

Исследование Земли из Космоса, 2021, № 4, стр. 28-45
Сезонная и межгодовая изменчивость поля концентрации хлорофилла-а в Беринговом море по спутниковым данным
К. К. Кивва a, *, А. А. Кубряков b
a Федеральное государственное бюджетное научное учреждение
“Всероссийский научно-исследовательский институт рыбного хозяйства и океанографии” (ФГБНУ “ВНИРО”)
Москва, Россия
b Федеральное государственное бюджетное учреждение науки Федеральный исследовательский центр
“Морской гидрофизический институт РАН” (ФГБУН ФИЦ МГИ РАН)
Севастополь, Россия
* E-mail: kirill.kivva@gmail.com
Поступила в редакцию 25.02.2021
Аннотация
В работе представлена характеристика пространственно-временной изменчивости поля концентрации хлорофилла-а в Беринговом море и прилегающей части Тихого океана, основанная на данных MODIS за 2003–2019 гг. Описана среднемноголетняя сезонная изменчивость концентрации хлорофилла-а, выявлены характерные сроки “цветения” фитопланктона в различных частях рассматриваемой акватории. Проанализирована межгодовая изменчивость сроков весеннего “цветения” в трех районах: в западной части моря к северо-востоку от Камчатского пролива, в северной части моря к югу от м. Наварин и в центральной части моря. При этом первые два из них близки по средним многолетним характеристикам. Показано, что сроки начала “цветения” в этих двух районах имели различную межгодовую изменчивость, а их размах в обоих случаях составил около 2 мес. В западной части моря наблюдается смещение “цветения” на более ранние сроки (март–апрель), а в северном районе сроки “цветения” наоборот сдвигаются к маю–июню. В центральной части моря весеннее “цветение” выражено значительно слабее, чем в остальных рассматриваемых районах, начинается в мае–июне и может продолжаться несколько недель. При этом относительно высокое содержание хлорофилла-а может наблюдаться там и в августе–октябре. В отдельные годы в центральном районе моря наблюдалось осеннее “цветение”, преобладающее по интенсивности (максимум концентрации) над “весенним”. Такая ситуация больше характерна для открытой части Тихого океана, а не для Берингова моря. Для всей рассматриваемой акватории выполнено сравнение хода развития весеннего “цветения” в 2010 и 2015 гг., которое показало, что пространственное развитие “цветения” в различные годы также может существенно отличаться. Межгодовые отличия в сроках начала “цветения” в различных районах моря связаны с особенностями физических процессов, определяющих формирование устойчивой стратификации вод (таяние льда или прогрев при слабом ветровом воздействии), а также могут быть связаны с изменчивостью течений.
ВВЕДЕНИЕ
Новое первичное органическое вещество (ОВ), синтезируемое фитопланктоном, служит основой пищевой цепи в морях и океанах. Бо́льшая часть этого ОВ на акваториях умеренных и высоких широт формируется в определенные периоды года, когда происходит быстрый и относительно продолжительный рост биомассы фитопланктона. Такие явления называются “цветениями” фитопланктона. Их возникновение связано с совпадением ряда условий, необходимых для активной вегетации фитопланктона: достаточная освещенность и наличие минеральных питательных солей (Sverdrup, 1953). Для накопления этой биомассы также необходимо, чтобы скорость ее прироста превышала скорость потребления фитопланктона зоопланктоном (Behrenfeld and Boss, 2014).
По классической теории Харольда Свердрупа, весеннее “цветение” возникает в субарктических и арктических морях, когда после зимнего возобновления запаса минеральных питательных солей в эвфотическом слое формируется устойчивая структура вод с сезонным пикноклином, располагающимся выше глубины критической освещенности (Sverdrup, 1953). То есть такого горизонта, при равномерном перемешивании до которого среднее количество света, получаемое клеткой фитопланктона, вовлеченной в это перемешивание, позволяет ей сохранять превышение темпов фотосинтеза над темпами деструкции ОВ при дыхании. Наступление таких условий требует уменьшения плотности верхнего слоя водной толщи (увеличения плавучести), и произойти это может за счет нескольких процессов: таяние льда, солнечный прогрев или поступление значительного количества материкового стока. В случае опреснения поверхностного слоя при таянии морского льда возникает прикромочное “цветение” фитопланктона (Дружкова, 2018; Perrette et al., 2011). В Беринговом море такое “цветение” наблюдается обычно в апреле−июне (Sigler et al., 2014). В отсутствие влияния материкового стока на акватории, где в холодный период года льда не было, устойчивая структура обязана своим появлением солнечному прогреву верхнего слоя вод. Такое “цветение”, иногда называемое классическим (Дружкова, 2018), обычно начинается несколько позже: в Беринговом море – в конце мая или в июне (Sigler et al., 2014). Вполне возможны ситуации, когда после прикромочного “цветения” формируется классическое. Причем они могут сливаться в единый период активной вегетации фитопланктона, а могут быть разнесены во времени (Niebauer et al., 1995; Sigler et al., 2014; Stabeno et al., 2001).
“Цветение” начинает постепенно затухать и прекращается, когда запас хотя бы одного из необходимых фитопланктону элементов иссякает или когда темп потребления биомассы фитопланктона зоопланктоном начинает превышать темп его прироста (Behrenfeld and Boss, 2014). При этом ниже верхнего продукционного слоя питательные соли (минеральные формы биогенных элементов) не расходуются, то есть формируется нутриклин – слой относительно резкого роста концентрации минеральных форм биогенных элементов. Поэтому на поздних стадиях “цветения” и после его затухания во многих районах Мирового океана, в частности полярных и субполярных, максимум биомассы фитопланктона смещается в район нутриклина, формируется подповерхностный максимум хлорофилла. Кроме того, часть нового ОВ, синтезированного в течение весеннего “цветения”, в виде детрита оседает в толще воды и, постепенно разлагаясь, частично вновь переходит в минеральную форму. Таким образом, ниже нутриклина биогенные элементы после “цветения” могут даже накапливаться. В случае если эпизодические события перемешивания водной толщи (например, шторма́) или подъема подповерхностных вод приводят к поступлению новых порций питательных веществ в эвфотический слой, вегетация фитопланктона может возобновиться (такие события иногда называют “вспышками” продуцирования или биомассы). На многих акваториях почти ежегодно наблюдается осеннее “цветение” фитопланктона, связанное с похожим процессом, поскольку осенью шторма́ усиливаются, а также начинается конвективное перемешивание (Sigler et al., 2014; Mikaelyan et al., 2017, 2018). Это приводит к возобновлению запаса минеральных питательных солей в эвфотическом слое. Перерывы в процессе перемешивания в то время, когда освещенность акватории еще значительна, могут приводить к повторному (или даже многократному) стратифицированию верхнего слоя вод и, как и в весеннее время, к осеннему “цветению” (Хен и др., 2015).
Несмотря на ежегодную повторяемость “цветений”, период их наступления не всегда строго соблюдается. Это может быть связано с особенностями протекания физических процессов в пределах каждого отдельно взятого года. Например, таяние морского льда (в тех районах, где присутствовал лед) и солнечный нагрев поверхности в разные годы могут начаться в разное время, протекать с различной интенсивностью или прерываться. Кроме того, формирование устойчивой структуры вод может быть нарушено ветровым перемешиванием. Поэтому сроки “цветений” год от года заметно сдвигаются, а объем их общего результата (например, суммарная продукция ОВ за период “цветения”) и качественный состав продуцируемого ОВ может меняться. Это, в свою очередь, может повлечь за собой изменения в состоянии зоопланктона и организмов более высоких трофических уровней, в том числе промысловых (Sigler et al., 2014; Hunt et al., 2011). Например, в случае позднего весеннего “цветения” может снижаться выживаемость молоди рыб (Platt et al., 2003; Hunt et al., 2011). Также от сроков “цветения” может зависеть пропорция нового ОВ, поступающего из пелагиали на дно (Coyle et al., 2011; Hunt et al., 2002; Hunt et al., 2011). В результате при разных условиях “цветения” бентосная часть экосистемы может получать различное количество пищи (Coyle et al., 2011; Hunt et al., 2011; Wassman, Reigstad, 2011).
Спутниковые данные высокого разрешения позволяют исследовать многие процессы и явления в океане (Алексанин и Ким, 2015; Андреев, 2017; Бондур и др., 2017, 2021; Бондур, Зубков, 2005). В исследованиях изменчивости поля концентрации хлорофилла-а (Хл) на поверхности Мирового океана широко используются спутниковые радиометрические наблюдения. В частности, они позволили изучить основные особенности биооптических характеристик многих морей России, включая Баренцево, Балтийское, Черное и Азовское (Kopelevich et al., 2008; Копелевич и др., 2018). В нескольких недавних работах выполнен анализ особенностей распределения хлорофилла-а в Охотском море (Андреев, Жабин, 2012; Жабин и др., 2017; Цхай, Хен, 2016, 2020). Однако для акватории Берингова моря многолетние данные обобщены только в работах (Okkonen et al., 2004; Iida, Saitoh, 2007; Andreev et al., 2020). В работе (Iida, Saitoh, 2007) приводится анализ сезонной и пространственной изменчивости поля Хл в 1998–2002 гг. Показано, что в отличие от шельфовых районов и западной части моря в центральной глубоководной части моря в течение всего года наблюдаются относительно низкие значения Хл. При этом, по крайней мере, в шельфовых районах западной части моря и вдоль восточной части материкового склона Хл в значительной степени зависит от особенностей циркуляции вод (Andreev et al., 2020; Okkonen et al., 2004).
Все перечисленные в отношении Берингова моря работы либо основаны на нескольких годах наблюдений, либо посвящены локальным особенностям поля Хл. То есть на текущий момент еще нет обобщения доступных спутниковых данных по этой акватории. Цель этой статьи – дать общую характеристику пространственно-временной структуры поля Хл на поверхности Берингова моря и прилегающей части Тихого океана, а также основных черт ее сезонной и межгодовой изменчивости на основе долговременного массива спутниковых оптических данных.
МАТЕРИАЛЫ И МЕТОДЫ
В основу работы положены спутниковые данные по Хл спектрорадиометра MODIS спутника Aqua за 2003–2019 гг. (исходное разрешение 4 км × 4 км × 8 дней) (https://oceancolor.gsfc.nasa.gov/) для акватории, ограниченной 162° в.д., 156° з.д., 46°–67° с.ш.. Использован стандартный продукт, полученный по алгоритму OC3M, данные загружены из архива NASA’s OceanСolor Web (O’Reily et al., 1998).
Отметим, что в прибрежных районах стандартный алгоритм может давать существенно завышенные абсолютные значения Хл из-за влияния относительно высоких концентраций окрашенного растворенного ОВ (ОРОВ) (Kopelevich et al., 2008). Известно, что для восточного шельфа Берингова моря стандартный алгоритм завышает низкие значения Хл и занижает высокие ((Naik et al., 2013, 2015). Завышение низких значений (ниже 0.5 мг/м3) там связано именно с влиянием ОРОВ (Naik et al., 2013). В абсолютном выражении разница между измеренной in situ и оцененной по спутниковым данным Хл может достигать 4–4.5 мг/м3 в мае и июне при медианном значении ниже 0.5 мг/м3, а в июле−октябре обычно не превышает 2 мг/м3 с медианой, близкой к 0 мг/м3 (Naik et al., 2013). В работе (Naik et al., 2015) получено среднее отношение между измеренной и оцененной по алгоритму OC3M величиной Хл ≈ 2.3. Соответствующее сравнение для северной части моря для мая–июня 2007 г. дало значение 2.04 (Lee et al., 2018), а для западной части моря для августа 2013 г. ≈1 (Салюк и др., 2016). Для центральной глубоководной части моря подобные оценки, насколько нам известно, отсутствуют. При этом высокие отклонения от in situ значений Хл чаще всего наблюдаются для концентрации меньше 0.5 мг/м3 (Lee et al., 2018; Naik et al., 2013), а особенности пространственного распределения значений Хл, оцененных по разным алгоритмам, в Беринговом море полностью совпадают (например, Lee et al., 2018).
Существует множество подходов к определению начала и окончания “цветения” фитопланктона с использованием спутниковых данных, разработанных различными авторами (Friedland et al., 2018). В связи с поставленной целью (общая характеристика процесса и его динамики) в данной работе “цветение” фитопланктона определялось методом экспертной оценки по данным о сезонной динамике Хл. Под “цветением” здесь и далее подразумевается ситуация превышения Хл значения, характерного для холодного сезона (ноябрь–февраль) на срок более 16 дней (двух интервалов по 8 дней). В качестве таких пороговых значений выбраны 0.5 мг/м3 для глубоководной части Берингова моря, удаленной от бровки шельфа более чем на 200 км, и 1.0 мг/м3 для шельфовых и прибрежных областей. Таким образом, сроки начала и окончания “цветения” в данной работе определены субъективно. Однако использованный подход позволил сосредоточиться на пространственной упорядоченности особенностей поля Хл без осреднения данных по относительно крупным ячейкам, как, например, в работе (Friedland et al., 2018). Для определения времени наступления пика цветения по среднемесячным картам, полученным по всему использованному массиву, рассчитан месяц, соответствующий максимуму Хл в каждом узле сетки массива.
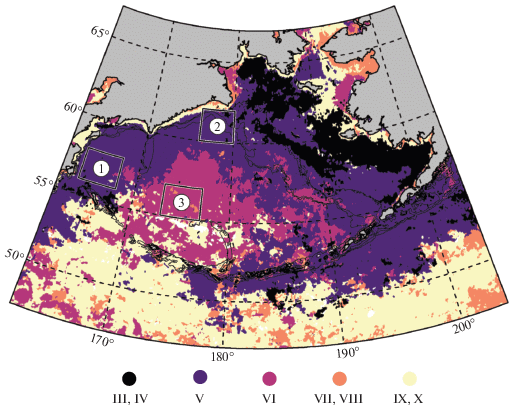
Для описания характеристик межгодовой изменчивости сезонного хода Хл выбраны три основных района: район в западной части моря к югу от Олюторского залива к северо-востоку от Камчатского пролива (западный: 56°–58° с.ш., 164°–168° в.д.) район на севере глубоководной части моря к югу от м. Наварин (северный: 60°–62° с.ш., 176°–180° в.д.), и район в центральной части моря (центральный: 55°–57° с.ш., 173°–177° в.д.). Расположение районов показано на рис. 1 и 6.
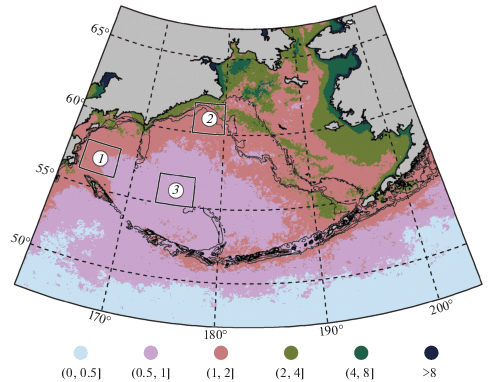
РЕЗУЛЬТАТЫ
Средняя многолетняя концентрация
Наиболее высокие средние многолетние значения Хл в Беринговом море приурочены к материковым побережьям Азии и Северной Америки, а также наблюдаются: в центре северной части шельфа; на восточном шельфе от каньона Наварин до о. Св. Матвея; в центральном шельфовом районе над глубинами порядка 70 м; вдоль материкового склона на участке от каньона Беринга до каньона Прибылова (рис. 1). При этом нужно принять во внимание, что за период с ноября по февраль данные по концентрации Хл для большей части исследуемой акватории полностью отсутствуют из-за облачности или морского льда. Поэтому средние многолетние значения концентрации там несколько завышены в связи с тем, что в них не учтены зимние данные, когда концентрация Хл в пределах исследуемой акватории предположительно наименьшая.
Сезонная динамика поля концентрации хлорофилла-а
Среднемноголетний сезонный ход Хл на исследуемой акватории характеризуется следующими особенностями. Относительно высокие значения Хл (выше 2 мг/м3) раньше всего наблюдается в районе к югу и юго-востоку от о. Нунивак и в Бристольском заливе в марте (рис. 2). Максимальные средние многолетние значения Хл в этом месяце там превышают 2 мг/м3, а на небольшой акватории даже 4 мг/м3, что свидетельствует о начале “цветения” фитопланктона в пределах исследуемой акватории. При этом большая часть восточного шельфа (приблизительно вся область севернее 60° с.ш. над глубинами менее 100 м) в марте чаще всего еще покрыта льдом (Baker et al., 2020). Вне восточного шельфа в это время среднее многолетнее значение Хл почти всюду составляет меньше 1 мг/м3, а на большей части акватории лежит в пределах 0.2–0.5 мг/м3.
Рис. 2.
Средняя многолетняя концентрация хлорофилла-а в Беринговом море в различные месяцы (мг/м3). Месяцы обозначены римскими цифрами в верхней части каждой панели (III – март, … X – октябрь). Сплошные линии – изобаты 100, 200 и 1000 м.
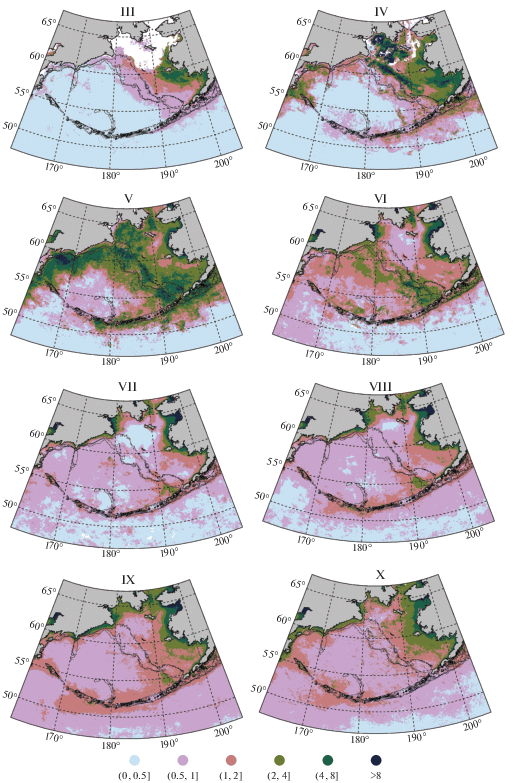
В апреле область высокой концентрации расширяется, охватывая практически весь восточный шельф (за исключением небольших участков, покрытых льдом) и западную часть моря, примыкающую к побережью Камчатки, Корякии и южной Чукотки. Наибольшие значения Хл (>8 мг/м3) наблюдаются преимущественно в северной и центральной частях восточного шельфа моря: приблизительно вдоль изобаты 100 м от м. Наварин почти до о-вов Прибылова, к западу от о. Св. Лаврентия и в Анадырском заливе. Относительно высока концентрация также в Бристольском заливе, в районе Олюторского и Карагинского заливов, в районе Камчатского пролива и к северу от Командорских о-вов. От марта к апрелю заметно повышается Хл также вокруг Алеутских островов, особенно в водах Аляскинского струйного течения. В мае на восточном шельфе моря концентрация обычно уже снижается, по крайней мере, над глубинами менее 100 м. “Цветение” постепенно смещается в более глубокую часть моря, в район бровки шельфа и материкового склона. Хотя над изобатами 100–200 м вдоль восточного шельфа по-прежнему сохраняются относительно высокие значения Хл (4–8 мг/м3), максимальные среднемноголетние значения (>10 мг/м3) наблюдаются, главным образом, вдоль материкового склона зал. Олюторского. Также заметно повышается Хл в центральной глубоководной части моря.
В июне “цветение” на восточном шельфе моря затухает, а в июле концентрация выше 1 мг/м3 наблюдается там только вокруг островов, в прибрежной зоне, в Анадырском заливе, в западной части бассейна Чирикова, а также над материковым склоном к северу от о. Уналашка и о. Умнак в районе каньона Беринга (Хл 2–4 мг/м3). При этом в июне “цветение” все больше продвигается в глубоководную часть моря. К северу от 57° с.ш. даже в глубоководной части моря среднее многолетнее значение концентрации составляет более 2 мг/м3. Однако на большой части акватории, удаленной от материкового склона и располагающейся примерно вдоль 55° с.ш., Хл даже в июне не превышает 1 мг/м3. К июлю на юге глубоководной части моря (над котловиной Бауэрс, в западной части Алеутской котловины, на юге Командорской котловины) и на северном шельфе моря (за исключением прибрежных районов, Анадырского залива и бассейна Чирикова) обычно формируются области с низкими значениями Хл (порядка 0.2–0.5 мг/м3). В августе такое распределение Хл по акватории почти полностью сохраняется. Отличается оно лишь тем, что области низких концентраций несколько размываются, а вдоль Алеутских островов концентрация возрастает. К югу от Алеутских островов от о. Агатту (восточный остров Ближних о-вов) до о. Атка, а также к северу от них от о. Амчитка до о. Сегуам концентрация в среднем составляет больше 1 мг/м3, а местами даже больше 2 мг/м3.
К сентябрю области повышенной концентрации вдоль Алеутских островов, побережья Аляски, Анадырского залива, от м. Наварин до м. Олюторского и далее вдоль всего камчатского побережья расширяются и объединяются в единую область. При этом в западной части Анадырского залива, а также в прибрежных районах западной части моря от м. Наварин до Карагинского залива средние многолетние значения Хл в узкой прибрежной полосе (порядка 50 км) местами превышают 4 мг/м3. К октябрю Хл вдоль Алеустких островов и п-ова Аляска убывает, среднемноголетнее значение концентрации там, за редким исключением, не превышает 2 мг/м3, а на большей части акватории находится в пределах 0.5–1.0 мг/м3. На севере Берингова моря, в западной части бассейна Чирикова, в Анадырском заливе и вдоль побережья от м. Наварин до м. Олюторского область повышенной концентрации сохраняется, но становится менее ярко выраженной. При этом области повышенной концентрации на всей протяженности внутренней части восточного шельфа, а также от зал. Олюторского на юг, включая акваторию вокруг Командорских островов, расширяются, и средние многолетние значения Хл в этих районах несколько повышаются. К ноябрю средние многолетние значения Хл убывают во всех районах (на рис. 2 не показано). Спутниковые данные по Хл за декабрь−февраль для большей части рассматриваемой акватории почти полностью отсутствуют из-за облачности или ледяного покрова.
Сроки пика “цветения” фитопланктона
Для характеристики сезонного смещения области пика “цветения” в данной работе использована группировка пикселей по среднему многолетнему месяцу максимума Хл (рис. 3).
Рис. 3.
Районы, в которых максимум среднего многолетнего сезонного хода концентрации хлорофилла-а приходится на месяцы с марта по октябрь. Месяцы обозначены римскими цифрами в верхней части каждой панели. Сплошная линия – изобата 200 м.
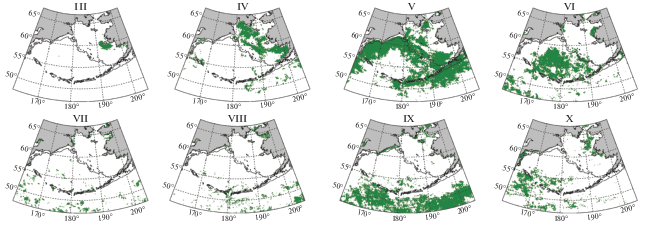
В марте сезонный максимум Хл отмечается во внутренней области восточного шельфа вокруг о. Нунивак за исключением зоны вдоль побережья материка. В апреле Хл достигает максимума на значительной части восточного шельфа моря: в Бристольском заливе, в центральной шельфовой области, в северной и восточной частях Анадырского залива, а также в области к югу и к западу от о. Св. Лаврентия. В мае сезонный пик Хл наблюдается в большей части Берингова моря, а также прилегающих вод Тихого океана. Это центральная область на юге восточного шельфа, район каньона Беринга, внешняя область шельфа практически на всем его протяжении, широкая полоса вдоль материкового склона моря от Алеутских о-вов на север до м. Наварин и далее на запад к Командорским о-вам, а также обширная область с океанской стороны п-ова Аляска и Алеутских островов.
В июне пик Хл достигается на обширной акватории глубоководной части моря на удалении более 500 км от свала глубин вдоль материков (но вдоль Командорско-Алеутской островной дуги – не везде). В июле–августе максимум Хл фиксируется лишь для относительно мелких участков исследуемой акватории, которые сложно объединить в какие-то обособленные районы. При этом сентябрь – месяц максимума Хл на обширной акватории открытой части Тихого океана к югу от 52° с.ш. В этом месяце максимум также достигается в относительно узкой прибрежной полосе шириной порядка 50 км на участках побережья от Анадырского пролива до залива Креста (восточная часть Анадырского залива) и от м. Наварин до восточной части Олюторского залива, а также вокруг о. Св. Лаврентия. В октябре максимум Хл наблюдается в восточной части бассейна Чирикова, к востоку от о. Св. Лаврентия, а также в западной части Олюторского залива и в Карагинском заливе.
Многолетняя динамика сроков “цветения”
Рассмотрим межгодовую динамику Хл на примере трех районов: в западной части моря к югу от Олюторского залива к северо-востоку от Камчатского пролива (западный район), на севере глубоководной части моря к югу от м. Наварин (северный район) и в центре моря (центральный район; см. рис. 1). Западный и северный районы близки по среднему многолетнему значению Хл (рис. 1), сезонному ходу Хл и сроку наступления максимума Хл (рис. 6), но многолетняя динамика Хл в них заметно различается. Хл в центральном районе во все месяцы заметно ниже, чем в двух других рассматриваемых районах, а максимум Хл приходится на более поздний срок.
В западном районе весеннее “цветение” фитопланктона обычно наблюдается в интервале с конца апреля по конец мая и длится 4–6 нед. (рис. 4). Однако в 2003, 2006 и 2010 гг. наблюдалось относительно позднее “цветение”, начавшееся в начале–середине мая и продлившееся до начала–середины июня. Обратная ситуация наблюдалась в 2015 и 2016 гг.: “цветение” началось в середине–конце марта. При этом оно было длительнее обычного и продолжалось до середины июня. В 2011 г. наблюдалось необычное для данного района “цветение”. Оно было слабо выражено, растянуто во времени с середины апреля по середину июля и имело несколько пиков. В целом с 2003 по 2019 г. наблюдается тенденция к смещению весеннего “цветения” на более ранние сроки. Осеннее “цветение” в западном районе отмечалось лишь в некоторые годы в октябре. В интервале лет 2003–2019 оно всегда было заметно менее интенсивным, чем весеннее “цветение”.
Рис. 4.
Динамика средней концентрации хлорофилла-а (мг/м3) в западном (а), северном (б) и центральном (в) районах в марте–октябре 2003–2019 гг. Для каждого района графики построены в одном масштабе, но для разных районов ис-пользован разный масштаб. Максимальные значения Хл указаны в верхней части каждого рисунка; даты максимумов – 9 мая 2009 г. в западном районе, 9 мая 2010 г. в северном и 17 мая 2019 г. в центральном районе. Пробелы на графиках отражают отсутствие данных.
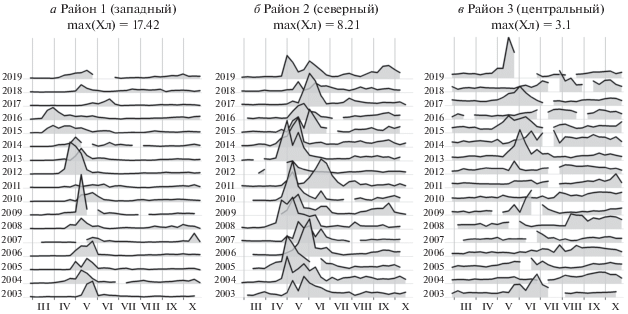
В северном районе “цветение” обычно начинается в конце апреля или в начале мая и длится 7–9 нед. То есть оно обычно происходит несколько позже, чем в западном районе, а также имеет бóльшую продолжительность. В 2005, 2008 и 2013 гг. в северном районе наблюдалось относительно раннее “цветение”, начавшееся в середине апреля. В 2011 и 2015 гг., напротив, цветение было относительно поздним. Пик цветения в эти годы пришелся на конец и начало июня соответственно.
Таким образом, в обоих рассматриваемых районах наблюдается значительная межгодовая изменчивость в сезонном ходе Хл. При этом разброс сроков пика “цветения” в 2003–2019 гг. составил около двух месяцев (рис. 4).
Средние по центральному району значения Хл существенно ниже, чем в рассмотренных ранее районах (до 2.7 мг/м3, а обычно даже во время “цветения” в интервале 1–1.5 мг/м3). При этом весеннее “цветение” фитопланктона там выражено гораздо слабее. Оно может начаться в мае (2003, 2009, 2013, 2017, 2019 гг.) или даже в конце апреля (2015 г.), а может быть сдвинуто к середине лета (2014, 2018 гг.). В этой части моря наблюдается осеннее “цветение”; в отдельные годы оно более ярко выражено, чем весеннее (например, 2004, 2008 и 2011 гг.). Там также возможен плавный переход от весеннего “цветения” к осеннему, и в таком случае период с относительно высокими значениями Хл может длиться несколько недель и даже месяцев (например, 2006, 2011, 2016 и 2018 гг.). То есть эта часть моря в зависимости от года может иметь черты сходства с другими районами, приуроченными к материковому склону, или с северной частью Тихого океана, где “цветение” обычно начинается в июле–августе и может длиться больше 50 дней (Friedland et al., 2018). Такие существенные межгодовые отличия в особенностях развития фитопланктона, предположительно, приводят к заметным изменениям в сообществе зоопланктона и более высоких трофических уровней.
Сравнение двух контрастных годов
Рис. 2–4 показывают, что на большей части рассматриваемой акватории весеннее “цветение” ежегодно достигает максимального развития в период с апреля по июнь. Рассмотрим развитие этого процесса на примере отличающихся друг от друга 2010 и 2015 гг. (рис. 5).
Рис. 5.
Сравнение распределения концентрации хлорофилла-а (мг/м3) в апреле (вверху), мае (в центре) и июне (внизу) 2010 и 2015 гг. (левый и правый столбцы соответственно). Белый цвет отмечает отсутствие данных (облачность или лед). Тонкие линии – изобаты 100, 200 и 1000 м.
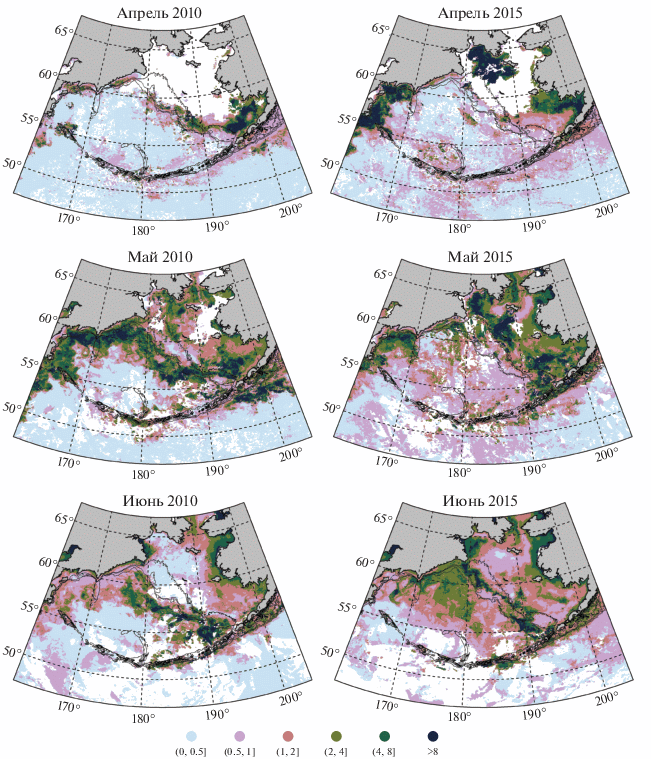
Рис. 6.
Месяцы среднего многолетнего максимума сезонного хода концентрации хлорофилла-а. Отмечены районы, по которым проанализирована межгодовая изменчивость (рис. 4). Месяцы обозначены римскими цифрами (III – март, … X – октябрь). Сплошные линии – изобаты 100, 200 и 1000 м.
В 2010 г. наиболее высокие значения Хл в апреле наблюдалась в Бристольском заливе и на прилегающей к нему центральной части юго-восточного шельфа моря, а также вдоль кромки льда, которая на участке шельфа от траверса каньона Прибылова до траверса каньона Первенец к концу апреля располагалась приблизительно вдоль изобаты 100 м (Baker et al., 2020). Там Хл местами превышала 4 мг/м3, а в южной части восточного шельфа даже 10 мг/м3. Практически вся остальная часть восточного шельфа до конца апреля еще полностью находилась подо льдом. Там “цветение” сформировалось только после освобождения акватории ото льда, что на большей части этой акватории произошло в мае. Вдоль западного шельфа моря в апреле преобладали значения в интервале 0.5–2 мг/м3.
В мае “цветение” было наиболее ярко выражено (Хл порядка 4–8 мг/м3) уже в глубокой части моря вдоль материкового свала глубин практически на всем его протяжении. На освободившейся ото льда части восточного шельфа преобладали значения концентрации в интервале 2–4 мг/м3. В этом месяце также сформировалось “цветение” фитопланктона к югу от п-ова Аляска (концентрация местами превышала 10 мг/м3).
К июню на большей части шельфа пик “цветения” уже был, судя по всему, пройден: Хл имела значения до 2 мг/м3. Однако в глубоководной части моря вдоль материкового свала глубин по-прежнему наблюдались высокие значения концентрации в интервале 2–8 мг/м3 и местами даже более 8 мг/м3. При этом полоса высоких значений в западной части моря (к западу от 180° в.д.) сместилась на юг и юго-восток от материкового склона.
В 2015 г. весеннее “цветение” фитопланктона развивалось по другому сценарию. Так же как и в 2010 г., в 2015 г. в апреле наблюдались высокие значения Хл в Бристольском заливе и на прилегающей акватории. Однако в отличие от 2010 г. большая часть северного шельфа моря начала освобождаться ото льда уже в апреле. В итоге к югу от о. Св. Лаврентия и в восточной части Анадырского залива уже в апреле наблюдались значения более 8 мг/м3. Другим ярким отличием от ситуации 2010 г. стало апрельское “цветение”, наблюдаемое в широкой полосе в западной части моря от Олюторского залива до Камчатского пролива.
К маю почти весь восточный шельф освободился ото льда. Так же как и в 2010 г., “цветение” на шельфе начало постепенно смещаться в сторону бровки шельфа. Однако в отличие от 2010 г. за пределы восточного шельфа область с концентрацией >2 мг/м3 распространилась только в районе каньона Беринга. Наиболее ярко “цветение” было выражено приблизительно вдоль изобаты 100 м. Хл там местами превышала 8 мг/м3. При этом в западной части моря (западнее 180° в.д.) наблюдались относительно высокие значения Хл (2–4 мг/м3, местами выше 4 мг/м3) даже на относительно большом удалении от бровки шельфа, а к югу от п-ова Аляска “цветение” было выражено гораздо слабее, чем в 2010 г.
В июне 2015 г. наиболее высокие значения Хл (выше 8 мг/м3) наблюдались на восточном шельфе в узкой полосе от о-вов Прибылова до м. Наварин а также в юго-восточной части моря за пределами шельфа в районе каньона Жемчуг. В отличие от 2010 г., концентрация вдоль материкового склона восточного шельфа была в основном <2 мг/м3, но при этом на обширной акватории глубоководной части моря в пределах 57°–62° с.ш. концентрация составляла >2 мг/м3. То есть “зеленый пояс” Берингова моря был выражен в основном в пределах шельфа, хоть и во внешней его части. При этом наблюдались необычно высокие значения концентрации в глубоководной части моря.
ОБСУЖДЕНИЕ
Пространственная структура
Сроки “цветения” фитопланктона относят к количественным экологическим индексам, характеризующим, в частности, особенности пелагических экосистем (Platt, Sathyendranath, 2008; Racault et al., 2012). Пространственная организация, упорядоченность морских экосистем в целом определяется широтной зональностью и может осложняться такими азональными факторами, как циркумконтинентальная секторность, влияние рельефа дна и циркуляция вод (Тамайчук, 2008; Петров, 2009). Очевидно, азональные факторы приобретают особенно заметное значение не только вблизи побережья океанов, материкового склона и в пределах окраинных морей, но также в районах с сезонным ледовым покровом. Например, ярко выраженная секторность наблюдается в фенологии “цветения” фитопланктона в пределах крупной полыньи в море Баффина: у восточного края полыньи “цветение” начинается в конце апреля или в начале мая, а у западного – в конце мая или даже позже (Marchese et al., 2017). Это явление связывают с различиями в физических условиях. По всей видимости, подобное различие в сроках физических явлений приводит к циркумконтинентальной и циркумшельфовой секторности распределения особенностей Хл и весеннего “цветения” фитопланктона в Беринговом море. Вероятно, этот фактор дополнительно осложняется наличием на восточном шельфе Берингова моря системы фронтальных зон (Flint et al., 2002).
Сезонная динамика
Наши результаты в целом согласуются с выводами А.М. Шпрингера с соавторами о повышенной продуктивности вдоль материкового склона Берингова моря (Springer et al., 1996). Однако они также указывают на метахронность этого явления: на различных участках т.н. “зеленого пояса” Берингова моря максимальное значение Хл наблюдается в различные периоды года, а в пределах одного участка “зеленого пояса” так же может наблюдаться в различные периоды в зависимости от года. Области высоких значений Хл могут существенно смещаться в пространстве год от года, а в отдельные годы этот “пояс” может трансформироваться в набор разрозненных фрагментов.
В сезонной динамике весеннего “цветения” фитопланктона в Беринговом море и на прилегающей акватории Тихого океана прослеживается следующая крупномасштабная структура (рис. 6). Весеннее “цветение” начинается в восточной части моря вокруг о. Нунивак в марте, распространяется из этого района в южном, юго-западном и западном направлениях как будто волной с апреля по май и затухает в центральной и южной глубоководной части моря.
В июле и августе активная вегетация наблюдается лишь в небольших районах, главным образом, вблизи побережий, а также в районе каньона Беринга. По всей видимости, в большинстве случаев это следствие таких процессов, как локальный ветровой апвеллинг или влияние материкового стока. Многие из этих районов, действительно, располагаются вблизи впадения рек. В частности, максимум Хл по данным спутникового зондирования в июне наблюдается в районах впадения в море таких крупных рек, как Анадырь и Юкон. Далее к июлю и августу эти области постепенно смещаются, по всей видимости, вдоль направления преобладающих в этих районах течений: на западе Анадырского залива на юг и, огибая м. Наварин, далее на запад, а в заливе Нортон на север, вокруг п-ова Сьюард. При этом материковый сток может содержать большое количество ОРОВ, имеющего схожие с хлорофиллом-а оптические свойства и влияющего на точность оценки Хл по спутниковым данным (Салюк и др., 2016; Matsuoka et al., 2007; Kopelevich et al., 2008). Поэтому значения Хл, оцененные по стандартному алгоритму OC3M, в районах, находящихся под влиянием материкового стока, могут быть существенно завышены. Для Берингова моря и близких акваторий в литературе встречаются оценки завышения в 1.3–1.5 раз (Салюк и др., 2016) или даже 2–5 раз (Lee et al., 2018; Naik et al., 2015).
Наличие высоких значений Хл вдоль побережья от м. Олюторского до м. Наварин и к северо-востоку от каньона Беринга подтверждается другими исследованиями. Вдоль побережья западной части моря в летние месяцы формируется локальный ветровой апвеллинг (Andreev et al., 2020). В районе каньона Беринга происходит подъем вод, связанный с совместным действием приливов и течений (Stabeno et al., 2002). Таким образом, высокие значения Хл в этих районах могут быть объяснены подъемом подповерхностных вод, богатых минеральными формами биогенных элементов, на поверхность.
Области открытой части моря, где максимум концентрации наблюдается в сентябре и октябре, следует отнести к областям с выраженным осенним “цветением” фитопланктона. По нашим данным в открытой части Тихого океана осеннее “цветение” обычно становится преобладающим. Это хорошо согласуется с результатами (Friedland et al., 2018). Однако в прибрежных районах Чукотки, Корякии и Камчатки, в Карагинском и Олюторском заливах, а также у побережья о. Св. Лаврентия ежегодно наблюдаются относительно высокие значения Хл (>2 мг/м3) в июне–августе, а в некоторые годы и в мае. То есть там ежегодно наблюдается ярко выраженное весеннее “цветение”.
В работах других авторов ранее уже было отмечено, что распространение прикромочного “цветения” фитопланктона вслед за отступающей кромкой льда во многих акваториях похоже на движение волны (Perrette et al., 2011). Наш анализ позволяет заключить, что подобным образом в Беринговом море распространяется “цветение” и на свободной ото льда акватории. То есть в Беринговом море, по всей видимости, наблюдаются две расходящихся волны “цветения”: волна прикромочного “цветения” движется в северном (северо-восточном) направлении вслед за отступающей кромкой льда, а волна классического “цветения” – в южном (юго-западном) направлении.
Межгодовая динамика
Очевидно, что особенности развития поля Хл в пределах любой акватории в отдельно взятый год будут зависеть от совокупности физических и биологических факторов (Кивва и др., 2020; Хен и др., 2015; Henson, 2007). Как упомянуто выше, большое значение имеют сезонная динамика теплового баланса поверхности моря, ветрового перемешивания и распространение льда. В связи с особенностями развития атмосферных процессов срок смены знака теплового баланса поверхности моря в одном и том же районе может существенно меняться год от года. В отдельные годы ведущую роль в скорости формирования устойчивой стратификации может играть ветровое перемешивание (Marchese et al., 2017).
Положение кромки льда в целом контролируется теплосодержанием океана (Bitz et al., 2005). То есть в случае положительной аномалии годового баланса тепла (с учетом адвективной составляющей) в пределах какой-либо отдельно взятой акватории при прочих равных кромка льда в среднем за холодный сезон будет располагаться севернее своего среднемноголетнего положения, а в обратном случае, соответственно, южнее. От зимнего распространения льда будет зависеть часть акватории, где потенциально сможет образоваться прикромочное “цветение” фитопланктона. При этом ширина прикромочной ледовой зоны также меняется: в Северном полушарии и, в частности, в Тихоокеанском секторе Северного Ледовитого океана на протяжении последних нескольких десятилетий она имеет тенденцию к увеличению (Strong, Rigor, 2013). По всей видимости, это может привести и к расширению области, на которой будет наблюдаться прикромочное цветение.
В 2010 г. в Беринговом море наблюдались аномальные ледовые условия. К примеру, 15 марта 2010 г. льдом был покрыт почти весь восточный шельф моря, включая Бристольский залив и исключая лишь относительно небольшой участок акватории на самом юге шельфа (Baker et al., 2020). 2015 г. значительно отличался: 15 марта 2015 г. кромка льда в пределах восточного шельфа располагалась приблизительно вдоль изобаты 100 м. Подобные различия в распространении и сезонной динамике ледового покрова в пределах моря не только напрямую определяют сроки формирования устойчивой структуры вод, но и отражают особенности теплового состояния вод моря, что значительно отражается на пространственно-временной изменчивости “цветения” фитопланктона (см. рис. 5).
Особенности Хл в центральной части Берингова моря
“Цветение” фитопланктона на восточном шельфе и вдоль материкового склона восточного шельфа относительно хорошо изучено (Sigler et al., 2014). Для западной части моря, примыкающей к материковому склону, натурных (in situ) исследований меньше (например, Мордасова, 1994). Но центральная часть моря остается наименее изученной в этом отношении; имеются лишь отрывочные данные о составе и распределении фитопланктона (Вентцель, 1991; Leblanc et al., 2005).
Срок начала классического “цветения” можно связать со сроком смены знака теплового баланса на границе море–атмосфера, то есть с моментом начала прогрева морской поверхности (Henson, 2007). Очевидно, что за счет географического положения в центральной глубоководной части Берингова моря прогрев морской поверхности начинается раньше (обычно с середины марта по середину апреля), чем в районе материкового склона у западной, северной и восточной границ глубоководной части моря (обычно с конца марта по конец апреля) (Гидрометеорология …, 1999). Однако согласно нашим результатам, “цветение” в глубоководной части моря наблюдается существенно позже – его пик в среднем приходится на июнь. Это может быть связано с несколькими факторами. Как отмечено выше, для формирования “цветения” необходимо наличие минеральных питательных веществ в эвфотическом слое, достаточная для фотосинтеза освещенность на всем вертикальном протяжении ВКС (Sverdrup, 1953) и превышение темпов деления клеток фитопланктона над темпами их потребления зоопланктоном (Behrenfeld, Boss, 2014).
Центральная часть моря может быть отнесена к регионам с высокой концентрацией основных биогенных элементов и низким уровнем хлорофилла-а (Banse, English, 1999; Shiomoto et al., 2002). Как и в северной части Тихого океана (а также во многих районах Южного океана), в этом районе моря развитие фитопланктона, скорее всего, ограничено концентрацией растворенного железа (Moore et al., 2013). В таком случае состав сообщества фитопланктона там может существенно отличаться от других районов моря (Banse, English, 1999). По всей видимости, вегетация диатомовых водорослей, формирующих основу весеннего “цветения” в других районах моря (Мордасова, 1994; Flint et al., 2002; Moran et al., 2012; Sukhanova, 1999), в пределах глубоководной части моря, удаленной от шельфовых районов, обычно затруднена недостатком железа. Такое предположение подтверждается распределением диатомей в верхнем слое донных осадков северной части Тихого океана (Ren et al., 2014), а также косвенно подтверждается результатами обобщения многолетних гидрохимических данных (Кивва, 2017). Показано, что в глубоководной части моря соотношение среднемноголетних сезонных величин убыли минеральных форм Si : N близко к единице. Это ближе к значениям для открытой части Тихого океана, чем для шельфовых районов моря, и указывает на меньшую долю диатомей в составе сообщества фитопланктона, чем в шельфовых районах.
Вопрос ограничения развития фитопланктона в центральной части моря лабильным растворенным железом требует обсуждения механизмов поступления этого элемента в центральную часть моря. По всей видимости, основными такими механизмами служат поступление атмосферного аэрозоля (Banse, English, 1999) и приток богатых железом вод из шельфовой части моря, интенсивность которого зависит, в частности, от формирования вихрей вдоль материкового склона (Aguilar-Islas et al., 2007; Tanaka et al., 2012). Вихревые структуры могут захватывать и переносить в глубоководную часть моря воды с высоким содержанием минеральных форм биогенных элементов. В них также может быть выше концентрация фитопланктона, продолжающего активно делиться при условии достаточного минерального питания. По всей видимости, с подобными процессами связана ситуация 2015 г., описанная выше, когда относительно высокие значения Хл наблюдались в глубоководной части моря. При этом похожие события наблюдались и в другие годы. В частности, значения Хл выше 2 мг/м3 отмечены на разных участках центральной части моря в июне–июле 2003, июне 2005, августе 2008, июне 2009, мае–июне 2013, июле 2014 и июне 2016 гг. Часть таких событий может быть связана не с адвекцией вод, а с выпадением атмосферного аэрозоля, в первую очередь вулканического происхождения.
Одной из причин позднего развития “цветения” в некоторых районах Мирового океана может быть преобладание темпа потребления фитопланктона зоопланктоном над темпом продуцирования ОВ на начальных стадиях “цветения” (Behrenfeld, Boss, 2014). Несмотря на то, что существующих данных о концентрации различных фракций зоопланктона и, тем более, о потреблении фитопланктона зоопланктоном в центральной части Берингова моря очень мало, существующие данные позволяют говорить о незначительной роли зоопланктона в контролировании биомассы фитопланктона (Banse, English, 1999).
Существенное ветровое перемешивание, несмотря на начало прогрева морской поверхности, может привести к более глубокому положению сезонного пикноклина в апреле и мае, чем глубина критической освещенности. Например, расположение нижней границы верхнего квазиоднородного слоя (ВКС) считается наиболее вероятной причиной позднего “цветения” в некоторых районах Южного океана (Ardyna et al., 2017). Среднемноголетняя мощность ВКС в центральной части Берингова моря в апреле составляет 80–120 м, а к маю уменьшается до 40–60 м (Лучин, 2019). Несмотря на то, что для Южного океана приводятся более значительные цифры, такое положение нижней границы ВКС в Беринговом море может быть одной из причин запаздывания “цветения” в центральной части моря.
Таким образом, относительно позднее развитие “цветения” в глубоководной части моря может быть связано с особенностями физических процессов формирования летней структуры вод и химического состава вод, определяющего состав сообщества фитопланктона.
ЗАКЛЮЧЕНИЕ
На основе анализа спутниковых данных по Хл мы показали, что в пределах Берингова моря “цветение” фитопланктона начинается на восточном шельфе в районе о. Нунивак и в Бристольском заливе. В 2003–2019 гг. это происходило в марте–апреле. Затем к апрелю–маю оно охватывало весь восточный и северный шельф и в мае–июне смещалось в район свала глубин (за исключением Командорско-Алеутской островной дуги). При этом сроки “цветения” год от года заметно сдвигаются во времени. Размах межгодовой изменчивости срока максимума Хл в западной части моря достигает, как минимум, 2 мес., а в центральной глубоководной части моря может составлять 5–6 мес.
Различия в ходе развития “цветения” проиллюстрированы на примере контрастных сезонов: 2010 и 2015 гг. В относительно холодном 2010 г., когда ледовитость моря была высокой, “цветение” развивалось по характерному для моря сценарию. Оно началось вдоль кромки льда в марте–апреле, распространилось вслед за отступающей кромкой льда на север и северо-восток, а также в район внешней части шельфа и материкового склона континентальной части моря к маю и в глубоководную часть моря вблизи материкового склона – к июню. В 2015 г. в апреле наблюдалось более интенсивное “цветение” в северной части шельфа, в заливах Олюторском и Карагинском, а также на прилегающей к ним части глубоководной акватории. В мае “цветение” вдоль материкового склона было менее интенсивным, чем в 2010 г. При этом в июне наблюдались высокие значения Хл в центральной глубоководной части моря, удаленной от материкового склона. Данные об изменчивости сроков очищения акватории Берингова моря ото льда (Baker et al., 2020) позволяют заключить, что такие различия в сезонном развитии и смещении “цветения” в северной части и на восточном шельфе моря были связаны с особенностями ледовой обстановки в марте–мае. Отличия в распределении Хл между 2010 и 2015 гг. в глубоководной части моря связаны с циркуляцией вод. Более детальный анализ влияния течений на поле Хл выходит за рамки данной статьи, но остается актуальной научной задачей.
Для глубоководной части моря, удаленной от материкового склона, во все месяцы характерны существенно менее высокие значения Хл, чем для шельфовых областей и части акватории, примыкающей к материковому склону. Обобщение данных о сезонном ходе минеральных форм биогенных элементов (Кивва, 2017), факторов, ограничивающих первичную продукцию в соседних районах Тихого океана (Moore et al., 2013), и доли диатомовых водорослей в донных отложениях северной части Тихого океана (Ren et al., 2014) позволяет заключить, что это связано с особенностями минерального питания фитопланктона. Низкая концентрация лабильного железа приводит к меньшей успешности в этом районе диатомовых водорослей по сравнению с шельфовыми и расположенными вблизи шельфа районами, где диатомеи формируют мощное весеннее “цветение”. При этом периодически происходят события переноса течениями воды в эту часть моря из района свала глубин материкового побережья моря или материкового склона Алеутских островов. Тогда там могут наблюдаться нехарактерно высокие значения Хл. По всей видимости, такое адвективное событие определило особенности пространственного распределения Хл на обширной акватории глубоководной части моря в 2015 г. Таким образом, в зависимости от особенностей циркуляции вод в разные годы центральная глубоководная часть моря имеет черты сходства с районами, расположенными вблизи материкового склона, или с открытой частью Тихого океана. В частности, там может наблюдаться заметное осеннее “цветение”, в отдельные годы (2004, 2008, 2011) имеющее бóльшую интенсивность, чем весеннее.
Особенности развития фитопланктона в течение года во многом определяют состояние других элементов экосистемы, в том числе промысловых популяций пелагических и донных видов рыб и беспозвоночных. От сроков весеннего “цветения” фитопланктона зависит доля первичной продукции, погружающаяся на дно, а также выживаемость молоди пелагических рыб. Бентосная часть экосистемы реагирует не только на количество поступающего ОВ, но и на его качественный состав (Grebmeier et al., 1988). Все это указывает на необходимость более детального изучения причин межгодовой изменчивости таких особенностей в основных промысловых районах Мирового океана.
Список литературы
Алексанин А.И., Ким В. Автоматическое обнаружение внутренних волн на спутниковых изображениях и оценка плотности перемешанного слоя // Исслед. Земли из космоса. 2015. № 1. С. 44–52.Андреев А.Г. Мезомасштабная циркуляция вод в районе Восточно-Сахалинского течения (Охотское море) // Исслед. Земли из космоса. 2017. № 2. С. 3–12.
Андреев А.Г. Циркуляция вод в северо-западной части Берингова моря по спутниковым данным // Исслед. Земли из космоса. 2019. № 4. С. 40–47.
Андреев А.Г., Жабин И.А. Мезомасштабные вихри и межгодовые изменения содержания хлорофилла-а в водах Охотского моря // Вестник ДВО РАН. 2012. № 6(166). С. 65–71.Бондур В.Г., Воробьев В.Е., Замшин В.В., Серебряный А.Н., Латушкин А.А., Ли М.Е., Мартынов О.В., Хурчак А.П., Гринченко Д.В. Мониторинг антропогенных воздействий на прибрежные акватории Черного моря по многоспектральным космическим изображениям // Исслед. Земли из космоса. 2017. № 6. С. 3–22.
Бондур В.Г., Замшин В.В., Чверткова О.И. Исследование из космоса экологического происшествия у полуострова Камчатка в сентябре–октябре 2020 года, связанного с красным приливом // Докл. АН. 2021. Т. 497. № 1. С. 61–68.
Бондур В.Г., Зубков Е.В. Выделение мелкомасштабных неоднородностей оптических характеристик верхнего слоя океана по многозональным спутниковым изображениям высокого разрешения. Часть 1. Эффекты сброса дренажных каналов в прибрежные акватории // Исслед. Земли из космоса. 2005. № 4. С. 54–61.
Вентцель М.В. Фитопланктон открытой и шельфовой областей Берингова моря // Океанология. 1991. Т. 31(2). С. 252–257.
Дружкова Е.И. Нанофитопланктон ледовой прикромочной зоны Баренцева моря в летний период 2017 г. // Труды КНЦ РАН. Океанология. 2018. Т. 4(9). Вып. 5. С. 29–44.
Кивва К.К. Гидрохимические условия первичного продуцирования в Беринговом море. Автореф. дис. канд. геогр. наук. М.: ВНИРО. 2017. 27 с.
Кивва К.К., Селиванова Ю.В., Писарева М.Н., Сумкина А.А. Роль физических процессов в формировании весеннего “цветения” фитопланктона в Беринговом море // Труды ВНИРО. 2020. Т 181. С. 206–222.
Копелевич О.В., Салинг И.В., Вазюля С.В., Глуховец Д.И., Шеберстов С.В., Буренков В.И., Каралли П.Г., Юшманова А.В. Биооптические характеристики морей, омывающих берега западной половины России, по данным спутниковых сканеров цвета 1998–2017 гг. М.: ООО “ВАШ ФОРМАТ”, 2018. 140 с.
Лучин В.А. Средние многолетние параметры верхнего квазиоднородного слоя Берингова моря (нижняя граница, температура, соленость) и их внутригодовая изменчивость // Изв. ТИНРО. 2019. Т. 199. С. 214–230.
Мордасова Н.В. Хлорофилл в западной части Берингова моря // Океанология. 1994. Т. 34. № 4. С. 557–563.
Петров К.М. Принципы физико-географической дифференциации арктических морей: Карское море // Из-в. РАН. Серия географическая. 2008. № 6. С. 19–30.
Салюк П.А., Степочкин И.Е., Букин О.А., Соколова Е.Б., Майор А.Ю., Шамбарова Ю.В., Горбушкин А.Р. Определение концентрации хлорофилла-а спутниковыми радиометрами MODIS Aqua и VIIRS в Восточной Арктике и Беринговом море // Исслед. Земли из космоса. 2016. № 1–2. С. 161–172.
Тамайчук АН. Основные закономерности региональной дифференциации Мирового океана // Уч. зап. Крымского фед. универ. им. В.И. Вернадского. География. Геология. 2008. Т. 21(60) № 2. С. 358–369.
Хен Г.В., Басюк Е.О., Матвеев В.И. Параметры верхнего квазиоднородного слоя и слоя скачка температуры и хлорофилл-а в западной глубоководной части Берингова моря летом и осенью 2002–2013 гг. // Известия ТИНРО. 2015. Т. 182. С. 115–131.
Aguilar-Islas A.M., Hurst M.P., Buck K.N., Sohst B., Smith G.J., Lohan M.C., Bruland K.W. Micro-and macronutrients in the southeastern Bering Sea: Insight into iron-replete and iron-depleted regimes // Prog. Oceanogr. 2007. V. 73(2). P. 99–126.
Andreev A.G., Budyansky M.V., Khen G.V., Uleysky M.Y. Water dynamics in the western Bering Sea and its impact on chlorophyll a concentration // Ocean Dyn. 2020. V. 70. P. 593–602.
Ardyna M., Claustre H., Sallée J.B., d’Ovidio F., Gentili B., Van Dijken G., d’Ortenzio F., Arrigo K.R. Delineating environmental control of phytoplankton biomass and phenology in the Southern Ocean // Geophys. Res. Lett. 2017. V. 44(10). P. 5016–5024.
Baker M.R., Kivva K.K., Pisareva M.N., Watson J.T., Selivanova J. Shifts in the physical environment in the Pacific Arctic and implications for ecological timing and conditions // Deep Sea Res., Part II, Top. Stud. Oceanogr. V. 177. 104802.
Banse K., English D.C. Comparing phytoplankton seasonality in the eastern and western subarctic Pacific and the western Bering Sea // Prog. Oceanogr. 1999. V. 43(2–4). P. 235–288.
Behrenfeld M.J., Boss E.S. Resurrecting the ecological underpinnings of ocean plankton blooms // Ann. Rev. Mar. Sci. 2014. V. 6. P. 167–94.
Bitz C.M., Holland M.M., Hunke E.C., Moritz R.E. Maintenance of the sea-ice edge // J. Clim. 2005. V. 18(15). P. 2903–2921.
Bliss A.C., Steele M., Peng G., Meier W.N., Dickinson S. Regional variability of Arctic sea ice seasonal change climate indicators from a passive microwave climate data record // Environ. Res. Lett. 2019. V. 14(4). P. 045003.
Coyle K.O., Eisner L.B., Mueter F.J., Pinchuk A.I., Janout M.A., Cieciel K.D., Farley E.V., Andrews A.G. Climate change in the southeastern Bering Sea: Impacts on pollock stocks and implications for the oscillating control hypothesis // Fish. Oceanogr. 2011. V. 20. P. 139–156.
Flint M.V., Sukhanova I.N., Kopylov A.I., Poyarkov S.G., Whitledge T.E. Plankton distribution associated with frontal zones in the vicinity of the Pribilof Islands // Deep Sea Res., Part II, Top. Stud. Oceanogr. 2002. V. 49(26). P. 6069–6093.
Friedland K.D., Mouw C.B., Asch R.G., Ferreira A.S.A., Henson S., Hyde K.J., Morse R.E., Thomas A.C., Brady D.C. Phenology and time series trends of the dominant seasonal phytoplankton bloom across global scales // Glob. Ecol. Biogeogr. 2018. V. 27(5). P. 551–569.
Grebmeier J.M., McRoy C.P., Feder H.M. Pelagic-benthic coupling on the shelf of the northern Bering and Chukchi seas. I. Food supply source and benthic biomass // Mar. Ecol. Prog. Ser. 1988. V. 48(1). P. 57–67.
Hunt Jr. G.L., Stabeno P., Walters G., Sinclair E., Brodeur R.D., Napp J.M., Bond N.A. Climate change and control of the southeastern Bering Sea pelagic ecosystem // Deep Sea Res., Part II. Top. Stud. Oceanogr. 2002. V. 49. P. 5821–5853.
Hunt Jr. G.L., Coyle K.O., Eisner L.B., Farley E.V., Heintz R.A., Mueter F., Napp J.M., Overland J.E., Ressler P.H., Salo S., Stabeno P.J. Climate impacts on eastern Bering Sea food webs: A synthesis of new data and an assessment of the Oscillating Control Hypothesis // ICES J. Mar. Sci. 2011. V. 68(6). P. 1230–1243.
Iida T., Saitoh S.I. Temporal and spatial variability of chlorophyll concentrations in the Bering Sea using empirical orthogonal function (EOF) analysis of remote sensing data // Deep Sea Res., Part II. Top. Stud. Oceanogr. 2007. V. 54. P. 2657–2671.
Kopelevich O.V., Burenkov V.I., Sheberstov S.V. Case studies of optical remote sensing in the Barents Sea, Black Sea and Caspian Sea // Remote sensing of the European seas / Eds. V. Barale, M. Gade. Dordrecht: Springer, 2008. P. 53–66.
Leblanc K., Hare C.E., Boyd P.W., Bruland K.W., Sohst B., Pickmere S., Lohan M.C., Buck K., Ellwood M., Hutchins D.A. Fe and Zn effects on the Si cycle and diatom community structure in two contrasting high and low-silicate HNLC areas // Deep Sea Res., Part I, Oceanogr. Res. Pap. 2005. V. 52(10). P. 1842–1864.
Lee S.H., Ryu J., Park J.W., Lee D., Kwon J.I., Zhao J., Son S. Improved Chlorophyll-a Algorithm for the Satellite Ocean Color Data in the Northern Bering Sea and Southern Chukchi Sea // Ocean Sci. J. 2018. V. 53(3). P. 475–485.
Marchese C., Albouy C., Tremblay J.É., Dumont D., D’Ortenzio F., Vissault S., Bélanger, S. Changes in phytoplankton bloom phenology over the North Water (NOW) polynya: a response to changing environmental conditions // Polar Biol. 2017. V. 40(9). P. 1721–1737.
Mikaelyan A.S., Kubryakov A.A., Silkin V.A., Pautova L.A., Chasovnikov V.K. Regional climate and patterns of phytoplankton annual succession in the open waters of the Black Sea // Deep Sea Res., Part I. Oceanogr. Res. Pap. 2018. V. 142. P. 44–57.
Mikaelyan A.S., Shapiro G.I., Chasovnikov V.K., Wobus F., Zanacchi M. Drivers of the autumn phytoplankton development in the open Black Sea // J. Mar. Syst. 2017. V. 174. P. 1–11.
Moore C.M., Mills M.M., Arrigo K.R., Berman-Frank I., Bopp L., Boyd P.W., Galbraith E.D., Geider R.J., Guieu C., Jaccard S.L., Jickells T.D. Processes and patterns of oceanic nutrient limitation // Nat. Geosci. 2013. V. 6(9). P. 701–710.
Moran S.B., Lomas M.W., Kelly R.P., Gradinger R., Iken K., Mathis J.T. Seasonal succession of net primary productivity, particulate organic carbon export, and autotrophic community composition in the eastern Bering Sea // Deep Sea Res., Part II, Top. Stud. Oceanogr. 2012. V. 65. P. 84–97.
Naik P., D’Sa E.J., Gomes H.D.R., Goés J.I., Mouw C.B. Light absorption properties of southeastern Bering Sea waters: Analysis, parameterization and implications for remote sensing // Remote Sens. Environ. 2013. V. 134. P. 120–134.
Naik P., Wang M., D’Sa E.J., Mordy C.W. Bering Sea optical and biological properties from MODIS // Remote Sens. Environ. 2015. V. 163. P. 240–252.
Okkonen S.R., Schmidt G.M., Cokelet E.D., Stabeno P.J. Satellite and hydrographic observations of the Bering Sea “Green Belt” // Deep Sea Res., Part II. Top. Stud. Oceanogr. 2004. V. 51. P. 1033–1051.
Perrette M., Yool A., Quartly G.D., Popova E.E. Near-ubiquity of ice-edge blooms in the Arctic // Biogeosciences. 2011. V. 8(2). P. 515–524.
Platt T., Fuentes-Yaco C., Frank K.T. Spring algal bloom and larval fish survival // Nature. 2003. 423(6938). P. 398–399.
Platt T., Sathyendranath S. Ecological indicators for the pelagic zone of the ocean from remote sensing // Remote Sens. Environ. 2008. V. 112(8). P. 3426–3436.
Racault M.F., Le Quéré C., Buitenhuis E., Sathyendranath S., Platt T. Phytoplankton phenology in the global ocean // Ecol. Indic. 2012. V. 14(1). P. 152–163.
Ren J., Gersonde R., Esper O., Sancetta C. Diatom distributions in northern North Pacific surface sediments and their relationship to modern environmental variables // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014. V. 402. P. 81–103.
Shiomoto A., Saitoh S.I., Imai K., Toratani M., Ishida Y., Sasaoka K. Interannual variation in phytoplankton biomass in the Bering Sea basin in the 1990s // Prog. Oceanogr. 2002. V. 55(1–2). P. 147–163.
Sigler M.F., Stabeno P.J., Eisner L.B., Napp J.M., Mueter F.J. Spring and fall phytoplankton blooms in a productive subarctic ecosystem, the eastern Bering Sea, during 1995–2011 // Deep Sea Res., Part II ,Top. Stud. Oceanogr. 2014. V. 109. P. 71–83.
Springer A.M., McRoy C.P., Flint M.V. The Bering Sea Green Belt: shelf-edge processes and ecosystem production // Fish. Oceanogr. 1996. V. 5(3–4). P. 205–223.
Stabeno P.J., Reed R.K., Napp J.M. Transport through Unimak Pass, Alaska // Deep Sea Res., Part II. Top. Stud. Oceanogr. 2002. V. 49. № 26. P. 5919–5930.
Strong C., Rigor I.G. Arctic marginal ice zone trending wider in summer and narrower in winter // Geophys. Res. Lett. 2013. V. 40(18). P. 4864–4868.
Sukhanova I.N. Spatial distribution and temporal variability of phytoplankton in the Bering Sea // Dynamics of the Bering Sea / Eds. T.R. Loughlin, K. Ohtani. 1999. P. 453–483.
Sverdrup H.U. On conditions for the vernal blooming of phytoplankton // J. Cons. Int. Explor. Mer. 1953. V. 18(3). P. 287–295.
Tanaka T., Yasuda I., Kuma K., Nishioka J. Vertical turbulent iron flux sustains the Green Belt along the shelf break in the southeastern Bering Sea // Geophys. Res. Lett. 2012. V. 39(8). L08603.
Wassmann P., Reigstad M. Future Arctic Ocean seasonal ice zones and implications for pelagic-benthic coupling // Oceanography. 2011. V. 24(3). P. 220–231.
Дополнительные материалы отсутствуют.
Инструменты
Исследование Земли из Космоса