

Исследование Земли из Космоса, 2021, № 5, стр. 85-93
Влияние суммарного значения температуры воды на размерно-массовые характеристики тихоокеанской устрицы Crassostrea gigas (Thunberg, 1793) в заливе Анива (о. Сахалин) по спутниковым данным
А. Ч. Ким a, *, Д. М. Ложкин a
a Сахалинский филиал ФГБНУ “ВНИРО” (“СахНИРО”)
Южно-Сахалинск, Россия
* E-mail: stasy.kim89@yandex.ru
Поступила в редакцию 07.04.2021
Аннотация
На основании массивов среднесуточных спутниковых данных по температуре поверхностного слоя воды за период 2010–2020 гг. в местах традиционного лова тихоокеанской устрицы Crassostrea gigas (бухта Лососей и лагуна Буссе) рассчитаны средние многолетние значения и аномалии длительности периода положительных температур и суммарные значения температуры воды. Выявлены зависимости размера раковины и общей массы Crassostrea gigas от температуры. В целом, в аномально теплые годы средние показатели высоты и массы значительно ниже нормы, а в аномально холодные годы – выше нормы. Это может свидетельствовать о том, что устрица, как вид умеренных широт, устойчива к холодным и умеренным температурам, но плохо развивается в условиях, повышенных в течение долгого периода температур. Исключение составляет масса моллюска в бухте Лососей, где наблюдается прямая зависимость от суммарного значения температуры воды.
ВВЕДЕНИЕ
В Сахалинской области одним из традиционных объектов промысла и любительского рыболовства является тихоокеанская (гигантская) устрица Crassostrea gigas (Thunberg, 1793). Ежегодное сверхлимитное ее освоение в рамках рекомендуемого вылова (РВ) привело к снижению запаса в лагуне Буссе и бухте Лососей (залив Анива, Охотское море). Огромный спрос к ресурсу, как к деликатесному двустворчатому моллюску, стал причиной сильного давления на его биологическое состояние. Как результат, в 2021 г. промысел устрицы в Восточно-Сахалинской подзоне закрыт. Однако, помимо антропогенного фактора, немаловажное влияние на жизненный цикл вида оказывают природные факторы окружающей среды, одним из которых является температура. Отсутствие работ, посвященных исследованию связи биологических параметров моллюска с изменениями термической обстановки в районах интенсивного лова на о. Сахалин, делает рукопись весьма актуальной. Поэтому целью данной работы является оценка влияния локальных температурных условий на высоту и вес раковины C. gigas в различных возрастных категориях.
Поскольку во многих районах не проводится круглогодичное наблюдение за факторами окружающей среды, на первое место выходят массивы спутниковых данных. Спектр применения спутниковых и прочих данных постоянно расширяется (Тихонов и др., 2017; Сизов, Кушнырь, 2018). Их практическому применению в области биологии посвящен целый ряд статей (Жабин и др., 2018; Ложкин и др., 2018; Пранц и др., 2020).
МАТЕРИАЛЫ И МЕТОДЫ
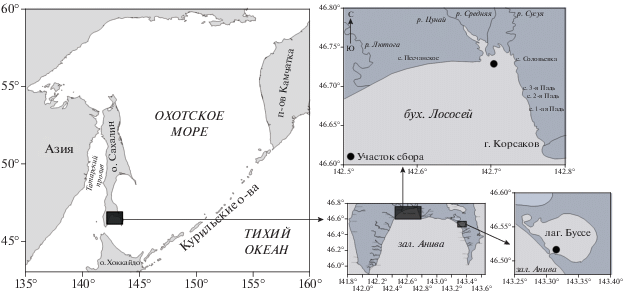
Работа основана на анализе размерно-массового и возрастного составов устрицы, собранной в 2010–2020 гг. в период учетных съемок и мониторинга в заливе Анива (включая лагуну Буссе). Районы исследований представлены на рис. 1. Координаты мест сбора материала: бухта Лососей (район с. Песчанское) – 46.728 518° с.ш., 142.703 118° в.д. и лагуна Буссе – 46.51 358 333° с.ш. 143.321 767° в.д.; 46.51 678 333° с.ш., 143.3178° в.д.; 46.51833333° с.ш., 143.318 183° в.д.; 46.52 195° с.ш., 143.311 283° в.д.; 46.52 366 667° с.ш., 143.311 917° в.д. Пробы отобраны и обработаны согласно общепринятым в гидробиологии методикам (Скарлато и др., 1964; Левин, Шендеров, 1975; Левин, 1994; Методы изучения …, 1990). Линейные измерения высоты раковины проводились при помощи штангенциркуля с точностью до 1 мм, общая масса животных определялась при помощи электронных весов A&D SK-2000WP (точность взвешивания 1 г). Построение диаграмм и необходимые расчеты осуществлялись посредством электронных таблиц Microsoft Excel.
Индивидуальный возраст устрицы определяется по радиальным срезам верхней створки (Золотарев, 1980; Tanabe, Oba, 1988; The age determination…, 1993; Kirby et al., 1998). В статье возраст установлен согласно указанным формулам: для бухты Лососей $t = \sqrt[{1.2251}]{{{M \mathord{\left/ {\vphantom {M {14.258}}} \right. \kern-0em} {14.258}}}}$, где t – возраст устрицы (лет) для бухты Лососей; M – общая масса тела (г) для каждой особи (Ким, Гон, 2020) и для лагуны Буссе $t = {{\left( {M - 0.3223} \right)} \mathord{\left/ {\vphantom {{\left( {M - 0.3223} \right)} {49.961~}}} \right. \kern-0em} {49.961~}},~$ где t – возраст устрицы (лет) для лагуны Буссе; M – общая масса тела (г) для каждой особи (Шпакова, Чумаков, 2013). Для возрастных групп от 1+ до 10+ лет по обоим районам (бухта Лососей и лагуна Буссе) были рассчитаны среднегодовые показатели высоты (H1, …, H10) и массы (M1, …, M10). Расчет производился только в том случае, когда количество экземпляров того или иного возраста, собранных в определенный год, составляло 6 и более. После чего были вычислены средние многолетние значения высоты и массы устрицы (далее приняты за нормы для отдельных возрастов) и их стандартные отклонения (σ) для каждого возраста. Объем обработанного материала составил 961 экз. в бухте Лососей и 2447 экз. в лагуне Буссе.
За исследуемый период плотность обитания моллюсков в заливе Анива варьировалась в широких пределах: в бухте Лососей – от 2 до 67 экз./м2, в лагуне Буссе – от 1 до 178 экз./м2.
Кроме того, использованы спутниковые данные по среднесуточной температуре поверхности океана (ТПО), полученные с помощью приемной станции TeraScan и хранящиеся в базе данных СахНИРО (более подробное описание приведено в работе (Новиненко, Шевченко, 2007)). Пространственное разрешение данных составляет около 2 км, однако для дальнейших расчетов было произведено пространственное осреднение (для района бухты Лососей и лагуны Буссе). Для заполнения пробелов, связанных с облачностью, был использован метод линейной интерполяции. Таким образом, был получен непрерывный ряд среднесуточных данных по ТПО, благодаря чему появилась возможность провести детальный статистический анализ.
Чтобы проследить зависимость размерно-массовых показателей от температуры, необходимо рассчитать соответствующие параметры. Для каждого года были условно выделены два периода (табл. 1): I период – от даты устойчивого перехода температуры через 0°C весной до 31 июля; II период – с 1 августа (массового нереста) до даты устойчивого перехода к отрицательным температурам. На основании этих данных вычислены длительности периодов (L1 и L2), суммарные значения температуры воды за указанные периоды (Q1 и Q2), среднепериодные температуры (t1w и t2w).
Таблица 1.
Даты перехода через 0°С (D1 – начало I периода; D2 – конец II периода)
| Год | D1 (Буссе) | D2 (Буссе) | D1 (Песчанское) | D2 (Песчанское) |
|---|---|---|---|---|
| 2010 | 07 апреля 2010 г. | 12 января 2011 г. | 16 апреля 2010 г. | 12 января 2011 г. |
| 2011 | 29 марта 2011 г. | 12 января 2012 г. | 05 апреля 2011 г. | 12 января 2012 г. |
| 2012 | 13 апреля 2012 г. | 27 декабря 2012 г. | 15 апреля 2012 г. | 20 декабря 2012 г. |
| 2013 | 18 апреля 2013 г. | 08 января 2014 г. | 15 апреля 2013 г. | 08 января 2014 г. |
| 2014 | 21 апреля 2014 г. | 22 декабря 2014 г. | 19 апреля 2014 г. | 25 декабря 2014 г. |
| 2015 | 27 марта 2015 г. | 14 декабря 2015 г. | 13 апреля 2015 г. | 23 декабря 2015 г. |
| 2016 | 09 апреля 2016 г. | 03 декабря 2016 г. | 21 апреля 2016 г. | 08 декабря 2016 г. |
| 2017 | 16 апреля 2017 г. | 06 декабря 2017 г. | 15 апреля 2017 г. | 19 декабря 2017 г. |
| 2018 | 16 апреля 2018 г. | 22 декабря 2018 г. | 10 апреля 2018 г. | 31 декабря 2018 г. |
| 2019 | 06 апреля 2019 г. | 21 декабря 2019 г. | 07 апреля 2019 г. | 18 декабря 2019 г. |
| 2020 | 11 апреля 2020 г. | 12 апреля 2020 г. |
ВАРИАЦИИ ТЕМПЕРАТУРЫ ПОВЕРХНОСТИ ВОДЫ
В табл. 2 представлены основные статистические параметры для ряда среднесуточных температур по годам, а в табл. 3 сведены средние многолетние (2010–2019 гг.) значения указанных параметров и их стандартные отклонения (σ). За исключением суммарного значения температуры воды за II период (Q2), температурные условия в лагуне Буссе подвержены большим межгодовым вариациям, чем в бухте Лососей. Что касается средних значений, их различия для двух районов невелики (находятся в пределах одного стандартного отклонения). В свою очередь, вариации всех параметров довольно значительны, разница между максимумом и минимумом превышает две величины стандартного отклонения.
Таблица 2.
Основные статистические параметры по годам
| Район лагуны Буссе | ||||||||
|---|---|---|---|---|---|---|---|---|
| год | L1, сут | Q1, °C | t1w, °C | L2, сут | Q2, °C | t2w, °C | Lсум, сут | Qсум, °C |
| 2010 | 116 | 1045.7 | 9.0 | 164 | 1595.7 | 9.7 | 280 | 2641.4 |
| 2011 | 125 | 924.6 | 7.4 | 164 | 1745.1 | 10.6 | 289 | 2669.6 |
| 2012 | 110 | 900.4 | 8.2 | 148 | 1693.4 | 11.4 | 258 | 2593.8 |
| 2013 | 105 | 946.2 | 9.0 | 161 | 1712.1 | 10.6 | 266 | 2658.3 |
| 2014 | 102 | 904.1 | 8.9 | 144 | 1637.3 | 11.4 | 246 | 2541.4 |
| 2015 | 127 | 1014.9 | 8.0 | 136 | 1571.3 | 11.6 | 263 | 2586.2 |
| 2016 | 114 | 1177.7 | 10.3 | 125 | 1519.8 | 12.2 | 239 | 2697.5 |
| 2017 | 107 | 1129.8 | 10.6 | 128 | 1556.3 | 12.2 | 235 | 2686.1 |
| 2018 | 107 | 1001.9 | 9.4 | 143 | 1683.9 | 11.8 | 250 | 2685.9 |
| 2019 | 117 | 1282.7 | 11.0 | 143 | 1603.1 | 11.2 | 260 | 2885.9 |
| 2020 | 112 | 1169.6 | 10.4 | |||||
| Район с. Песчанское | ||||||||
| год | L1, сут | Q1, °C | t1w, °C | L2, сут | Q2, °C | t2w, °C | Lсум, сут | Qсум, °C |
| 2010 | 107 | 1128.6 | 10.5 | 164 | 1717.2 | 10.5 | 271 | 2845.7 |
| 2011 | 118 | 918.0 | 7.8 | 164 | 1812.8 | 11.1 | 282 | 2730.9 |
| 2012 | 108 | 1020.8 | 9.5 | 142 | 1736.5 | 12.2 | 250 | 2757.3 |
| 2013 | 108 | 1057.3 | 9.8 | 161 | 1638.4 | 10.2 | 269 | 2695.7 |
| 2014 | 104 | 987.4 | 9.5 | 147 | 1674.1 | 11.4 | 251 | 2661.6 |
| 2015 | 110 | 1051.2 | 9.6 | 145 | 1561.8 | 10.8 | 255 | 2613.0 |
| 2016 | 102 | 1090.0 | 10.7 | 130 | 1569.4 | 12.1 | 232 | 2659.4 |
| 2017 | 108 | 1053.8 | 9.8 | 141 | 1529.6 | 10.8 | 249 | 2583.5 |
| 2018 | 113 | 1015.5 | 9.0 | 153 | 1710.8 | 11.2 | 266 | 2726.3 |
| 2019 | 116 | 1169.6 | 10.1 | 140 | 1636.9 | 11.7 | 256 | 2806.5 |
| 2020 | 111 | 1169.4 | 10.5 | |||||
Примечание. L1 – длительность I периода; Q1 – суммарное значение температуры воды за I период; t1w – среднепериодная температура за I период; L2 – длительность II периода; Q2 – суммарное значение температуры воды за II период; t2w – среднепериодная температура за II период; Lсум – длительность за I и II периоды; Qсум – суммарное значение температуры воды за I и II периоды.
Таблица 3.
Сводка статистических параметров за 2010–2019 гг.
| Район лагуны Буссе | Район с. Песчанское | |||||||
|---|---|---|---|---|---|---|---|---|
| минимум | максимум | среднее | σ | минимум | максимум | среднее | σ | |
| L1, сут. | 102 | 127 | 113 | 8 | 102 | 118 | 110 | 5 |
| Q1, °C | 900.4 | 1282.7 | 1045.2 | 128.3 | 918.0 | 1169.6 | 1060.1 | 76.6 |
| t1w, °C | 7.4 | 11.0 | 9.3 | 1.2 | 7.8 | 10.7 | 9.7 | 0.8 |
| L2, сут. | 125 | 164 | 146 | 14 | 130 | 164 | 149 | 11 |
| Q2, °C | 1519.8 | 1745.1 | 1631.8 | 74.4 | 1529.6 | 1812.8 | 1658.8 | 88.9 |
| t2w, °C | 9.7 | 12.2 | 11.3 | 0.8 | 10.2 | 12.2 | 11.2 | 0.7 |
Рассмотрим наиболее значимые отклонения от приведенной в табл. 3 нормы. В районе бухты Лососей раньше всего прогрев до положительных температур произошел в 2011 и 2019 гг., наиболее поздний прогрев наблюдался в 2014 и 2016 гг. Наибольшая длительность II периода в этом районе отмечена в 2010, 2011 и 2013 гг., в то время как в 2016 г. охлаждение произошло на 19 дней раньше обычного.
В районе лагуны Буссе, в отличие от бухты Лососей, 2019 г. по датам прогрева и охлаждения близок к норме. В 2017 г. длительность II периода была аномально низкой. В 2016 г. значение длительности первого периода практически совпадает с нормой, а II период на 3 недели короче обычного. 2015 г., напротив, в районе бухты Лососей можно характеризовать, как нормальный, в то время как в районе Буссе длительность обоих периодов далека от нормы.
Что касается суммарного значения температуры воды, видно, что чаще всего отрицательная аномалия в одном периоде компенсируется положительной аномалией в другом. Таким образом, суммарное значение температуры воды за два периода меняется менее значительно, чем в отдельные периоды. В обоих районах, как нормальные по этому признаку, можно характеризовать 2010 и 2018 гг., а аномально холодным является 2011 г. Аномально теплыми в обоих районах в I периоде оказались 2019 и 2020 гг. В 2012 и 2014 гг. в районе Буссе в I периоде наблюдалась значительная отрицательная аномалия Q1, а в 2016 г. – значительная положительная аномалия. В бухте Лососей параметр Q2 был аномально низким в 2015 и 2017 гг.
ЗАВИСИМОСТЬ РАЗМЕРА И МАССЫ ОСОБЕЙ ОТ НАКОПЛЕННОГО ТЕПЛА
В табл. 4 и 5 приведены минимальные, максимальные и средние многолетние значения, а также стандартные отклонения (σ) высоты и массы особей для каждой возрастной группы C. gigas (от 1+ до 10+) за весь период наблюдения.
Таблица 4.
Основные статистические параметры размера и массы особей по возрастам в районе с. Песчанское
| Район с. Песчанское | ||||||||
|---|---|---|---|---|---|---|---|---|
| возраст, лет | высота, мм | масса, г | ||||||
| минимум | максимум | среднее | σ | минимум | максимум | среднее | σ | |
| 1+ | 35 | 102 | 57 | 7 | 7 | 23 | 16 | 3 |
| 2+ | 48 | 109 | 70 | 4 | 24 | 44 | 35 | 3 |
| 3+ | 60 | 107 | 83 | 6 | 44 | 66 | 56 | 2 |
| 4+ | 70 | 127 | 92 | 7 | 67 | 90 | 79 | 2 |
| 5+ | 76 | 143 | 99 | 8 | 91 | 115 | 104 | 1 |
| 6+ | 83 | 169 | 108 | 9 | 116 | 141 | 127 | 2 |
| 7+ | 92 | 161 | 116 | 8 | 142 | 168 | 155 | 2 |
| 8+ | 100 | 194 | 127 | 14 | 169 | 195 | 181 | 3 |
| 9+ | 96 | 177 | 130 | 10 | 197 | 224 | 209 | 2 |
| 10+ | 110 | 209 | 142 | 14 | 225 | 254 | 242 | 1 |
Таблица 5.
Основные статистические параметры размера и массы особей по возрастам в районе лагуны Буссе
| Район лагуны Буссе | ||||||||
|---|---|---|---|---|---|---|---|---|
| возраст, лет | высота, мм | масса, г | ||||||
| минимум | максимум | среднее | σ | минимум | максимум | среднее | σ | |
| 1+ | 44 | 187 | 89 | 26 | 26 | 75 | 55 | 8 |
| 2+ | 52 | 197 | 100 | 15 | 76 | 125 | 100 | 6 |
| 3+ | 65 | 209 | 114 | 14 | 126 | 175 | 153 | 5 |
| 4+ | 101 | 197 | 126 | 9 | 176 | 225 | 200 | 4 |
| 5+ | 91 | 234 | 132 | 13 | 226 | 275 | 250 | 4 |
| 6+ | 105 | 224 | 148 | 8 | 276 | 325 | 302 | 2 |
| 7+ | 113 | 247 | 154 | 10 | 326 | 375 | 350 | 2 |
| 8+ | 115 | 228 | 162 | 9 | 376 | 425 | 403 | 3 |
| 9+ | 130 | 248 | 163 | 6 | 426 | 474 | 449 | 5 |
| 10+ | 123 | 250 | 173 | 5 | 475 | 524 | 498 | 4 |
Как высота, так и масса особей в каждой возрастной группе в пределах съемки в один и тот же год подвержена значительным вариациям. Разброс значений достигал 50 мм по высоте и 25 г по массе особей в районе бухты Лососей и около 100 мм по высоте и 50 г по массе в районе лагуны Буссе. Стоит заметить, что вариации среднегодовых показателей менее значительны, однако с их помощью можно установить, как влияет продолжительность теплого периода (L1 и L2) и суммарное значение температуры воды (Q1 и Q2) на размер и массу особей.
В табл. 6 показаны значимые отклонения от нормы, когда значение того или иного параметра (Li, Qi, Hi, Mi) было выше или ниже нормы на величину большую или равную σ для данного параметра. Плюсом обозначены положительные аномалии, минусом – отрицательные. Серым цветом показаны годы, в которые параметр находился в пределах нормы, а заштрихованные ячейки означают, что по указанной возрастной группе было отобрано 5 и менее экземпляров. Из этой таблицы можно увидеть, что в аномально теплые годы средние показатели высоты значительно ниже нормы, а в аномально холодные годы – выше нормы. Это может свидетельствовать о том, что устрица, как вид умеренных широт, устойчива к холодным и умеренным температурам, но плохо развивается в условиях, повышенных в течение долгого периода температур.
Таблица 6.
Отклонения среднегодовых показателей от нормы
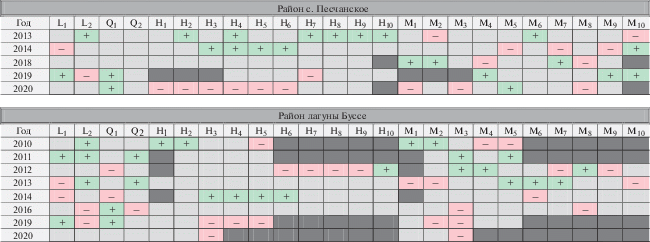
Примечание. L1 – длительность I периода; L2 – длительность II периода; Q1 – суммарное значение температуры воды за I период; Q2 – суммарное значение температуры воды за II период; H1, …, H10 – высота раковины в возрасте 1+ –10+ лет; М1, …, М10 – общая масса тела моллюска в возрасте 1+...10+ лет;  – положительная аномалия;
– положительная аномалия;  – отрицательная аномалия;
– отрицательная аномалия;  – параметр в пределах нормы;
– параметр в пределах нормы;  – количество экземпляров в возрастной группе ≤5.
– количество экземпляров в возрастной группе ≤5.
Рассмотрим в качестве примера графики аномалий высоты и массы в двух изучаемых районах: 4-леток для Бухты Лососей и 3-леток для лагуны Буссе (рис. 2). На рисунке столбцы означают аномалии соответствующих биологических параметров, а красной линией показана аномалия суммарного значения температуры воды за первый период (Q1). Синим пунктиром обозначено стандартное отклонение высоты или массы, а красным пунктиром – стандартное отклонение Q1. Возрастная группа подобрана таким образом, чтобы было достаточное количество экземпляров во все годы. К сожалению, 2015 г. для района бухты Лососей и 2018 г. для района лагуны Буссе пришлось исключить ввиду недостатка материала по большей части возрастных групп.
Рис. 2.
График аномалий основных исследуемых параметров в указанный год. 1 – аномалия высоты раковины (ΔН) или общей массы тела моллюска (ΔМ); 2 – аномалия суммарного значения температуры воды за I период (ΔQ1); 3 – стандартное отклонение ΔН или ΔМ; 4 – стандартное отклонение ΔQ1; 5 – малая выборка (9–10 экземпляров).
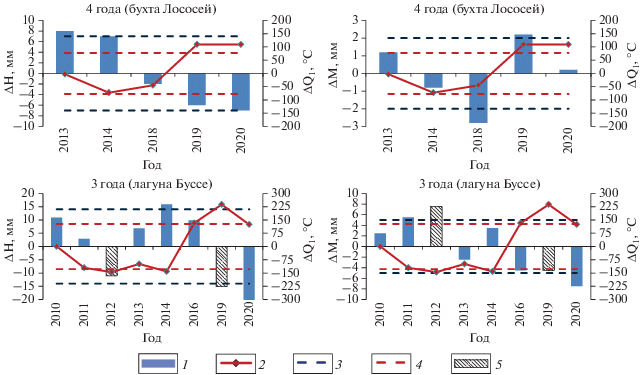
Норма высоты четырехлетних особей для бухты Лососей составляет 92 мм, в то время как норма массы – 79 г. Как видно из графика, два параметра (∆H и ∆Q1) практически находятся в противофазе, коэффициент корреляции составляет –0.77. Что касается массы, то в бухте Лососей наблюдается положительная корреляция (0.67).
В лагуне Буссе норма трехлетних особей по высоте составляет 114 мм, а по массе – 153 г. Сложно судить о корреляции высоты и массы особей и суммарного значения температуры воды, ведь даже для этой возрастной группы в 2012 и 2019 гг. было собрано лишь 9 и 10 экземпляров соответственно (поэтому на рис. 2 эти два года заштрихованы). Однако даже в этом случае наблюдается отрицательная корреляция и по высоте (–0.45), и по массе (–0.81).
Подтверждение сезонных и межгодовых изменений роста устрицы описано в работах (Раков, 1984; Холодов и др., 2017), где одним из основных факторов, ограничивающих активный рост, является температура воды. Последнее влияет на норму потребления кислорода и при 32°C наблюдается значительное увеличение смертности (Bougrier et al., 1995).
ЗАКЛЮЧЕНИЕ
Температурные условия в указанных районах подвержены значительным межгодовым вариациям, в самые суровые годы длительность теплого периода (с температурой воды выше 0°C) может отличаться от таковой в мягкие годы почти на два месяца. Интенсивность прогрева также зависит от текущих гидрометеорологических условий и может существенно отличаться из года в год. Естественно, подобные различия могут оказывать влияние на вид, чья активность зависит от температуры окружающей среды.
В целом, в аномально теплые годы средние показатели высоты и массы устрицы значительно ниже нормы, а в аномально холодные годы – выше нормы. Это может свидетельствовать о том, что устрица, как вид умеренных широт, устойчива к холодным и умеренным температурам, но плохо развивается в условиях, повышенных в течение долгого периода температур. Исключение составляет масса моллюска в бухте Лососей, где наблюдается прямая зависимость от суммарного значения температуры воды.
Полученные результаты можно считать лишь начальной оценкой, ведь температура является лишь одним из множества абиотических факторов среды, которые влияют на жизненный цикл объекта (таких как соленость, содержание кислорода, тип грунта, турбулентность воды, глубина обитания и т.п.), однако даже по столь грубой оценке видно, что она является немаловажным фактором окружающей среды.
Список литературы
Жабин И.А., Лукьянова Н.Б., Дубина В.А. Структура и динамика вод морской акватории национального парка “Шантарские острова” (Охотское море) по данным спутниковых наблюдений // Исслед. Земли из космоса. 2018. № 5. С. 3–14.
Золотарев В.Н. Продолжительность жизни двустворчатых моллюсков Японского и Охотского морей // Биология моря. 1980. № 6. С. 3–12.
Ким А.Ч., Гон Р.Т. Размерно-возрастная структура устрицы гигантской Crassostrea gigas в бухте Лососей (залив Анива): отчет о НИР (промежуточный). Южно-Сахалинск: СахНИРО, 2019. 38 с. Науч. арх. № 12693 н/а.
Левин В.С. Промысловая биология морских донных беспозвоночных и водорослей. С-Пб.: ПКФ “ОЮ-92”, 1994. 240 с.
Левин В.С., Шендеров Е.Л. Некоторые вопросы методики количественного учета макробентоса с применением водолазной техники // Биология моря. 1975. № 1. С. 64–70.
Ложкин Д.М., Цхай Ж.Р., Шевченко Г.В. Спутниковый мониторинг температурных условий в районах устьев нерестовых рек южной части о. Сахалин // Исслед. Земли из космоса. 2018. № 5. С. 15–22.
Методы изучения двустворчатых моллюсков: Труды Зоологического института АН СССР. Т. 219 / Под ред. Г.Л. Шкорбатова, Я.И. Старобогатова. Л.: ЗИН, 1990. 206 с.
Новиненко Е.Г., Шевченко Г.В. Пространственно-временная изменчивость температуры поверхности Охотского моря по спутниковым данным // Исслед. Земли из космоса. 2007. № 5. С. 50–60.
Пранц С.В., Кулик В.В., Будянский М.В., Улейский М.Ю. О связи мест промысла сайры с крупномасштабными когерентными структурами в океане по спутниковым данным // Исслед. Земли из космоса. 2020. № 4. С. 18–26.
Раков В.А. Биологические основы культивирования тихоокеанской устрицы Crassostrea gigas Thunberg в заливе Петра Великоrо. Автореферат дисс. на соискание ученой степени к. б. н. – Владивосток, 1984. 24 с.
Сизов О.С., Кушнырь О.В. Анализ подходов к систематизации тематических задач, решаемых с помощью данных // Исслед. Земли из космоса. 2018. № 2. С. 77–86.
Скарлато О.А., Голиков А.Н., Грузов Е.Н. Водолазный метод гидробиологических исследований // Океанология. М.: Наука, 1964. № 4. С. 710–711.
Тихонов В.В., Хвостов И.В., Романов А.Н., Шарков Е.А. Анализ изменений ледяного покрова пресноводных водоемов по данным SMOS // Исслед. Земли из космоса. 2017. № 6. С. 46–53.
Холодов В.И., Пиркова А.В., Ладыгина Л.В. Выращивание мидий и устриц в Черном море. Воронеж: ООО “Издат-принт”, 2017. 508 с.
Шпакова Т.А., Чумаков Д.Е. Размерно-возрастная структура промысловых скоплений устрицы в лагуне Буссе: отчет о НИР (промежуточный). Южно-Сахалинск: СахНИРО, 2013. 24 с. Науч. арх. № 11841 н/а.
Bougrier S., Geairon P., Deslous-Paoli J., Bacher C., Jonquières G. Allometric relationships and effects of temperature on clearance and oxygen consumption rates of Crassostrea gigas (Thunberg) // Aquaculture. 1995. V. 134(1). P. 143–154. https://doi.org/10.1016/0044-8486(95)00036-2
Kirby M.X., Soniat T.M., Spero H.J. Stable Isotope Sclerochronology of Pleistocene and Recent Oyster Shells (Crassostrea virginica) // Palaios. 1998. V. 13(6). P. 560–569.
Tanabe K., Oba T. Latitudinal variation in shell growth patterns of Phacosoma japonicum (Bivalvia: Veneridae) from the Japanese coast // Marine Ecology Progress Series. 1988. V. 47. № 1. P. 75–82.
The age determination and growth rate of the European flat oyster, Ostrea edulis, in British waters determined from acetate peels of umbo growth lines / C.A. Richardson, S.A. Collis, K. Ekaratne et al. (eds) // ICES J. of Mar. Sci. 1993. V. 50(4). P. 493–500.
Дополнительные материалы отсутствуют.
Инструменты
Исследование Земли из Космоса