

Вопросы ихтиологии, 2023, T. 63, № 2, стр. 199-208
Особенности питания девятииглой колюшки Pungitius pungitius (Gasterosteidae) в водоёмах северо-запада России
Н. А. Березина 1, *, Н. Н. Жгарева 2, А. П. Стрельникова 2
1 Зоологический институт РАН – ЗИН РАН
Санкт-Петербург, Россия
2 Институт биологии внутренних вод РАН – ИБВВ РАН
Ярославская область, пос. Борок, Россия
* E-mail: nadezhda.berezina@zin.ru
Поступила в редакцию 09.06.2022
После доработки 26.07.2022
Принята к публикации 27.07.2022
- EDN: EXVTQO
- DOI: 10.31857/S0042875223020029
Аннотация
Изучено питание девятииглой колюшки Pungitius pungitius двух водоёмов северо-запада России – субарктического озера Кривое (Карелия) и эстуария реки Нева (Ленинградская область). Личинки двукрылых – хирономид (Chironomidae), мокрецов (Ceratopogonidae), симулид (Simuliidae) и лимониид (Limoniidae), а также планктонные ракообразные составляли основу пищевого комка рыб, их соотношение по численности и массе различалось между изученными водоёмами. Выявлена сезонная смена доминирующих групп пищевых организмов колюшки оз. Кривое. Кроме беспозвоночных и икры рыб, которые были основой питания колюшки в зимний период, летом в её питании присутствовали водные мхи, нитчатые и диатомовые водоросли. Анализ изотопного состава азота в мышцах колюшки, окуня и в беспозвоночных выявил, что в трофической сети оз. Кривое колюшка располагается в пределах третьего трофического уровня, приближаясь по величинам δ15N (5.2–8.6‰) к хищным рыбам. Особенностью питания P. pungitius является переход от разнообразного рациона в летне-осенний период к монотрофному животному питанию (личиками хирономид и икрой рыб) в подлёдный зимне-весенний период.
Девятииглая колюшка Pungitius pungitius (L.) – циркумполярный вид, широко распространённый в озёрах, морях и реках бассейнов Атлантического, Северного Ледовитого и Тихого океанов (Атлас …, 2003). Эта мелкая (5–9 см) стайная рыба обитает в прибрежных биотопах пресных и морских экосистем. В последние годы девятииглая колюшка активно расширяет свой ареал в России (Аськеев и др., 2010; Клевакин и др., 2011). Она практически не имеет промыслового значения, человек её не употребляет в пищу, а рыболовы зачастую считают сорной рыбой. Вместе с тем девятииглая колюшка входит в рацион промысловых рыб, водных млекопитающих и околоводных птиц. Например, она отмечена в питании арктического гольца Salvelinus alpinus (L.), кижуча Oncorhynchus kisutch (Walbaum) (Бугаев и др., 2007), речного окуня Perca fluviatilis L. (Березина и др., 2021), стерха Grus leucogeranus (Pallas) (Дегтярев и др., 2013), уток (Mergus merganser (L.) и M. serrator (L.)) (Бойко, Бианки, 2009), куликов (Pluvialis fulva (Gmelin), Calidris melanotos (Vieillot)) (Бобырь, Антипин, 2017; Антипин, 2018) и беломорской кольчатой нерпы Pusa hispida Schreber (Светочев, Светочева, 2015).
Основой питания девятииглой колюшки являются организмы планктона и бентоса (Thorman, Wiederholm, 1983). Причём преобладание в её питании какой-либо одной группы организмов является вполне обычным, так как, по мнению некоторых исследователей (Черешнев, 2008), эти рыбы, вероятно, потребляют в первую очередь пищевые объекты, образующие скопления в местах их обитания. В связи с этим спектр питания вида очень вариабелен. Так, в питании девятииглой колюшки, обитающей на мелководных участках эстуария р. Брёлвен (западное побережье Швеции) и в прибрежье Ботнического залива Балтийского моря, обнаружены организмы инфауны, эпибентические животные и зоопланктон (Thorman, Wiederholm, 1983). В нижнем течении р. Большая (Камчатка) девятииглая колюшка питалась многочисленными в этом водоёме бокоплавами (Amphipoda), мизидами (Mysida) и кумовыми (Cumacea) ракообразными (Максименков, Токранов, 1994). По данным Введенской (2008), в пище этого вида из бассейна р. Таловка (Северо-Западная Камчатка) отмечены ветвистоусые ракообразные – босмины (Bosmina), хидорусы (Chydorus), дафнии (Daphnia), а также остракоды (Ostracoda). В оз. Киси бассейна р. Ола (Магаданская область) девятииглая колюшка в основном потребляла моллюсков (47% массы) и остракод (24%), в меньшей степени – личинок хирономид (Chironomidae) и ручейников (Trichoptera): соответственно 13.5 и 15% (Хаменкова, 2011). В оз. Халактырское (побережье Авачинского залива) доля (по массе) личинок хирономид в её питании составляла 41–47% (Погорелова и др., 2020). Нередко эта колюшка потребляла икру и молодь рыб (Максименков, Токранов, 1994). Таким образом, обладая гибкостью в пищевых предпочтениях, девятииглая колюшка при высокой численности может конкурировать за пищевые ресурсы с молодью промысловых рыб. В связи с этим изучение её роли в трофической сети водоёмов имеет немаловажное значение.
О питании девятииглой колюшки в разнотипных водоёмах северной части Европы сравнительно немного данных, которые касаются субарктических озёр (Cameron et al., 1973) и заливов Балтийского моря (Thorman, Wiederholm, 1983; Науменко и др., 2020). В эвтрофном эстуарии р. Нева (бассейн Балтийского моря) недавно было проведено детальное изучение трофических связей рыб прибрежной зоны (Demchuk et al., 2021), но поскольку девятииглая колюшка входила в группу второстепенных видов, её питанию было уделено мало внимания.
Цель работы – выявить спектр питания девятииглой колюшки в субарктическом олиготрофном оз. Кривое (Карелия) и эвтрофном эстуарии р. Нева (Ленинградская область), определить по изотопному составу азота в мышцах позицию этой рыбы в трофической сети оз. Кривое в разные периоды года, сравнить полученные данные с составом питания этого вида в разнотипных водоёмах (озёрах и эстуариях рек) других регионов России для выявления возможных общих закономерностей.
МАТЕРИАЛ И МЕТОДИКА
Изученные местообитания
Озеро Кривое – небольшое субарктическое озеро в Северной Карелии (общая площадь 0.5 км2), расположенное на берегу губы Чупа Белого моря у м. Картеш (табл. 1). Начиная с первых лет исследований этого озера (Winberg et al., 1973) оно сохраняет черты олиготрофной экосистемы с низким содержанием питательных веществ и первичной продукции фитопланктона, а также слабым развитием макрофитов. Прибрежная зона (глубина 0–3 м) составляет 20% общей площади озера. Донные отложения в профундали состоят из серого и бурого ила, глин, минеральных и органических веществ, в то же время для литорали типичны илистый песок, камни и детрит.
Таблица 1.
Некоторые характеристики воды на станциях отлова девятииглой колюшки Pungitius pungitius в оз. Кривое ( 1) и в эстуарии р. Нева (2–5)
| Станция № | Координаты, с.ш./в.д. |
Концентрация в воде | |
|---|---|---|---|
| сумма солей, г/л | общий фосфор, мкг/л | ||
| 1. Оз. Кривое | 66°21′/33°38′ | 0.07–0.08 | 18−25 |
| 2. Приморск | 60°20′/28°43′ | 1.98−2.03 | 88−125 |
| 3. Ольгино | 59°59′/30°05′ | 0.05−0.07 | 23−80 |
| 4. Устье р. Систа | 59°48′/28°54′ | 2.82−3.30 | 45−50 |
| 5. Графская бухта | 59°58′/29°12′ | 2.00–2.30 | 100−148 |
Эстуарий р. Нева расположен в восточной части Балтийского моря, его условно разделяют на пресную (Невскую губу), с содержанием солей в воде 0.05–0.07 г/л, и солоноватоводную (внешний эстуарий, 0.3–5 г/л) части. В последние десятилетия в эстуарии происходит эвтрофирование прибрежной зоны из-за поступления фосфора и азота с водами впадающей Невы. На хорошо прогреваемых мелководьях высока продукция прибрежных растений и нитчатых водорослей, а достигабщая 800–900 г С/м2 в год (Berezina et al., 2005). Две станции отлова колюшки (Ольгино, устье р. Систа) располагались в опреснённой части (табл. 1, рис. 1). Для них характерно высокое развитие зарослей тростника Phragmites sp. и рдестов Potamogeton spp. Также были отловлены рыбы на двух точках в солоноватой части эстуария (станции Графская бухта и Приморск) в зоне нитчатых водорослей Cladophora glomerata (Linnaeus) Kützing и Ulva intestinalis Linnaeus. Субстрат в литорали, где отлавливали рыб, состоял из песка, ила, гальки и валунов.
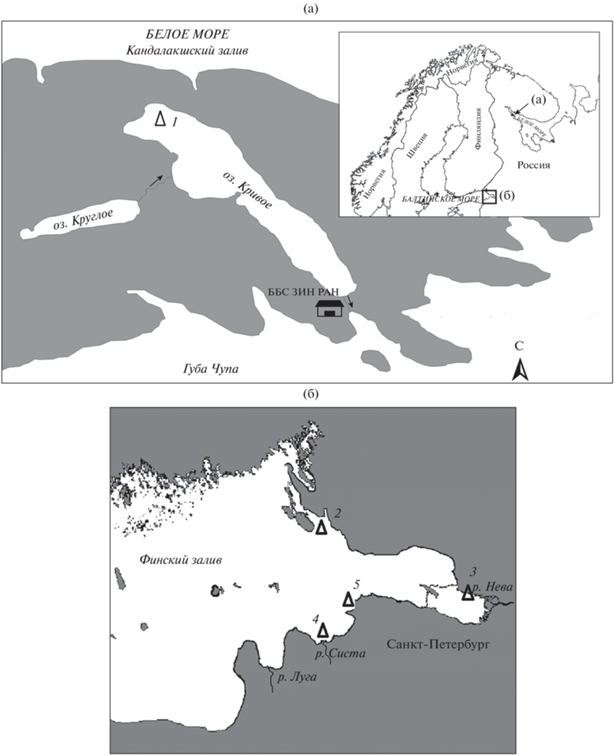
Рис. 1.
Карта-схема расположения станций (△) отлова девятииглой колюшки Pungitius pungitius в оз. Кривое (а) и эстуарии р. Нева (б). Станции: 1 – оз. Кривое, 2 – Приморск, 3 – Ольгино, 4 – устье р. Систа, 5 – Графская бухта; ББС ЗИН РАН – Беломорская биологическая станция Зоологического института РАН; (→) – направление течения.
Сбор материала
Отлов рыб для анализа состава пищевого комка проводили в оз. Кривое в июне, сентябре 2019 г. и феврале, апреле 2020 г.; в эстуарии р. Нева – в июле 2019 г. Летом и осенью отлов проводили в прибрежной зоне (глубины 0.1–0.8 м) сачком (диаметр входного отверстия 30 см) и мальковой волокушей с размером ячеи 4–6 мм, в зимний период – в сублиторали озера (креветочными ловушками в виде зонтика-верши с шестью входными отверстиями). Отловленных рыб фиксировали 4%-м формалином, последующие измерения и анализ желудков проводили в лаборатории. Общая длина проанализированных рыб была в пределах 16–60 мм (табл. 2).
Таблица 2.
Объём материала и биологические характеристики девятииглой колюшки Pungitius pungitius из оз. Кривое (июнь и сентябрь) и эстуария р. Нева (июль) в 2019 г.
| Показатель | Локальность, станция (месяц) | |||||
|---|---|---|---|---|---|---|
| оз. Кривое | эстуарий р. Нева | |||||
| 1 (VI) | 1 (IX) | 2 (VII) | 3 (VII) | 4 (VII) | 5 (VII) | |
| Число рыб, экз. | 12 | 10 | 10 | 10 | 9 | 11 |
| Общая длина, мм | $\frac{{16{\kern 1pt} --{\kern 1pt} 35}}{{23.4 \pm 7.6}}$ | $\frac{{30{\kern 1pt} --{\kern 1pt} 46}}{{36.0 \pm 4.9}}$ | $\frac{{45{\kern 1pt} --{\kern 1pt} 60}}{{50.8 \pm 5.5}}$ | $\frac{{40{\kern 1pt} --{\kern 1pt} 55}}{{44.1 \pm 3.9}}$ | $\frac{{42{\kern 1pt} --{\kern 1pt} 55}}{{49.0 \pm 3.5}}$ | $\frac{{40{\kern 1pt} --{\kern 1pt} 54}}{{45.9 \pm 4.0}}$ |
| Масса рыбы, г | $\frac{{0.03{\kern 1pt} --{\kern 1pt} 0.20}}{{0.14 \pm 0.05}}$ | $\frac{{0.30{\kern 1pt} --{\kern 1pt} 0.90}}{{0.50 \pm 0.17}}$ | $\frac{{0.40{\kern 1pt} --{\kern 1pt} 1.30}}{{0.90 \pm 0.30}}$ | $\frac{{0.50{\kern 1pt} --{\kern 1pt} 1.90}}{{0.90 \pm 0.40}}$ | $\frac{{0.50{\kern 1pt} --{\kern 1pt} 2.00}}{{1.10 \pm 0.50}}$ | $\frac{{0.40{\kern 1pt} --{\kern 1pt} 0.90}}{{0.70 \pm 0.16}}$ |
| Индекс наполнения желудка, ‱ | $\frac{{96{\kern 1pt} --{\kern 1pt} 160}}{{102.9 \pm 14.5}}$ | $\frac{{47{\kern 1pt} --{\kern 1pt} 333}}{{90.0 \pm 58.5}}$ | $\frac{{55{\kern 1pt} --{\kern 1pt} 114}}{{62.0 \pm 13.0}}$ | $\frac{{35{\kern 1pt} --{\kern 1pt} 90}}{{51.0 \pm 17.2}}$ | $\frac{{60{\kern 1pt} --{\kern 1pt} 167}}{{144.0 \pm 26.6}}$ | $\frac{{0{\kern 1pt} --{\kern 1pt} 161}}{{41.7 \pm 13.0}}$ |
Для анализа трофической позиции (оз. Кривое) отбирали колюшку общей длиной 30‒40 мм и окуня (16‒18 см), по 4‒6 экз. каждого вида рыб, и по 20 экз. беспозвоночных – личинок хирономид и подёнок (Ephemeroptera). После отбора всех животных помещали на поднос со льдом и отправляли в лабораторию, где были немедленно отобраны образцы тканей спинных мышц рыб. Беспозвоночных предварительно очищали от ила и детрита, обмывали дистиллированной водой и отбирали по 3‒4 экз. в одну пробу. Все образцы высушивали 48 ч в термостате при температуре 55°С.
Анализ питания рыб
Измерение рыб и обработку содержимого желудков проводили по общепринятой методике (Методическое пособие …, 1974). Для оценки интенсивности питания использовали общий индекс наполнения желудка – выраженное в продецимилле отношение общей массы потреблённых организмов к массе рыбы. Из содержимого желудка выбирали всех животных и растений, объединяли их по крупным таксонам, просчитывали и определяли массу. Дальнейшую идентификацию беспозвоночных до вида или рода проводили под микроскопом. В связи с этим в тексте статьи указаны видовые таксоны, а в сводной табл. 3 представлены суммированные данные по крупным таксонам на уровне семейств и отрядов, в которые были объединены отдельные виды при взвешивании. Рассчитывали процентные доли массы (M, %) основных групп беспозвоночных в общей массе пищевого комка. Учитывали число всех пищевых компонентов (N, экз.) в содержимом желудка, а затем для каждой группы объектов питания рассчитывали средние доли численности (% общего числа компонентов). Средние величины указывали со стандартной ошибкой средних.
Таблица 3.
Состав пищи девятииглой колюшки Pungitius pungitius по численности (N, %) и массе (M, %) в летний период в изученных водоёмах в 2019 г.
| Компонент пищи | Станция | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | ||||||
| N | M | N | M | N | M | N | M | N | M | |
| Oligochaeta | 4 | 1 | 4 | 1 | 5 | 3 | ||||
| Bivalvia | 4 | 7 | ||||||||
| Gastropoda | 5 | 6 | ||||||||
| Copepoda | 5 | 4 | 8 | 2 | 15 | 3 | 11 | 1 | 32 | 10 |
| Cladocera | 5 | 9 | 8 | 1 | ||||||
| Ostracoda | 8 | 4 | ||||||||
| Hydrachnidiae im. | 2 | 1 | 8 | 3 | 5 | 4 | ||||
| Cybaeide | 7 | 3 | ||||||||
| Odonata lv. | 4 | 5 | ||||||||
| Plecoptera lv. | 2 | 4 | ||||||||
| Ephemeroptera lv. | 9 | 9 | 4 | 5 | 9 | 6 | ||||
| Trichoptera lv. | 5 | 13 | ||||||||
| Crambidae lv., im. | 4 | 4 | 8 | 4 | 5 | 5 | ||||
| Coleoptera lv., im. | 9 | 6 | 5 | 2 | 26 | 26 | ||||
| Chironomidae lv., pp. | 18 | 38 | 12 | 37 | 28 | 40 | 9 | 18 | 22 | 54 |
| Ephydridae lv., im. | 4 | 1 | 10 | 21 | ||||||
| Simuliidae, lv., pp. | 4 | 2 | 4 | 4 | 19 | 10 | ||||
| Ceratopogonidae lv. | 9 | 2 | 12 | 15 | 19 | 17 | 10 | 24 | ||
| Corixidae im. | 8 | 9 | ||||||||
| Limoniidae lv., im. | 9 | 4 | 8 | 8 | 15 | 11 | 12 | 6 | ||
| Икра рыб | 5 | 5 | 5 | 5 | 20 | 10 | ||||
| Прочие | 5 | 2 | 6 | 3 | 1 | 2 | ||||
Анализ стабильных изотопов азота
Ткани рыб (колюшки и окуня) и беспозвоночных (по 200‒300 мкг) помещали в оловянные капсулы на аналитические весы Mettler Toledo MX5 (“Mettler Toledo”, США) с точностью ± 1 мкг. Определение изотопного состава тканей беспозвоночных и мышц рыб проводили в Центре коллективного пользования Института проблем экологии и эволюции РАН (г. Москва) на комплексе, состоящем из изотопного масс-спектрометра Thermo Delta V Plus (“Thermo Scientific”, США) и элементарного анализатора Thermo Flash EA 1112. Изотопный состав азота выражали в тысячных долях отклонения (‰) от международного стандарта δNi = [(Ni − Nst)/Nst] × 1000, где N – атомное соотношение тяжёлого и лёгкого изотопов азота в пробе (i) и стандарте (st). Для азота стандартом служит N2 атмосферного воздуха.
Величины δ15N представлены в виде арифметических средних со стандартными отклонениями. Для оценки роли рыб в трофической сети озера рассчитывали их трофическую позицию (TL) по значениям δ15N (Post, 2002): TL = (δ15Nf − δ15Nb)/3.4 + 2, где δ15Nf – соотношение изотопов азота в колюшке или окуне, а δ15Nb – в первичных консументах (личинки хирономид). Для выявления влияния сезона на величины δ15N применяли однофакторный дисперсионный анализ Краскела–Уоллиса. Оценку значимости различий полученных величин между сезонами года проводили в статистическом пакете Statistica 8.0 с использованием критерия Манна–Уитни.
РЕЗУЛЬТАТЫ
Состав пищи девятииглой колюшки в изученных водоёмах
Состав пищи в летний период, наиболее продуктивный в изученных водоёмах, демонстрирует наибольшее разнообразие пищевого спектра колюшки. В этот период средние индексы наполнения желудков рыб из оз. Кривое составили 102.9‱, а у особей с различных станций в эстуарии р. Нева варьировали от 42 до 144‱ (табл. 2). Индексы наполнения желудков рыб со станции 5 (Графская бухта) в среднем были наименьшими, у одной особи желудок был пуст (табл. 2). В рационе колюшки оз. Кривое и эстуария р. Нева отмечена 21 группа водных животных (табл. 3). Значимой разницы между количеством и массой различных компонентов питания рыб, отловленных на разных станциях в эстуарии р. Нева и оз. Кривое, не обнаружено (тест Краскела–Уоллиса, p > 0.05), при этом значение разных групп беспозвоночных в питании рыб существенно различалось. В пище колюшки оз. Кривое и со всех станций эстуария р. Нева доминировали личинки хирономид, составляя 18‒54% общей массы пищи (табл. 3). Доля по численности этих организмов также была высока (9‒22%), в желудке одной рыбы регистрировали до 23 экз. личинок хирономид. Среди них у рыб из обоих водоёмов доминировали несколько видов: Cryptochironomus viridulus Fabricius, Polypedilum nubeculosum Meigen, Endochironomus albipennis Meigen и Tanytarsus gregarius Kieffer. Другие виды (Anatopynia plumipes (Fries), Micropsectra praecox (Meigen) и Pagastiella orophila Edwards) были единичны. В среднем по обоим водоёмам доля хирономид (по массе) в пище колюшки составила 37.1 ± 5.8%, эта группа составляет основу питания колюшки (табл. 4). Личинки мокрецов также были значимы в питании девятииглой колюшки, составляя в среднем 11.6 ± 4.6%. Наиболее часто встречающимся видом среди мокрецов был Palpomyia lineate (Meigen).
Таблица 4.
Общий состав пищи девятииглой колюшки Pungitius pungitius по численности (N, %) и массе (M, %) и значимость пищевых компонентов в изученных водоёмах
| Группа пищевых компонентов | N | SE | M | SE | Значимость |
|---|---|---|---|---|---|
| Chironomidae | 15.8 | 2.3 | 37.1 | 5.8 | Д |
| Ceratopogonidae | 8.0 | 2.1 | 11.6 | 4.6 | Д |
| Cladocera + Copepoda | 16.6 | 6.4 | 5.8 | 3.5 | Суб Д |
| Ephemeroptera | 4.4 | 2.0 | 4.0 | 1.7 | Суб Д |
| Coleoptera | 8.0 | 4.8 | 6.9 | 4.9 | Суб Д |
| Limoniidae | 8.8 | 2.5 | 5.7 | 1.8 | Суб Д |
| Ephydridae | 2.2 | 1.4 | 4.3 | 4.1 | Суб Д |
| Икра рыб | 6.0 | 3.7 | 4.0 | 1.9 | Суб Д |
| Trichoptera | 1.0 | 1.0 | 3.2 | 3.2 | Р |
| Crambidae | 3.4 | 1.5 | 2.6 | 1.0 | Р |
| Oligochaeta | 4.6 | 2.7 | 1.5 | 1.1 | Р |
| Mollusca | 2.0 | 1.8 | 2.5 | 2.5 | Р |
| Simuliidae | 5.4 | 3.5 | 3.2 | 1.8 | Р |
| Corixidae | 1.6 | 1.6 | 1.7 | 1.7 | Р |
| Ostracoda | 1.6 | 1.6 | 0.8 | 0.8 | Ед |
| Hydrachnidiae | 3.0 | 1.5 | 1.6 | 0.8 | P |
| Cybaeide | 1.4 | 1.4 | 0.7 | 0.7 | Ед |
| Odonata | 0.8 | 0.8 | 1.0 | 1.0 | P |
| Plecoptera | 0.4 | 0.4 | 0.8 | 0.8 | Ед |
| Прочие | 5.0 | 2.5 | 1.0 | 0.6 | P |
Представители водных насекомых, таких как подёнки, лимонииды (Limoniidae), жуки Coleoptera и эфидриды (Ephydridae), входили в группу субдоминантных компонентов, составляя в пище рыб в среднем 4–7% её массы (табл. 4). В пище колюшки оз. Кривое отмечены личинки подёнок Nigrobaetis niger (L.) и Caenis undosa Tiensuu и лимониид Dicranota bimaculata (Schummel), личинки и имаго жуков Helophorus aquaticus L. и Rhantus fennicus Hulden. Икра рыб также в среднем составляла 4% массы пищевого комка девятииглой колюшки.
Важный вклад в питание колюшки вносили и планктонные ракообразные – копеподы семейств Chydoridae и Bosminidae и кладоцеры Macrocyclops и Cyclops sp., они доминировали по численности (17 ± 6.4%) и достигали 5.7% массы пищевого комка. Планктонные ракообразные Macrocyclops albidus (Jurine), Cyclops kolensis Lilljeborg, Mesocyclops crassus (Fischer) и Acanthocyclops viridis (Jurine) преобладали в пище колюшки, отловленной на наиболее эвтрофной станции в эстуарии р. Нева – станции 5 (табл. 3).
Остальные представители насекомых (Trichoptera, Crambidae, Simuliidae и Corixidae) были второстепенными компонентами в питании колюшки, хотя у рыб с отдельных станций в эстуарии р. Нева их доля по массе была высока (например, личинок и куколок мошек на станции 4). Гусеницы и взрослые особи бабочек семейства Crambidae: прудовая Nymphula nitidulata (Hufnagel), кувшинковая Elophila nymphaeata (L.) и телорезовая Paraponyx stratiotata L. огнёвки встречались в пище колюшки оз. Кривое. Редкими компонентами её питания были мелкие двустворчатые моллюски семейства Sphaeriidae и брюхоногие моллюски семейства Bithyniidae, а также олигохеты Tubifex tubifex (O.F. Müller) у рыб из эстуария р. Нева и Lumbriculus variegates (Muller) у особей из оз. Кривое. Единично и только у колюшки с отдельных станций в эстуарии р. Нева отмечены личинки веснянок (Isoperla diffomis Klapalek), ручейников (Agraylea multipunctata Curtis), ракушковые рачки и изоподы Asellus aquaticus L. В пище колюшки обнаружены также случайно попадающие в воду наземные насекомые – муравьи семейства Formicidae и мухи семейства Muscidae. В вегетационный период в пище колюшек были отмечены растительные остатки (мхи, диатомовые и нитчатые водоросли).
Сезонная динамика состава пищи колюшки (оз. Кривое)
Питание колюшки оз. Кривое в осенний период (сентябрь) и тем более в зимний период (подо льдом) было менее разнообразно, чем летом (рис. 2). Осенью в питании уменьшалась доля личинок и куколок хирономид (с 38 до 22% массы), но возрастала доля других двукрылых (мошек Simuliidae) (28%) и планктонных ракообразных (22%). Также несколько увеличивалась доля личинок бабочек семейства Crambidae и водяных клещей. В феврале и апреле в пище колюшек были представлены в основном личинки хирономид Sergentia coracina (Zetterstedt) и икра/личинки ряпушки Coregonus albula (L.).

Трофическая позиция колюшки
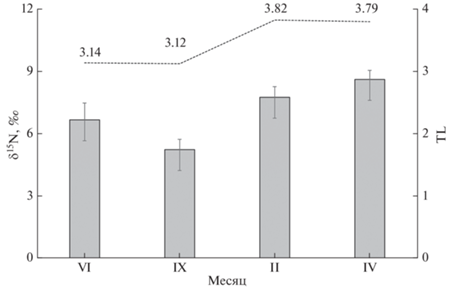
Средние величины δ15N в мышцах девятииглой колюшки оз. Кривое варьировали от 5.2 до 8.6‰, значимо (H = 13.8, р = 0.003) возрастая гидрологической зимой, т.е. в подлёдный период (рис. 3). Между этим и летним периодом различия были недостоверны (p > 0.05), в то время как разница между величинами δ15N в сентябре и в подлёдный период (февраль и апрель), согласно критерию Манна–Уитни, была значимой (p = 0.03). Величины δ15N у первичных консументов (личинок хирономид и подёнок) в течении года варьировали от 1.4 до 2.8‰. Рассчитанная для колюшки позиция в трофической сети была сходной в течение тёплого сезона (лето–осень) – 3.1 и несколько повышалась в холодный сезон (подо льдом) – 3.8. Таким образом, колюшку можно отнести к типичным зоофагам, её позиция в трофической сети соответствовала третьему уровню. В зимний период она потребляла хирономид и икру ряпушки, по величинам δ15N приближаясь к хищнику второго порядка – окуню (δ15N 9.1‰, TL 3.85).

ОБСУЖДЕНИЕ
В субарктическом олиготрофном озере девятииглая колюшка питалась преимущественно личинками (и в меньшей степени куколками) мелких двукрылых (хирономид, мокрецов, мошек) и других насекомых. Доминирование бентосных компонентов в питании субарктических рыб отмечали и ранее как характерную черту для северных озёр (Eloranta et al., 2010; Berezina et al., 2018), что зачастую связано с бедностью кормовой базы (в том числе зоопланктона) в этих водоёмах. Спектр питания девятииглой колюшки в бассейне Чебоксарского и Куйбышевского водохранилищ, а также рек Камчатки и Курильских о-вов в основном состоял из личинок мелких хирономид, мошек и других донных организмов (Пичугин и др., 2004; Травина, Введенская, 2009; Травина, Ярош, 2009; Логинов и др., 2014). Видовое разнообразие хирономид в пище было наиболее высоким и включало виды, которые доминировали в зообентосе (Травина, Ярош, 2009). Сравнение спектра питания девятииглой колюшки в эстуарии р. Чёрная (Кандалакшский залив, Белое море) с составом сообществ беспозвоночных показало, что там, где основу бентоса составляли личинки хирономид, эти организмы и доминировали в пищевом комке (Пономарев и др., 2003).
Анализ полученных результатов по питанию девятииглой колюшки в прибрежной зоне эстуариев Балтийского моря, включая наши данные по эстуарию р. Нева, в Ботническом заливе (Thorman, Wiederholm, 1983) и эвтрофном Куршском заливе (Науменко и др. 2020) свидетельствует о смешанном характере питания вида как планктонными, так и бентосными беспозвоночными. Планктонные ракообразные, главным образом веслоногие Eudiaptomus graciloides (Lilljeborg) (30% по численности), ветвистоусые Daphnia longispina (O.F. Müller) (29%) и их яйца, часто доминировали в питании колюшки Куршского залива (Науменко и др., 2020). Активное потребление колюшкой планктонных ракообразных (ветвистоусых, веслоногих) отмечено и для отдельных эвтрофных станций эстуария р. Нева. В питании колюшки рек Охотского моря (р. Большая) (Максименков, Токранов, 1994) была выявлена пространственная динамика в смене пищевых объектов – доминирование личинок хирономид в нижнем течении реки и переход на питание ракообразными (бокоплавы и мизиды) в приустьевом участке эстуария.
Отмечали также сезонное возрастание (в осенний период) доли планктонных ракообразных в пище колюшки северного озера (оз. Кривое) и других водоёмов – р. Большая, р. Малая Юнга, Куршский залив (Травина, Ярош, 2009; Логинов и др., 2014; Науменко и др., 2020). По-видимому, доминирование тех или иных групп зообентоса и зоопланктона в рационе рыб связано с их доступностью в водоёмах. Таким образом, для состава пищи девятииглой колюшки характерны существенные различия между эстуариями, реками и озёрами. Однако в большинстве водоёмов она потребляет личинок двукрылых, в меньшей степени других насекомых и планктонных ракообразных.
Доминирование личинок хирономид в питании колюшки во многих водоёмах и в разные периоды года может свидетельствовать о её пищевой избирательности этой группы водных животных. Экспериментальное исследование вкусовых предпочтений девятииглой колюшки подтверждает эту особенность (Михайлова, Касумян, 2015) – была выявлена высокая привлекательность агар-агаровых гранул со вкусом личинок хирономид для колюшки из москворецкой, беломорской и охотоморской популяций.
На основании анализа изотопного состава азота в мышцах девятииглая колюшка располагается в трофической сети водоёма в пределах третьего трофического уровня (TL 3.12‒3.82), приближаясь в зимний период к уровню хищников второго порядка (3.85). Сезонной особенностью спектра питания колюшки является переход от потребления разнообразной животной пищи с примесью растительных компонентов в тёплый летне-осенний период года к хищничеству в подлёдный период (зимой и весной), что приводит к некоторому увеличению её трофической позиции. Трофическая позиция этой мелкоразмерной рыбы (длина до 6 см) оказалась близка к другим хищным рыбам (окунь), что можно объяснить интенсивным потреблением (20–30% по массе) икры/личинок рыб (рис. 2). Икра рыб была обнаружена в питании девятииглой колюшки не только для изученных нами водоёмах, но для многих других (Введенская, 1993; Максименков, Токранов, 1994). Такая же черта характерна и для питания близкого вида – трёхиглой колюшки Gasterosteus aculeatus L. (Ярош и др., 2009).
Таким образом, в изученных нами водоёмах девятииглая колюшка, как и большинство видов рыб в водоёмах России, является зоофагом со смешанным типом питания, основу которого составляют личинки мелких двукрылых насекомых и ракообразные. Перенося вещества и энергию от первичных консументов (зоопланктон, зообентос) на другой более высокий трофический уровень (хищные рыбы, птицы, млекопитающие) этот вид может быть важным звеном трофической сети водных экосистем арктической зоны с низким богатством ихтиофауны, особенно при его высокой численности. Будучи при этом очень пластичной в питании, девятииглая колюшка может конкурировать за пищевые ресурсы с молодью промысловых рыб (окунь, ряпушка). Питание икрой рыб приближает колюшку по положению в трофической сети к высшим хищникам.
Сравнение питания колюшки разных размерных групп (в частности молоди и взрослых особей) в рамках данного исследования не проводили из-за ограниченности имеющегося материала, но в дальнейшем планируется проведение таких исследований. По крайней мере в летний период, когда был выявлен широкий спектр питания колюшки, можно ожидать различия в питании особей разного размера. В дальнейшем также необходим анализ пищевых предпочтений девятииглой колюшки в связи с составом и структурой сообществ планктона и бентоса в водоёме, что позволит выявить причины её предпочтительного питания теми или иными организмами.
Список литературы
Антипин М.А. 2018. Бурокрылая ржанка Pluvialis fulva ловит и ест девятииглую колюшку Pungitius pungitius // Рус. орнитол. журн. Т. 27. № 1664. С. 4408–4409.
Аськеев О.В., Аськеев И.В., Ананин А.Н., Тишин Д.В. 2010. Обнаружение девятииглой колюшки (Pungitius pungitius Linnaeus, 1758) в бассейне р. Камы // Поволж. экол. журн. № 1. С. 103–106.
Атлас пресноводных рыб России. 2003. М.: Наука, 253 с.
Березина Н.А., Литвинчук Л.Ф., Максимов А.А. 2021. Связь пищевого спектра рыб с составом зоопланктона и зообентоса в субарктическом озере // Биология внутр. вод. № 4. С. 406–416. https://doi.org/10.31857/S0320965221040069
Бобырь И.Г., Антипин М.А. 2017. Первый зарегистрированный случай питания дутыша Calidris melanotos рыбой – девятииглой колюшкой Pungitius pungitius // Рус. орнитол. журн. Т. 26. № 1498. С. 3838–3839.
Бойко Н.С., Бианки В.В. 2009. Особенности питания некоторых уток Белого моря // Рус. орнитол. журн. Т. 18. № 474. С. 540‒542.
Бугаев В.Ф., Вронский Б.Б., Заварина Л.О. и др. 2007. Рыбы реки Камчатка. Петропавловск-Камчатский: Изд-во КамчатНИРО, 459 с.
Введенская Т.Л. 1993. Пищевые рационы рыб в литорали оз. Дальнего // Исследования биологии и динамики численности промысловых рыб камчатского шельфа. № 2. С. 130–139.
Введенская Т.Л. 2008. Некоторые черты биологии рыб бассейна р. Таловки (Северо-Западная Камчатка) // Матер. IX науч. конф. “Сохранение биоразнообразия Камчатки и прилегающих морей”. Петропавловск-Камчатский: Камчатпресс. С. 41–44.
Дегтярев В.Г., Слепцов С.М., Пшенников А.Е. 2013. Ихтиофагия восточной популяции стерха (Grus leucogeranus) // Зоол. журн. Т. 92. № 5. С. 588–595. https://doi.org/10.7868/S0044513413030070
Клевакин А.А., Логинов В.В., Морева О.А., Тарбеев М.Л. 2011. Биологические особенности девятииглой колюшки Pungitius pungitius (Linnaeus, 1758) локальной популяции реки Ушаковка // Рос. журн. биол. инвазий. № 2. С. 86–106.
Логинов В.В., Клевакин А.А., Морева О.А. и др. 2014. Морфологическая характеристика и питание девятииглой колюшки (Pungitius pungitius Linnaeus 1758) бассейна Чебоксарского водохранилища // Рос. журн. биол. инвазий. № 2. С. 96–109.
Максименков В.В., Токранов A.M. 1994. Питание девятииглой колюшки Pungitius pungitius (Gasterosteidae) в эстуарии и нижнем течении реки Большая (Западная Камчатка) // Вопр. ихтиологии. Т. 34. № 5. С. 697–702.
Методическое пособие по изучению питания и пищевых взаимоотношений рыб в естественных условиях. 1974. М.: Наука, 244 с.
Михайлова Е.С., Касумян А.О. 2015. Вкусовые предпочтения и пищевое поведение девятииглой колюшки Pungitius pungitius трёх географически удалённых популяций // Вопр. ихтиологии. Т. 55. № 5. С. 541–564. https://doi.org/10.7868/S0042875215050112
Науменко Е.Н., Ушакова А.Ю., Голубкова Т.А. 2020. Питание сеголеток рыб Куршского залива Балтийского моря в 2016 году // Тр. ВНИРО. Т. 179. С. 60–77. https://doi.org/10.36038/2307-3497-2020-179-60-77
Пичугин М.Ю., Сидоров Л.К., Стыгар В.М. 2004. Биологические и морфологические особенности девятииглых колюшек рода Pungitius (Gasterosteiformes) Курильских островов // Вопр. ихтиологии. Т. 44. № 1. С. 15–26.
Погорелова Д.П., Хивренко Д.Ю., Улатов А.В. 2020. Некоторые сведения о питании рыб в литоральной зоне озера Халактырского // Матер. XXI науч. конф. “Сохранение биоразнообразия Камчатки и прилегающих морей”. Петропавловск-Камчатский: Камчатпресс. С. 106–109.
Пономарев С.А., Бурковский И.В., Столяров А.П., Новиков Г.Г. 2003. Особенности питания трех- и девятииглой колюшек с учетом их микробиотопического распределения в эстуарии (Кандалакшский залив, Белое море) // Усп. соврем. биологии. Т. 123. № 6. С. 609–617.
Светочев В.Н., Светочева О.Н. 2015. Питание и пищевые отношения настоящих тюленей в Белом море // Вестн. КНЦ РАН. Т. 3. № 22. С. 93–101.
Травина Т.Н., Введенская Т.Л. 2009. Роль хирономид (Diptera, Chironomidae) в питании молоди лососей и других видов рыб в реке Большая (Западная Камчатка) // Евразиат. энтомол. журн. Т. 8. Прил. 1. С. 102–108.
Травина Т.Н., Ярош Н.В. 2009. К вопросу о питании девятииглой колюшки Pungitus pungitius в нижнем течении р. Большая в 2007–2008 гг. // Исслед. вод. биол. ресурсов Камчатки и сев.-зап. части Тихого океана. № 15. С. 88–93.
Хаменкова Е.В. 2011. Трофические взаимоотношения рыб оз. Киси бассейна реки Ола (Магаданская область) // Чтения памяти В.Я. Леванидова. Вып. 5. Владивосток: Дальнаука. С. 563–570.
Черешнев И.А. 2008. Пресноводные рыбы Чукотки. Магадан: Изд-во СВНЦО ДВО РАН, 324 с.
Ярош Н.В., Травина Т.Н., Введенская Т.Л. 2009. Питание трехиглой колюшки Gasterosteus aculeatus (Leiurus) в нижнем течении реки Большая // Исслед. вод. биол. ресурсов Камчатки и сев.-зап. части Тихого океана. № 15. С. 94–102.
Berezina N.A., Golubkov S.M., Gubelit J.I. 2005. Grazing effects of alien amphipods on macroalgae in the littoral zone of the Neva Estuary (eastern Gulf of Finland, Baltic Sea) // Oceanol. Hydrobiol. Stud. V. 34. Suppl. 1. P. 63–82.
Berezina N.A., Strelnikova A.P., Maximov A.A. 2018. The benthos as the basis of vendace, Coregonus albula, and perch, Perca fluviatilis, diets in an oligotrophic sub-Arctic lake // Polar Biol. V. 41. № 9. P. 1789–1799. https://doi.org/10.1007/s00300-018-2319-0
Cameron J.N., Kostoris J., Penhale P.A. 1973. Preliminary energy budget of the ninespine stickleback (Pungitius pungitius) in an Arctic Lake // J. Fish. Res. Board Can. V. 30. № 8. P. 1179–1189. https://doi.org/10.1139/f73-188
Demchuk A.S., Uspenskiy A., Golubkov S.M. 2021. Abundance and feeding of fish in the coastal zone of the Neva Estuary, eastern Gulf of Finland // Boreal Environ. Res. № 26. P. 1–16.
Eloranta A.P., Kahilainen K.K., Jones R.I. 2010. Seasonal and ontogenetic shifts in the diet of Arctic charr Salvelinus alpinus in a subarctic lake // J. Fish Biol. V. 77. № 1. P. 80–97. https://doi.org/10.1111/j.1095-8649.2010.02656.x
Post D.M. 2002. Using stable isotopes to estimate trophic position: Models, methods, and assumptions // Ecology. № 83. № 3. P. 703–718. https://doi.org/10.2307/3071875
Thorman S., Wiederholm A.-M. 1983. Seasonal occurrence and food resource use of an assemblage of nearshore fish species in the Bothnian Sea, Sweden // Mar. Ecol. Prog. Ser. V. 10. № 3. P. 223–229. https://doi.org/10.3354/meps010223
Winberg G.G., Alimov A.F., Boullion V.V. et al. 1973. Biological productivity of two subarctic lakes // Freshw. Biol. V. 3. № 2. P. 177–197. https://doi.org/10.1111/J.1365-2427.1973.TB00071.X
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии