ||||||
| Pyrosomatidae | 0.4 | 4.2 | 0.2 | 1.1 | ||||||
| Salpa sp. | 4.9 | 16.0 | 10.4 | 19.2 | 33.5 | 40.0 | 5.4 | 17.9 | ||
| Туникаты (всего) | 0.4 | 4.9 | 11.3 | 33.5 | 5.7 | |||||
| Osteichthyes | 12.7 | 29.2 | 30.0 | 12.0 | 11.0 | 7.7 | 4.4 | 10.0 | 16.4 | 14.7 |
| Bathylagidae | 1.6 | 4.2 | 0.8 | 1.1 | ||||||
| Myctophidae | 2.8 | 4.2 | 6.3 | 8.0 | 21.7 | 7.7 | 23.2 | 5.0 | 8.1 | 6.3 |
| Diaphus sp. | 5.0 | 8.3 | 2.5 | 2.1 | ||||||
| Lampadena sp. | 7.0 | 4.0 | 1.9 | 1.1 | ||||||
| Stomiidae | 2.4 | 3.8 | 0.4 | 1.1 | ||||||
| Melanostomias sp. | 1.0 | 4.2 | 0.5 | 1.1 | ||||||
| Chauliodus sp. | 6.8 | 4.0 | 1.9 | 1.1 | ||||||
| Epigonus sp. | 12.6 | 12.5 | 6.3 | 3.2 | ||||||
| Nansenia sp. | 2.6 | 4.0 | 0.7 | 1.1 | ||||||
| Paralepis sp. | 3.4 | 4.0 | 0.9 | 1.1 | ||||||
| Maurolicus muelleri | 1.6 | 8.3 | 0.8 | 2.1 | ||||||
| Chlorophthalmus agassizi | 2.5 | 4.2 | 1.3 | 1.1 | ||||||
| Tetragonurus cuvieri | 27.0 | 8.3 | 13.6 | 2.1 | ||||||
| Рыбы (всего) | 66.8 | 56.1 | 35.1 | 27.6 | 56.1 | |||||
| Переваренная пища | 0.2 | 4.0 | 0.3 | 7.7 | 0.5 | 2.1 | ||||
| Число желудков общее/с пищей | 25/24 | 25/25 | 30/26 | 27/20 | 107/95 | |||||
| Ожирение внутренностей, баллы | $\frac{{1.9 \pm 0.2}}{{0.9}}$ | $\frac{{1.7 \pm 0.1}}{{0.7}}$ | $\frac{{1.9 \pm 0.1}}{{0.7}}$ | $\frac{{1.9 \pm 0.2}}{{0.9}}$ | $\frac{{1.9 \pm 0.1}}{{0.8}}$ | |||||
| Переваренность пищи, баллы | $\frac{{2.6 \pm 0.1}}{{0.5}}$ | $\frac{{2.4 \pm 0.1}}{{0.6}}$ | $\frac{{2.5 \pm 0.1}}{{0.6}}$ | $\frac{{2.4 \pm 0.1}}{{0.6}}$ | $\frac{{2.5 \pm 0.1}}{{0.6}}$ | |||||
| Индекс наполнения желудков, ‱ | $\frac{{98.2 \pm 13.0}}{{63.6}}$ | $\frac{{68.7 \pm 14.2}}{{71.0}}$ | $\frac{{38.6 \pm 9.3}}{{47.5}}$ | $\frac{{30.5 \pm 7.6}}{{33.9}}$ | $\frac{{61.5 \pm 6.3}}{{61.0}}$ | |||||
Примечание. Здесь и в табл. 2, 3: М – доля компонента по массе, %, ЧВ – частота встречаемости компонента, %; * донный трал, в остальных случаях – пелагический. Здесь и в табл. 2–4: над чертой среднее значение и его ошибка, под чертой – стандартное отклонение; “+” – доля компонента менее 0.1%.
Основу питания берикса составляли рыбы (56.1% массы пищи). Доля рыб в пище постепенно снижалась с 66.8% вечером до 27.6% в утренние часы. Среди рыб в питании берикса преобладали миктофиды, в том числе Diaphus sp. и Lampadena sp. Наибольшее число видов рыб в пище берикса отмечено в вечернее время, когда в желудках встречались представители семейств Bathylagidae и Myctophidae, Epigonus sp., Melanostomias sp., Chlorophthalmus agassizi, Maurolicus muelleri и Tetragonurus cuvieri; к утру число видов рыб в пище берикса снижалось.
Значение ракообразных в пище увеличивалось с вечера и достигало максимума во второй половине ночи (51.3% по массе), когда ракообразные становились главной пищей берикса, и снова снижалось до 38.9% в утренние часы. Креветки Oplophorus novaezeelandiae были доминирующим (29%) видом среди ракообразных в пище берикса во второй половине ночи, однако в утреннее время в питании отсутствовали. Креветки Acanthephyra acanthitelsonis (16.8%) встречалась в пище только в вечернее время. Доля креветок Funchalia woodwardi в пище берикса возрастала с вечера и достигала максимума (10.1%) во второй половине ночи. В утренние часы креветки F. woodwardi в пище берикса отсутствовали. Масса и частота встречаемости остальных ракообразных были небольшими. Креветка Robustosergia robusta встречалась в желудках в начале ночи, креветки семейства Sergestidae – в вечернее и ночное время. Ночью в желудках берикса в небольшом количестве встречались креветки рода Systellaspis sp. и Plesionika richardi. Гиперииды были второстепенным объектом пищи берикса по массе. Доля гипериид Platyscelus ovoides увеличивалась с вечера и достигала максимума (16.2%) в утренние часы. Некоторые особи P. ovoides находились внутри сальп. Vibilia sp. встречались в желудках в вечерние и ночные часы. Туникаты в желудках берикса были представлены Appendicularia, Pyrosomatinae и Salpa sp. Доля по массе и частота встречаемости сальп в пище берикса увеличивались в ночное время и достигали максимума (соответственно 33.5 и 40.0%) в утренние часы. Другие туникаты (Appendicularia и Pyrosomatinae) встречались в пище единично. Кальмаров фиксировали в желудках берикса в вечернее и ночное время, в утренние часы они отсутствовали, их доля была невелика и составляла 2.5% (табл. 1).
Центральный блок подводных поднятий
Пробы для изучения питания берикса были взяты из уловов донного трала на подводном поднятии банка Щедрая и на подводной горе Вальдивия. На банке Щедрая пробы взяты поздним вечером и ночью. Траление проводили по дну по скоплениям, фиксируемым эхолотом в виде “щёточки”. На Вальдивии траления были выполнены в конце дня по пелагическим скоплениям, фиксируемым эхолотом в виде “облачка”. Ночью провели траление по дну (с низкой результативностью) под наблюдаемыми в пелагиали скоплениями рыбы. В утренние часы траление проводили по скоплению на дне, фиксируемому эхолотом в виде “щёточки”. Уловы состояли из кабан-рыбы Pentaceros richardsoni – 80% (по массе), берикса – 5%, беспузырного окуня Helicolenus mouchezi – 12%, красноглазки Emmelichthys nitidus nitidus – 3%. Единично встречались рувета, рыба-сабля и акулы семейства Etmopteridae. Берикс в уловах был длиной 19–56 см и массой 60–1740 г. На подводном поднятии Вальдивия самки берикса имели половые продукты II (83.3%) и III (16.7%) стадий зрелости, самцы – II (70.0%) и III (30.0%) стадий. На подводном поднятии банка Щедрая половые продукты самок были II (2.8%) и III (97.2%) стадий зрелости, самцов – III (33.3%) и IV (66.7%) стадий.
В водах подводного поднятия Вальдивия средний ИНЖ берикса составил 23.1‱. Максимальный ИНЖ (39.5‱) отмечен в конце дня при средней переваренности пищи 2.1 балла, минимальный (7.1‱) – в утренние часы при средней переваренности 2.9 балла. На поднятии банка Щедрая средний ИНЖ берикса был 28.9‱, максимальный (37.0‱) отмечен в конце вечера при средней переваренности пищи 2.5 балла. Главной пищей берикса Центрального блока поднятий были ракообразные (38.2% по массе), второстепенной – рыбы (33.0%), кальмары (11.8%) и туникаты (11.6%) (табл. 2). В водах поднятия Вальдивия главной пищей берикса были рыбы (29.4%), среди которых преобладал M. muelleri (26.5%), он зарегистрирован в желудках в конце дня с ЧВ 15.7%, остальная рыба была сильно переварена.
Таблица 2.
Питание берикса Beryx splendens на поднятиях Центрального блока (горы Щедрая, Вальдивия)
| Компонент пищи и другие показатели |
Щедрая | Вальдивия | Всего | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Дата (время начала и глубина траления) |
Итого | Дата (время начала и глубина траления) |
Итого | |||||||||||||
| 26.12.89 г. (21:15, 400 м)* |
29.12.89 г. (01:15, 390 м)* |
30.12.89 г. (18:30, 230 м)* |
31.12.89 г. (23:15, 230 м)* |
01.01.90 г. (07:15, 220 м)* |
||||||||||||
| М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | |
| Teuthida | 0.3 | 27.3 | 23.2 | 30.8 | 8.5 | 19.4 | 5.0 | 6.3 | 34.9 | 33.3 | 32.3 | 25.0 | 22.0 | 19.6 | 11.8 | 24.3 |
| Кальмары (всего) | 0.3 | 23.2 | 8.5 | 5.0 | 34.9 | 32.3 | 22.0 | 11.8 | ||||||||
| Crustacea | 3.0 | 9.1 | 7.7 | 23.1 | 4.6 | 11.1 | 3.7 | 18.8 | 41.8 | 44.4 | 33.8 | 41.7 | 24.5 | 31.4 | 10.0 | 28.6 |
| Decapoda | 18.9 | 36.4 | 5.1 | 7.7 | 14.3 | 13.9 | 6.0 | 11.1 | 2.4 | 3.9 | 11.0 | 10.0 | ||||
| Oplophorus novaezeelandiae | 31.6 | 27.3 | 22.5 | 8.3 | 15.2 | 4.3 | ||||||||||
| Euphausiidae | 5.4 | 23.1 | 1.8 | 8.3 | 1.3 | 4.3 | ||||||||||
| Hyperiidae | 0.6 | 6.3 | 0.2 | 2.0 | 0.1 | 1.4 | ||||||||||
| Phronima sp. | 0.2 | 9.1 | 0.1 | 2.8 | + | 1.4 | ||||||||||
| Platyscelus ovoides | 11.3 | 8.3 | 2.0 | 2.0 | 0.6 | 1.4 | ||||||||||
| Ракообразные (всего) | 53.7 | 18.2 | 43.3 | 4.3 | 47.8 | 45.1 | 29.1 | 38.2 | ||||||||
| Tunicata | 1.8 | 9.1 | 4.6 | 7.7 | 2.7 | 5.6 | 2.0 | 2.9 | ||||||||
| Salpa sp. | 2.1 | 18.2 | 1.4 | 15.6 | 26.9 | 31.3 | 17.2 | 22.2 | 18.1 | 19.6 | 6.0 | 15.7 | ||||
| Pyrosoma atlanticum | 7.4 | 9.1 | 4.9 | 2.8 | 3.6 | 1.4 | ||||||||||
| Туникаты (всего) | 11.3 | 4.6 | 9.0 | 26.9 | 17.2 | 18.1 | 11.6 | |||||||||
| Osteichthyes | 15.9 | 16.7 | 2.9 | 3.9 | 0.8 | 2.9 | ||||||||||
| Myctophidae | 3.7 | 9.1 | 2.5 | 2.8 | 1.8 | 1.4 | ||||||||||
| Diaphus sp. | 31.0 | 9.1 | 33.6 | 7.7 | 32.8 | 5.6 | 23.1 | 2.9 | ||||||||
| Maurolicus muelleri | 63.8 | 50.0 | 26.5 | 15.7 | 7.3 | 11.4 | ||||||||||
| Рыбы (всего) | 34.7 | 33.6 | 35.3 | 63.8 | 15.9 | 29.4 | 33.0 | |||||||||
| Переваренная пища | + | 9.1 | 20.4 | 30.8 | 3.9 | 5.8 | 0.1 | 11.1 | 6.7 | 8.3 | 1.4 | 2.0 | 5.4 | 1.4 | ||
| Число желудков общее/с пищей | 24/11 | 29/13 | 53/24 | 25/16 | 25/18 | 25/12 | 75/30 | 128/70 | ||||||||
| Ожирение внутренностей, баллы | $\frac{{1.2 \pm 0.1}}{{0.6}}$ | $\frac{{2.1 \pm 0.1}}{{0.4}}$ | $\frac{{2.8 \pm 0.1}}{{0.5}}$ | $\frac{{1.6 \pm 0.1}}{{0.6}}$ | $\frac{{1.5 \pm 0.1}}{{0.7}}$ | $\frac{{2.7 \pm 0.1}}{{0.5}}$ | $\frac{{1.7 \pm 0.1}}{{1.1}}$ | $\frac{{1.5 \pm 0.1}}{{0.8}}$ | ||||||||
| Переваренность пищи, баллы | $\frac{{2.5 \pm 0.2}}{{0.7}}$ | $\frac{{3.0 \pm 0.2}}{{0.8}}$ | $\frac{{2.9 \pm 0.2}}{{0.7}}$ | $\frac{{2.1 \pm 0.2}}{{0.7}}$ | $\frac{{2.9 \pm 0.1}}{{1.3}}$ | $\frac{{2.9 \pm 0.3}}{{1.1}}$ | $\frac{{2.7 \pm 0.1}}{{0.6}}$ | $\frac{{2.7 \pm 0.1}}{{0.7}}$ | ||||||||
| Индекс наполнения желудков, ‱ | $\frac{{37.0 \pm 18.16}}{{0.2}}$ | $\frac{{20.9 \pm 7.12}}{{5.6}}$ | $\frac{{28.3 \pm 9.1}}{{44.5}}$ | $\frac{{39.5 \pm 5.7}}{{5.5}}$ | $\frac{{16.5 \pm 3.8}}{{16.3}}$ | $\frac{{7.1 \pm 1.6}}{{5.7}}$ | $\frac{{23.1 \pm 5.8}}{{38.7}}$ | $\frac{{15.5 \pm 3.4}}{{35.1}}$ | ||||||||
Доля ракообразных (29.1%) была соизмерима с долей рыб. Все ракообразные в пище были представлены сильно переваренными фрагментами и становились главной пищей берикса в ночное время (47.8%) и в утренние часы (45.1%). Утром в пище встречались гиперииды P. ovoides (11.3%) с ЧВ 8.3%. Переваренность большинства особей P. ovoides составляла 1.1 балла. Кальмары (22.0%) в желудках берикса были сильно переварены. Туникаты, представленные в пище берикса сальпами (18.1%) с ЧВ 19.6%, встречались в конце дня (26.9%) и в ночное время (17.2%) (табл. 2). В водах банки Щедрая главной пищей берикса были ракообразные (43.2%), их доля в вечерние часы увеличивалась до 53.7% за счёт креветки O. novaezeelandiae (31.6%) с ЧВ 27.3%. В ночные часы главной пищей берикса становились рыбы (33.6%) и прежде всего миктофиды, среди которых доминировал Diaphus sp. Доля Diaphus sp. вечером составляла 31.0%, ночью – 33.6%; ЧВ соответственно – 9.6 и 7.7%. В ночное время в пище берикса возрастала доля кальмаров (23.2%), а ракообразных – снижалась до 18.2%. Туникаты в вечернее время в пище берикса были представлены фрагментами пиросомы Pyrosoma atlanticum (7.4%) и сальпами (2.1%) с ЧВ соответственно 9.1 и 18.2%. Ночью доля туникат составляла 4.6%.
Южный блок подводных поднятий
На подводном поднятии Альфа-2 пробы были взяты из уловов донным тралом вечером и пелагическим тралом ночью. Траления выполняли по скоплениям рыбы, фиксируемым эхолотом. На подводном поднятии Бета пробы взяты из уловов пелагическим тралом в конце дня и в вечерние часы. Уловы состояли из кабан-рыбы – 45% (по массе), берикса – 25%, красноглазки – 18%, беспузырного окуня – 10%, руветы – 2%. Единично встречались рыба-сабля и акулы семейства Etmopteridae. Берикс в уловах был длиной 25–50 см и массой 230–1650 г. В водах подводного поднятия Альфа-2 самки берикса имели половые продукты II (60%), III (38%) и IV (2%) стадий зрелости, самцы – II (65%), III (30%) и IV (5%) стадий. На поднятии Бета самки берикса имели гонады II (53%), III (46%) и IV (1%) стадий зрелости, самцы – II (52%), III (46%) и IV (2%) стадий. Средний ИНЖ берикса Южного блока составил 80.3‱. На подводном поднятии Альфа-2 максимальный ИНЖ (106.6‱) был отмечен в октябре 1988 г. В январе 1990 г. максимальный ИНЖ (70.0%) отмечен у берикса, взятого из улова пелагическим тралом в ночное время. Через сутки, в ночное время, ИНЖ составил 31.3‱. Вечером накормленность берикса снизилась до 49.4‱. На подводном поднятии Бета максимальный ИНЖ (84.2‱) был у берикса, пойманного вечером пелагическим тралом, степень переваренности пищи составила 2.3 балла. Минимальная накормленность (ИНЖ 7.8‱) отмечена в дневное время у берикса, пойманного пелагическим тралом у дна подводного поднятия. Степень переваренности пищи была 2.9 балла (табл. 3).
Таблица 3.
Питание берикса Beryx splendens на поднятиях Южного блока (горы Бета, Альфа-2)
| Компонент пищи и другие показатели |
Бета | Альфа-2 | Всего | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Дата (время начала и глубина траления) |
Итого | Дата (время начала и глубина траления) |
Итого | |||||||||||||||
| 01.01.1990 г. (16:30, 510 м) |
01.01.1990 г. (21:10, 630 м) |
27.10.88 г. (21:45, 490 м)* |
3.01.90 г. (03:15, 150 м) |
03.01.90 г. (21:00, 600 м)* |
04.01.90 г. (01:50, 570 м) |
|||||||||||||
| М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | М | ЧВ | |
| Teuthida | 33.5 | 27.3 | 1.3 | 15.0 | 2.1 | 10.9 | 0.3 | 15.4 | 6.1 | 7.1 | 8.8 | 41.2 | 7.9 | 64.0 | 2.8 | 11.5 | 6.0 | 28.9 |
| Oegopsida | 6.9 | 3.6 | 3.5 | 4.0 | 3.1 | 1.1 | 3.2 | 1.8 | ||||||||||
| Abraliopsis sp. | 0.5 | 5.0 | 0.5 | 1.8 | 0.1 | 3.6 | + | 4.0 | 4.3 | 14.9 | 0.1 | 2.6 | ||||||
| Кальмары (всего) | 33.5 | 1.8 | 2.6 | 0.3 | 13.1 | 8.8 | 11.4 | 10.2 | 9.3 | |||||||||
| Crustacea | 34.5 | 45.5 | 0.2 | 5.0 | 1.2 | 10.9 | 0.7 | 7.7 | 9.7 | 32.1 | 0.3 | 5.9 | 6.2 | 80.0 | 3.4 | 9.2 | 5.2 | 32.5 |
| Copepoda | 0.5 | 2.6 | 2.2 | 10.9 | 0.1 | 30.8 | + | 40.0 | 0.1 | 5.7 | + | 3.5 | ||||||
| Decapoda | 15.5 | 15.0 | 15.0 | 5.5 | + | 15.4 | 1.0 | 17.9 | 0.5 | 28.0 | 0.4 | 8.0 | 2.6 | 14.9 | ||||
| Sergestidae | 0.5 | 7.7 | 0.1 | + | 0.1 | 2.3 | 0.1 | 0.9 | ||||||||||
| Pasiphaea sp. | 76.3 | 92.3 | 16.6 | 52.0 | 14.5 | 13.8 | 15.1 | 21.9 | ||||||||||
| Gennadas sp. | + | 4.0 | + | + | + | 0.9 | ||||||||||||
| Funchalia woodwardi | 43.9 | 60.0 | 40.6 | 21.8 | 45.2 | 32.1 | 6.1 | 5.9 | 23.7 | 40.0 | 20.4 | 10.3 | 27.9 | 28.1 | ||||
| Oplophorus novaezeelandiae | 0.8 | 7.1 | 0.4 | 12.0 | 1.1 | 3.4 | 0.4 | 4.4 | ||||||||||
| Euphausiidae | 0.3 | 5.0 | 0.3 | 1.8 | 0.2 | 15.4 | + | 8.0 | 0.5 | 9.2 | 0.1 | 4.4 | ||||||
| Thysanoessa sp. | 0.4 | 38.5 | 1.1 | 8.0 | 1.0 | 8.0 | 0.5 | 6.1 | ||||||||||
| Eucopia sp. | 0.5 | 5.0 | 0.5 | 1.8 | 0.1 | 9.6 | ||||||||||||
| Mysida | 7.5 | 11.8 | 0.1 | 20.0 | + | 6.7 | 0.2 | 6.1 | ||||||||||
| Hyperiidae | 0.1 | 5.0 | 0.1 | 1.8 | + | 15.4 | 0.1 | 5.9 | 0.5 | 28.0 | + | 3.4 | 0.2 | 9.6 | ||||
| Vibilia sp. | + | 5.0 | + | 1.8 | 0.3 | 3.6 | 2.4 | 47.1 | 0.6 | 60.0 | 0.6 | 6.9 | 0.5 | 21.9 | ||||
| Platyscelus armatus | 0.2 | 5.0 | 0.2 | 1.8 | + | 0.9 | ||||||||||||
| Platyscelus ovoides | 11.5 | 9.1 | 0.3 | 1.8 | + | 3.6 | 0.5 | 28.0 | 0.4 | 17.2 | 0.3 | 8.8 | ||||||
| Cyllopus magellanicus | + | 3.6 | + | 11.8 | 0.4 | 12.0 | 0.3 | 3.4 | 0.2 | 2.6 | ||||||||
| Ostracoda | + | 5.0 | + | 1.8 | + | 4.0 | + | 0.3 | + | 1.8 | ||||||||
| Isopoda | + | 15.4 | + | 8.0 | 5.4 | 24.1 | + | 3.5 | ||||||||||
| Ракообразные (всего) | 46.5 | 60.7 | 60.4 | 78.2 | 57.0 | 16.4 | 50.7 | 48.2 | 53.4 | |||||||||
| Tunicata | 2.4 | 2.9 | 10.0 | 2.8 | 3.6 | 19.7 | 84.6 | 8.2 | 14.3 | 8.5 | 64.0 | 8.6 | 14.9 | 8.2 | 28.9 | |||
| Appendicularia | 0.2 | 4.0 | 3.7 | 11.5 | 0.1 | 0.9 | ||||||||||||
| Salpa sp. | 6.9 | 35.0 | 5.7 | 12.7 | 24.2 | 52.9 | 8.5 | 100.0 | 3.9 | 5.7 | 6.7 | 36.8 | ||||||
| Pyrosoma atlanticum | 12.8 | 5.0 | 12.4 | 1.8 | 1.1 | 3.6 | 24.2 | 35.3 | 4.1 | 28.0 | 4.6 | 18.4 | 5.3 | 12.3 | ||||
| Туникаты (всего) | 2.4 | 22.6 | 20.9 | 19.7 | 9.3 | 48.4 | 21.3 | 20.8 | 20.3 | |||||||||
| Osteichthyes | 14.2 | 18.2 | 0.4 | 5.0 | 0.8 | 5.5 | 0.9 | 15.4 | 7.1 | 7.1 | 5.2 | 24.0 | 5.0 | 26.4 | 4.2 | 11.4 | ||
| Myctophidae | 12.6 | 15.0 | 12.3 | 5.5 | 0.4 | 7.7 | 8.1 | 7.1 | 26.2 | 11.8 | 8.4 | 20.0 | 8.3 | 8.0 | 9.1 | 11.4 | ||
| Diaphus dumerilii | 5.1 | 3.6 | 2.6 | 4.0 | 7.1 | 5.7 | 2.4 | 1.8 | ||||||||||
| Vinciguerria sp. | 0.7 | 5.0 | 0.7 | 1.8 | 0.1 | 0.9 | ||||||||||||
| Maurolicus muelleri | 1.1 | 5.0 | 1.1 | 1.8 | 0.2 | 0.9 | ||||||||||||
| Рыбы (всего) | 14.2 | 14.8 | 14.9 | 1.3 | 20.3 | 26.2 | 16.2 | 20.4 | 16.0 | |||||||||
| Икра | 0.1 | 7.7 | + | 4.0 | + | 2.3 | + | 1.8 | ||||||||||
| Переваренная пища | 3.4 | 9.1 | 0.1 | 5 | 1.2 | 3.6 | 0.4 | 7.7 | 0.3 | 10.7 | 0.2 | 0.4 | 20 | 0.4 | 5.7 | 1.0 | 9.6 | |
| Число желудков общее/с пищей | 20/11 | 25/20 | 45/31 | 29/13 | 31/28 | 20/17 | 26/25 | 106/83 | 151/114 | |||||||||
| Ожирение внутренностей, баллы | $\frac{{2.8 \pm 0.1}}{{0.5}}$ | $\frac{{2.6 \pm 0.2}}{{0.9}}$ | $\frac{{2.8 \pm 0.3}}{{0.8}}$ | – | $\frac{{2.1 \pm 0.2}}{{0.9}}$ | $\frac{{2.1 \pm 0.1}}{{0.6}}$ | $\frac{{2.6 \pm 0.1}}{{0.5}}$ | $\frac{{2.4 \pm 0.1}}{{0.6}}$ | $\frac{{2.4 \pm 0.2}}{{0.7}}$ | |||||||||
| Переваренность пищи, баллы | $\frac{{2.9 \pm 0.2}}{{0.7}}$ | $\frac{{2.3 \pm 0.1}}{{0.4}}$ | $\frac{{2.5 \pm 0.3}}{{0.5}}$ | $\frac{{2.4 \pm 0.1}}{{0.7}}$ | $\frac{{2.4 \pm 0.2}}{{0.8}}$ | $\frac{{2.3 \pm 0.1}}{{0.5}}$ | $\frac{{2.8 \pm 0.1}}{{0.7}}$ | $\frac{{2.5 \pm 0.2}}{{0.6}}$ | $\frac{{2.5 \pm 0.2}}{{0.6}}$ | |||||||||
| Индекс наполнения желудков, ‱ | $\frac{{7.8 \pm 5.7}}{{5.5}}$ | $\frac{{84.2 \pm 15.1}}{{67.5}}$ | $\frac{{43.2 \pm 18.1}}{{60.3}}$ | $\frac{{106.6 \pm 17.0}}{{170.5}}$ | $\frac{{70.0 \pm 13.0}}{{68.7}}$ | $\frac{{49.4 \pm 11.0}}{{45.4}}$ | $\frac{{31.3 \pm 4.0}}{{19.9}}$ | $\frac{{64.3 \pm 13.5.2}}{{53.2}}$ | $\frac{{80.3 \pm 11.6}}{{145.4}}$ | |||||||||
Главной пищей берикса Южного блока поднятий были ракообразные (53.9%). Второстепенной пищей – туникаты (20.3%), рыбы (16.0%) и кальмары (9.3%) (табл. 3). В пище берикса на поднятии Альфа-2 основу питания составляли ракообразные (48.2%), среди которых преобладали креветка F. woodwardi (20.4%) с ЧВ 10.3% и креветки рода Pasiphaea (14.5%). Доля и частота встречаемости изопод составляли 5.4 и 24.1%. Доля других креветок, мизид, эвфаузиид в пище берикса суммарно не превышала 3.2%. Гиперииды видов P. armatus, P. ovoides, Cyllopus magellanicus и Vibilia sp. суммарно составляли 1.3% по массе. Туникаты (20.8%) в пище берикса включали пиросом P. atlanticum (4.6%), сальп и аппендикулярией. Туникаты были найдены в желудках рыб, пойманных в вечернее и ночное время, и были главной пищей (48.4%) берикса в пробе из донного траления вечером.
Среди рыбы (20.4% по массе) в желудках берикса преобладали неидентифицированные до вида миктофиды (8.3%) с ЧВ 8.0% и Diaphus dumerilii (7.1%) с ЧВ 5.7%. Кальмары (10.2%) в пище берикса встречались в вечернее и ночное время. Большинство кальмаров было сильно переварено, они были представлены клювами, крючьями и фрагментами мантии, что затрудняло их определение (табл. 3). Главной пищей берикса на поднятии Бета были ракообразные (60.4%), в основном декаподы, среди которых креветка F. woodwardi (40.6%) с ЧВ 21.8% занимала ведущее место. Второстепенной пищей были туникаты (20.9%) с ведущим видом P. atlanticum (12.4%) с ЧВ 1.8%. Сальпы (5.7% по массе) доминировали по частоте встречаемости (12.7%). Доля рыб составляла 14.9%, среди них преобладали миктофиды (12.3%). Доля Vinciguerria sp. и M. muelleri была невелика. Кальмары встречались в небольшом количестве (2.6%), но днём в пробе желудков берикса из улова пелагического траления у дна вершины подводного поднятия их доля составила 33.5% М.
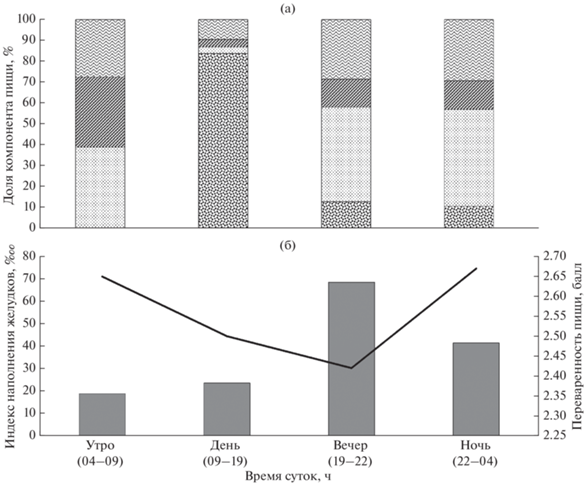
Питание берикса в различные периоды суток различалось по накормленности, составу пищевых организмов и переваренности пищи. Имеющийся материал позволил объединить данные по периодам суток (рис. 1). Накормленность берикса в утренний период была минимальна (ИНЖ 18.8‱) при средней степени переваренности 2.65 балла. Пища берикса в этот период состояла из ракообразных (38.9%), туникат (33.5%) и рыб (27.5%). В дневное время накормленность берикса немного возрастала (ИНЖ 23.6‱), степень переваренности пищи снижалась до 2.50 балла. В этот период в пище преобладали кальмары (83.7%), рыбы (9.5%), туникаты (3.6%) и ракообразные (3.2%) служили второстепенной пищей. Вечерний период характеризовался максимальной накормленностью (ИНЖ 68.5‱) берикса при минимальной средней степени переваренности пищи 2.42 балла. В этот период в пище доминировали ракообразные (45.2%), рыбы (28.5%), туникаты (13.4%) и кальмары (12.9%) являлись второстепенной пищей. В ночной период накормленность (ИНЖ 41.5‱) немного снижалась, но возрастала средняя степень переваренности пищи до 2.67 балла. Состав пищи берикса был сходен с таковым в вечернее время и представлен ракообразными (46.4%), рыбой (29.3%), туникатами (13.6%) и кальмарами (10.7%).
Рис. 1.
Суточная динамика питания берикса Beryx splendens на подводных поднятиях Китового хребта: а – спектры питания (доля компонента пищи
по массе): ( ) – кальмары, (
) – кальмары, ( ) – ракообразные, (
) – ракообразные, ( ) – туникаты, (
) – туникаты, ( ) – рыбы; б – индексы наполнения желудков (
) – рыбы; б – индексы наполнения желудков ( ) и среднее значение переваренности пищи (─).
) и среднее значение переваренности пищи (─).
Состав пищи берикса по размерным группам. Видовой и размерный состав пищи берикса разных размерных групп различался. Ракообразные (88.1% по массе) служили главной пищей мелкого берикса TL 190–249 мм. Среди ракообразных ведущим видом была креветка Pasiphaea sp. (66.3%). Доля креветок O. novaezeelandiae и F. woodwardi невысока – 3%. Гиперииды, представленные P. ovoides, C. magellanicus, P. armatus и Vibilia sp., в пище берикса суммарно составляли 10.0%. Среди туникат (6.1%) в пище преобладали неидентифицированные фрагменты оболочников (4.8%) и целые сальпы (1.3%). Из рыб (2.6%) в желудках берикса встречались Nansenia sp. (1.9%) и M. muelleri (0.7%). Доля кальмаров была невелика (1.3%). Накормленность берикса этой размерной группы была максимальной (ИНЖ 141.0‱) по сравнению с другими: 84% особей берикса питались. Средняя переваренность пищи была минимальной (2.1 балла) по сравнению с другими размерными группами (табл. 4).
Таблица 4.
Состав пищи (% по массе) различных размерных групп берикса Beryx splendens Китового хребта в 1989–1990 гг.
| Компонент пищи и другие показатели |
Размерная группа, мм | ||||||
|---|---|---|---|---|---|---|---|
| 190–249 | 250–299 | 300–349 | 350–399 | 400–449 | 450–4990 | 500–559 | |
| Oegopsida | 8.2 | ||||||
| Abraliopsis atlantica | 0.1 | 0.3 | |||||
| Teuthida | 1.3 | 2.7 | 3.5 | 4.0 | 23.0 | 2.6 | 4.3 |
| Кальмары (всего) | 1.3 | 2.7 | 3.5 | 4.1 | 23.3 | 10.7 | 4.3 |
| Copepoda | 0.1 | + | 0.4 | ||||
| Acanthephyra acanthitelsonis | 4.4 | 4.3 | |||||
| Systellaspis sp. | 0.4 | ||||||
| Oplophorus novaezeelandiae | 1.9 | 23.2 | 10.2 | 7.1 | 5.5 | ||
| Funchalia woodwardi | 1.1 | 8.8 | 13.7 | 6.9 | 47.4 | 67.2 | |
| Pasiphaea sp. | 66.3 | 4.6 | |||||
| Sergestes sp. | 4.0 | 0.3 | |||||
| Plesionika richardi | 0.5 | ||||||
| Gennadas sp. | + | ||||||
| Decapoda | 2.5 | 3.2 | 2.4 | 1.8 | 3.1 | 11.7 | |
| Thysanoessa sp. | 0.4 | ||||||
| Eucopia sp. | 0.3 | ||||||
| Mysida | 0.5 | 0.2 | |||||
| Euphausiidae | 0.8 | + | 0.4 | 0.1 | |||
| Platyscelus armatus | 0.2 | 0.1 | |||||
| Platyscelus ovoides | 5.4 | 1.7 | 0.8 | + | |||
| Vibilia sp. | 0.5 | 0.2 | 2.4 | 0.2 | 0.1 | ||
| Cyllopus magellanicus | 1.9 | ||||||
| Hyperiidae | 2.0 | 0.3 | 0.2 | 0.1 | |||
| Crustacea | 6.2 | 10.5 | 8.2 | 2.2 | 4.7 | 3.9 | |
| Ракообразные (всего) | 88.1 | 40.3 | 42.9 | 30.5 | 25.4 | 63.1 | 67.2 |
| Appendicularia | + | ||||||
| Pyrosoma atlanticum | 0.9 | 1.7 | 11.6 | 6.6 | |||
| Salpa sp. | 1.3 | 13.5 | 11.3 | 10.5 | 9.5 | 1.0 | |
| Tunicata | 4.8 | 10.3 | 4.9 | 1.0 | |||
| Туникаты (всего) | 6.1 | 14.4 | 23.3 | 10.5 | 26.0 | 8.6 | + |
| Chauliodus sp. | 2.2 | ||||||
| Vinciguerria sp. | 0.5 | ||||||
| Maurolicus muelleri | 0.7 | 9.7 | 5.7 | 2.0 | |||
| Stomiatidae | 1.0 | ||||||
| Diaphus sp. | 5.9 | 1.5 | 7.5 | ||||
| Diaphus dumerilii | 28.5 | ||||||
| Lampadena sp. | 2.2 | ||||||
| Myctophidae | 12.7 | 7.8 | 18.9 | 5.2 | 7.0 | ||
| Tetragonurus cuvieri | 9.7 | 4.8 | |||||
| Chlorophthalmus agassizi | 6.3 | ||||||
| Bathylagidae | 0.8 | ||||||
| Paralepis sp. | 1.1 | ||||||
| Epigonus sp. | 7.6 | 3.3 | |||||
| Nansenia sp. | 1.9 | 1.3 | |||||
| Рыбы неопределённые | 7.4 | 5.4 | 16.4 | 2.3 | 10.1 | ||
| Рыбы (всего) | 2.6 | 42.0 | 27.5 | 54.5 | 25.2 | 17.1 | 28.5 |
| Переваренная пища | 1.9 | 0.6 | 2.8 | 0.4 | 0.1 | 0.5 | |
| Переваренность пищи, баллы | $\frac{{2.1 \pm 0.2}}{{0.8}}$ | $\frac{{2.2 \pm 0.2}}{{0.7}}$ | $\frac{{2.6 \pm 0.9}}{{0.7}}$ | $\frac{{2.6 \pm 0.1}}{{0.5}}$ | $\frac{{2.3 \pm 0.2}}{{0.8}}$ | $\frac{{2.3 \pm 0.1}}{{0.7}}$ | 2 |
| Масса рыбы, г | $\frac{{170.6 \pm 5.9}}{{42.3}}$ | $\frac{{256.2 \pm 6.1}}{{54.9}}$ | $\frac{{452.2 \pm 7.6}}{{65.5}}$ | $\frac{{712.8 \pm 16.1}}{{126.0}}$ | $\frac{{1018.8 \pm 14.4}}{{122.9}}$ | $\frac{{1447.7 \pm 43.2}}{{193.1}}$ | $\frac{{2120.0 \pm 123.4}}{{213.8}}$ |
| Длина рыбы (TL), мм | $\frac{{237.0 \pm 0.2}}{{1.7}}$ | $\frac{{278 \pm 0.2}}{{1.5}}$ | $\frac{{330.3 \pm 0.2}}{{1.5}}$ | $\frac{{380.0 \pm 0.2}}{{1.5}}$ | $\frac{{428.0 \pm 0.1}}{{1.3}}$ | $\frac{{478.0 \pm 0.3}}{{1.5}}$ | $\frac{{540.0 \pm 1.0}}{{1.7}}$ |
| Индекс наполнения желудков, ‱ | $\frac{{141.8 \pm 31.1}}{{224.0}}$ | $\frac{{28.0 \pm 4.5}}{{40.6}}$ | $\frac{{34.3 \pm 4.8}}{{38.7}}$ | $\frac{{63.1 \pm 9.2}}{{67.3}}$ | $\frac{{49.9 \pm 7.7}}{{53.0}}$ | $\frac{{74.1 \pm 18.9}}{{84.0}}$ | $\frac{{72.7 \pm 42.9}}{{74.4}}$ |
| Число желудков общее/с пищей | 52/44 | 80/56 | 74/65 | 61/50 | 74/48 | 20/15 | 3/3 |
| Размер компонента пищи, мм | $\frac{{21.3 \pm 2.5}}{{16.2}}$ | $\frac{{33.7 \pm 3.6}}{{21.6}}$ | $\frac{{23.0 \pm 1.9}}{{17.9}}$ | $\frac{{43.0 \pm 3.4}}{{32.2}}$ | $\frac{{44.6 \pm 4.0}}{{32.9}}$ | $\frac{{50.9 \pm 5.0}}{{23.9}}$ | $\frac{{42.0 \pm 23.4}}{{46.8}}$ |
| Индекс длины жертвы, % | $\frac{{0.09 \pm 0.01}}{{0.07}}$ | $\frac{{0.12 \pm 0.01}}{{0.08}}$ | $\frac{{0.10 \pm 0.01}}{{0.05}}$ | $\frac{{0.11 \pm 0.09}}{{0.08}}$ | $\frac{{0.10 \pm 0.01}}{{0.08}}$ | $\frac{{0.11 \pm 0.01}}{{0.05}}$ | $\frac{{0.08 \pm 0.04}}{{0.09}}$ |
Рыбы (42.0%) составляли основу питания берикса TL 250–299 мм. В желудках встречались неидентифицированные миктофиды (12.7%), Diaphus sp. (5.9%), M. muelleri (9.7%) и Chlorophthalmus agassizi (6.3%). Доля ракообразных в пище была чуть меньше – 40.0%, среди них доминировала креветка O. novaezeelandiae (23.2%). Туникаты (14.4%), в основном сальпы (13.5%), составили третью по массе пищевую группу берикса. Доля кальмаров (2.7%) была незначительной; 71% особей берикса этой размерной группы питались, среднее значение ИНЖ составило 28.0‱ при средней переваренности пищи в 2.2 балла.
Берикс TL 300–349 мм питался ракообразными (42.9%), которые были главной пищей, рыбой (27.5%), туникатами (23.3%) и кальмарами (3.5%). Среди ракообразных в желудках берикса преобладали креветки O. novaezeelandiae (10.2%) и F. woodwardi (8.8%). Миктофиды были доминирующими среди рыб (7.8%). В сходных долях встречались Epigonus sp. (7.6%) и M. muelleri (5.7%). Туникаты в желудках берикса были представлены сальпами (11.3%) и P. atlanticum. Доля кальмаров (3.5%) в пище была невелика; 87% особей берикса питались, ИНЖ составил 34.3‱ при средней переваренности пищи в 2.6 балла.
Главной пищей берикса TL 350–399 мм были рыбы (54.5%), второстепенной – ракообразные (30.5%), туникаты (10.5%) и кальмары (4.1%). Среди рыб преобладали миктофиды (18.9%) и Tetragonurus cuvieri (9.7%). Доля остальных видов рыб была небольшой. В пище берикса среди ракообразных ведущим видом была креветка F. woodwardi (13.7%).
Доля остальных ракообразных была незначительной. Среди туникат в пище берикса доминировали сальпы (10.5%). Кальмары (4.1%) были представлены в основном сильно переваренными, неидентифицированными особями и Abraliopsis atlantica (0.1%).
Питались 81.9% особей берикса, ИНЖ составил 63.1‱ при средней переваренности пищи в 2.6 балла.
Пища берикса TL 400–449 мм состояла главным образом из туникат (26.0%). Ракообразные (25.3%), рыба (25.2%) и кальмары (23.3%) находились в желудках в сходных долях. Пиросома P. atlanticum (11.6%) и сальпы (9.5%) доминировали среди туникат. Креветки F. woodwardi (6.9%) и O. novaezeelandiae (5.5%) имели наибольшую долю среди ракообразных. Рыбы в пище берикса были представлены Diaphus sp. (7.5%), неидентифицированными миктофидами (5.2%), T. cuvieri (4.8%) и другими видами. Неидентифицированные кальмары (23.0%) составляли бо́льшую часть массы кальмаров в пище берикса; 71.6% особей берикса питались, ИНЖ составил 49.9‱ при средней переваренности пищи в 2.3 балла.
Ракообразные (63.1%) были главной пищей берикса TL 450–499 мм, среди которых доминировала креветка F. woodwardi (47.4%). Второстепенную пищу составляли рыбы (17.1%), кальмары (10.7%) и туникаты (8.7%). Представители отряда Oegopsida (8.2%) имели наибольшую долю среди кальмаров. Пиросома P. atlanticum (6.6%) доминировала среди туникат; 75% берикса питались, ИНЖ составил 74.1‱ при средней переваренности пищи в 2.3 балла.
Питание берикса размерной группы TL 500–559 мм было изучено по данным трёх желудков и носит справочный характер (табл. 4).
ОБСУЖДЕНИЕ
Состав пищи берикса в водах Атлантического океана сходен на уровне основных таксонов: классов, отрядов и семейств. Берикс акваторий Углового и Азорского поднятий питается мезопелагическими рыбами, кальмарами, ракообразными, иглокожими и полихетами (Пшеничный и др., 1986; Винниченко, 1997; Козлов, 2004; Виноградов др., 2005). В водах Канарских островов пища берикса состоит из мелкой рыбы, ракообразных и головоногих моллюсков (Dürr, González, 2002). У берегов Бразилии берикс питается мезопелагическими ракообразными, кальмарами и рыбой (Muto et al., 2005). Сходный спектр питания был в районе подводного поднятия Китового хребта, где берикс питался мезопелагическими ракообразными, рыбами, туникатами, головоногими и крылоногими моллюсками, полихетами (Дудочкин, Котляр, 1989). Подобный состав пищевых организмов берикса на поднятиях Китового хребта подтверждает настоящее исследование по данным 1988–1990 гг.
Во всех акваториях в пище берикса доминируют мезо- и макропланктонные организмы, входящие в состав фауны звукорассеивающих слоёв (ЗРС). При определенных условиях, когда на вершинах подводных поднятий находится богатая донная фауна, берикс частично использует в пищу донные организмы (Пшеничный и др., 1986). На подводных поднятиях Китового хребта донные организмы в пище берикса в 1988–1990 гг. обнаружены не были, но есть сведения о находке полихет (0.5%) в пище берикса на одном из поднятий Китового хребта (Дудочкин, Котляр, 1989). Авторы этого сообщения не отнесли найденных полихет к донному или пелагическому сообществам фауны, что не позволяет исключить донные организмы из пищи берикса Китового хребта.
Спектр питания берикса, изученный по материалам ловов на пяти поднятиях Китового хребта, состоит из 45 компонентов (таксонов, до которых возможно было провести идентификацию) пищи: ракообразные (23), рыбы (15), туникаты (4) и кальмары (3). Ранее Дудочкин и Котляр (1989) обнаружили в питании берикса на одном из поднятий Китового хребта 44 компонента пищи: ракообразные (28), рыбы (9); моллюски (5), включая головоногих и крылоногих моллюсков; туникаты (1) и полихеты (1). Выявленные отличия связаны с большим количеством исследованных желудков берикса, собранного на пяти подводных поднятиях, что расширило спектр питания берикса в водах Китового хребта.
Особенности питания берикса изученного района. Оценка сходства питания берикса по компонентам пищи на разных подводных поднятиях Китового хребта показала, что степень сходства была низкой между поднятием банка Северная и поднятиями Щедрая, Вальдивия, Бета, Альфа-2. Высокая степень сходства была между поднятиями Щедрая и поднятиями Бета и Альфа-2, между поднятиями Бета и Альфа-2 (табл. 5). Вероятно, между поднятиями разная степень сходства питания берикса связана с составом доступной кормовой фауны, зависящим от условий среды на каждом из поднятий.
Таблица 5.
Степень сходства питания берикса (индекс Чекановского–Сёренсена) на подводных поднятиях Китового хребта
| Подводное поднятие | Число компонентов пищи | Подводное поднятие | ||||
|---|---|---|---|---|---|---|
| Северная | Щедрая | Вальдивия | Бета | Альфа-2 | ||
| Северная | 33 | 0.29 | 0.23 | 0.35 | 0.31 | |
| Щедрая | 11 | 0.29 | 0.35 | 0.43 | 0.41 | |
| Вальдивия | 8 | 0.23 | 0.35 | 0.33 | 0.29 | |
| Бета | 20 | 0.35 | 0.43 | 0.33 | 0.41 | |
| Альфа-2 | 26 | 0.31 | 0.41 | 0.29 | 0.41 | |
Ракообразные в пище берикса составляли наибольшую долю по массе на поднятиях Южного блока (53.9%), меньшую – в водах Центрального (38.2%) и Северного (35.2%) блоков (рис. 2). Рыбы были главной пищей берикса (56.0%) на поднятии Северного блока. В водах Центрального (33.0%) и Южного (16.0%) блоков поднятий доля рыбы в пище берикса снижалась. Туникат в пище берикса чаще встречался на поднятиях Южного блока (20.3%), а его доля в водах Центрального блока составляла 11.6%, в Северном блоке – 5.7%. Кальмары в небольшой массе встречались в пище берикса на всех блоках поднятий Китового хребта: на Северном комплексе (2.5%), на Центральном (11.8%), на Южном (9.3%). Все обнаруженные пищевые организмы берикса характерны для мезо- и макропланктона фауны ЗРС (Парин, 1968; Ломакина, 1978; Несис, 1982; Овен и др., 1984; Parin, 1986).
Рис. 2.
Спектры питания (доля компонента пищи по массе) берикса Beryx splendens на подводных блоках поднятий Китового хребта: 1 – Северный, 2 – Центральный, 3 – Южный, 4 – Китовый хребет в целом (по: Дудочкин, Котляр, 1989); ( ) – прочая пища, ост. обозначения см. на рис. 1.
) – прочая пища, ост. обозначения см. на рис. 1.

Сравнение спектра питания берикса в декабре–январе 1989–1990 гг. с данными, приводимыми Дудочкиным и Котляром (1989) для октября 1979 г., показывает значительное сходство с питанием берикса в водах Центрального блока подводных поднятий. При сходстве пищи на уровне высших таксонов видовой состав пищевых организмов на уровне родов и видов различается (табл. 1–3, рис. 2). Эти различия, во-первых, могут быть связаны с сезоном лова: сентябрь–октябрь (Дудочкин, Котляр, 1989) и декабрь–январь в нашем сообщении. Во-вторых, в 1989–1990 гг. изучено питание более крупного берикса (190–560 мм), чем у Дудочкина и Котляра (1989), – 160–330 мм. В-третьих, состав потребляемых пищевых организмов берикса во многом зависит от их доступности, зависящей от мозаичности распределения планктона (Беклемишев, 1969; Механизмы …, 2017) в водах подводных поднятий, которая в свою очередь связана с турбулентностью вод (Дарницкий, Болдырев, 1987; Stavn, 1971), концентрацией организмов мезо- и макропланктона ЗРС во время их опускания на подводное поднятие в процессе суточной вертикальной миграции, а также с тем, что планктонные организмы могут быть нанесены течением на подводное поднятие (Кашкин, 1977; Пахоруков и др., 2014). Имеются сообщения, что состав пищевых объектов (креветок, рыб) берикса во многом зависит от фазы луны (Винниченко, 1997; Salini et al., 2001). Винниченко (1997) обратил внимание на разделение слоёв ЗРС и их состав. Верхний слой состоит преимущественно из эвфаузиид, а нижний – из скопления мелких мезопелагических рыб. Мелкий берикс питается в верхнем слое ЗРС, крупный – в нижнем (Винниченко, 1997). Разделение фауны ЗРС по слоям может дать объяснение присутствию в пище берикса небольшого количества копепод, мизид и эвфаузиид из верхних слоев ЗРС, так как более крупный берикс Китового хребта выбирает более крупную пищу и питается в более глубоких слоях ЗРС. Немаловажным фактором является большее количество материала, использованного для данной работы, что позволило выявить достаточно редкие компоненты пищи берикса, не учтённые ранее. Таким образом, разная степень сходства в питании берикса из различных блоков поднятий скорее всего связана со всем многообразием факторов, влияющих на пищевое поведение берикса (состав доступной кормовой фауны; условия среды данного комплекса, которые определяют миграции ЗРС, размеры и физиологическое состояние берикса), а также со временем лова и объёмом собранного материала.
Особенности питания берикса разного размера. Переход от мелких жертв к более крупным характерен для низкотелого берикса (Дудочкин, Котляр, 1989; Dürr, González, 2002; Виноградов и др., 2005; Horn et al., 2010), что находит подтверждение в материалах нашей работы. По мере увеличения размеров берикса от 190 до 449 мм доля ракообразных снижается с 88.1 до 25.4%, а возрастает доля туникат и кальмаров (табл. 4). Такие изменения связаны с увеличением доступности более крупных жертв по мере увеличения размеров берикса. Берикс выбирает для питания организмы размером 0.9–130.0 (в среднем 36.1) мм (стандартная ошибка среднего 1.7, стандартное отклонение 28.3), при этом диапазон соотношения размеров жертв и размеров берикса для всех его размерных групп достаточно узок и составляет 9.7–11.5%.
Временны́е (суточные) изменения питания берикса. Большинство авторов связывают питание берикса и его вертикальные миграции с суточными вертикальными миграциями организмов ЗРС (Галактионов, 1984; Винниченко, 1997; Horn et al., 2010). Хорн с соавторами (Horn et al., 2010) указывают на отсутствие связи между временем суток и питанием берикса на месте исследования, но добавляют, что берикс питался и днём, и ночью. Существуют работы, связывающие миграции и питание рыб с фазами луны (Salini et al., 2001) и освещённостью, определяющей доступность пищи берикса в течение суток (Винниченко, 1997). Таким образом, связь между питанием берикса и его вертикальными миграциями существует, но детально механизм этой связи пока не изучен.
Выполнить рекомендованную методиками суточную станцию по изучению динамики питания не удалось. Представление об изменениях в питании берикса в течение суток было получено, когда были сгруппированы все имеющиеся данные по составу пищи, накормленности и степени переваренности пищи по периодам суток: утро–день–вечер–ночь. Наибольшая накормленность (ИНЖ 68‱) берикса при самой низкой переваренности пищи (2.40 балла) отмечена в вечерние часы. Накормленность снижалась в ночное время (ИНЖ 41‱) при минимальной переваренности пищи (2.42 балла). В вечернее время берикс образовывал промысловые скопления в пелагиали над подводным поднятием. Такие скопления распадаются во второй половине ночи, и уловы берикса падают. Состав пищи в вечерний и ночной периоды был сходен: основные объекты – ракообразные, второстепенные – рыбы, туникаты и кальмары (рис. 1). Минимальная накормленность (ИНЖ 18.8‱) при высокой переваренности пищи (2.65 балла) отмечена в утренние часы. Состав пищи в этот период был сходным с таковым в вечернее и ночное время. В некоторых случаях в светлое время суток эхолот показывал скопления рыб на дне поднятий в виде “щёточки”. Подобные скопления состояли из беспузырного окуня и кабан-рыбы, берикс в уловах встречался единично (Гущин, 2021). Днём главной пищей берикса были кальмары (83.7%), второстепенной – рыбы, туникаты и ракообразные. Масса кальмаров в пище берикса в дневное время представляется завышенной, так как на этот период пришлись всего две пробы. Таким образом подтверждается связь питания берикса с миграциями организмов ЗРС, когда максимум накормленности и минимальная переваренность пищи берикса совпадают с подъёмом организмов ЗРС в верхние горизонты воды.
ЗАКЛЮЧЕНИЕ
Подтверждается, что берикс, обитающий в зоне подводных поднятий Китового хребта, имеет широкий пищевой спектр, состоящий из 45 компонентов, включая ракообразных, рыб, туникат и кальмаров, относящихся к мезо- и макропланктону мезопелагиали и совершающих суточные вертикальные миграции в составе ЗРС. По составу пищи высокая степень сходства (индекс Чекановского–Сёренсена) была между пищей берикса в водах поднятия Щедрая и поднятий Бета, Альфа-2, между поднятиями Бета и Альфа-2. Низкая степень сходства была между питанием берикса на поднятии Северная и на поднятиях Щедрая, Вальдивия, Бета, Альфа-2. Предполагается, что разная степень сходства в пище берикса на этих поднятиях связана с различиями в составе фауны, доступностью пищевых организмов во время их суточной вертикальной миграции, условиями среды блоков поднятий, размерами берикса, сезоном и временем лова. Донные организмы в пище берикса отсутствовали на всех поднятиях. Ракообразные по массе служили главной пищей берикса (53.9%) в водах Южного и Центрального (38.2%) блоков поднятий. Рыбы были главной пищей берикса (56.0%) на поднятии Северного блока. Туникаты и кальмары встречались в виде второстепенной пищи берикса всех блоков поднятий. Берикс выбирает для питания организмы размером 0.9–130.0 (в среднем 36.1) мм. По мере увеличения размеров берикса увеличиваются и размеры его жертв, что вызывает смену видов пищевых организмов, но при этом соотношение размеров жертв и особей берикса изменяется мало и составляет 9.7–11.5%. Подтверждена связь питания берикса с суточными миграциями мезо- и макропланктона в составе ЗРС. Наибольшая накормленность берикса и минимальная переваренность пищи приходились на вечер и начало ночи, когда организмы ЗРС начинают подъём в верхние горизонты воды. Главной пищей берикса в вечернее и ночное время являются ракообразные.
Список литературы
Беклемишев К.В. 1969. Экология и биогеография пелагиали. М.: Наука, 291 с.
Винниченко В.И. 1997. Вертикальные суточные миграции низкотелого берикса Beryx splendens (Berycidae) на подводных возвышенностях открытой части северной Атлантики // Вопр. ихтиологии. Т. 37. № 4. С. 490–496.
Виноградов В.И., Козлов Д.А., Кукуев Е.И. 2005. О питании берикса-альфонсина Beryx splendens Lowe (Berycidae) на подводных возвышенностях в районе Азорских островов // Океанология. Т. 45. № 4. С. 567–573.
Галактионов Г.3. 1984. Особенности стайного поведения берикса-альфонсина Beryx splendens Lowe (Berycidae) талассобатиали Атлантического океана // Вопр. ихтиологии. Т. 24. № 5. С. 863–865.
Гущин А.В. 2021. Питание рыб подводных поднятий Китового хребта (Юго-восточная Атлантика). 1. Беспузырный окунь Helicolenus mouchezi, кабан-рыба Pentaceros richardsoni, красноглазка Emmelichthys nitidus подводной горы Вальдивия // Там же. Т. 61. № 2. С. 178–184. https://doi.org/10.31857/S0042875221020107
Болдырев В.З., Дарницкий В.Б., Куликов М.Ю. 1987. Формирование биологической продуктивности в районах поднятий океанского ложа // Биологические ресурсы открытого океана. М.: Наука. С. 31–64.
Дубравин В.Ф. 2001. Поверхностные водные массы и формирование зон биологической продуктивности Атлантического океана. СПб.: Гидрометеоиздат, 125 с.
Дубравин В.Ф. 2013. Атлас термохалинной и биогеографической структур вод Атлантического океана. Калининград: Капрос, 471 с.
Дудочкин А.С., Котляр А.Н. 1989. О питании берикса Beryx splendens // Вопр. ихтиологии. Т. 29. № 1. С. 135–141.
Ильин А.В. 1976. Геоморфология Атлантического океана. М.: Наука, 232 с.
Инструкция по производству биологических работ и первичной обработке данных на судах Запрыбпромразведки. 1977. Калининград: Запрыбпромразведка, 200 с.
Кашкин Н.И. 1977. Фауна звукорассеивающих слоев // Биология океана. Биологическая структура океана. Т. 1. М.: Наука. С. 299–317.
Козлов Д.А. 2004. Биология и промысел низкотелого берикса (Beryx splendens Lowe, 1833) на подводных возвышенностях северной части Атлантики: Автореф. дис. … канд. биол. наук. Калининград: КГТУ, 24 с.
Ломакин И.Э. 2014. Террасы подводных гор и тектоника дна Мирового океана. Саарбрюккен: LAP Lambert Acad. Publ., 103 с.
Ломакина Н.Б. 1978. Эуфаузииды Мирового океана. Л.: Наука, 222 с.
Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях. 1974. М.: Наука, 254 с.
Механизмы образования скоплений и функционирования планктона в экосистемах Индийского океана. 2017. Белгород: Константа, 360 с.
Несис К.Н. 1982. Краткий определитель головоногих моллюсков Мирового океана. М.: Лег. и пищ. пром-сть, 358 с.
Овен Л.С., Салехова Л.П., Никольский В.Н. 1984. Сравнительная характеристика фауны звукорассеивающих слоев тропической зоны Атлантического океана // Экология моря. Т. 18. С. 8–17.
Парин Н.В. 1968. Ихтиофауна океанской эпипелагиали. М.: Наука, 186 с.
Пахоруков Н.П. 1980. Глубоководные придонные рыбы Китового хребта и прилежащих районов // Рыбы открытого океана. М.: Изд-во ИО РАН. С. 19–31.
Пахоруков Н.П. 2003. Рыбы подводных гор тропической зоны мирового океана: состав фауны, поведение и экологическая классификация: Автореф. дис. … канд. биол. наук. М.: ИО РАН, 24 с.
Пахоруков Н.П., Парин Н.В., Царин С.А., Данилюк О.Н. 2014. Результаты визуальных наблюдений рыб из ПОА “Север-2” на хребте Вавилова (Атлантический океан) // Мор. экол. журн. Т. 13. № 4. С. 15–28.
Песенко Ю.А. 1982. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 287 с.
Пшеничный Б.Н., Котляр А.Н., Глухов А.А. 1986. Рыбные ресурсы талассобатиали Атлантического океана // Биологические ресурсы Атлантического океана. М.: Наука. С. 230–252.
Федоров В.В., Карамышев А.К. 1991. Трофическая структура бентоса вершин Китового хребта и условия её формирования. М.: Изд-во ВНИРО. С. 6–18
Dürr J., González J.A. 2002. Feeding habits of Beryx splendens and Beryx decadactylus (Berycidae) off the Canary Islands // Fish. Res. V. 54. № 3. P. 363–374. https://doi.org/10.1016/S0165-7836(01)00269-7
Horn P. L., Forman J., Dunn M.R. 2010. Feeding habits of alfonsino Beryx splendens // J. Fish Biol. V. 76. № 10. P. 2382–2400. https://doi.org/10.1111/j.1095-8649.2010.02630.x
Muto E.Y., da Silva M.H.C., Vera G.R. et al. 2005. Alimentaçao e relações tróficas de peixes demersais da plataforma continental externa e talude superior da Região Sudeste-Sul do Brasil. São Paulo: Intituto Oceanográfico USP, 65 p.
Parin N.V. 1986. Distribution of mesobenthopelagic fishes in slope waters and around submarine rises // Pelagic Biogeography. Paris: UNESCO. P. 226–229.
Salini J., Brewer D., Farmer M., Jones P. 2001. Lunar periodicity of prawns and by-catch in trawls from the Gulf of Carpentaria, northern Australia // Mar. Biol. V. 138. № 5. P. 975–983. https://doi.org/10.1007/s002270000528
Stavn R.N. 1971. The horizontal-vertical distribution hypothesis: Langmuir circulation and Daphnia distribution // Limnol. Ocean. V. 16. № 2. P. 453–466. https://doi.org/10.4319/lo.1971.16.2.0453
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии