

Вопросы ихтиологии, 2022, T. 62, № 6, стр. 680-685
Новые данные по морфологии и распространению двух редких видов ошибнеобразных рыб: Diplacanthopoma japonicum (Bythitidae) и Pycnocraspedum microlepis (Neobythitidae)
А. М. Прокофьев *
Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 28.02.2022
После доработки 14.03.2022
Принята к публикации 15.03.2022
- EDN: QJJXRM
- DOI: 10.31857/S0042875222060297
Аннотация
Уточнены морфология и распространение Diplacanthopoma japonicum и Pycnocraspedum microlepis. Вид D. japonicum впервые отмечен в водах Северо-Западной Австралии. Предполагается возможность синонимии номинальных видов D. japonicum и D. nigripinne. Пять экземпляров P. microlepis, пойманных на хребте Кюсю-Палау, позволяют уточнить изменчивость некоторых диагностических признаков вида. Составлен ключ для определения видов рода Pycnocraspedum.
В сборах отечественных рыбопоисковых экспедиций, ныне хранящихся в Институте океанологии (ИО) РАН, Москва, обнаружено несколько экземпляров редких видов ошибнеобразных рыб из родов Diplacanthopoma Günther, 1887 и Pycnocraspedum Alcock, 1889. Сведения о видах этих родов крайне ограничены, их идентификация затруднительна, а характер внутривидовой изменчивости практически не изучен. В свете этого описание нового материала представляет несомненный интерес, тем более что новый экземпляр D. japonicum (Steindachner et Döderlein, 1887) существенно расширяет известный ареал вида, а P. microlepis (Matsubara, 1943) представлен в изученной коллекции серией из пяти экземпляров (больше, чем было описано в литературе за всё предшествующее время).
Традиционное разделение отряда Ophidiiformes на подотряды живородящих (Bythithoidei) и икромечущих (Ophidioidei) ошибнеобразных (Cohen, Nielsen, 1978; Nielsen et al., 1999) неоднократно было поставлено под сомнение (Howes, 1992; Прокофьев, 2004а, 2004б). По результатам изучения комплекса передних позвонков, ассоциированных с плавательным пузырём (Прокофьев, 2004б), из сборного семейства Ophidiidae sensu Cohen, Nielsen (1978) было выделено семейство Neobythitidae в составе прежних подсемейств Neobythitinae и Brotulotaeniinae и трибы Sirembini в классификации Коэна и Нильсена, но с исключением рода Hypopleuron Smith et Radcliffe, 1913, который был выделен в особое подсемейство в составе Ophidiidae s. str. (Прокофьев, 2004а, 2004б). Более детальный морфологический анализ (Ohashi, 2014) полностью подтвердил выделение семейства Neobythitidae в указанных границах, показав его сестринские отношения с комплексом, включающим Carapidae, Hypopleuron и Ophidiidae s. str. На основании этого я считаю неправомочным объединение Ophidiidae и Neobythitidae в одном семействе и их противопоставление Carapidae, что продолжает фигурировать в самых последних сводках (Nelson et al., 2016). Такое объединение формально допустимо лишь при слиянии всех семейств икромечущих ошибнеобразных в одно, что вряд ли оправдано.
МАТЕРИАЛ И МЕТОДИКА
Описанные экземпляры принадлежат коллекции ИО РАН. Методика изучения соответствует общепринятой для группы (Cohen, Nielsen, 1978). Подсчёт числа лучей в непарных плавниках и позвонков выполнен по рентгенограммам, изготовленным автором; при подсчёте позвонков уростиль не учтён. Подсчёты, различающиеся на разных сторонах одной и той же рыбы, разделены знаком “/”. “Ложным хвостом” названо состояние, возникающее при регенерации утраченного хвостового конца тела, когда С замещается регенерирующими лучами вертикальных плавников (Iwamoto, 1970). Поры сейсмосенсорных каналов головы с крупными зияющими отверстиями именуются вслед за Андрияшевым (1955) ноздревидыми. В тексте использованы следующие сокращения: D, A, P, V и C – спинной, анальный, грудные, брюшные и хвостовой плавники; vert. – число позвонков (туловищных (vert. abd.) + хвостовых), TL и SL – абсолютная и стандартная длина, lc – длина головы, H и HA – максимальная высота тела и его высота на уровне начала А; aD, aA и aV – соответственно предорсальное, преанальное и превентральное расстояние; lP и lV – длина Р и V, ao – длина рыла, oo – горизонтальный диаметр орбиты, io – ширина костного межглазничного промежутка, lmx и hmx – длина верхней челюсти и высота пластинки maxillare, БМРТ – большой морозильный рыболовный траулер.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Diplacanthopoma japonicum (Steindachner et Döderlein, 1887)
(рис. 1)
Рис. 1.
Diplacanthopoma japonicum TL 245+ мм, ИО РАН № 03620: а – общий вид, б – рентгенограмма, в – свободные невромасты
на дорсальной поверхности рыла, г – кожная лопасть ( ) “Diplacanthopoma-flap” – по: Прокофьев, 2004б. Масштаб: 15 мм.
) “Diplacanthopoma-flap” – по: Прокофьев, 2004б. Масштаб: 15 мм.
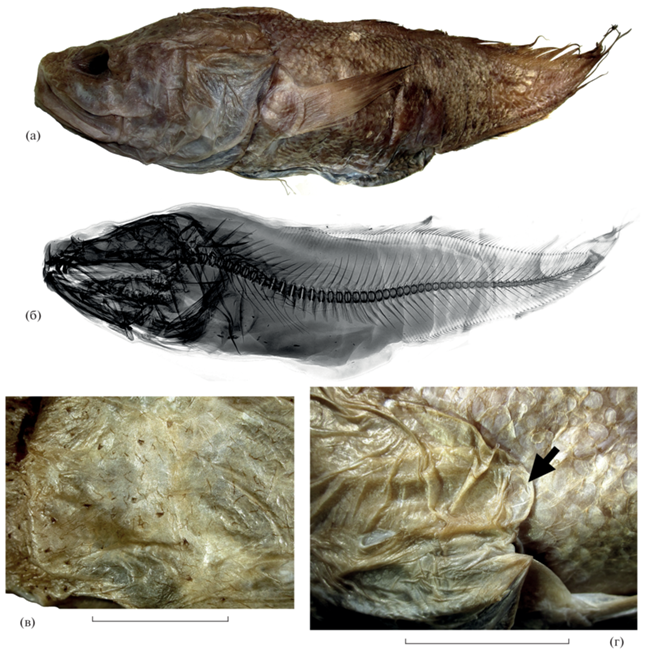
Материал. ИО РАН № 03620, 1 экз. TL 245+ мм, 09°01′12″ ю.ш. 130°59′30″ в.д., БМРТ “Академик Берг”, донный трал, глубина 540 м.
Описание. P 29/31, V 1; vert. abd. 20. Хвостовой конец рыбы утрачен при жизни, формирует “ложный хвост” (рис. 1а, 1б), в связи с чем корректный подсчёт числа лучей в вертикальных плавниках и общего числа позвонков невозможен. Голова массивная, широкая; рыло несколько уплощённое, межглазничное пространство вдавлено между орбитами, верхний костный край орбит приподнят. Дорсальный профиль головы резко повышается от глаз к затылку, максимальная высота тела приходится на предорсальную область, в 1.55 раза меньше длины головы. Две ноздри; передняя меньшего размера, открывается очень короткой трубочкой над верхней челюстью на равном расстоянии от вершины рыла и переднего края орбиты. Задняя ноздря расположена выше передней, открывается непосредственно перед передним краем орбиты крупным овальным отверстием с приподнятыми верхним, передним и нижним краями. Рот большой, верхняя челюсть оканчивается позади вертикали заднего края глаза; пластинка maxillare свободная, сильно расширена кзади, её задний край слабо вогнутый; имеется хорошо развитое supramaxillare. При закрытом рте бо́льшая часть maxillare прикрыта снаружи широкими инфраорбитальными костями, оставляющими видимым только нижнезадний конец пластинки maxillare. Озубление praemaxillare видно снаружи при закрытом рте. На челюстях, сошнике и нёбных костях мелкие конические зубы, расположенные полосками; сошниковое зубное пятно в виде широкой V-образной поперечной полосы; базибранхиальные зубные пластинки отсутствуют. В супраорбитальном канале две поры, первая очень крупная, ноздревидная, открывается у вершины рыла медиальнее передней ноздри; задняя пора небольшая, открывается на дорсальной поверхности головы несколько позади вертикали заднего края орбиты. В инфраорбитальном канале шесть ноздревидных пор, из них первая – наименьшая и расположена выше остальных, третья и шестая – наибольшие; передние три сближены, пятая расположена над задним концом maxillare, шестая – перед передним краем praeoperculum на уровне нижнего края орбиты. В темпоральном канале две крупные поры, расположенные над жаберной крышкой. В предкрышечно-нижнечелюстном канале семь ноздревидных пор; первая пара пор широко разобщена, пятая пора расположена над нижнечелюстным суставом, две последние поры гигантских размеров, соизмеримы с диаметром зрачка, открываются на praeoperculum. Корональная и супратемпоральная комиссуры отсутствуют. Кожа верха и боков головы сплошь покрыта разреженными волосовидными тёмноокрашенными папиллами и более крупными и редкими (но образующими заметные сгущения на верхней поверхности рыла и вокруг глаз) узкими клапановидными тёмноокрашенными невромастами (рис. 1в). На вершине рыла у передненижнего края первой супраорбитальной поры и под передней ноздрёй имеется несколько коротких толстых кожных выростов, вероятно, являющихся рудиментами ростральной (этмоидной) комиссуры (Балушкин, Прокофьев, 2005). Туловищная боковая линия представлена легко обрывающимися свободными невромастами. Praeoperculum с неровным гладким задним краем, покрытым кожей, без шипов. Оперкулярная лопасть к вершине сужена и заострена, оперкулярный шип изогнут кверху, его конец достигает заднего края operculum. Над верхним краем operculum хорошо развита широкая закруглённая на свободном конце кожная лопасть (“Diplacanthopoma-flap” – по: Прокофьев, 2004б) (рис. 1г). На первой жаберной дуге три (1 + 2) развитые жаберные тычинки, помимо них имеется 16 (3 + 1 + 12) бугорковидных. Ложножабра рудиментарная, представлена тремя короткими и широкими элементами. Язык массивный, его конец свободный.
Начало D расположено заметно позади основания Р, начало V – под praeoperculum, концы лучей V заметно заходят за вертикаль основания Р. Основание Р образует очень короткую и широкую лопасть с мясистым выростом у верхнего края, конец прижатого к телу Р достигает вертикали ануса. Анус расположен у начала А. Начало А расположено на вертикали 31-го луча D. Тело покрыто некрупной циклоидной чешуёй, на голове чешуя полностью отсутствует. Около 18 поперечных рядов чешуй в предорсальной области.
Окраска фиксированной рыбы после длительного хранения в формалине светло-коричневая, белеющая на нижней поверхности головы, груди и брюхе; дистальная половина Р и дистальные края вертикальных плавников заметно темнее, V светлые, почти белые. Ротовая полость светлая, жаберная – тёмная.
Измерения. В мм: lc 85, H 55, HA 47, aD 95, aA 155, lP 51, lV 39, ao 18, oo 15, io 24, lmx 35, hmx 13. В % lc: ao 21.2, oo 17.7, io 28.2, lmx 41.2, hmx 15.3.
Сравнительные замечания. Виды рода Diplacanthopoma редки в коллекциях, и их таксономия совершенно не разработана. Было описано восемь номинальных видов (название D. alcockii Goode et Bean, 1896, согласно работе Коэна и Нильсена (Cohen, Nielsen, 2002), является nomen nudum): D. brachysoma Günther, 1887 (Западная Атлантика); D. brunneum Smith et Radcliffe, 1913 (Филиппины); D. japonicum (Южная Япония); D. jordani Garman, 1899 (тропическая Восточная Пацифика); D. kreffti Cohen et Nielsen, 2002 (Северо-Западная Австралия); D. nigripinne Gilchrist et von Bonde, 1924 (Южная Африка); D. raniceps Alcock, 1898 (Андаманское море) и D. riversandersoni Alcock, 1895 (Аравийское море). Эти виды было предложено разделить на две группы по величине глаза (Cohen, Nielsen, 2002): малоглазые (oo 14.8–18.9% lc), включающие D. japonicum, D. kreffti и D. riversandersoni, и большеглазые (oo 20.0–25.4% lc) – все остальные. Это разделение представляется весьма условным, а пределы внутривидовой, в том числе возрастной, изменчивости данного признака остаются невыясненными ни у одного вида. Исследованный экземпляр, характеризующийся oo 17.7% lc, попадает в группу малоглазых видов. От единственного ранее известного в австралийских водах представителя рода (D. kreffti, шельф северо-запада Австралии) он хорошо отличается заметно более короткой и высокой верхней челюстью (lmx и hmx – 41.2 и 15.3 против 49.3 и 10.6% lc), которая большей частью свободна (прикрыта складкой кожи у D. kreffti). По данным Коэна и Нильсена (Cohen, Nielsen, 2002), D. kreffti также отличается от D. japonicum меньшим числом лучей D и А (соответственно 117 и 86 против 132–134 и 100–101), но у описываемой рыбы эти признаки неустановимы из-за утраты хвостового конца тела. Однако число лучей D перед началом А у изученного экземпляра (31) также меньше, чем у голотипа D. kreffti (37). Изученный экземпляр заметно отличается от голотипа и единственного известного экземпляра D. kreffti меньшей шириной межглазничного промежутка и длиной Р, но большей длиной V (соответственно 28.2, 60.0 и 45.9 против 34.6, 68.6 и 36.9% lc) и бо́льшим числом развитых жаберных тычинок на первой дуге (четыре против трёх).
Изученный экземпляр не может быть отождествлён с D. riversandersoni, так как имеет заметно больше лучей Р (29–30 против 24) и не столь расширенную кзади пластинку maxillare (hmx 15.3 против 18.2% lc) (Alcock, 1895; Cohen, Nielsen, 2002). Все признаки исследованной рыбы хорошо согласуются с данными, приводимыми разными авторами (Steindachner, Döderlein, 1887; Machida, 1988; Cohen, Nielsen, 2002; Nakabo, 2002) для D. japonicum, на основании чего я считаю австралийский экземпляр конспецифичным указанному виду.
Вместе с тем нужно отметить, что южноафриканский вид D. nigripinne занимает промежуточное положение между большеглазыми и малоглазыми видами, имея oo 20% lc согласно первоописанию (Gilchrist, von Bonde, 1924; Cohen, Nielsen, 2002). При указываемой для D. japonicum изменчивости по данному признаку в 15.9–18.9% это значение вполне может представлять частный случай индивидуальной изменчивости. Голотип D. nigripinne, как и многие другие типы Гилкриста, стал жертвой вандализма некоего Хогбена (L.T. Hogben) (Pietsch, 1972). Второй экземпляр этого вида был кратко охарактеризован Коэном (Cohen, 1986). Его промеры указаны не были, а число лучей в D и А приведено соответственно как ca.120 и ca.70. Это существенно меньше, чем известно для D. japonicum и среди всех видов рода приближается только к D. raniceps, однако число лучей Р (29) выходит за рамки значений, известных для D. raniceps (21–26) (Cohen, Nielsen, 2002). В то же время Коэн (Cohen, 1986) указывает лишь приблизительное число лучей и не приводит число позвонков. Для видов Diplacanthopoma характерно резкое истончение каудального конца тела, который часто утрачивается ещё при жизни и замещается ложным хвостом (Cohen, Nielsen, 2002). Если приблизительный подсчёт числа лучей в вертикальных плавниках у второго южноафриканского экземпляра объясняется наличием ложного хвоста (приводимый в работе рисунок рыбы не даёт полной уверенности, что это не так), то никаких других признаков, различающих D. japonicum и D. nigripinne не остаётся. Эти виды вполне могут оказаться конспецифичными. К сожалению, обсуждаемый экземпляр также утерян ещё до инвентаризации коллекции при переносе её в другое здание в 2007 г. (О. Гон, личное сообщение, октябрь 2019 г.).
Распространение. Вид был до сих пор известен по двум находкам у тихоокеанского побережья Японии (у г. Токио и у м. Урадо) (Steindachner, Döderlein, 1887; Machida, 1988; Cohen, Nielsen, 2002; Nakabo, 2002). Новая находка в Тиморском море между о-вами Танимбар и северо-западным побережьем Австралии значительно расширяет известный ареал вида, однако, если предположение о синонимии D. nigripinne c D. japonicum верно, то он может быть распространён на батиальных глубинах по всей тропической и субтропической Индо-Вест-Пацифике.
Pycnocraspedum microlepis (Matsubara, 1943)
(рис. 2)

Материал. ИО РАН № 03619, 5 экз. TL 185–330 мм, SL 167–315 мм, 26°05′ с.ш. 135°49′ в.д., глубина 320–340 м, 07.01.1982 г.
Описание. D 95–98, A 70–71, P 23–27, V 2, C 10; vert. 13 + 39–41 = 52–54. Голова умеренной величины, укладывается 4.0–4.6 раза в SL, 1.8–2.0 раза – в преанальном расстоянии. Рыло притупленное, широко закруглённое, равно или едва короче горизонтального диаметра глаза, который в 1.3 (SL 167 мм) или 1.6 (SL 213–315 мм) раза меньше ширины межглазничного промежутка. Две ноздри, лежащие на одной горизонтали на уровне середины глаза. Нижняя челюсть короче верхней, озубление praemaxillaria видно при закрытом рте; верхняя челюсть оканчивается далеко позади заднего края орбиты, её пластинка высокая с вогнутым задним краем; supramaxillare хорошо развито. На челюстях, сошнике, нёбных костях и базибранхиальных зубных пластинках очень мелкие зубы, расположенные полосками. Базибранхиальных пластинок две, медиальные; передняя в 3–4 раза длиннее задней, 3.3–3.9 раза содержится в длине головы. В супраорбитальном канале две поры, первая крупная, ноздревидная, расположена позади вершины рыла непосредственно над верхней губой; вторая небольшого размера, открывается на уровне ноздрей. В инфраорбитальном и преоперкуло-мандибулярном каналах по девять пор, мандибулярных пор пять, преоперкулярных – четыре. Темпоральный канал открывается одной крупной порой, расположенной на свободной кожной лопасти (“Diplacanthopoma-flap”) над верхним краем жаберного отверстия. Корональная и супратемпоральная комиссуры отсутствуют. Туловищный канал отчётливый, жёлобовидный, фрагментируется и исчезает в задней четверти тела. Praeoperculum с тремя–четырьмя короткими шипами; оперкулярный шип короткий, далеко не достигает заднего края оперкулярной лопасти. На первой жаберной дуге четыре развитые жаберные тычинки, помимо них имеется 3–4 + 3 + + 10–15 зубных пластинок (перед, между и позади развитых жаберных тычинок). Ложножабра, как правило, отсутствует, но у 1 экз. обнаружено четыре коротких расставленных элемента. Пилорических придатков 18 (по 1 экз.).
Начало D расположено впереди вертикали основания Р, начало V – под praeoperculum; концы лучей V нитевидные; конец прижатого к телу Р не достигает ануса. Анус расположен у начала А. Тело покрыто некрупной циклоидной чешуёй, на голове доходящей до вершины рыла.
Окраска фиксированных рыб коричневая (там, где чешуя облетела – светлая), туловищный канал боковой линии отчётливо затемнён, вертикальные плавники черноватые. Ротовая полость светлая, жаберная – тёмная. Желудок в передней половине черноватый, в задней – светлый.
Измерения. В % SL: lc 22.2–25.2, H 19.2–21.6, HA 18.0–20.2, aD 18.0–22.2, aA 43.6–44.9, aV 16.2–19.1, lP 16.0–19.7 (у 1 экз. – 14.9), lV 12.1–18.8, ao 3.9–4.4, oo 4.2–4.8, io 6.0–7.1, lmx 12.0–14.3, hmx 4.5–5.1. В % TL: SL 90.3–95.5.
Сравнительные замечания. В составе рода описан единственный атлантический вид P. phyllosoma (Parr, 1933), известный только по метаморфозирующему экземпляру, и четыре индо-тихоокеанских: P. armatum Gosline, 1954 (Гавайи); P. fulvum Machida, 1984 (жёлоб Окинава); P. microlepis (Южная Япония) и P. squamipinne Alcock, 1889 (от Восточной Африки до Новой Каледонии; возможно, сборная группа) (Nielsen et al., 1999; Прокофьев, 2005). Отнесение описанных экземпляров к виду P. microlepis основано на числе лучей в вертикальных плавниках, шипов предкрышки, пилорических придатков; отсутствии латеральных базибранхиальных зубных пластин и относительно коротких Р, не достигающих ануса (Gosline, 1954; Machida, 1984; Nakabo, 2002; Прокофьев, 2005; Teena et al., 2021). От имеющихся описаний P. microlepis изученные рыбы отличаются лишь большей изменчивостью по числу лучей Р (23–27 против 25–28) и шипов предкрышки (3–4 против 3) (Matsubara, 1943; Machida, 1984; Nakabo, 2002). От всех других индо-тихоокеанских представителей рода P. microlepis отличается бо́льшим числом пилорических придатков (18–20 против 12–13), близким к таковому у атлантического P. phyllosoma (21). Кроме того, от P. armatum он отличается короткими Р, не достигающими ануса (у P. armatum заходят за анус), и более задним положением начала D (позади вертикали заднего края praeoperculum, тогда как у P. armatum – на этой вертикали) (Gosline, 1954), а от P. fulvum – отсутствием латеральных базибранхиальных зубных пластин и ложножабры, более развитым ошиплением praeoperculum и бо́льшим числом лучей в вертикальных плавниках (Machida, 1984; Nakabo, 2002). Для определения видов Pycnocraspedum можно предложить следующий ключ.
1 (2) Развитых жаберных тычинок пять или шесть ………................……….………..……. P. armatum
2 (1) Развитых жаберных тычинок четыре ..… 3
3 (4) Два шипа на praeoperculum; латеральные базибранхиальные пластинки имеются (P. fulvum, не известно для P. phyllosoma) …………………..….… 5
4 (3) Три или четыре шипа на praeoperculum; латеральные базибранхиальные пластинки, как правило, отсутствуют (в виде исключения, редуцированная пластинка может быть обнаружена с одной стороны (Прокофьев (2005)) …………….…. 7
5 (6) D 83, A 63; пилорических придатков 13; Тихий океан к югу от Японии (жёлоб Окинава) …………....…………………………………………… P. fulvum
6 (5) D 97, A 71; пилорических придатков 21; Атлантический океан у Багамских о-вов ..................................................…… P. phyllosoma
7 (8) Пилорических придатков 12 или 13; голова укладывается 3.0–3.9 раза в SL ........... P. squamipinne
8 (7) Пилорических придатков 18–20; голова укладывается 4.0–4.6 раза в SL ………. P. microlepis
Распространение. Вид известен из тихоокеанских вод Японии от моря Кумано Нада до зал. Тоса и с хребта Кюсю-Палау (Nakabo, 2002), также был указан для Восточно-Китайского моря (Nielsen et al., 1999). Наши экземпляры пойманы на хребте Кюсю-Палау.
Список литературы
Андрияшев А.П. 1955. Обзор угревидных ликодов [Lycenchelys Gill (Pisces, Zoarcidae) и близкие формы] морей СССР и сопредельных вод // Тр. ЗИН АН СССР. Т. 18. С. 349–384.
Балушкин А.В., Прокофьев А.М. 2005. Новый вид рода Cataetyx (Ophidiiformes: Bythitidae) с банок Геракла (Южно-Тихоокеанское поднятие) // Вопр. ихтиологии. Т. 45. № 4. С. 554–558.
Прокофьев А.М. 2004а. О таксономическом положении и родственных отношениях Hypopleuron caninum Smith & Radcliffe, 1913 (Teleostei: Ophidiiformes) // Объед. науч. журн. № 14 (106). С. 64–69.
Прокофьев А.М. 2004б. Строение и таксономическое значение комплекса передних абдоминальных позвонков у представителей отряда Ophidiiformes (Pisces, Paracanthopterygii), и вопросы классификации отряда // Естеств. и тех. науки. № 2 (11). С. 129–142.
Прокофьев А.М. 2005. О некоторых редких видах ошибнеобразных рыб из Южной Атлантики и Индо-Вест-Пацифики с описанием нового рода Megacataetyx gen. novum (Teleostei: Ophidiiformes) // Там же. № 2 (16). С. 111–128.
Alcock A. 1895. Natural history notes from H.M. Indian marine survey steamer “Investigator”. Ser. II. № 18. On a new species of viviparous fish of the family Ophidiidae // Ann. Mag. Nat. Hist. Ser. 6. V. 16. P. 144–146.
Cohen D.M. 1986. Bythitidae // Smith’s Sea fishes. Johannesburg: McMillan South Africa. P. 354–356.
Cohen D.M., Nielsen J. 1978. Guide to the identification of genera of the fish order Ophidiiformes with a tentative classification of the order // NOAA Tech. Rep. NMFS Circ. 417. Seattle: NOAA; NMFS. vii + 72 p. https://doi.org/10.5962/bhl.title.63242
Cohen D.M., Nielsen J. 2002. Diplacanthopoma kreffti (Pisces, Bythitidae), a new species from the Northwest Australian shelf, with comments on the name D. alcockii Goode and Bean, 1896 // Arch. Fish. Mar. Res. V. 50. № 1. P. 11–15.
Gilchrist J.D.F., von Bonde C. 1924. Deep-sea fishes procured by the S.S. “Pickle” (Pt. 2) // Rept. Fish. Mar. Biol. Surv. Union of South Africa. V. 3. № 7. P. 1–24, Pls. 1–6.
Gosline W.A. 1954. Fishes killed by the 1950 eruption of Mauna Loa. II. Brotulidae // Pac. Sci. V. 8. № 1. P. 68–83.
Iwamoto T. 1970. Macrourid fishes of the Gulf of Guinea // The R/V Pillsbury deep-sea biological expedition to the Gulf of Guinea, 1964–65. Ser. Stud. Trop. Oceanogr. № 4. Pt. 2. Miami: UM. P. 316–431.
Machida Y. 1984. Pycnocraspedum fulvum // Fishes of the Okinawa Trough and the adjacent waters. V. 1. Tokyo: JFRCA. 247 p. + 173 Pl.
Machida Y. 1988. An additional specimen of an imperfectly known bythitid fish, Diplacanthopoma japonicum (Bythitidae, Ophidiiformes) // Rep. Usa Mar. Biol. Inst. Kochi Univ. № 10. P. 69–73.
Matsubara K. 1943. Ichthyological annotations from the depth of the Sea of Japan. I–VII // J. Sigenkagaku Kenkyusyo. V. 1. № 1. P. 37–82. Pl. 1.
Nakabo T. 2002. Ophidiiformes // Fishes of Japan with pictorial keys to the species. V. 1. Tokyo: Tokai Univ. Press. P. 436–451.
Nielsen J.G., Cohen D.M., Markle D.F., Robins C.R. 1999. Ophidiiform fishes of the world. An annotated and illustrated catalogue of pearlfishes, cusk-eels, brotulas and other ophidiiform fishes known to date // FAO Spec. catalogue. V. 18. Rome: FAO, 178 p.
Nelson J.S., Grande T.C., Wilson M.V.H. 2016. Fishes of the world. Hoboken: John Wiley and Sons, 707 p. https://doi.org/10.1002/9781119174844
Ohashi S. 2014. Comparative morphology and phylogenetic systematics of the family Ophidiidae and related taxa (Teleostei: Ophidiiformes): Ph.D. Thesis, Hokkaido: HUSCAP, 5 p. (http://hdl.handle.net/2115/55362. Version 03/2022).
Pietsch T.W. 1972. Ergebnisse der Forschungsreisen des FFS “Walter Herwig” nach Südamerika. XIX. Systematics and distribution of ceratioid fishes of the genus Dolopichthys (family Oneirodidae), with the description of a new species // Arch. Fishereiwiss. V. 23. Pt. 1. P. 1–28.
Steindachner F., Döderlein L. 1887. Beiträge zur Kenntniss der Fische Japan’s (IV) // Denksch. K. Akad. Wiss. Wien, Math.-Wiss. Cl. V. 53. P. 257–296.
Teena J.T.K., Murugan A., Kumar A.T.T., Lal K.K. 2021. Redescription of a rare cusk eel, Pycnocraspedum squamipinne (Actinopterygii, Ophidiiformes, Ophidiidae), from Bay of Bengal // Acta Ichthyol. Piscat. V. 51. № 1. P. 77–83. https://doi.org/10.3897/aiep.51.63469
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии