

Исследование Земли из Космоса, 2022, № 1, стр. 38-52
Пространственно-временная изменчивость цветений цианобактерий на основе их автоматической идентификации по данным MODIS
П. Н. Лишаев a, А. А. Алескерова a, *, А. А. Кубряков a, Н. В. Василенко a, С. В. Станичный a
a Морской гидрофизический институт РАН
Севастополь, Россия
* E-mail: annete08@mail.ru
Поступила в редакцию 09.04.2021
- EDN: BASIFE
- DOI: 10.31857/S020596142201002X
Аннотация
В работе предложен автоматической алгоритм выделения областей цветения цианобактерий (ЦБ) в Азовском море на основе анализа оптических спектров спутниковых данных MODIS. Валидация алгоритма проведена на основе сопоставления с квазисинхронными данными высокого разрешения Landsat. На основе результатов применения алгоритма были исследованы пространственная и временная изменчивость цветений ЦБ в Азовском море в 2003–2019, даны примеры эволюции цветений в отдельные годы. Цветение ЦБ в Азовском море наблюдается с марта по ноябрь с максимумом в августе. Наиболее интенсивные и продолжительные цветения ЦБ наблюдаются в Таганрогском заливе. Отсюда, в весенние месяцы они переносится на восток вдоль северного берега. В июне–июле ЦБ начинают интенсивно распространяться на юг вдоль восточного берега, после чего в некоторые годы проникают в центр бассейна под действием антициклонических течений. Анализ межгодовой изменчивости показал, что интенсивность цветений была наиболее высокой в 2004–2011, с максимумом в 2008–2009 гг., и значительно упала в последние годы (2011–2019). Возможной причиной такого уменьшения является снижение стока рек Дона и Кубани, а также рост скорости ветра, отмечаемый в последние годы.
ВВЕДЕНИЕ
Азовское море является мелководным водоемом внутреннего типа, для которого характерна относительно низкая соленость вод, интенсивный прогрев, активное ветровое перемешивание, а также выраженное влияние пресноводного стока рек Дон и Кубань, несущего биогенные элементы (Ильин и др., 2009; Сорокина, Бердников, 2018). Такие условия способствуют ежегодным сезонным цветениям фитопланктона в водах акватории, в том числе цветениям цианобактерий (ЦБ) (Ковалева, 2020).
Наличие ЦБ, а также отдельные случаи их цветений в Азовском море, были описаны в ряде работ (Ковалева, 2006; Матишов и др., 2009; Селифонова, 2008; Selifonova, 2010). По данным контактных измерений было установлено, что наиболее интенсивные цветения ЦБ наблюдаются в зонах конвергенции вод. Так, в Таганрогском заливе в период гидрологического лета ЦБ представляют до 90% всей биомассы фитопланктона (Ковалева, 2020; Никитина, 2010), в Темрюкском заливе до 80% всей биомассы (Ясакова, 2015). В остальных районах акватории ЦБ также фиксируются, однако в значительно меньшем количестве.
Цветения ЦБ могут оказывать негативное воздействие на состояние экосистемы региона: в период таких цветений продукция кислорода значительно меньше его поглощения, что приводит к гипоксии в толще воды (Матишов и др., 2003). ЦБ образуют толстые поверхностные пленки, которые снижают доступность света и растворимость кислорода, что препятствует росту биоты в более глубоких слоях (Paerl & Otten, 2013). Также отдельные виды ЦБ способны продуцировать нейро- и гепотоксины в окружающие воды, что представляет дополнительную опасность для живых организмов. Они могут вызвать гибель зоопланктона и рыб (Андреева и др., 2020, Sivonen, Jones, 1999; Harke et al., 2016). Таким образом, интенсивные цветения ЦБ могут приводить к массовым заморным явлениям (Матишов, Футшей. 2003).
Контактные измерения позволили обнаружить некоторые регионы нахождения ЦБ и получить информацию о составе, морфометрии и других характеристиках этих видов. Однако такие измерения нерегулярны по времени и пространству, что критично для изучения пространственного распространения цветения ЦБ.
Возможность регулярной идентификации областей цветения ЦБ могут дать мультиспектральные спутниковые измерения. ЦБ оказывают значительное влияние на отражательную способность морской поверхности и имеют выраженные спектральные особенности, которые являются основой методов выделения областей их цветений по данным спутниковых оптических сенсоров. В настоящее время существует несколько таких методов для выделения ЦБ в различных акваториях Мирового океана (Vincent et al., 2004; Kutser et al., 2006; Kahru et al., 2007; Blondeau-Patissier et al., 2014; Карабашев и др., 2015; и др). Исследование изменчивости цветений ЦБ в Азовском море по спутниковым данным проводилось в работе (Aleskerova et al., 2019). В этой работе на основе оптических данных высокого разрешения Landsat был представлен обобщающий анализ особенностей пространственной изменчивости характеристик цветений за 1999–2016 гг., описаны экстремальные цветения в различные годы исследуемого периода. Данные Landsat дают возможность наблюдать за особенностями цветения нитчатых плавающих водорослей. Однако они имеют ряд ограничений, связанные прежде всего, с малым временным разрешением (16 дней) этих измерений, которое с учетом облачности часто не дает возможность наблюдать за эволюцией цветений ЦБ.
В настоящей работе на основе сопоставления данных среднего разрешения MODIS и высокого разрешения Landsat проводиться анализ особенностей оптических спектров в области цветения ЦБ. На основе этого анализа предлагается алгоритм автоматической идентификации цветений ЦБ в Азовском море, основанный на ежесуточных данных 10 оптических каналов сканера MODIS. Разработанный алгоритм используется для исследования пространственной, сезонной и межгодовой изменчивость цветений ЦБ за период 2003–2019 гг., особенностей их эволюции и распространения в отдельные годы.
ДАННЫЕ
В работе использовался массив данных сенсоров спутников серии Landsat 4, 5, 7, 8 с разрешением 15–30 м (для построения RGB изображений true color), которые были получены из архива https://glovis.usgs.gov/. Для анализа спектров области цветений использовались данные об отражательной способности RRS (Remote Sensing Reflectance) MODIS в 10 каналах, центрированных на 412, 443, 469, 488, 531, 547, 555, 645, 667, 678 нм, которые имеют разрешение 1 км и получены из архива http://oceandata.sci.gsfc.nasa.gov/. Для анализа скорости ветра использовались данные реанализа ERA-Interim (пространственное разрешение – 0.25 градуса, дискретность по времени – 6 ч). Данные о стоках рек Дон и Кубань были взяты из массива GloFAS-ERA5 (Harrigan et al., 2020).
МЕТОД ИДЕНТИФИКАЦИИ ЦИАНОБАКТЕРИЙ
Цветение ЦБ определялось визуально по снимкам Landsat на основе следующих критериев. ЦБ имеют значительно повышенные значения яркости в ближнем инфракрасном диапазоне (NIR) по сравнению с чистой водой (Blondeau-Patissier et al., 2014) за счет большой биомассы и положительной плавучести. Наличие хлорофилла “а” в клетках ЦБ приводит к повышенному поглощению света в синей части спектра по сравнению с мутными водами, содержащими взвесь минерального происхождения. При построении композитных изображений RGB области цветения ЦБ будут обладать насыщенным зеленым цветом. Дополнительным критерием являлось проявление пространственных структур в NIR канале (см. пример на рис. 1, б). На рис. 1, б красным выделяются плавающие нитевидные структуры ЦБ, которые аккумулируются в конвергентных зонах.

Для разработки автоматического алгоритма идентификации ЦБ по виду спектра яркости, был выполнен совместный анализ данных Landsat и MODIS. На снимках Landsat в видимом диапазоне были отобраны сцены, на которых явно отмечались участки, подверженные цветению ЦБ. Далее, используя квазисинхронные данные MODIS за те же даты, проводился анализ спектров в точках, соответствующей цветению, и вне его. Рассмотрим спектры, полученные по данным MODIS на примере 16 июля 2013 г. На снимке Landsat четко наблюдается обширная область цветения ЦБ в районе юго-восточной части Азовского моря (рис. 2, а). Эта область имеет форму вытянутой с севера на юг изгибающейся полосы шириной не более 10 км и длиной около 70 км. Повышенные значения яркости для этой области характерны и для карты яркости MODIS (рис. 2, б).
Рис. 2.
а – RGB снимок Landsat за 16 июля 2013 г.; б – RGB снимок MODIS за 16 июля 2013 г.; в – пространственное распределение значений яркости в 10-канале MODIS; г – спектры выбранных точек.
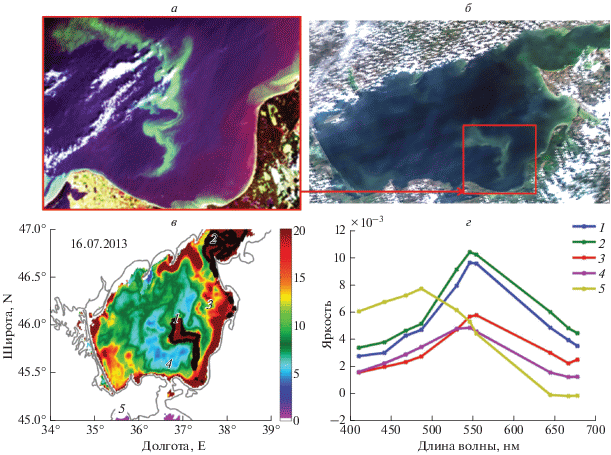
Рассмотрим спектры для отмеченных точек: 1, 2– цветений ЦБ; 3, 4 – повышенные значения яркости вне зоны цветения ЦБ (цветение фитопланктона или взвесь); 5 – относительно “чистые” воды. Для спектра точки 5 в Черном море (условно “чистая” вода) максимум наблюдается в 3–4 каналах на длинах волн 469–488 нм, а яркость в длинноволновой части спектра стремится к 0. Спектры точек 1–4 характеризуются максимумом в 6–7 каналах (547–555 нм), свидетельствующим о высоких значениях обратного рассеяния взвеси. Еще одной особенностью спектра области цветения цианобактерий точек 1, 2 является “ступенька”, сформированная между 3 и 4 каналами (469 и 488 нм, соответственно). Возможной причиной образования данной “ступеньки” можно назвать наличие каротиноидов, которые играют фотозащитную функцию для ЦБ (Paerl et al., 1983, 1985), давая преимущество для роста в поверхностном слое вод с очень высокими значениями светового потока. Максимум поглощения каротиноидов приходится на интервал 460–510 нм (Holt, Krogmann, 1981). Таким образом, в области цветения ЦБ наблюдается сильное поглощение в 4 канале (488 нм). Эта особенность отсутствует для спектров 3–4, что говорит о недостаточным количеством поглощающих пигментов, характерных для ЦБ. Повышенные концентрации взвешенного вещества в этих точках, по-видимому, связаны с взмучиванием придонных осадков в мелководном Азовском море.
Данным методом было проанализировано более чем 20 квазисинхронных сцен Landsat/MODIS. Анализ “характерных” спектров ЦБ позволил разработать набор флагов, представленных в табл. 1, для алгоритма автоматической идентификации ЦБ по данным сканера MODIS. Флаги были подобраны таким образом, чтобы избежать детектирования ложных цветений. Каждый из флагов тщательно подбирался и проверялся на всем массиве данных MODIS за 2003–2019 гг.
Таблица 1.
Набор флагов для алгоритма автоматической идентификации ЦБ по данным сканера MODIS
| Номер | Условия для спектра, необходимые при идентификации ЦБ |
|---|---|
| 1 | RRS(678) > 0.001 |
| 2 | RRS(443)./RRS(547) < 0.8 |
| 3 | (RRS(488)–RRS(469))./(RRS(531)–RRS(488)) < 0.4*(488–469)/(531–488) |
| 4 | (RRS(488)–RRS(469))./(RRS(469)–RRS(443) < (488–469)/(469–443) |
| 5 | RRS(645)./RRS(547) < 0.7 |
| 6 | RRS(488)./RRS(547) < 0.8 |
| 7 | RRS(469) > 0.85*RRS(488) |
Наличие ЦБ в воде приводит к изменению спектра отражательной способности, особенно в диапазоне длин волн 1–6 каналов (412–550 нм), соответствующих сине-зеленому спектру, что может быть вызвано наличием в составе ЦБ определенных пигментов.
Рассмотрим предложенную систему флагов.
1. Флаг 1 используется для идентификации вод с большим содержанием взвешенного вещества или ЦБ, поглощающего в ближнем ИК спектре.
2. Флаги 2, 6 позволяют “отсечь” воды, в которых недостаточно влияние хлорофилла (“чистые” воды или воды с повышенным содержанием взвешенного вещества).
3. Флаги 3, 4 используются для идентификации “ступеньки”, формирующейся, скорее всего за счет поглощения каротиноидов в канале 3 (469 нм) и 4 (488 нм). Также необходимо отслеживать угол, образованный линиями, соединяющими значения яркости в каналах 2, 3, 4 и 5.
4. Флаг 7 отвечает за “горизонтальное” положение “ступеньки”, сформированной значениями отражения между 3 и 4 каналами.
5. Флаг 5 позволяет отметить мелководные области, а также участки суши, в которых коэффициент отражения в ближнем ИК-канале выше, чем в более коротковолновой области спектра.
Примеры значений флагов для спектров, изображенных на рис. 2, приведены в табл. 2. Спектры условно мутных вод точек 3 и 4 не удовлетворяют флагам 3, 4 и 7, а спектры относительно чистых Черноморских вод точки 5 – условиям 1, 2, 4 и 6.
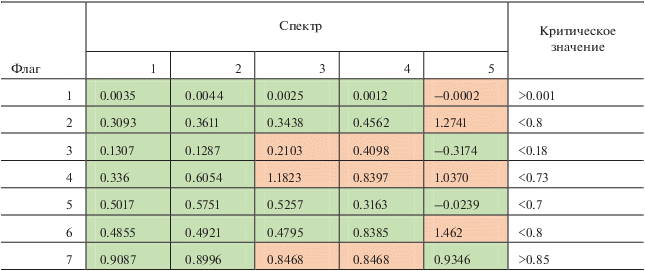
В табл. 2 зеленым цветом отмечены критерии, которые выполняются, а красным – которые не выполняются. Номера столбцов соответствуют номерам спектров, которые представлены на рис. 2, в. Строки – значения, полученные для каждого спектра и сравниваемые с приведенными критериями в табл. 1
На рис. 3 представлены еще два примера сопоставления RGB-снимков со спутника Landsat с картами распределения ЦБ, полученных по данным MODIS. На спутниковом снимке Landsat за 16 августа 2003 г. наблюдается область цветения ЦБ в центральной части Азовского моря, которая также выделяется и на картах MODIS, построенных по предложенному нами алгоритму (см. черные контурные линии на рис. 3, б). Максимальное по интенсивности цветение наблюдается в Таганрогском заливе. Отсюда воды подверженные цветению распространяются от залива в южном направлении до 45.6° с.ш. При этом вдоль восточного берега наблюдается более чистые воды, за исключением районов устьев рек Кубань, Протока и Ахтарского лимана.
Рис. 3.
а – RGB снимок Landsat-7 за 16 августа 2003 г.; б – контуры выделенных областей цветения ЦБ, которые наложены на карту MODIS хлорофилла А за 16 августа 2003 г.; в – RGB снимок Landsat-7 за 2 сентября 2015 г.; г – контуры выделенных областей цветения ЦБ, которые наложены на карту MODIS хлорофилла А за 2 сентября 2015 г.
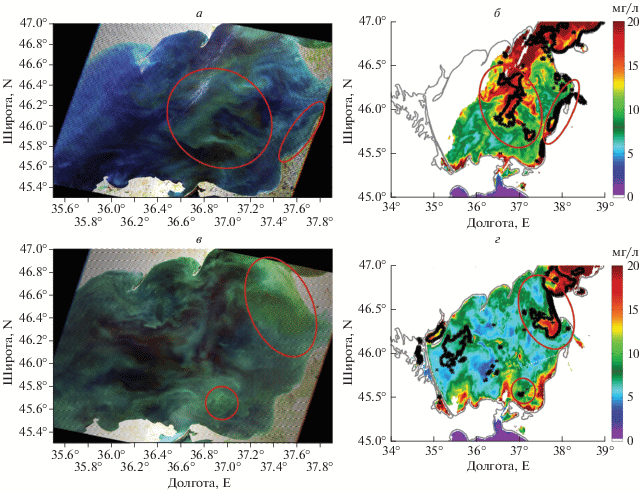
24 августа 2009 года цветение наблюдается в восточной части Азовского моря (рис. 3, в, 3, г). Максимальная концентрация ЦБ наблюдалась у стока реки Кубань, где водоросли образовывали нитчатую структуру на фронте речных вод. Подобные нитевидные структуры наблюдались так же у Обиточной и Бердянской кос, и по центру Азовского моря. При сопоставлении RGB-снимка и карты распределения ЦБ, полученной по алгоритму, наблюдается достаточно хорошее соответствие. Алгоритм показывает отсутствие водорослей напротив Ясенского залива и отфильтровывает взмученные воды Арабатского залива.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Пространственное распределение и сезонная изменчивость
На основе разработанного алгоритма были построены карты вероятности наблюдения ЦБ: выделенная область цветения отмечалась значением 1, все остальные пиксели – 0. Затем осреднение за выбранный промежуток времени определяло вероятность наблюдения ЦБ (P) в данной точке.
Карта СКО (среднеквадратичного отклонения) вероятности наблюдения ЦБ в Азовском море, полученная по предложенному алгоритму, показывают, что наиболее часто ЦБ отмечается в Таганрогском заливе, поскольку гидрологические условия в этом районе наиболее благоприятны для их развития. Это подтверждается и данными биологических наблюдений (Матишов, Фуштей, 2003; Макаревич, Ларионов, 2006; Никитина, 2010). Однако, под действием течений, области цветения могут распространяться далеко вдоль восточного или западного берегов моря, откуда затем могут проникать в центральную часть моря (рис. 4, а). При этом наиболее встречаемое направление распространения южное – вдоль юго-восточного берега.
Рис. 4.
а – Карта СКО вероятности наблюдения ЦБ в Азовском море, полученная по данным сканера MODIS; б – сезонный ход вероятности обнаружения ЦБ за 2003–2019 гг.
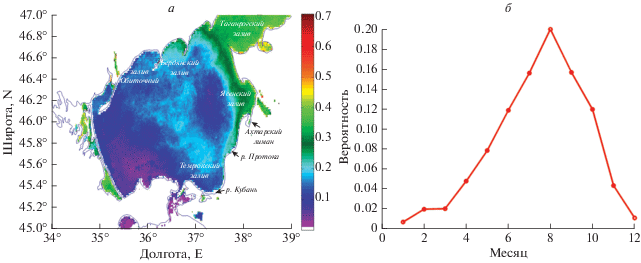
Сезонный ход вероятности наблюдения ЦБ (рис. 4, б) в Азовском море характеризуется наибольшими значениями с июня по октябрь, с выраженным пиком в августе, что согласуется с результатами анализа спутниковых данных высокого разрешения (Алескерова и др., 2019). Наименьшие значения наблюдаются в зимние месяцы, с декабря по март. Среднемесячные климатические карты P (рис. 5) показывают, что цветение обычно начинается в марте в Таганрогском заливе, откуда под действием циклонической циркуляции ЦБ переносятся на запад. В результате, в весенние месяцы чаще цветению подвержены воды северного побережья. В июне–июле ЦБ от выхода из Таганрогского залива начинают интенсивно распространяться на юг в центральную часть моря. В это время вдоль восточного побережья значения P достигают P = 0.5, т.е. ЦБ фиксируются в половине случаев. Интенсивные цветения цианобактерий Microcystis aeruginosa Kutz. Emend Elenk. И Lyngbia limnetica Lemm. в центральной восточной части Азовского моря отмечались, в частности, по данным натурных измерений в июне 2005 г. (Селифонова, 2005). В августе ЦБ достигают юго-восточной части Азовского моря, откуда выносятся в центральную часть бассейна. Как показано в работе (Алескерова и др., 2019), такое распространение связано с ячейкой антициклонической циркуляции течений, которая возникает в восточной части Азовского моря, вероятно под влиянием интенсивного речного стока Дона и Кубани. Максимальные величины занимаемой площади цветения наблюдаются в летне–осенний период с июня по октябрь (рис. 5). В ноябре ЦБ наблюдаются редко, по большей части в Таганрогском заливе и вдоль восточного берега Азовского моря с вероятностью не более P = 0.2. В некоторые годы спутниковые измерения отмечает наличие ЦБ в Таганрогском заливе и в зимние месяцы, что также было отмечено в работе (Теренько 2010) по измерениям в декабре 2009 г.
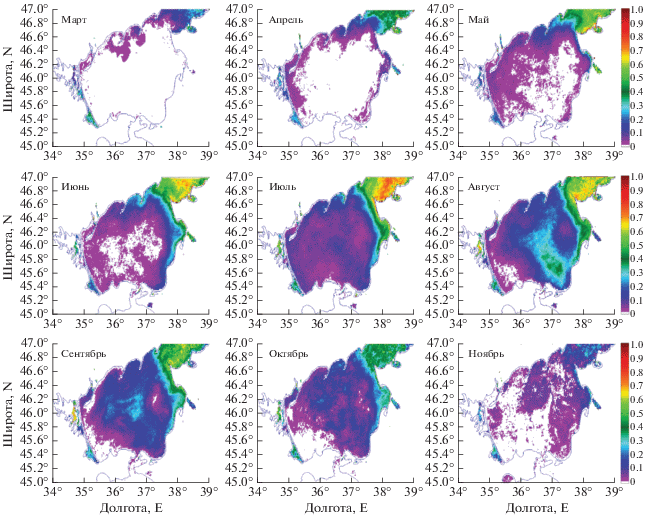
Примеры эволюции цветений в различные годы
Рассмотрим распространение цветения ЦБ в Азовском море на примере 2012 г. (рис. 6). В апреле 2012 г. ЦБ наблюдаются в Таганрогском заливе. Вместе с водами залива они распространяются вдоль северного берега на восток до Бердянска. Локальный очаг цветения ЦБ в это же время наблюдается у Ахтарского лимана. Такая же карта вероятности наблюдения ЦБ повторяется и в мае. Но в мае, кроме раннее обозначенных районов, ЦБ встречаются в небольшой области в Темрюкском заливе, около стока реки Кубань с вероятностью Р = 0.3. К июню цветение распространилось вдоль всего восточного побережья вплоть до Темрюкского залива. Источниками цветения здесь можно выделить Ахтарский лиман, реки Кубань и Протока. В июле цветению подвержено практически все Азовское море. Максимальная вероятность их наблюдения сохраняется для Таганрогского залива и вдоль восточного побережья.
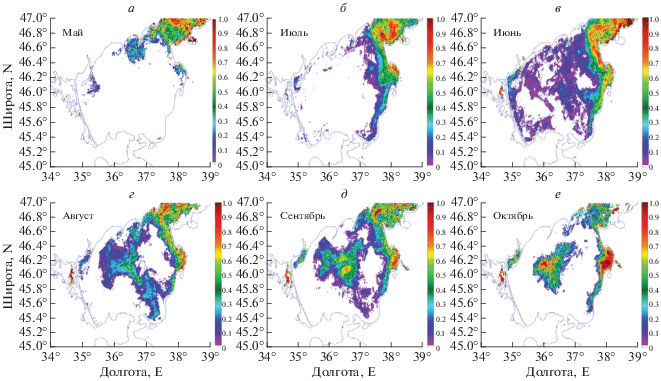
В августе цветение ЦБ под действием антициклонических течений распространилось в центральную часть Азовского моря. В сентябре происходит разрушение возникшей антициклонической ячейки и формируется локальная область цветения в центре моря с максимумом наблюдения ЦБ около Р = 0.8. В октябре выделяются две зоны, где сохраняется цветение ЦБ: в центральной части моря и вдоль восточного берега.
Детальный анализ показывает, что цветения характеризуются более короткопериодной изменчивостью. Рассмотрим пример эволюции цветения ЦБ в 2013 г. (рис. 7). Цветение ЦБ первоначально возникает в мае в Таганрогском заливе. В начале июня воды, подверженные цветению распространяются с Таганрогского залива вдоль восточного берега Азовского моря до Ясенского залива. К середине июня площадь ЦБ значительно увеличивается. Водоросли наблюдаются в Таганрогском заливе, вдоль северного берега Азовского моря до Обиточной косы, в Ясенском заливе и у стока рек Кубань и Притока. После 19 июня (рис. 7, в) цветение ЦБ разрушается. Это резкое уменьшение вероятно связано с действием шторма 19 июня, когда скорость ветра достигла 8–10 м/с. Ветровое воздействие приводит к усилению перемешивания. В результате верхний слой, в котором создаются наиболее благоприятные условия для азотфиксирующих ЦБ, разрушается, что приводит к ослаблению цветений (см. например Hunter et al., 2008). К концу июня и начала июля сохраняются две локальные области с высоким содержанием ЦБ: Таганрогский и Ясенский заливы. Цветение ЦБ в Азовском море снова усиливается к середине июля (рис. 7, д). В данный период времени ЦБ наблюдаются вдоль всего восточного побережья Азовского моря и начинают концентрироваться в локальной области в центре моря, которая отчетливо идентифицируется 16 июля. Более детально цветение ЦБ в это время наблюдается на RGB-композите, полученном по спутниковому снимку Landsat-8 с высоким разрешением (рис. 7, е). Протяженность данной области ориентировочно составляет около 70 км с севера на юг, а ширина – около 12 км. Площадь данного цветения составила около 1200 км2. С 19 июля образовавшаяся локальная область цветения начинает разрушаться, и воды, содержащие ЦБ, распространяются к северным берегам Азовского моря. Разрушение данного цветения в середине июля вероятно связано с увеличением скорости ветра до 10 м/с 19 и 21 июля.
Рис. 7.
Динамика развития цветения ЦБ в летний период 2013 г. а–д, ж–и – цветения ЦБ отмечены черным контуром, которые наложены на карту MODIS хлорофилла А (цветовая шкала); е – RGB снимок Landsat-8 за 18 июля 2013 г.
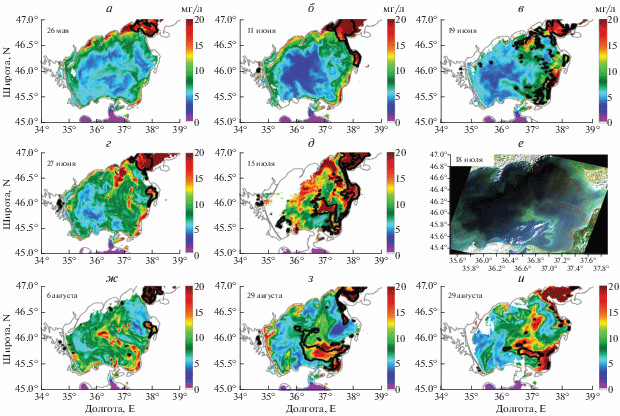
К началу августа ЦБ вновь наблюдаются только в Таганрогском и Ясенском заливах. Следующая вспышка цветения ЦБ в центральной части Азовского моря наблюдается к середине августа. Максимальная область распространения ЦБ фиксируется 22 августа (рис. 7, з). К 29 августа площадь данного цветения значительно сокращается. В сентябре ЦБ наблюдаются только в Таганрогском и Ясенском заливах.
Межгодовая изменчивость цветения ЦБ
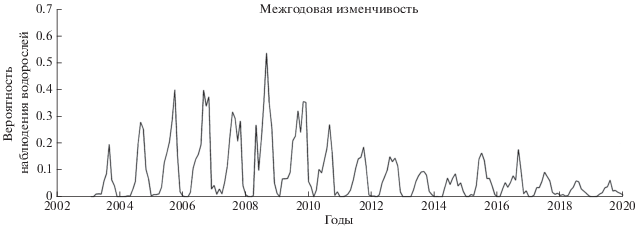
Предложенный спектральный алгоритм идентификации позволил оценить межгодовую изменчивость площади, занятой цветениями ЦБ в Азовском море за период 2003–2019 гг. Доля площади определялась как количество пикселей индетифицированных, как цветение ЦБ, деленное на общую площадь Азовского моря.
На графике межгодовой изменчивости (рис. 8 – черная линия) выделяется период 2004–2011 гг. с высокими значениями площадей цветения ЦБ, за которым следует резкое снижение в 2011–2020 гг. Максимальное количество ЦБ наблюдалось в 2008 г. со значениями в летний период около 0.55. В 2005–2007 и 2009 гг. максимальная площадь, занятая ЦБ, составляла порядка 0.4. Из графика видно, что площадь цветения ЦБ в летний период снизилась в последние годы от значений 0.3–0.4 (30–40%) до 0.05–0.1 (5–10%). Небольшое увеличение вероятности наблюдения ЦБ, (Р = 1.5) в последние годы наблюдалось в 2015–2016 гг.
Рис. 8.
График межгодовой изменчивости площади, занятой цветениеми ЦБ в Азовском море по данным MODIS.
На рис. 9 представлены среднегодовые карты вероятности наблюдения ЦБ в некоторые годы, полученные по данным сканера MODIS. Максимальные цветения ЦБ в 2006, 2008 и 2009 гг. практически полностью покрывают всю поверхность моря. В 2006 и 2009 (рис. 9, г) году четко наблюдается распространение ЦБ вдоль юго-восточного берега и их последующий вынос в центральную часть моря. В ходе экспедиции в декабре 2009 г. на станции, расположенной в Бердянском заливе были отобраны пробы фитопланктона, в которых наблюдалось практически полное доминирование ЦБ, составляющие по численности более 65%. Доминирование ЦБ (до 93%) было отмечено в северном и центральном районах моря, где отмечалось цветение O. amphybia (22.71 г/м3) (Теренько, 2010). Эти результаты согасуется с анализом данных MODIS (рис. 9, г), что свидетельствует о пригодности разработанного алгоритма для идентификации цветении ЦБ.
Рис. 9.
Среднегодовое пространственное распределение вероятности наблюдения ЦБ в некоторые годы по данным MODIS.
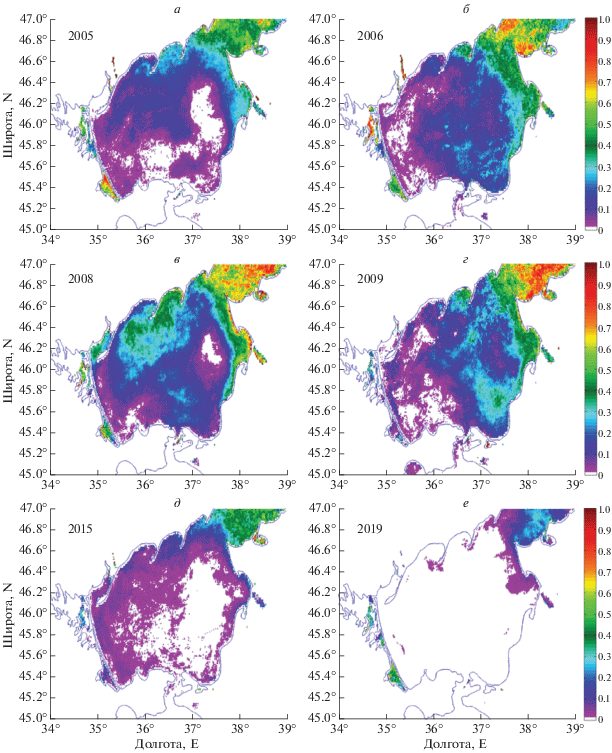
В 2008 г. в Азовском море наблюдалось максимальное цветение ЦБ вдоль северного и восточного берегов (рис. 9, в). В юго-восточной части они проникали в центральную часть Азовского моря, покрывая в итоге почти всю его акваторию.
В 2005, 2015 г. наблюдается отчетливое преобладание распространения ЦБ вдоль северного берега, что менее характерно для Азовского моря (рис. 4, а; рис. 5). В июне 2005 г. была проведена экспедиция в центральной части Азовского моря и вдоль его восточного берега в ходе которой была измерена средняя биомасса фитопланктона, которая достигала 2.7 г/м3. В его составе доминировали ЦБ Microcystis aeruginosa Kutz. emend Elenk и Lyngbia limnetica Lemm. (70–99% биомассы). На отдельных станциях их биомасса достигала 8 г/м3 (Селифонова, 2008). Таким образом полученное распределение на рис. 9, а согласуется с данными контактных измерений (Селифонова, 2008).
Минимальные по площади цветения наблюдались в последние годы исследуемого периода – 2018, 2019 гг. Для примера, в 2019 году (рис. 9, е) цветение ЦБ в основном наблюдалось только в Таганрогском заливе, с небольшим проявлением в южном направлении до Ясенского залива и возле Ахтарского лимана.
Влияние физических факторов на временную изменчивость цветения ЦБ
На изменчивость цветения влияет несколько факторов, среди которых важное значение играет речной сток рек, который определяет соленость и приток биогенов, и скорость ветра, определяющая устойчивость верхнего слоя (Berg, Sutula, 2015). Анализ межгодовой изменчивости среднего речного стока (красная линия на рис. 10), показывает, что речной сток значительно снизился после 2008 г. Его максимальные значения были отмечены в 2004–2006 гг. (1200 м3/с), за которым следовало резкое уменьшение. В 2009–2018 гг. его значения были практически в 2 раза ниже (600 м3/с). Локальное увеличение стока до 900 м3/с отмечается также в 2008 г., в котором наблюдается максимальное значение площади цветений ЦБ за 2003–2019 гг. Совпадение наблюдающихся отрицательных трендов и локальных пиков свидетельствует о том, что уменьшение речного стока послужило важной причиной снижения интенсивности цветений ЦБ.
Рис. 10.
График межгодовой изменчивости площади, занятой цветениеми ЦБ в Азовском море по данным MODIS (черная линия – вероятность наблюдения ЦБ, суммарного среднегодового стока рек Дона и Кубани (фиолетовая линия), среднегодовой скорости ветра за май–сентябрь над бассейном (синяя линия).
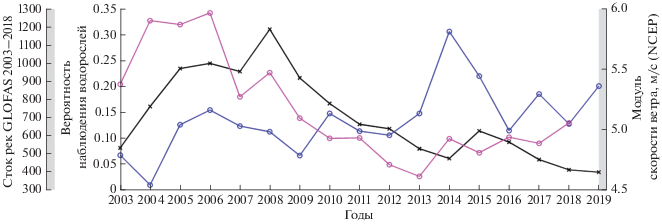
Еще одним важным фактором, влияющим на цветения ЦБ, может выступать изменения скорости ветра (синяя линия на рис. 10). Развитию ЦБ благоприятствуют условия устойчивой стратификации, которая позволяет им оставаться в поверхностном слое (Hunter et al., 2008; Wynne et al., 2010). В последние годы (после 2013, см. рис. 10) в регионе отмечается усиление скорости ветра (Kubryakov et al., 2019) в теплый период времени, которое вероятно также частично способствует уменьшению цветений. Воздействие ветра вызывает разрушение стратификации в верхних слоях, которое способствует перемешиванию вод, и как следствие разрушение цветений. Наименьшие значения площади ЦБ отмечены в 2014, 2018–2019 гг. Скорость ветра в летние периоды этих лет значительно возрастала. Поэтому несмотря на то небольшое увеличение стока рек, площади цветений были минимальны. Таким образом, изменение условий ветрового перемешивания также является одним из факторов, вызвавших ослабление цветений ЦБ в Азовском море. Это подтверждается подробным анализом динамики цветения в 2013 г. (рис. 7), который показывает, что цветение ЦБ разрушается при усилении ветра и наоборот, его локальные очаги возникают при безветренной погоде.
ВЫВОДЫ
Предложенный спектральный алгоритм автоматической идентификации ЦБ по данным сканера MODIS позволил оценить пространственную, сезонную и межгодовую изменчивость ареалов цветения ЦБ в Азовском море за исследуемый период 2003–2019 гг. С декабря по март встречаемость ЦБ близка к нулю, а в теплый период возрастает с максимумом в августе. В зависимости от года, цветение может быть как локализовано в Таганрогском заливе (2019 г.), так и распространится по всей акватории моря (2008, 2009 гг.). При этом в центральную часть моря ЦБ попадают в результате их антициклонического движения вдоль восточного берега. Полученные данные о районах встречаемости ЦБ согласуются с результатами контактных исследований (Селифонова, 2008; Matishov et al., 2009). Спутниковые данные свидетельствуют, что влияние потенциально токсичных ЦБ на Азовское море в последние годы уменьшается. Анализ межгодовой изменчивости показал, что цветения ЦБ были максимальны в 2007–2009 гг., после чего наблюдается резкое снижение их интенсивности. Возможными причинами этого является уменьшение стока рек и рост скорости ветра, которые ухудшают условия для развития ЦБ в верхнем слое.
Список литературы
Андреева Н.А., Мельников В.В., Снарская Д.Д. Роль цианобактерий в морских экосистемах // Биология моря. 2020. Т. 46. № 3. С. 161–173.
Ильин Ю.П., Фомин В.В., Дьяков Н.Н., Горбач С.Б. Гидрометеорологические условия морей Украины. Т. 1. Азовское море //Севастополь: ЭКОСИ-Гидрофизика. 2009. С. 400.
Карабашев Г.С., Евдошенко М.А. Спектральные признаки цветения цианобактерий в Балтийском море по данным сканера MODIS // Современные проблемы дистанционного зондирования Земли из космоса. 2015. Т. 12. № 3. С. 158.
Ковалева Г.В. Микроводоросли бентоса, перифитона и планктона прибрежной части Азовского моря. 2006.
Ковалёва Г.В. Проблема “цветения” воды в Азовском море // Труды Южного научного центра Российской академии наук. 2020. Т. 8. С. 122–148.
Макаревич П.Р., Ларионов В.В. Особенности строения фитопланктонных сообществ в зонах градиентов солености бассейна Азовского моря // Альгология. 2006. № 16. № 2. С. 216–226.
Матишов Г.Г., Фуштей Т.В. К проблеме вредоносных “цветений воды” в Азовском море // Исследовано в России. 2003. Т. 6. С. 213–225.
Никитина А.В. Численное решение задачи динамики токсичных водорослей в Таганрогском заливе // Известия Южного федерального университета. Технические науки. 2010. Т. 107. № 6. С. 113–116.
Селифонова Ж.П. Функционирование экосистемы Азовского моря // Биология внутренних вод. 2008. № 3. С. 3–7.
Сорокина В.В., Бердников С.В. Биогенная нагрузка Дона и Кубани на экосистему Азовского моря // Водные ресурсы. 2018. Т. 45. № 6. С. 670–684.
Теренько Г.В., Грандова М.А. Современное состояние фитопланктонного сообщества Украинского сектора Азовского моря в декабре 2009 г // Наукові записки Тернопільського національного педагогічного університету імені Володимира Гнатюка. Серія: Біологія. 2010. № 3. С. 44.
Ясакова О.Н. Динамика фитопланктона в районе Темрюкского залива Азовского моря // Ботанический журнал: Санкт-Петербург. 2015. Т. 100. № 10. С. 1015–1029.
Aleskerova A.A., Kubryakov A.A., Stanichny S.V., Lishaev P.N., Mizyuk A.I. Cyanobacteria Bloom in the Azov Sea According to Landsat Data // Izvestiya, Atmospheric and Oceanic Physics. 2019. T. 55. № 9. C. 1416–1426.
Berg M., Sutula M. Factors affecting the growth of cyanobacteria with special emphasis on the Sacramento-San Joaquin Delta // Southern California Coastal Water Research Project Technical Report. 2015. C. 869.
Blondeau-Patissier D., Gower J.F., Dekker A.G., Phinn S.R., Brando V.E. A review of ocean color remote sensing methods and statistical techniques for the detection, mapping and analysis of phytoplankton blooms in coastal and open oceans // Progress in oceanography. 2014. T. 123. C. 123–144.
Harke M.J., Steffen M.M., Gobler C.J., Otten T.G., Wilhelm S.W., Wood S.A., Paerl H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae. 2016. 54. C. 4–20.
Harrigan S., Zsoter E., Alfieri L., Prudhomme C., Salamon P., Wetterhall F., … Pappenberger F. GloFAS-ERA5 operational global river discharge reanalysis 1979–present // Earth System Science Data. 2020. T. 12. № 3. C. 2043–2060.https://doi.org/10.5194/essd-2019-232)
Hunter P.D., Tyler A.N., Willby N.J., Gilvear D.J. The spatial dynamics of vertical migration by Microcystis aeruginosa in a eutrophic shallow lake: A case studyusing high spatial resolution time-series airborne remote sensing. Limnol. Oceanogr. 2008. 53: 2391–2406.
Kahru M., Savchuk O.P., Elmgren R. Satellite measurements of cyanobacterial bloom frequency in the Baltic Sea: interannual and spatial variability // Marine Ecology Progress Series. 2007. T. 343. C. 15–23.
Kutser T., Metsamaa L., Strömbeck N., Vahtmäe E. Monitoring cyanobacterial blooms by satellite remote sensing //Estuarine, Coastal and Shelf Science. 2006. T. 67. № 1–2. C. 303–312.
Kubryakov A., Stanichny S., Shokurov M., Garmashov A. Wind velocity and wind curl variability over the Black Sea from QuikScat and ASCAT satellite measurements. Remote sensing of environment. 2019. 224, 236–258.https://doi.org/10.1016/j.rse.2019.01.034
Lavrova O.Y., Mityagina M.I. Manifestation specifics of hydrodynamic processes in satellite images of intense phytoplankton bloom areas // Izvestiya, Atmospheric and Oceanic Physics. 2016. T. 52. № 9. C. 974–987.
Matishov G.G. Matishov D.G., Stepanian O.V., Aksenov D.S. Complex studies of the Azov, Black, and Caspian seas performed on the Deneb survey vessel in 2007 // Oceanology. 2009. T. 49. № 2. C. 290.
Mizyuk A.I. Korotaev G.K., Grigoriev A.V., Puzina O.S., Lishaev P.N. Long-Term Variability of Thermohaline Characteristics of the Azov Sea Based on the Numerical Eddy-Resolving Model // Physical Oceanography. 2019. T. 26. № 5. C. 438–450.
Paerl H.W., Otten T.G.. Harmful cyanobacterial blooms: causes, consequences, and controls // Microbial ecology. 2013. 65(4). C. 995–1010.
Selifonova Z.P. The role of zooplankton in the functioning of the Taganrog Gulf ecosystem in the Sea of Azov // Inland Water Biology. 2010. T. 3. № 4. C. 335–343.
Sivonen K., Jones G.J. Cyanobacteria toxins. In: Chorus I., Betram, (Eds.), Toxic Cyanobacteria in Water: A Guide to Public Health, Significance, Monitoring and Management. The World Health Organization / E and F.N. Spon. 1999. C. 41–51.
Vincent R.K., Qin X., McKay R.M.L., Miner J., Czajkowski K., Savino J., Bridgeman T. Phycocyanin detection from LANDSAT TM data for mapping cyanobacterial blooms in Lake Erie // Remote Sensing of Environment. 2004. T. 89. № 3. C. 381–392.
Wynne T.T., Stumpf R.P., Tomlinson M.C., Dyble J. Characterizing a cyanobacterial bloom in western Lake Erie using satellite imagery and meteorological data. Limnology and Oceanography. 2010. 55(5). C. 2025–2036.
Дополнительные материалы отсутствуют.
Инструменты
Исследование Земли из Космоса