

Вопросы ихтиологии, 2022, T. 62, № 5, стр. 500-523
Морские петухи рода Lepidotrigla (Triglidae) залива Нячанг и сопредельных акваторий (Вьетнам, Южно-Китайское море). Виды с короткими грудными плавниками
А. М. Прокофьев *
Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 27.01.2022
После доработки 11.03.2022
Принята к публикации 14.03.2022
- EDN: UOWMWY
- DOI: 10.31857/S0042875222050174
Аннотация
Приведён обзор морских петухов рода Lepidotrigla залива Нячанг и сопредельных вод. В указанной акватории встречено шесть видов с короткими грудными плавниками, не заходящими за шестой луч анального плавника: L. deasoni, L. firmisquamis, L. marisinensis, L. punctipectoralis, L. cf. spiloptera и L. venusta. Приведены их детальные иллюстрированные описания. Виды L. deasoni и L. marisinensis впервые отмечены в ихтиофауне Вьетнама. Выявлен ряд отличий вьетнамских экземпляров от типичных L. spiloptera из индо-австралийских вод, не позволяющих исключать принадлежность вьетнамского материала к неописанному виду. Помимо указанных видов, в изученной акватории известны представители рода с длинными грудными плавниками, относимые к группе видов “L. japonica”, систематика которой нуждается в серьёзной ревизии. Составлен ключ для определения видов Lepidotrigla, известных из Южно-Китайского моря.
Род Lepidotrigla Günther, 1860 является наиболее богатым видами в семействе Triglidae: в настоящее время считаются валидными 54 вида, населяющие умеренные и тропические воды Восточной Атлантики и Индо-Вест-Пацифики, с максимальным видовым разнообразием в водах Южной Японии, Южно-Китайского моря, Индо-Австралийского архипелага и Австралии (Fricke et al., 2022). Видовой состав этого рода до сих пор окончательно не установлен и до настоящего времени продолжаются находки новых для науки видов (Gomon, Kawai, 2018; Gomon, Psomadakis, 2018; Prokofiev, Yato, 2020). В фауне Вьетнама ранее было отмечено 10 видов (Nguyen, 1999), хотя ряд определений могут быть ошибочными. Так, указания на присутствие в Южно-Китайском море японского вида L. kanagashira Kamohara, 1936, по данным Ли (Li, 1981), относятся к описанным им видам L. lepidojugulata и L. longimana. Тем не менее, в будущем можно ожидать новые находки для фауны Вьетнама, так как всего в Южно-Китайском море встречено не менее 18 видов рода (Randall, Lim, 2000; Prokofiev, Yato, 2020), причём в их перечень не были включены виды L. kishinoueyi Snyder, 1911 и L. microptera Günther, 1873, указываемые для Тонкинского залива Нгуеном (Nguyen, 1999), хотя L. microptera приводится для Южно-Китайского моря и в ряде других источников (Ochiai, Yatou, 1985; Yamada, 2002). Виды рода Lepidotrigla отличаются от представителей других родов триглид присутствием развитых ростральных выступов (отростков), крупной чешуёй, выступающими наружу в виде щитков и/или шипов пластинчатыми разрастаниями птеригиофоров как колючей, так и мягкой части D и наличием посторбитальной бороздки (Richards, Jones, 2002).
В собранной мною коллекции прибрежно-морских рыб южной части Центрального Вьетнама (заливы Нячанг, Ванфонг и Фантьет) присутствуют представители шести видов рода Lepidotrigla. Настоящая публикация посвящена описанию этого материала. Не считая недавнего описания нового вида по этим сборам (Prokofiev, Yato, 2020), сведения о присутствии видов Lepidotrigla во Вьетнаме до сих пор относились исключительно к Тонкинскому заливу (Nguyen, 1999). Все рассматриваемые в данной работе виды относятся к короткоплавниковым представителям Lepidotrigla, у которых Р не заходит далее середины длины основания А (Prokofiev, Yato, 2020). Помимо них в рассматриваемой акватории известны виды с длинными Р, простирающимися за середину длины основания (иногда за конец) А, и с не покрытой чешуёй превентральной областью. В фауне Вьетнама такие морские петухи обычно определяются как L. japonica (Bleeker, 1854) (Nguyen, 1999; Prokofiev, Yato, 2020; Shao, 2022). Хотя в моих сборах из зал. Нячанг длинноплавниковые виды не обнаружены, один экземпляр был найден в коллекции Института океанографии (г. Нячанг) (рис. 1а). Ещё один длинноплавниковый экземпляр Lepidotrigla (ювенильный) добыт у о-вов Кото в южной части зал. Халонг (Тонкинский залив) (рис. 1б). От экземпляра из коллекции Института океанографии он отличается гораздо более короткими и широкими ростральными отростками и наличием тёмного (при жизни – красного) пятна в заднем отделе колючей части D. Группа видов “L. japonica” нуждается с серьёзной ревизии, определение её номинальных видов затруднительно (Gomon, Kawai, 2018). Учитывая крайнюю ограниченность собственного материала и неразработанность систематики группы, в настоящий обзор длинноплавниковые виды не включены.

МАТЕРИАЛ И МЕТОДИКА
Изученный материал хранится в коллекции Института океанологии РАН, Москва. Рыбы были получены из траловых уловов местного промысла, в связи с чем при перечислении материала я ограничиваюсь указанием города, на рынках которого был куплен соответствующий экземпляр. Молодь Lepidotrigla достаточно часто встречалась при проведении траловой съёмки в прибрежье Вьетнама, однако из-за значительной морфологической трансформации в онтогенезе и совершенно неразработанных критериев диагностики ювенильных особей этот материал оставлен за рамками настоящей статьи. Признаки, приведённые в видовых описаниях, относятся только к изученному материалу, однако в диагнозах учтены данные литературы по соответствующему виду. Анализ меристических и пластических признаков проведён согласно описанной ранее методике (Hubbs, Lagler, 1958; Richards, Saksena, 1977; Proko-fiev, Yato, 2020). Последний разделённый до основания луч вертикальных плавников принят за один. Номенклатура гребней головы дана в соответствии с приведённой ранее (Gomon, Kawai, 2018; Prokofiev, Yato, 2020). Длина шипов головы измерена от основания гребня шипа. Типы строения ростральных отростков даны по работе Прокофьева и Ято (Prokofiev, Yato, 2020). В тексте использованы следующие сокращения: SL – стандартная длина; D, A, P, V и С – соответственно спинной, анальный, грудные, брюшные и хвостовой плавники; LL, LL–D и LL–A – соответственно боковая линия, число чешуй между LL и началами D и А; USNM – Национальный музей, Смитсоновский институт, Вашингтон (США); MNHN – Парижский национальный музей естественной истории, Франция.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Lepidotrigla deasoni Herre et Kauffman, 1952
Рис. 2.
Lepidotrigla deasoni SL 204 мм, г. Нячанг (а, б, г, д) и голотип USNM 202504 (в): а – общий вид; б, в – ростральные выступы; г – голова, вид сверху; д – шипы и гребни головы, дорсолатерально. Шипы: cl – клейтральный, op – оперкулярный, ptt – посттемпоральный, pos – посторбитальные, prs – преорбитальные, ss – сфенотикальный; гребни: pc – париетальный, sc – сфенотикальный; po – посторбитальная бороздка, ro – ростральные выступы. Масштаб, мм: б – 22, в – 10; г, д – 18, линейка общая .

Рис. 3.
Lepidotrigla deasoni SL 204 мм, г. Нячанг: а – аберрантная колючка D (accs), б – пигментация перепонки колючей части D, в – внутренняя сторона Р. Масштаб: 15 мм.

Материал. 1 экз. SL 204 мм, г. Нячанг (рис. 2а).
Сравнительный материал. USNM 202504 – голотип L. deasoni, фотографии в разных ракурсах.
Диагноз. Ростральные выступы короткие треугольные, с дополнительными вершинами кнутри от главной; заглазничная бороздка широко прерванная; преорбитальные и посторбитальные шипы имеются, сфенотикальные имеются или отсутствуют, париетальные шипы отсутствуют; P короткий, его внутренняя сторона у фиксированных рыб с очень размытым просветлением над интенсивно-чёрным участком пигментации; чешуя легко опадающая, на груди отсутствует; LL 55–58.
Описание (рис. 2а). D VIII (+I) + 15 (в скобках указана аберрантно расположенная колючка), A 15, P 11 + 3, V I + 5, LL 58, LL–D 3, LL–A 14.
Ростральные выступы (рис. 2б–2г) короткие, в форме равнобедренного треугольника; их вершины расходящиеся, но за счёт того, что выступы сравнительно короткие и с широким основанием, соизмеримы с длиной выступа, степень их расхождения кажется небольшой (в действительности же угол, образуемый продольной осью отростка и срединно-брюшной осью тела, составляет около 25°). Боковые края ростральных выступов практически прямые; наружный край несёт мелкие зубчики, едва превышающие по величине бугорки скульптуры покровных костей черепа; внутренний край с тремя маленькими выступами, постепенно уменьшающимися по величине от ближайшего к главной вершине к наиболее дальнему от неё, за которым следует ряд из более мелких бугорков краевой скульптуры предглазничной кости. Интерростральный промежуток заметно превышает межглазничный. Передний край интеррострального промежутка вогнут посередине. На щеке имеется слабый продольный гребень, начинающийся на уровне конца передней трети орбиты, переходящий на praeoperculum и оканчивающийся тупым выступом (с правой стороны – маленьким шипом). Надглазничные гребни слабо обособлены, параллельны, межглазничный промежуток между ними глубоко вогнут. Преорбитальных шипов два, небольшие, но отчётливые (задний – чуть крупнее), острые, расставленные (рис. 2г). Посторбитальные шипы с левой стороны очень малы, с правой – практически не выражены, в виде бугорков. Верхний посторбитальный шип направлен назад, нижний – вбок и немного вниз. Сфенотикальный шип с левой стороны маленький, но отчётливый, острый, с правой отсутствует (здесь не выражен даже его гребень) (у голотипа отсутствует с обеих сторон). Париетальные гребни малы, едва заметны; париетальные шипы отсутствуют. Посторбитальная бороздка глубокая, но короткая, не заходит за медиальную границу надглазничного гребня (рис. 2г, 2д). Оперкулярный и посттемпоральный шипы хорошо выражены, острые, соизмеримые по величине. Оперкулярный шип направлен вершиной назад и немного вверх (слева) или вниз (справа). Вершина оперкулярного шипа расположена позади уровня вершины посттемпорального шипа. Клейтральный шип хорошо развит, острый, заметно длиннее остальных шипов, расположен вентрально от оперкулярного шипа, его вершина оканчивается на вертикали основания шестой колючки D. Покровные кости черепа и плечевого пояса густо усеяны мелкими бугорками, организованными в ряды.
Челюсти равной длины. Верхняя челюсть оканчивается на вертикали переднего края орбиты. Симфиз praemaxillaria отчётливо вогнут между передними концами зубных рядов соседних сторон, в эту выемку входит вершина симфиза dentalia. В челюстях мелкие бугорковидные зубы полосками; озубление praemaxillaria полностью видно при закрытом рте. Сошник и нёбные кости без зубов. Язык очень массивный, широкий, едва обособленный. Жаберных тычинок на 1-й дуге в наружном ряду 10 (1 + 6 + iii) (семь развитых + три бугорковидных; кроме них, между тычинками на ceratobranchiale с первой по шестую имеется по плоскому бугорку), во внутреннем – восемь; в наружном ряду на 2-й дуге – 10 (i + 7 + ii). Есть щель позади четвёртой жаберной дуги. Ложножабра крупная, из 29 элементов.
Вдоль оснований обоих спинных плавников парный ряд из 23 костных щитков, из которых восемь под основанием колючей части D, один в интердорсальном промежутке, а остальные – под основанием мягкой части D (последний – под его 14-м лучом); начиная с третьего щитки оканчиваются шипами, спереди многовершинными (на первых двух щитках они заменены бугорками), а с шестого (справа – с седьмого) – одним острым и сильным шипом. В колючей части D первые три колючки наибольшие и значительно толще и массивней остальных, особенно первая и вторая; вторая колючка самая длинная, в 1.2 раза длиннее третьей и в 1.3 раза длиннее первой; последующие колючки постепенно уменьшаются. С левой стороны в основании первой колючки (постеролатерально) присутствует аберрантная дополнительная маленькая колючка (в 4.5 раза меньше длины основной колючки), которая нормально сочленена с первым дорсальным птеригиофором (рис. 3а, 3б). Передняя и переднебоковые поверхности первых двух колючек D мелко зазубрены, на переднебоковых сторонах третьей колючки прослеживаются нечёткие мелкие бугорки; поверхность последующих колючек гладкая. Колючки D, прижатые назад, не достигают вершинами начала мягкой части D. Первые два мягких луча D неветвистые; в А близ самых вершин разветвляются только четыре последних луча. Нет непарного щитка перед началом D и колючих щитков LL. Р заходят за уровень начала А, достигая основания его четвёртого луча; пятый и шестой соединённые лучи Р наибольшие. Наибольший (верхний) свободный луч Р заметно короче наибольших лучей соединённого Р, слева достигает, справа немного не доходит до вершины V; средний – длиннее нижнего свободного луча Р. V прикрепляются под основанием Р, достигают начала А. Колючка V значительно короче его наибольших (третьего и четвёртого) ветвистых лучей. Анус расположен у начала А. С с неглубокой выемкой, его нижняя лопасть слегка длиннее верхней.
Ктеноидная чешуя на туловище выше уровня клейтрального шипа, а близ конца вертикальных плавников и на хвостовом стебле – выше LL, кроме задней половины хвостового стебля, где чешуя циклоидная, как и на других участках тела ниже вышеуказанного уровня. Циклоидная чешуя более опадающая, чем ктеноидная. На горле, груди и между основаниями V, в основании Р и на небольшом участке позади и над ним, и между основаниями P и V чешуи нет. Предорсальная область покрыта чешуёй (узкий участок непосредственно перед первой колючкой D голый), предорсальных чешуй ~8, не образуют срединного ряда. Канал боковой линии ветвится на каждой чешуйке, с расходящимися ответвлениями (от пяти до семи на чешуях в передней части LL, три – на самых задних чешуях). Вокруг хвостового стебля ~14 чешуй.
Измерения приведены в таблице.
Морфометрические признаки шести видов рода Lepidotrigla из зал. Нячанг и сопредельных вод
| Признак | L. deasoni | L. firmisquamis | L. marisinensis | L. punctipectoralis | L. cf. spiloptera | L. venusta | |
|---|---|---|---|---|---|---|---|
| n = 1 | n = 7 | n = 2 | n = 2 | n = 1 | n = 8 | n = 8 | |
| SL, мм | 204 | 72–125 | 113–130 | 87–92 | 206 | 90–148 | 90–114 |
| В % SL | |||||||
| Длина головы | 33.8 | $\frac{{34.8{\kern 1pt} --{\kern 1pt} 38.9}}{{36.6}}$ | 32.3–33.6 | 32.6–34.5 | 35.0 | $\frac{{33.1{\kern 1pt} --{\kern 1pt} 37.8}}{{35.0}}$ | $\frac{{30.7{\kern 1pt} --{\kern 1pt} 33.3}}{{31.6}}$ |
| Длина рыла | 13.7 | $\frac{{13.4{\kern 1pt} --{\kern 1pt} 17.5}}{{15.8}}$ | 13.8–15.9 | 13.0–13.8 | 17.5 | $\frac{{11.8{\kern 1pt} --{\kern 1pt} 15.5}}{{13.9}}$ | $\frac{{9.6{\kern 1pt} --{\kern 1pt} 11.7}}{{10.8}}$ |
| Горизонтальный диаметр глаза | 8.8 | $\frac{{9.0{\kern 1pt} --{\kern 1pt} 10.4}}{{9.5}}$ | 9.2–9.5 | 9.8–10.1 | 5.3 | $\frac{{8.6{\kern 1pt} --{\kern 1pt} 9.7}}{{9.0}}$ | $\frac{{9.4{\kern 1pt} --{\kern 1pt} 10.4}}{{9.9}}$ |
| Ширина межглазничного промежутка | 7.8 | $\frac{{6.8{\kern 1pt} --{\kern 1pt} 9.0}}{{7.5}}$ | 9.2–9.7 | 7.1–8.0 | 7.3 | $\frac{{6.8{\kern 1pt} --{\kern 1pt} 8.1}}{{7.4}}$ | $\frac{{6.8{\kern 1pt} --{\kern 1pt} 7.2}}{{7.0}}$ |
| Интерростральная ширина | 10.8 | $\frac{{11.9{\kern 1pt} --{\kern 1pt} 14.5(17.4){\kern 1pt} *}}{{13.2}}$ | 15.0–15.4 | 8.7–9.2 | 12.1 | $\frac{{11.3{\kern 1pt} --{\kern 1pt} 13.3}}{{12.4}}$ | $\frac{{9.5{\kern 1pt} --{\kern 1pt} 10.8}}{{9.9}}$ |
| Длина рострального отростка | 2.7 | $\frac{{6.7{\kern 1pt} --{\kern 1pt} 9.7}}{{7.6}}$ | 6.9–7.7 | 3.3–4.0 | 5.8 | $\frac{{4.8{\kern 1pt} --{\kern 1pt} 6.1}}{{5.3}}$ | $\frac{{3.7{\kern 1pt} --{\kern 1pt} 4.5}}{{4.1}}$ |
| Ширина основания рострального отростка | 2.7 | $\frac{{4.5{\kern 1pt} --{\kern 1pt} 4.8(5.2){\kern 1pt} *}}{{4.8}}$ | 4.0–4.6 | 2.7–4.6 | 3.4 | $\frac{{3.6{\kern 1pt} --{\kern 1pt} 4.8}}{{4.1}}$ | $\frac{{3.3{\kern 1pt} --{\kern 1pt} 3.7}}{{3.5}}$ |
| Длина верхней челюсти | 13.2 | $\frac{{11.6{\kern 1pt} --{\kern 1pt} 13.6}}{{12.7}}$ | 11.5–12.0 | 13.0–14.4 | 12.6 | $\frac{{12.8{\kern 1pt} --{\kern 1pt} 13.5}}{{13.0}}$ | $\frac{{12.1{\kern 1pt} --{\kern 1pt} 13.2}}{{12.5}}$ |
| Преорбитальная высота | 14.2 | $\frac{{11.6{\kern 1pt} --{\kern 1pt} 15.2}}{{12.8}}$ | 12.3–12.8 | 10.9–12.1 | 13.1 | $\frac{{12.2{\kern 1pt} --{\kern 1pt} 13.7}}{{12.8}}$ | $\frac{{8.8{\kern 1pt} --{\kern 1pt} 10.8}}{{10.0}}$ |
| Суборбитальная высота | 11.8 | $\frac{{9.7{\kern 1pt} --{\kern 1pt} 12.0}}{{11.3}}$ | 10.0–10.6 | 9.8–10.9 | 12.1 | $\frac{{10.7{\kern 1pt} --{\kern 1pt} 11.5}}{{11.0}}$ | $\frac{{10.1{\kern 1pt} --{\kern 1pt} 11.7}}{{10.8}}$ |
| Длина клейтрального шипа | 9.8 | $\frac{{10.5{\kern 1pt} --{\kern 1pt} 13.2}}{{11.7}}$ | 10.8–13.7 | 8.1–9.2 | 12.1 | $\frac{{9.4{\kern 1pt} --{\kern 1pt} 12.2}}{{11.2}}$ | $\frac{{8.0{\kern 1pt} --{\kern 1pt} 10.5}}{{9.4}}$ |
| Длина посттемпорального шипа | 6.4 | $\frac{{7.1{\kern 1pt} --{\kern 1pt} 8.5(9.4){\kern 1pt} *}}{{7.7}}$ | 6.9–8.0 | 7.6–8.6 | 7.8 | $\frac{{7.4{\kern 1pt} --{\kern 1pt} 8.9}}{{7.9}}$ | $\frac{{6.3{\kern 1pt} --{\kern 1pt} 7.4}}{{6.7}}$ |
| Длина оперкулярного шипа | 3.9 | $\frac{{6.7{\kern 1pt} --{\kern 1pt} 8.4}}{{7.5}}$ | 6.1–6.5 | 5.4–7.5 | 7.8 | $\frac{{7.2{\kern 1pt} --{\kern 1pt} 7.9}}{{7.6}}$ | $\frac{{6.6{\kern 1pt} --{\kern 1pt} 7.2}}{{7.1}}$ |
| Затылочно-предорсальная длина | 5.9 | $\frac{{4.2{\kern 1pt} --{\kern 1pt} 4.9}}{{4.6}}$ | 4.8–5.3 | 5.7–6.0 | 5.1 | $\frac{{4.7{\kern 1pt} --{\kern 1pt} 6.8}}{{5.6}}$ | $\frac{{4.7{\kern 1pt} --{\kern 1pt} 5.7}}{{5.3}}$ |
| Первое предорсальное расстояние | 33.8 | $\frac{{33.0{\kern 1pt} --{\kern 1pt} 38.9}}{{35.6}}$ | 33.1–35.4 | 32.6–34.5 | 33.5 | $\frac{{33.8--35.1}}{{34.4}}$ | $\frac{{29.8{\kern 1pt} --{\kern 1pt} 32.4}}{{31.0}}$ |
| Второе предорсальное расстояние | 52.9 | $\frac{{52.6{\kern 1pt} --{\kern 1pt} 57.0}}{{55.5}}$ | 54.6–54.9 | 56.3–56.5 | 55.3 | $\frac{{52.7{\kern 1pt} --{\kern 1pt} 55.9}}{{54.1}}$ | $\frac{{52.6{\kern 1pt} --{\kern 1pt} 54.1}}{{53.2}}$ |
| Преанальное расстояние | 52.9 | $\frac{{54.5{\kern 1pt} --{\kern 1pt} 59.0}}{{56.5}}$ | 54.6–57.1 | 53.3–58.6 | 53.4 | $\frac{{52.7{\kern 1pt} --{\kern 1pt} 59.1}}{{54.8}}$ | $\frac{{52.6{\kern 1pt} --{\kern 1pt} 53.2}}{{52.8}}$ |
| Превентральное расстояние | 29.4 | $\frac{{28.6{\kern 1pt} --{\kern 1pt} 33.3}}{{31.7}}$ | 27.7–31.0 | 23.9–27.6 | 28.15 | $\frac{{29.0{\kern 1pt} --{\kern 1pt} 30.4(24.4){\kern 1pt} *{\kern 1pt} *}}{{29.6}}$ | $\frac{{25.5{\kern 1pt} --{\kern 1pt} 27.4}}{{26.6}}$ |
| Длина соединённого Р | 29.4 | $\frac{{30.7{\kern 1pt} --{\kern 1pt} 36.0}}{{32.6}}$ | 28.5–31.0 | 32.1–34.5 | 29.1 | $\frac{{28.4{\kern 1pt} --{\kern 1pt} 33.3}}{{30.1}}$ | $\frac{{32.1{\kern 1pt} --{\kern 1pt} 34.8}}{{33.1}}$ |
| Длина первого свободного луча Р | 26.5 | $\frac{{23.2{\kern 1pt} --{\kern 1pt} 25.2}}{{24.3}}$ | 23.9–24.6 | 21.7–25.3 | 25.2 | $\frac{{23.3{\kern 1pt} --{\kern 1pt} 26.9}}{{25.7}}$ | $\frac{{25.5{\kern 1pt} --{\kern 1pt} 27.6}}{{26.6}}$ |
| Длина второго свободного луча Р | 22.1 | $\frac{{18.8{\kern 1pt} --{\kern 1pt} 22.4}}{{20.3}}$ | 19.5–21.5 | 18.5–21.8 | 21.4 | $\frac{{20.1{\kern 1pt} --{\kern 1pt} 22.3}}{{21.4}}$ | $\frac{{21.4{\kern 1pt} --{\kern 1pt} 23.4}}{{22.5}}$ |
| Длина третьего свободного луча Р | 16.2 | $\frac{{13.6{\kern 1pt} --{\kern 1pt} 15.8(19.4){\kern 1pt} *}}{{15.2}}$ | 15.0–16.9 | 15.2–18.4 | 17.0 | $\frac{{15.5{\kern 1pt} --{\kern 1pt} 16.9}}{{16.1}}$ | $\frac{{18.7{\kern 1pt} --{\kern 1pt} 19.8}}{{19.1}}$ |
| Длина V | 25.5 | $\frac{{25.7{\kern 1pt} --{\kern 1pt} 29.0}}{{27.2}}$ | 24.6–26.6 | 23.9–27.6 | 24.8 | $\frac{{25.0{\kern 1pt} --{\kern 1pt} 28.0}}{{26.2}}$ | $\frac{{26.3{\kern 1pt} --{\kern 1pt} 29.5}}{{27.9}}$ |
| Высота первой колючки D | 13.2 | $\frac{{13.2{\kern 1pt} --{\kern 1pt} 17.1}}{{14.9}}$ | 13.8–15.9 | 15.8–17.2 | 17.0 | $\frac{{14.8{\kern 1pt} --{\kern 1pt} 18.9}}{{16.4}}$ | $\frac{{15.8{\kern 1pt} --{\kern 1pt} 18.4}}{{16.8}}$ |
| Высота второй колючки D | 16.9 | $\frac{{16.1{\kern 1pt} --{\kern 1pt} 20.0}}{{17.8}}$ | 15.4–18.1 | 19.6–20.7 | 20.4 | $\frac{{19.6{\kern 1pt} --{\kern 1pt} 21.1}}{{20.5}}$ | $\frac{{17.5{\kern 1pt} --{\kern 1pt} 20.5}}{{19.4}}$ |
| Высота третьей колючки D | 14.2 | $\frac{{16.1{\kern 1pt} --{\kern 1pt} 20.9}}{{18.1}}$ | 15.7–16.8 | 18.5–19.5 | 18.45 | $\frac{{17.0{\kern 1pt} --{\kern 1pt} 20.9}}{{19.2}}$ | $\frac{{16.7{\kern 1pt} --{\kern 1pt} 18.9}}{{18.3}}$ |
| Длина C | 22.1 | $\frac{{20.8{\kern 1pt} --{\kern 1pt} 24.8}}{{22.9}}$ | 20.8–22.1 | 21.7–23.6 | 19.9 | $\frac{{19.4{\kern 1pt} --{\kern 1pt} 23.7}}{{21.4}}$ | $\frac{{22.8{\kern 1pt} --{\kern 1pt} 23.7}}{{23.3}}$ |
| Максимальная высота тела | 23.5 | $\frac{{20.2{\kern 1pt} --{\kern 1pt} 27.4}}{{23.8}}$ | 23.1–23.9 | 21.8–30.7 | 21.8 | $\frac{{21.7{\kern 1pt} --{\kern 1pt} 24.7}}{{23.1}}$ | $\frac{{21.9{\kern 1pt} --{\kern 1pt} 25.3}}{{23.7}}$ |
| Минимальная высота тела | 5.9 | $\frac{{6.3{\kern 1pt} --{\kern 1pt} 7.0}}{{6.7}}$ | 6.7–7.1 | 5.2–5.4 | 5.3 | $\frac{{5.4{\kern 1pt} --{\kern 1pt} 6.5}}{{5.9}}$ | $\frac{{5.4{\kern 1pt} --{\kern 1pt} 6.3}}{{5.8}}$ |
| Длина хвостового стебля | 15.7 | $\frac{{13.2{\kern 1pt} --{\kern 1pt} 16.1}}{{14.8}}$ | 13.1–15.0 | 13.6–17.2 | 16.5 | $\frac{{14.3{\kern 1pt} --{\kern 1pt} 16.9}}{{15.1}}$ | $\frac{{13.5{\kern 1pt} --{\kern 1pt} 15.8}}{{14.5}}$ |
Окраска фиксированных рыб светлая, желтоватая, голые (не покрытые чешуёй) участки кожи ярко-белые; выступающие наружу покровные кости головы темнее кожи; края чешуйных карманов в передней половине туловища (примерно до конца прижатого к телу Р) выше уровня клейтрального шипа, далее назад, выше LL, – тёмные, сероватые. Участки нечёткого тёмного пигмента на межлучевой перепонке между первой и второй, второй и третьей и третьей и четвёртой колючками D, вероятно, при жизни формировали продольную субдистальную полосу (рис. 3б). Мягкие части D, A и V не окрашены.
В средней части верхней и нижней лопасти С прослеживаются очень слабые размытые прерывистые скопления сероватых меланофоров, маркирующих возможную срединную перевязь на С при жизни. Внутренняя сторона Р серо-чёрная, интенсивно зачернена близ оснований пятого–десятого лучей и далее назад между восьмым, девятым и десятым лучами, лишена оформленного светлого рисунка, но с размытым просветлением в проксимальной половине выше участка интенсивно-чёрной пигментации; перепонка между 10-м и 11-м лучами не пигментирована (рис. 3в). Ротожаберная полость светлая.
Сравнительные замечания. Описанный экземпляр хорошо соответствует признакам голотипа L. deasoni, за исключением широкого и полого вогнутого интеррострального промежутка (рис. 2б, 2г), тогда как у голотипа он имеет вид глубокой V-образной вырезки (рис. 2в). Поскольку подобные вариации отмечены в моём материале для L. cf. spiloptera, я не склонен придавать большого значения этому различию. Оно может быть связано с ростом, так как описанная рыба значительно крупнее экземпляров типовой серии (SL 80–97 мм). К тому же, голотип L. deasoni зафиксирован с приподнятыми жаберными крышками, что может искажать пропорции. Главная вершина рострального выступа у голотипа L. deasoni более массивная, чем у вьетнамского экземпляра (рис. 2б, 2в), но это отличие также лежит в пределах изменчивости других видов рода. Изученный экземпляр позволяет добавить к характеристике вида два признака пигментации, которые, к сожалению, не могут быть проверены на голотипе, поскольку последний почти полностью обесцветился. На перепонке колючей части D прослеживается очень неясная продольная субдистальная полоса, которая при жизни, очевидно, была красной. На внутренней стороне P в дорсопроксимальной части прослеживается размытый участок просветления, возможно, видоспецифичный, поскольку такие участки не обнаружены у других видов, после фиксации имеющих внутреннюю сторону Р без рисунка (L. firmisquamis, L. marisinensis, L. venusta). В первоописании внутренняя сторона Р охарактеризована лишь как “большей частью чёрная (largely blackish)” (Herre, Kauffman, 1952. P. 29).
Среди видов, встречающихся за пределами акватории Вьетнама, L. deasoni наиболее близок к L. abyssalis Jordan et Starks, 1904 и L. hime Matsubara et Hiyama, 1932, известными от Южной Японии и из Восточно-Китайского моря (Yamada, 2002), отличаясь от перечисленных видов лишь более короткими ростральными отростками и перепонкой соединённого Р, непигментированной только между двумя самыми нижними лучами (против трёх или четырёх у сравниваемых видов). Кроме того, в отличие от L. abyssalis у описываемого вида меньше чешуй в LL (55–58 против 60–64), а в отличие от L. hime –интерростральный промежуток шире межглазничного, а не наоборот, и верхняя челюсть не достигает переднего края зрачка. Интересно отметить, что для сравниваемых видов также характерна красноватая субдистальная полоса на перепонке колючей части D.
Распространение. Для фауны Вьетнама указывается впервые. Вид был ранее известен только по типовой серии из Манильского залива (о. Лусон, Филиппины).
Lepidotrigla firmisquamis Prokofiev et Yato, 2020
Материал. Типовая серия (Prokofiev, Yato, 2020).
Диагноз. Ростральные выступы длинные треугольные, внутренние дополнительные вершины, если имеются, намного короче главной; заглазничная бороздка глубокая, широко прервана; преорбитальные и посторбитальные шипы имеются, сфенотикальные имеются или отсутствуют, париетальные шипы отсутствуют; P короткий, его внутренняя сторона без светлого рисунка у фиксированных рыб; чешуя крепко сидящая, на груди отсутствует; LL 54–58.
Измерения приведены в таблице.
Замечания. Детальное описание вида опубликовано ранее (Prokofiev, Yato, 2020). От большинства видов из региона, за исключением L. alata и L. marisinensis, он легко отличим по крепко сидящей чешуе и LL–D 4, а от L. alata и L. marisinensis – по наличию развитых пре- и посторбитальных шипов и глубокой, хотя и широко прерванной, заглазничной бороздки.
Распространение. Известен из заливов Нячанг и Халонг, Вьетнам.
Lepidotrigla marisinensis (Fowler, 1938)
(рис. 4)
Рис. 4.
Lepidotrigla marisinensis SL 130 мм, г. Нячанг: а – общий вид, б – ростральные выступы, в – гребни и шипы крыши черепа (не развиты), г – внутренняя сторона Р. Масштаб, мм: б, г – 10; в – 12.

Материал. 2 экз. SL 113 мм (г. Фантьет) и SL 130 мм (г. Нячанг).
Диагноз. Ростральные выступы длинные треугольные, без внутренних дополнительных вершин; заглазничная бороздка почти полностью редуцирована; преорбитальные, посторбитальные, сфенотикальные и париетальные шипы отсутствуют; P короткий, его внутренняя сторона без светлого рисунка у фиксированных рыб; чешуя плотно сидящая, на груди отсутствует; LL 58–62.
Описание (рис. 4а). D IX + 15–16, A 15–16, P 11 + 3, V I + 5, LL 58–60, LL–D 4, LL–A 17–20.
Ростральные выступы длинные, уплощённые, в форме равнобедренного треугольника; ширина их основания явственно меньше длины; вершины заострённые, отчётливо расходящиеся (угол, образуемый продольной осью отростка и срединно-брюшной осью тела, составляет 25°–35°). Ростральный выступ с единственной центральной вершиной, боковые края которой несут мелкие бугорки, такие же как на покровных костях (рис. 4б). Интерростральный промежуток заметно превышает по ширине межглазничный, от полого вогнутого до V-образного с угловатой срединной выемкой; симфизный конец нижней челюсти немного не достигает симфиза praemaxillaria. У экз. SL 113 мм очень слабый продольный гребень прослеживается лишь в задней части щеки, позади вертикали середины орбиты, переходит на praeoperculum, но не образует на конце шипа; у экз. SL 130 мм отсутствует. Надглазничные гребни пологие, валиковидные, параллельные, межглазничный промежуток между ними глубоко вогнут. Преорбитальные и посторбитальные шипы отсутствуют (рис. 4в), у задневерхнего края орбиты имеются очень слабые, маленькие, трудноразличимые вырезки, маркирующие остатки посторбитальной бороздки, таким образом, совершенно не развитой. Сфенотикальный гребень едва заметен, в виде валика, протягивающегося от конца надглазничного гребня до основания посттемпорального, покрыт обычной скульптурой, без каких-либо выделяющихся шипиков. Париетальные гребни и шипы отсутствуют. Оперкулярный и посттемпоральный шипы короткие, на вершинах заострённые; посттемпоральные шипы сверху и снизу несут расширения, за счёт которых приобретают листовидную форму. Оперкулярный шип короче посттемпорального, оканчивается далеко перед вершиной последнего, направлен вершиной назад и немного вверх. Клейтральный шип наиболее длинный, острый, расположен на равном расстоянии от основания оперкулярного шипа и верхнего края основания Р, его вершина оканчивается на вертикали основания пятой колючки D. Кости черепа и плечевого пояса густо скульптированы мелкими бугорками, организованными в ряды; на ветвях нижней челюсти они образуют продольные ряды медиальнее и латеральнее нижнечелюстного канала.
Симфиз верхней челюсти нависает над вершиной нижней, озубление praemaxillaria полностью видно при закрытом рте. Верхняя челюсть оканчивается под передней третью орбиты. В челюстях мелкие бугорковидные зубы полосками; сошник и нёбные кости без зубов. Язык очень массивный, широкий, едва обособленный. Жаберных тычинок на первой дуге в наружном ряду 10–11 (1 + 6–7 + iii), во внутреннем – 7–8; в наружном ряду на 2-й дуге – 9 (0–1 + 6–7 + ii). Есть щель позади четвёртой жаберной дуги. Ложножабра крупная, из 20–25 элементов.
Вдоль оснований обоих спинных плавников парный ряд из 23–24 костных щитков, из которых восемь под основанием колючей части D, один в интердорсальном промежутке, а остальные – под основанием мягкой части D (последний – под его предпоследним или последним лучом). Первые восемь щитков с множественными выступами (первые два слиты и покрыты многочисленными бугорками), начиная с интердорсального, оканчиваются одним острым и сильным шипом. Первые четыре колючки D заметно толще и массивней остальных (особенно первая и вторая); вторая или вторая и третья колючки – наибольшие, но лишь в 1.1 раза длиннее первой; последующие колючки постепенно уменьшаются по длине. Передняя и переднебоковые поверхности первых двух и передняя поверхность третьей колючки D мелко зазубрены; поверхность последующих колючек гладкая. Колючки D, прижатые назад, далеко не достигают вершинами начала мягкой части D. Первые два мягких луча D неветвистые; в А близ самых вершин разветвляются четыре–пять последних лучей. Нет непарного щитка перед началом D. P заходят за начало А, достигая вертикали основания его четвёртого луча; четвёртый–шестой соединённые лучи Р наибольшие. Наибольший (верхний) свободный луч Р заметно короче наибольших лучей соединённого Р, достигает или не достигает вершины V (но в последнем случае всё же достигает ануса); средний – длиннее нижнего свободного луча Р. V прикрепляется под основанием Р, достигает или едва заходит за начало А. Колючка V значительно короче наибольших ветвистых лучей, самые длинные ветвистые лучи этого плавника – третий и четвёртый. Анус расположен вблизи начала А. C слабовыемчатый, его лопасти равной длины.
Чешуя ктеноидная, но ктении слабо выражены на чешуях вентральной стороны туловища, совсем пропадают на чешуях на участке позади оснований Р и V. Чешуя повсеместно крепко сидящая. Горло, грудь, участки между и перед основаниями Р и V и позади оснований V голые; брюхо покрыто чешуёй. Предорсальная область покрыта чешуёй (очень узкий участок непосредственно перед D голый), предорсальных чешуй шесть или семь. Канал боковой линии ветвится на каждой чешуйке, с расходящимися ответвлениями (до восьми на чешуях в передней половине туловища, от четырёх до двух на самых задних чешуях). Вокруг хвостового стебля 21 чешуя.
Измерения приведены в таблице.
Окраска фиксированных рыб довольно тёмная, коричневато-жёлтая, покровные кости окрашены практически одинаково с телом, голые (не покрытые чешуёй) участки кожи – ярко-белые. Плавники, кроме соединённой части Р, светлые. Внутренняя сторона Р серо-чёрная, более интенсивно-чёрная в нижней половине, без рисунка; перепонка между двумя нижними соединёнными лучами (у меньшего экземпляра – также в проксимальных двух третях между девятым и десятым лучами) светлая (рис. 4г). Ротожаберная полость светлая.
Сравнительные замечания. Этот вид очень близок к L. alata, известному от Японии до Южно-Китайского моря, и отличия между ними нуждаются в дальнейшем уточнении, хотя и представляются реальными. Они суммированы в нижеприведённом определительном ключе.
Распространение. Для фауны Вьетнама отмечен впервые. Ранее был известен по голотипу, пойманному в Южно-Китайском море в районе 21°42′ с.ш. и 114°50′ в.д. (Fowler, 1938).
Lepidotrigla punctipectoralis Fowler, 1938
Рис. 5.
Lepidotrigla punctipectoralis SL 206 мм (а, в, г) и SL 87 мм (б): а, б – общий вид; в – ростральные выступы, г – внутренняя сторона Р. Масштаб, мм: в – 12, г – 15.

Рис. 6.
Lepidotrigla punctipectoralis SL 87 мм (а) и SL 206 мм (б, в), вооружение головы. Вид: а, б – сверху; в – сбоку; mpos – сплошная посторбитальная бороздка, pc – париетальные гребни. Масштаб, мм: в – 12, г – 15.
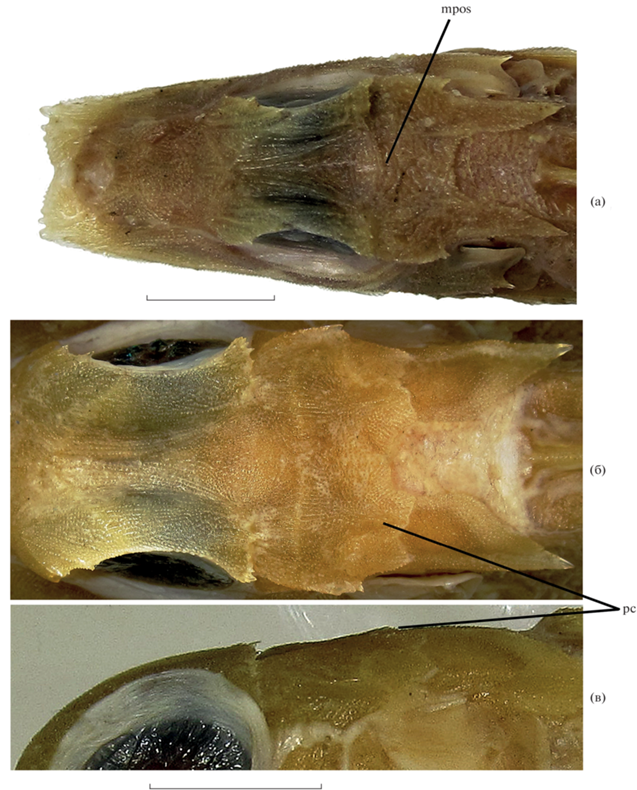
Материал. 3 экз. SL 87–206 мм, г. Нячанг.
Диагноз. Ростральные выступы от конических у молоди до длинных треугольных у взрослых рыб, с дополнительными вершинами кнутри от главной; заглазничная бороздка широко прерванная (у молоди сохраняются следы сплошной бороздки); преорбитальные, посторбитальные и сфенотикальные шипы имеются, париетальные отсутствуют; P короткий, его внутренняя сторона у фиксированных рыб с диагональными пятнышками, не заходящими за границу интенсивно-чёрной пигментации; чешуя легко опадающая, на груди отсутствует; LL 59–64.
Описание (рис. 5а, 5б). D IX–X + 16, A 15–17, P 11 + 3, V I + 5, LL 59–64, LL–D 3, LL–A 12–14.
Ростральные выступы умеренно длинные, на вершинах заострённые, вершины расходящиеся; угол, образуемый продольной осью отростка и срединно-брюшной осью тела, составляет ~30°. У экз. SL 206 мм ростральные выступы по форме близки к прямоугольному треугольнику, ширина их основания заметно меньше длины (рис. 5в). У молоди SL 87 и 92 мм ростральные выступы более широкие и короткие, ширина их основания едва больше или едва меньше длины (рис. 6а). Внешний боковой край рострального выступа спрямлён, в дистальной половине гладкий, в проксимальной несёт ряд зубчиков, несколько более крупных, чем бугорки скульптуры покровных костей черепа. Главная вершина рострального выступа у экземпляра SL 206 мм в виде сильного уплощённого шипа с гладкими в дистальной половине краями, вовнутрь от неё расположено три зубчика гораздо меньшего размера, прогрессивно уменьшающихся по величине от ближайшего к главной вершине к наиболее дальнему от неё, за последним следует ряд из более мелких зубчиков, переходящих в бугорки краевой скульптуры предглазничной кости (рис. 5в). У молоди главная вершина лишь немногим длиннее следующего зубца, дополнительные зубцы прогрессивно уменьшаются в размере от наружных к внутренним (рис. 6а). Интерростральный промежуток заметно превышает межглазничный. Передний край интеррострального промежутка вогнутый. На щеке имеется слабый продольный гребень, начинающийся на уровне вертикали переднего края орбиты, переходящий на praeoperculum и оканчивающийся небольшим преоперкулярным шипом (острым с левой стороны и притуплённым с правой). У молоди этот гребень в суборбитальной области отчётливо вогнут книзу. Надглазничные гребни слабо обособлены, параллельны, межглазничный промежуток между ними глубоко вогнут. Преорбитальных шипов два (рис. 6б), небольшие, но отчётливые, их выступающие концы соизмеримы по величине, острые, расставленные. У экземпляра SL 206 мм верхний посторбитальный шип маленький, но отчётливый, направлен назад и немного вбок. У двух меньших экземпляров верхний посторбитальный шип не выражен. Нижний посторбитальный шип у экз. SL 92 мм отчётливый, у двух других образует массивный треугольный выступ заднебокового конца надглазничного гребня, несущий мелкие шипики, которые слабо различаются по величине и лишь немногим превышают размер бугорков скульптуры костей (но у экз. SL 206 мм с правой стороны один из бугорков увеличен). Сфенотикальный шип хорошо выражен, с сильным шельфовидным боковым гребнем. Париетальные гребни слабые, короткие, зазубренные; париетальные шипы не развиты (рис. 6б, 6в). Посторбитальная бороздка глубокая, но короткая, у экземпляра SL 206 мм не заходит за медиальную границу надглазничного гребня (рис. 6б). У молоди глубокие вырезки у задних концов надглазничных гребней соединяются очень слабой поперечной бороздкой (рис. 6а). Оперкулярный и посттемпоральный шипы хорошо выражены, острые, направлены назад, оперкулярный короче посттемпорального у молоди и равен ему у экземпляра SL 206 мм, его вершина не достигает вертикали вершины посттемпорального шипа. Клейтральный шип хорошо развит, острый, заметно длиннее остальных шипов, расположен вентрально от оперкулярного шипа, его вершина оканчивается на вертикали основания четвёртой–шестой колючки D. Кости черепа и плечевого пояса густо усеяны мелкими бугорками, организованными в ряды; на ветвях нижней челюсти скульптура слабо выражена у рыб SL 87 и 92 мм.
Челюсти равной длины. Верхняя челюсть оканчивается на вертикали переднего края орбиты. У экземпляров SL 92 и 206 мм симфиз praemaxillaria отчётливо вогнут между передними концами зубных рядов соседних сторон, в эту выемку входит вершина симфиза dentalia (у экз. SL 87 мм нижнечелюстной симфиз оканчивается перед этой выемкой). В челюстях мелкие бугорковидные зубы полосками; озубление praemaxillaria полностью видно при закрытом рте. Сошник и нёбные кости без зубов. Язык очень массивный, широкий, едва обособленный. Жаберных тычинок на 1-й дуге в наружном ряду 10–12 (0–1 + 6–7 + iii–v) (7 развитых + 3–5 бугорковидных; кроме них между тычинками на ceratobranchiale с первой по шестую имеется по плоскому бугорку), во внутреннем – 8 (7 + i); в наружном ряду на 2-й дуге – 9–10 (9 + i или i + 8). Есть щель позади четвёртой жаберной дуги. Ложножабра крупная, из 24–32 элементов (их число увеличивается с ростом рыб).
Вдоль основания D парный ряд из 24–25 костных щитков, из которых восемь или девять под основанием колючей части D, один в интердорсальном промежутке, а остальные – под основанием мягкой части D (последний – под его последним или предпоследним лучом). Два передних щитка покрыты бугорками, на трёх последующих переходящих во множественные шипы; последующие щитки оканчиваются одним острым и сильным шипом. Первые три колючки D заметно длиннее, толще и массивней остальных; вторая колючка наибольшая, в 1.1 раза длиннее третьей и в 1.2 раза длиннее первой; последующие колючки постепенно уменьшаются по длине. Передняя и переднебоковые поверхности первой, второй и дистальной трети третьей (у молоди – только первой и второй) колючек D мелко зазубрены; поверхность последующих колючек гладкая. Колючки D, прижатые назад, достигают вершинами начала мягкой части этого плавника, но не заходят за него. Первые два мягких луча D неветвистые; в А близ вершин разветвляются только четыре последних луча. Нет непарного щитка перед первой колючкой D. P заходят за вертикаль начала А, достигая основания его четвёртого–шестого луча; пятый и шестой лучи соединённого Р наибольшие. Наибольший (верхний) свободный луч Р заметно короче наибольших лучей соединённого Р, почти достигает своим концом вершины брюшных плавников; средний длиннее нижнего свободного луча Р. V прикрепляется под основанием Р, достигает начала А. Колючка V значительно короче его наибольших ветвистых лучей, второй–четвёртый или третий и четвёртый ветвистые лучи самые длинные. Анус расположен у начала А. С с неглубокой выемкой; его лопасти равной длины.
Чешуя на туловище ктеноидная выше уровня клейтрального шипа, а близ конца вертикальных плавников и на хвостовом стебле – выше LL, на более низких участках и на дорсальной поверхности задней половины хвостового стебля переходит в циклоидную. Циклоидная чешуя сильнее опадающая, чем ктеноидная. На горле, груди и между основаниями V, в основании Р и на небольшом участке над ним и позади него, а также между основаниями Р и V чешуи нет. Предорсальная область покрыта чешуёй (участок непосредственно перед началом D голый), предорсальных чешуй от 5 (SL 206 мм) до ~8 (SL 87–92 мм). Канал LL ветвится на каждой чешуйке, с расходящимися ответвлениями (обычно четыре или пять на передних чешуях, к хвостовому стеблю их число снижается до трёх). Вокруг хвостового стебля 14 чешуй.
Измерения приведены в таблице.
Окраска фиксированных рыб светлая, желтоватая, голые (не покрытые чешуёй) участки кожи ярко-белые; выступающие наружу покровные кости головы и плечевого пояса более тёмные; края чешуйных карманов в передней половине туловища (примерно до конца прижатого к телу Р) выше уровня клейтрального шипа, далее назад – выше LL, тёмные, сероватые. Плавники, кроме Р и С, не окрашены. У экз. SL 206 мм посередине С прослеживаются размытые скопления сероватых меланофоров, вероятно, образовывавшие сплошную срединную перевязь. У экз. SL 92 мм помимо срединной прослеживается поперечная перевязь в основании С, на обеих перевязях пигментация заметно сильнее выражена на дорсальной лопасти. У экз. SL 87 мм такая пигментация распределена по всей дистальной части С. Внутренняя сторона Р серо-чёрная, с интенсивно-чёрным диагонально вытянутым пятном между четвёртым и девятым лучами; перепонка между 9-м и 11-м лучами белая. На интенсивно-чёрном участке пигментации имеются немногочисленные размытые (у молоди более резкие и чёткие) белые пятнышки, не заходящие за границы пятна, организованные в нечёткие диагональные ряды (рис. 5г). Ротожаберная полость светлая.
Сравнительные замечания. Этот вид отличается от других западнотихоокеанских представителей рода, имеющих короткие Р и не покрытую чешуёй превентральную область, за исключением L. kishinouyei Snyder, 1911 и L. spiloptera Günther, 1880, наличием у фиксированных рыб белых пятен на внутренней стороне Р. От L. kishinouyei он легко отличим длинным верхним свободным лучом Р, который у L. kishinouyei далеко не достигает вершины V. В отличие от L. spiloptera у описываемого вида белые пятна более редкие и ограничены в своём распространении только участком интенсивно-чёрной пигментации. Эта характерная особенность пигментации внутренней стороны Р была подмечена уже в первоописании (Fowler, 1938). Помимо этого L. punctipectoralis отличается от L. spiloptera отсутствием париетальных шипов и, возможно, несколько более мелкой чешуёй (LL 59–64 против 57–61). В литературе оба вида, по-видимому, часто смешиваются (например: Shen et al., 1993. Pl. 63. Fig. 8).
Среди видов, встречающихся за пределами акватории Вьетнама, белые пятна на участке интенсивно-чёрной пигментации внутренней стороны Р у фиксированных рыб присутствуют ещё только у L. faurei Gilchrist et Thompson, 1914 из западной части Индийского океана, но этот вид существенно отличается от L. punctipectoralis по форме ростральных отростков, имеющих вид закруглённых ошиплённых выступов без главной вершины (Richards, Saksena, 1977).
Распространение. Описан из вод Филиппин (пролив Сулу и между о-вами Самар и Масбате). Во Вьетнаме указывался для Тонкинского залива (Nguyen, 1999). Также был отмечен в водах Южной Японии (Ochiai, Yatou, 1985; Yamada, 2002), Тайваня (Shao et al., 2008) и в Андаманском море у берегов Юго-Западного Таиланда (Satapoomin, 2011). Указания не сопровождаются морфологическим описанием, либо приводимые иллюстрации в действительности изображают другой вид (Shen et al., 1993; Yamada, 2002). Вероятно, вид является эндемиком Южно-Китайского моря, внутренних морей Филиппин и прилегающих акваторий.
Lepidotrigla cf. spiloptera Günther, 1880
Рис. 7.
Lepidotrigla cf. spiloptera SL 140 мм (а, в, д), SL 138 мм (г) и SL 97 мм (б): а, б – общий вид; в, г – варианты строения ростральных выступов; д – внутренняя сторона Р. Масштаб: 10 мм.

Рис. 8.
Lepidotrigla cf. spiloptera SL 140 мм, гребни и шипы крыши черепа. Вид: а – сверху, б – сбоку; ps – париетальный шип. Масштаб: 12 мм, линейка общая.
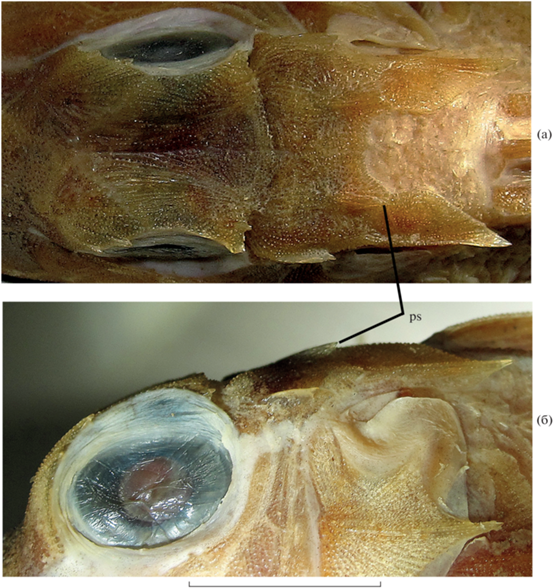
Материал. 12 экз. SL ~80–148 мм, г. Нячанг.
Диагноз. Ростральные выступы длинные треугольные, внутренние дополнительные вершины короткие или отсутствуют; заглазничная бороздка сплошная; преорбитальные, посторбитальные, сфенотикальные и париетальные шипы хорошо выражены; P короткий, его внутренняя сторона у фиксированных рыб с мелкими чётко очерченными светлыми пятнышками по всей поверхности; чешуя легко опадающая, на груди отсутствует; LL 57–61.
Описание (рис. 7а, 7б). D IX + 15–16, A 15–16, P 11 + 3, V I + 5, LL 57–61, LL–D 2½–3, LL–A 16–18.
Ростральные выступы умеренно длинные, треугольные; ширина их основания явственно меньше длины; вершины заострённые, расходящиеся (угол, образуемый продольной осью отростка и срединно-брюшной осью тела, составляет ~30°). Внешние края ростральных выступов практически прямые, в проксимальной половине мелко зазубрены или внутренний край несёт серию мелких зубцов (до десятка), постепенно уменьшающихся по величине от самого наружного к наиболее внутреннему; в дистальной половине ростральный выступ явственно резче сужается к вершине, не так как в проксимальной, и его края гладкие (рис. 7в, 7г). Интерростральный промежуток заметно превышает межглазничный. Передний край интеррострального промежутка с треугольной выемкой посередине. На щеке имеется слабый продольный гребень, начинающийся на уровне конца передней трети орбиты, переходящий на praeoperculum и оканчивающийся небольшим, но отчётливым преоперкулярным шипом. Иногда в месте перехода суборбитального гребня с infraorbitalia на praeoperculum (на уровне задней трети или заднего края орбиты) также имеется выступ или крошечный шип, но чаще этот гребень ровный. Надглазничные гребни слабо обособлены, параллельны, межглазничный промежуток между ними глубоко вогнут. Преорбитальных шипов два, хорошо выражены, их выступающие концы соизмеримы по величине или задний несколько крупнее, острые, расставленные. Позади задневерхнего края орбиты надглазничный гребень образует треугольную площадку между верхним и нижним посторбитальными шипами; последние небольшие, но отчётливые. Верхний шип направлен назад, а нижний вбок или вбок и вниз, редко вбок и назад (или его вершина отогнута назад); обычно нижний шип несколько крупнее верхнего, реже они равны. Вершины посторбитальных шипов варьируют от заострённых до тупых и широких, иногда они замещены группой мелких шипиков или нижний шип разделён на две вершины, иногда с дополнительными очень мелкими шипиками в их основаниях. Встречаются экземпляры, у которых по всему заднему краю треугольника, образованного верхним и нижним шипами, расположены многочисленные шипики. Сфенотикальный шип хорошо развит, иногда с дополнительным выступом перед вершиной, с сильным гребнем. Париетальные шипы отчётливо выражены, с хорошо развитым гребнем (рис. 8). Посторбитальная бороздка сплошная, глубокая до медиального края надглазничного гребня, далее вовнутрь снижается, но всегда заметна (рис. 8а). Оперкулярный и посттемпоральный шипы хорошо выражены, острые, соизмеримые по величине. Оперкулярный шип направлен вершиной назад или назад и немного вверх. Вершина оперкулярного шипа расположена более или менее позади вертикали вершины посттемпорального шипа. Клейтральный шип хорошо развит, острый, заметно длиннее остальных шипов, расположен вентрально от оперкулярного шипа, его вершина оканчивается на вертикали основания пятой колючки D. Покровные кости черепа и плечевого пояса густо усеяны мелкими бугорками, организованными в ряды; скульптура на ветвях нижней челюсти слабо развита, а у рыб SL > 100 мм может совершенно отсутствовать.
Челюсти равной длины. Верхняя челюсть оканчивается на вертикали переднего края орбиты. Симфиз praemaxillaria с выемкой или без, в последнем случае вершина симфиза dentalia не видна сверху. В челюстях мелкие бугорковидные зубы полосками; озубление praemaxillaria полностью видно при закрытом рте. Сошник и нёбные кости без зубов. Язык очень массивный, широкий, едва обособленный. Жаберных тычинок на 1-й дуге в наружном ряду 9–11 (0–i + 6–7 + iii), во внутреннем – 7–9 (7–8 + 0–ii); в наружном ряду на 2-й дуге – 8–11 (8–9 + 0–ii). Есть щель позади четвёртой жаберной дуги. Ложножабра крупная, из 23–35 элементов (их число увеличивается с ростом рыб).
Вдоль основания D парный ряд из 23–24 костных щитков, восемь из которых под основанием колючей части D, один в интердорсальном промежутке, а остальные – под основанием мягкой части D (последний – под его последним лучом). От двух до четырёх (чаще три) самых передних щитков с множественными шипами, последующие оканчиваются одним острым и сильным шипом. Первые три колючки D значительно длиннее, толще и массивней остальных, первая и вторая – наиболее толстые, вторая – самая длинная, в 1.00–1.15 раза длиннее третьей (как исключение – равна третьей) и в 1.30–1.35 раза длиннее первой (при SL > 130 мм – в (1.1)1.2–1.3 раза); последующие колючки постепенно уменьшаются по длине. Передняя и переднебоковые поверхности первых трёх колючек D мелко зазубрены; поверхность последующих колючек гладкая. Колючки D, прижатые назад, достигают вершинами начала мягкой части D или едва заходят за него. Первые два (очень редко – один) луча мягкой части D неветвистые; в А близ самых вершин разветвляются последние пять лучей. Нет непарного щитка перед началом D. Р заходят за начало А, достигая основания его четвёртого–шестого луча; пятый–седьмой соединённые лучи Р наибольшие. Наибольший (верхний) свободный луч Р заметно короче наибольших лучей соединённого Р, достигает вершины прижатых к брюху V; средний – длиннее нижнего свободного луча Р. V прикрепляются под основанием Р, достигают начала А, реже заходят за него (до уровня второго луча). Колючка V значительно короче наибольших ветвистых лучей; третий–четвёртый, реже второй–четвёртый ветвистые лучи этого плавника самые длинные. Анус расположен у начала А. С с неглубокой выемкой, его лопасти равной длины.
Чешуя на туловище ктеноидная от уровня клейтрального шипа, а позади вертикали середины мягкой части D и А – выше LL (но на хвостовом стебле, по-видимому, циклоидная); на более нижних участках – циклоидная. Чешуя сравнительно легко опадающая, особенно циклоидная. На горле, груди и между основаниями V, в основании Р и на небольшом участке над и позади него, а также между основаниями Р и V чешуи нет. Предорсальная область покрыта чешуёй (участок непосредственно перед первой колючкой D голый), от четырёх до шести предорсальных чешуй. Канал боковой линии ветвится на каждой чешуйке, с расходящимися ответвлениями (от трёх до пяти, на чешуях в передней части LL ответвлений больше). Вокруг хвостового стебля 14–16 (обычно 16) чешуй.
Измерения приведены в таблице.
Окраска фиксированных рыб светлая, желтоватая, голые (не покрытые чешуёй) участки кожи – ярко-белые; выступающие наружу покровные кости, напротив, гораздо темнее; края чешуйных карманов в передней половине туловища (примерно до конца прижатого к телу Р) выше уровня клейтрального шипа, далее назад – выше LL, тёмные, сероватые. Скопления сероватых меланофоров на С очень слабо развиты, не образуют сколь-либо заметной поперечной перевязи. Внутренняя сторона Р чёрно-серая, интенсивно-чёрная между четвёртым и девятым–десятым лучами, с мелкими чётко-очерченными белыми пятнышками, распределёнными по всей поверхности (их густота несколько варьирует индивидуально) (рис. 7д). Остальные плавники не окрашены. Ротожаберная полость светлая.
Сравнительные замечания. Описанные рыбы характеризуются специфической пигментацией внутренней стороны Р, которая сплошь покрыта равномерно распределёнными мелкими чётко очерченными белыми пятнышками, хорошо сохраняющимися у фиксированных рыб. Среди всех видов рода подобный рисунок отмечен только для двух: L. spiloptera Günther, 1880 и L. macrobrachium Fowler, 1938. Последний вид резко отличается очень длинным Р, соединённая часть которого заходит за конец основания А, а наибольший свободный луч почти достигает середины длины этого основания. Сильно приподнятые надглазничные гребни у голотипа L. macrobrachium, отображённые на рисунке первоописания (Fowler, 1938. Fig. 52 ), по-видимому, не имеют диагностического значения и связаны с его ювенильной природой, поскольку “длина” голотипа составляет всего 64 мм.
Хотя в литературе имеется много указаний на нахождения L. spiloptera в различных районах Индо-Вест-Пацифики, как показано Гомоном и Каваи (Gomon, Kawai, 2018), достоверно к этому виду можно отнести лишь находки в северо-восточной части Индийского океана от Малых Зондских о-вов до Северо-Западной Австралии. Указанные авторы дали переописание этого вида на основании голотипа и серии дополнительных экземпляров. Изученный вьетнамский материал в целом хорошо соответствует признакам, описанным Гомоном и Каваи, за исключением формы ростральных отростков и, как правило, более глубокой в срединной своей части посторбитальной борозды, которая у индо-австралийских рыб совсем слабая (Gomon, Kawai, 2018. Fig. 14B ). Ростральные отростки у индо-австралийских экземпляров варьируют от коротких треугольных до конических (Prokofiev, Yato, 2020), их внутренние дополнительные зубцы немногим короче главной вершины, тогда как у вьетнамских рыб они длинные треугольные, а внутренние дополнительные зубцы, если и развиты, то значительно короче главной вершины. При этом форма ростральных отростков у вьетнамских рыб весьма постоянна и не подвержена возрастной изменчивости, по крайней мере, при SL > 80 мм. Учитывая, что форма ростральных выступов является важным диагностическим признаком в пределах рода, конспецифичность вьетнамских и индо-австралийских рыб вызывает сомнения. В то же время я пока воздерживаюсь от выделения нового вида, учитывая значительное сходство вьетнамских рыб и “типичных” L. spiloptera по большинству других признаков. Необходимо отметить, что указываемый для вод Южной Японии под названием “L. punctipectoralis” вид в действительности также, вероятно, относится к комплексу “L. spiloptera”, поскольку для него также характерны равномерно рассеянные по внутренней поверхности Р светлые пятна (Yamada, 2002). Ростральные отростки у японских рыб изображены в виде широких треугольных выступов, отдельные вершины в которых слиты между собой (Yamada, 2002). Этим они больше походят на вьетнамских, чем на индо-австралийских рыб. Рыбы от берегов Тайваня, приводимые под названием “L. punctipectoralis”, судя по опубликованным фотографиям (Shen et al., 1993. Pl. 63. Figs. 7, 8), конспецифичны L. cf. spiloptera из вод Вьетнама. По всей видимости, L. spiloptera представляет собой комплекс близких видов, нуждающийся в детальной ревизии.
Распространение. Вид описан из моря Банда (у о-вов Кай) (Günther, 1880), в последующем указывался для Вьетнама (Тонкинский залив) (Nguyen, 1999), Филиппин (Herre, Kauffman, 1952), Юго-Западного Таиланда (Satapoomin, 2011) и Южной Индии от побережья Мадраса – “Madras Coast” (Menon, Rama Rao, 1963. P. 49), вод к югу от о-вов Бали и Ломбок и у северо-западных берегов Австралии (арх. Дампир) (Gomon, Kawai, 2018). Находки за пределами индоокеанских вод Восточной Индонезии и Северо-Западной Австралии, вероятнее всего, относятся к особым видам комплекса “L. spiloptera” или представляют собой ошибочные определения других видов.
Lepidotrigla venusta Fowler, 1938
Рис. 9.
Lepidotrigla venusta SL 114 мм (б – SL 106 мм): а – общий вид; б, в – варианты строения ростральных выступов; г – чешуйный покров превентральной области, д – внутренняя сторона Р. Масштаб, мм: б–г – 5, д – 9.5.

Рис. 10.
Lepidotrigla venusta SL 114 мм, гребни и шипы крыши черепа. Вид: а – сверху, б – дорсолатерально. Обозначения см. на рис. 2. Масштаб: 11 мм.

Материал. 10 экз. SL 90–114 мм, г. Нячанг.
Диагноз. Ростральные выступы конические (Prokofiev, Yato, 2020. Fig. 1C), с хорошо развитыми внутренними дополнительными вершинами; заглазничная бороздка широко прервана; преорбитальные, посторбитальные и сфенотикальные шипы имеются, париетальные отсутствуют; P короткий, его внутренняя сторона без светлого рисунка у фиксированных рыб; чешуя легко опадающая, хорошо развита на груди; LL 52–60.
Описание (рис. 9а). D IX + 15, A 15, P 11 + 3, V I + 5, LL 56–60, LL–D (2) 2½, LL–A 9–11.
Ростральные выступы умеренно длинные, треугольные; ширина их основания меньше длины; вершины уплощённые, заострённые, слабо расходящиеся (угол, образуемый продольной осью отростка и срединно-брюшной осью тела, не превышает 15°). Наружный край рострального выступа практически прямой, в проксимальной половине–трети своей длины несёт мелкие зазубрины, соизмеримые или лишь слегка превышающие по величине бугорки скульптуры покровных костей черепа; внутренний край сильно скошен примерно от половины длины выступа, с одним–тремя отчётливыми, постепенно уменьшающимися дополнительными зубцами, медиальнее которых располагаются ещё два или три более мелких зубца или серия зубчиков, соизмеримых по величине с бугорками скульптуры покровных костей черепа. Самый наружный дополнительный зубец (первый от главной вершины) составляет от двух третей до трети длины главной вершины (чаще всего не более половины этой длины), внутренний край главной вершины в дистальной половине может иметь небольшое боковое расширение или выступ (рис. 9б, 9в). Интерростральный промежуток превышает межглазничный. Передний край интеррострального промежутка с более или менее отчётливой выемкой посередине, симфизный конец нижней челюсти при взгляде сверху в ней не виден. На щеке имеется очень слабый продольный гребень, начинающийся примерно на вертикали середины орбиты, переходящий на praeoperculum и оканчивающийся небольшим выступом, редко – маленьким шипом. Надглазничные гребни слабо обособлены, параллельны, межглазничный промежуток между ними глубоко вогнут. Преорбитальных шипов два, оба хорошо развиты, острые, расставленные, их выступающие концы соизмеримы по величине. Посторбитальные шипы образуют треугольную площадку позади задневерхнего края орбиты, внутренний (верхний) и наружный (нижний) выступы которой могут иметь вид небольших шипов или группы шипиков. Сфенотикальный гребень относительно короткий, но отчётливо шельфовидный, зазубренный, сзади оканчивается несколькими более крупными шипиками, реже – отчётливым терминальным шипом. Париетальные шипы отсутствуют, гребни едва намечены. Посторбитальная бороздка глубокая до медиального края надглазничного гребня или даже не доходит до него, затем резко сглаживается и становится едва заметной, но, тем не менее, сплошная. Оперкулярный и посттемпоральный шипы небольшие, острые, соизмеримые по величине (оперкулярный шип может быть едва короче). Оперкулярный шип направлен вершиной назад или назад и немного вверх. Вершина оперкулярного шипа расположена на одной вертикали с вершиной посттемпорального шипа или немного позади неё. Клейтральный шип относительно небольшой, острый, расположен вентрально от оперкулярного шипа, его вершина оканчивается на вертикали основания третьей колючки D (рис. 10). Покровные кости черепа и плечевого пояса густо усеяны мелкими бугорками, организованными в ряды. Скульптура ветвей нижней челюсти даже у самых крупных рыб представлена всего одним–двумя рядами бугорков.
Нижняя челюсть чаще немного короче верхней, реже челюсти равной длины. Верхняя челюсть оканчивается на уровне передней трети–середины орбиты. Симфиз praemaxillaria без выемки, вершина симфиза dentalia не видна сверху. В челюстях мелкие бугорковидные зубы полосками; озубление praemaxillaria полностью видно при закрытом рте. Сошник и нёбные кости без зубов. Язык очень массивный, широкий, едва обособленный. Жаберных тычинок на первой дуге в наружном ряду 8–10 (пять или шесть развитых), во внутреннем – 7; в наружном ряду на второй дуге – 9–10 (6–7 + ii–iii). Есть щель позади четвёртой жаберной дуги. Ложножабра крупная, из 18–23 элементов.
Вдоль основания D парный ряд из 23 костных щитков, восемь из которых под основанием колючей части D, один в интердорсальном промежутке, а остальные – под основанием мягкой части D (последний – под его терминальным лучом). Один или два (как исключение, три) самых передних щитка с несколькими выступами, последующие оканчиваются одним острым и сильным шипом (иногда все субдорсальные щитки оканчиваются единственным шипом). Первые три колючки D наибольшие и значительно толще и массивней остальных, из них самые толстые первая и вторая, наиболее длинная – вторая колючка, в 1.1 раза длиннее третьей и в 1.1–1.2 раза длиннее первой; последующие колючки постепенно уменьшаются по длине. Передняя и переднебоковые поверхности первой, второй и базальной половины третьей колючек D мелко зазубрены; поверхность последующих колючек гладкая. Колючки D, прижатые назад, явственно не достигают вершинами начала мягкой части D. Первые два мягких луча D неветвистые; в А близ самых вершин разветвляются пять последних лучей. Нет непарного щитка перед началом D. Р заходят за вертикаль начала А, достигая основания его пятого–шестого луча; четвёртый–шестой, реже только пятый и шестой соединённые лучи Р наибольшие. Наибольший (верхний) свободный луч Р заметно короче наибольших лучей соединённого Р, достигает или немного не достигает вершины прижатых к брюху V; средний длиннее нижнего свободного луча Р. V прикрепляются под основанием Р, достигают начала А. Колючка V значительно короче наибольших (третьего и четвёртого) ветвистых лучей. Анус расположен у начала А. С отчётливо вырезан, верхняя лопасть чаще слегка длиннее нижней или обе лопасти равной длины.
Чешуя легко опадающая, на туловище ктеноидная от уровня клейтрального шипа, а позади вертикали середины мягкой части D и А – выше LL (но на хвостовом стебле, по-видимому, циклоидная); на более нижних участках – циклоидная. Как циклоидная, так и ктеноидная чешуя легко опадает и частично утрачена у коллекционных экземпляров (обычно сохраняется лишь в передней части туловища (до уровня передней трети А) ниже LL). Чешуи LL, напротив, крепко сидящие. Грудь сплошь покрыта чешуёй (рис. 9г), голыми остаются лишь горло (вершина истмуса, но в некоторых случаях чешуйный покров доходит практически до вершины), участок между и перед основаниями свободных лучей Р и основанием V, внутренняя сторона основания Р и участок непосредственно позади основания V. Обычно голые участки позади оснований V соседних сторон разделены сплошной срединной полоской чешуек, соединяющей очешуенные участки груди и брюха, но изредка эта чешуйная полоска прерывается, и голый участок позади оснований V сплошной. Предорсальная область покрыта чешуёй (кроме голого участка непосредственно перед первой колючкой D), предорсальных чешуй от трёх до шести, обычно пять. Канал LL ветвится на каждой чешуйке, с расходящимися ответвлениями (обычно четыре, редко три, как исключение – пять на чешуях в передних двух третях LL и три на последующих чешуях). Чешуй вокруг хвостового стебля 13–15 (обычно 14).
Измерения приведены в таблице.
Окраска фиксированных рыб светлая, желтовато-розоватая, голые (не покрытые чешуёй) участки кожи – ярко-белые; выступающие наружу покровные кости головы и плечевого пояса, напротив, гораздо темнее основного фона; верхние две трети туловища сплошь покрыты тёмными меланофорами, хорошо заметными под бинокуляром, расположенными как на чешуйных карманах, так и подкожно. Плавники, кроме соединённой части Р, не окрашены, но при увеличении на лучах мягкой части спинного, хвостового и иногда на свободных лучах грудного плавника может быть различима мелкая меланофорная пигментация. Перепонка колючей части D не пигментирована. Внутренняя сторона Р серо-чёрная, более интенсивно-чёрная в нижней половине, без рисунка; перепонка между первым, вторым и дистальной частью третьего и между 9-м и 11-м лучами светлая (рис. 9д). Ротожаберная полость светлая.
Сравнительные замечания. От других видов, отмеченных в зал. Нячанг и прилегающих акваториях, легко отличим наличием чешуйного покрова в превентральной области. Сравнение с подобными видами из других районов Южно-Китайского моря дано в определительном ключе, приведённом ниже. Среди видов, известных за пределами акватории Вьетнама, L. venusta наиболее близок к L. kanagashira Kamohara, 1936 из зал. Тоза (Южная Япония), отличаясь, насколько можно судить из описаний (Kamohara, 1936; Ochiai, Yatou, 1985; Yamada, 2002), лишь более короткими Р, не заходящими за шестой луч А (тогда как у L. kanagashira достигают девятого луча А). Отличия между этими видами нуждаются в дальнейшем уточнении. По личному сообщению Т. Ято (T. Yato, Kobe, Japan), изменчивость L. venusta также нуждается в изучении – в пользу этого свидетельствуют и различия в числе чешуй LL между изученными экземплярами (56–60), рыбами типовой серии (52–54) и материалом из вод Тайваня (59–63) (Fowler, 1938; Prokofiev, Yato, 2020. Tabl. 2).
Распространение. Известен от Тайваня до Вьетнама и Филиппин. Во Вьетнаме ранее указывался для Тонкинского залива (Nguyen, 1999). Экземпляры из Манильской бухты (Herre, Kauffman, 1952), возможно, принадлежат не к этому виду, а к одному из описанных Ли (Li, 1981).
КЛЮЧ ДЛЯ ИДЕНТИФИКАЦИИ ВИДОВ LEPIDOTRIGLA ЮЖНО-КИТАЙСКОГО МОРЯ
1 (8) Чешуя на груди имеется ……………..…..…. 2
2 (5) Пятый луч соединённого P заметно удлинён по сравнению с соседними (Li, 1981) …........ 3
3 (4) Мягкая часть D с двумя рядами тёмных пятен; наибольший свободный луч Р не заходит за анус …………………………. L. lepidojugulata Li, 1981
4 (3) Мягкая часть D не пигментирована; наибольший свободный луч Р заходит за начало А ………………………………………… L. longimana Li, 1981
5 (2) Пятый луч соединённого P соизмерим с соседними …..…..................……………………………. 6
6 (7) Конец соединённого Р достигает 11-го луча А; мягкая часть D с двумя рядами тёмных пятен ……….….……..…………..… L. argyrosoma Fowler, 1938
7 (6) Конец соединённого Р не заходит за шестой луч А; мягкая часть D не пигментирована …………….………………………… L. venusta Fowler, 1938
8 (1) Грудь не покрыта чешуёй …………..…….… 9
9 (12) Р длинный, заходит за середину длины основания А …….................…………………………. 1011
10 (11) Внутренняя сторона Р у фиксированных рыб без рисунка ............ L. gr. japonica (Bleeker, 1854)22
11 (10) Внутренняя сторона Р у фиксированных рыб со светлыми пятнами (Fowler, 1938) ………..…………………. L. macrobrachium Fowler, 1938
12 (9) Р короткий, не заходит за середину длины основания А …………..…………….…................. 13
13 (16) Наибольший свободный луч Р далеко не достигает вершины наибольшего луча V ……. 14
14 (15) Внутренняя сторона P с интенсивно-чёрным пятном, несущим рисунок из мелких светлых пятнышек; главная вершина рострального выступа не более трёх раз содержится в диаметре глаза (Yamada, 2002) ..…….......……. L. kishinouyei Snyder, 191133
14 (15) Внутренняя сторона P без чёрного пятна; главная вершина рострального выступа более трёх раз содержится в диаметре глаза (Yamada, 2002) ………………………. L. microptera Günther, 1873
16 (13) Наибольший свободный луч Р слегка не достигает или едва заходит за вершину наибольшего луча V ….………………....…………………………. 17
17 (30) Внутренняя сторона Р у фиксированных рыб без оформленного светлого рисунка ..……….. 18
18 (23) Посторбитальная бороздка редуцирована до слабых выемок у заднебокового конца надглазничных гребней или полностью отсутствует; преорбитальные шипы редуцированы или отсутствуют …………………...……..…………………… 19
19 (20) Глаз крупный, его диаметр превышает преорбитальную длину; вентральная часть туловища ниже уровня клейтрального шипа у фиксированных рыб ярко-серебристая ......... L. oglina Fowler, 1938
20 (19) Глаз не столь большой, его диаметр меньше преорбитальной длины; покрытые чешуёй участи тела у фиксированных рыб окрашены одинаково …………...................................………… 21
21 (22) Перепонка колючей части D c тёмным (при жизни – красным) скоплением пигмента в задней части; щитки с одиночными шипами начинаются от задней половины колючей части D; чешуя на вентральной стороне тела циклоидная ….......………………………..… L. alata (Houttuyn, 1782)
22 (21) Перепонка колючей части D не пигментирована; щитки с одиночными шипами под колючей частью D отсутствуют; циклоидная чешуя обнаруживается лишь на ограниченном участке позади оснований Р и V …...............L. marisinensis (Fowler, 1938)
23 (18) Посторбитальная бороздка всегда глубокая позади надглазничных гребней; преорбитальные шипы хорошо развиты …………………… 24
24 (25) Ростральные отростки в виде округлых ошиплённых выступов; по телу разбросаны мелкие чёрные пятнышки ............… L. longifaciata Yato, 1981
25 (24) Ростральные отростки всегда с хорошо развитой главной вершиной; на теле нет пятен ............................................................................. 26
26 (27) Вторая колючка D сильно удлинена по сравнению с соседними ... L. guentheri Hilgendorf, 1879
27 (26) Вторая колючка D ненамного длиннее соседних ……………….…....................………………. 28
28 (29) Ростральные отростки короткие треугольные (Prokofiev, Yato, 2020. Fig. 1B); внутренняя сторона Р с размытым просветлением неопределённых очертаний в проксимальной части над интенсивно-чёрным участком, перепонка между 10-м и 11-м лучами светлая; чешуя легко опадающая; LL–D 3 ………..........…… L. deasoni Herre et Kauffman, 1952
29 (28) Ростральные отростки длинные треугольные (Prokofiev, Yato, 2020. Fig. 1A); внутренняя сторона Р однотонно серо-чёрная над интенсивно-чёрным участком, перепонка между 9-м и 11-м лучами светлая; чешуя крепко сидящая; LL–D 4 …………………. L. firmisquamis Prokofiev et Yato, 2020
30 (17) Внутренняя сторона Р у фиксированных рыб с белыми пятнами ………….…..........….. 31
31 (32) Белые пятна на внутренней стороне Р у фиксированных рыб немногочисленные, ограничены участком интенсивно-чёрной пигментации в нижней части плавника; главная вершина рострального отростка удлиняется с ростом рыб; париетальные шипы отсутствуют; заглазничная бороздка прервана, по меньшей мере, у взрослых рыб (у молоди могут прослеживаться следы сплошной бороздки)…......….. L. punctipectoralis Fowler, 1938
32 (31) Белые пятна распределены по всей внутренней стороне Р у фиксированных рыб; главная вершина рострального отростка значительно длиннее дополнительных даже у молоди; париетальные шипы хорошо развиты; заглазничная бороздка сплошная и отчётливая на всех стадиях онтогенеза …………..........……… L. cf. spiloptera Günther, 1880
Список литературы
Fowler H.W. 1938. Descriptions of new fishes obtained by the United States Bureau of Fisheries steamer Albatross, chiefly in Philippine seas and adjacent waters // Proc. US Natl. Mus. V. 85. № 3032. P. 31–135. https://doi.org/10.5479/si.00963801.85-3032.31
Fricke R., Eschmeyer W.N., van der Laan R. (eds.). 2022. Eschmeyer’s catalog of fishes: genera, species, references (http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Version 03/2022).
Gomon M.F., Kawai T. 2018. A review of Indonesia’s Indian Ocean species of Lepidotrigla gurnards (Teleostei: Scorpaeniformes: Triglidae) with descriptions of three new species from southern coastal waters // Raffles Bull. Zool. V. 66. P. 624–651. https://doi.org/10.5281/zenodo.5360523
Gomon M.F., Psomadakis P.N. 2018. Review of the Lepidotrigla gurnards (Teleostei: Scorpaeniformes: Triglidae) in the Bay of Bengal and Andaman Sea of Myanmar with a description of a new species // Ibid. V. 66. P. 66–77. https://doi.org/10.5281/zenodo.5360658
Günther A. 1880. Report on the shore fishes procured during the voyage of H. M. S. Challenger in the years 1873–1876 // Zoology. V. 1. Pt. 6. P. 1–82.
Herre A.W.C.T., Kauffman D.E. 1952. New and little known Philippine triglids // Proc. Biol. Soc. Wash. V. 65. P. 27–30.
Hubbs C.L., Lagler K.F. 1958. Fishes of the Great Lakes region // Cranbrook Inst. Sci. Bull. № 26. 213 p.
Li S.-Z. 1981. Two new Lepidotrigla species from the South China Sea // Zool. Res. V. 2. № 4. P. 295–300.
Kamohara T. 1936. On two new species of fishes found in Japan // Dobutsugaku Zasshi (Zool. Mag. Tokyo). V. 48. № 12. P. 1006–1008.
Menon A.G.K., Rama Rao K.V. 1963. Notes on three rare marine fishes from Madras // Ann. Zool. V. 4. № 5. P. 47–51.
Nguyen H.P. 1999. Checklist of marine fishes in Vietnam. V. 5. Hochiminh: Agricult. Publ. House, 305 p.
Ochiai A., Yatou T. 1985. Family Triglidae gurnards // The fishes of the Japanese Archipelago. Pt. 1. Tokyo: Tokai Univ. Press. P. 333–334.
Prokofiev A.M., Yato T. 2020. A new species of Lepidotrigla from the South China Sea off Vietnam (Teleostei: Scorpaeniformes: Triglidae) // Zootaxa. V. 4877. № 2. P. 373–385. https://doi.org/10.11646/zootaxa.4877.2.9
Randall J.E., Lim K.K.P. 2000. A checklist of the fishes of the South China Sea // Raffles Bull. Zool. Suppl. № 8. P. 569–667.
Richards W.J. 1992. Comments on the genus Lepidotrigla (Pisces: Triglidae) with descriptions of two new species from the Indian and Pacific Oceans // Bull. Mar. Sci. V. 51. № 1. P. 45–65.
Richards W.J., Jones D.L. 2002. Preliminary classification of the gurnards (Triglidae: Scorpaeniformes) // Mar. Freshw. Res. V. 53. № 2. P. 274–282. https://doi.org/10.1071/MF01128
Richards W.J., Saksena V.P. 1977. Systematics of the gurnards, genus Lepidotrigla (Pisces, Triglidae) from the Indian Ocean // Bull. Mar. Sci. V. 27. № 2. P. 208–222.
Satapoomin U. 2011. The fishes of southwestern Thailand, the Andaman Sea – a review of research and a provisional checklist of species // Phuket Mar. Biol. Cent. Res. Bull. № 70. P. 29–77.
Shao K.-T. 2022. The Fish Database of Taiwan. http://fishdb.sinica.edu.tw. Version 03/2022
Shao K.-T., Ho H.-C., Lin P.-L. et al. 2008. A checklist of the fishes of southern Taiwan, northern South China Sea // Raffles Bull. Zool. Suppl. № 19. P. 233–271.
Shen S.-C., Chen C.-H., Lee S.-C. et al. 1993. Fishes of Taiwan. Taipei: Natl. Taiwan Univ. Press, 960 pp.
Yamada U. 2002. Triglidae // Fishes of Japan with pictorial keys to the species. V. 1. Tokyo: Tokai Univ. Press. P. 604–609.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии