

Исследование Земли из Космоса, 2022, № 1, стр. 80-91
Использование гиперспектральных спутниковых данных для идентификации цианобактерий в Чёрном и Азовском морях
Е. А. Морозов *
Морской гидрофизический институт РАН
Севастополь, Россия
* E-mail: frisman@list.ru
Поступила в редакцию 29.10.2020
- EDN: IYTVAA
- DOI: 10.31857/S0205961422010055
Аннотация
Использованы данные гиперспектрального спутникового датчика Hyperspectral Imager for the Coastal Ocean (HICO, NASA) для идентификации и качественной оценки содержания цианобактерий (ЦБ) в прибрежных акваториях Черного и Азовского морей. Применены три алгоритма обработки данных: два использующие анализ формы спектра и один полуаналитический. Первый алгоритм использует определение минимума спектра коэффициента отражения для дистанционного зондирования в области спектра около 680 нм, так называемый индекс присутствия ЦБ. На основе данного алгоритма разработан второй алгоритм для идентификации присутствия в воде фикоцианина, являющегося пигментом-маркером для ЦБ. Второй алгоритм анализирует минимум спектра коэффициента отражения для дистанционного зондирования (Rrs) в области 620 нм. Третий полуаналитический алгоритм позволяет определять концентрацию фикоцианина. Произведено сопоставление результатов автоматической идентификации присутствия ЦБ с визуальным анализом спектров. Произведено сопоставление результатов применения трех алгоритмов к изображениям. Сделан вывод, что фикоцианин присутствует в исследованных акваториях, и что его присутствие является следствием наличия ЦБ.
ВВЕДЕНИЕ
Исследования фитопланктона с помощью спутниковых данных шагнуло далеко за пределы определения концентрации хлорофилла-а (Хл-а). Активно разрабатываются как методы, нацеленные на исследования других параметров фитопланктона, например, размерной структуры, видового состава, идентификации вредоносных цветений водорослей, так и идентификации других пигментов помимо, Хл-а (Uitz et al., 2008; IOCCG, 2014; Bracher et al., 2017). Такое развитие данного научного направления связано как с необходимостью извлекать максимум информации из спутниковых данных о цвете океана (IOCCG, 2014), так и с увеличением количества и качества разнообразных датчиков цвета океана на околоземной орбите, включая новые гиперспектральные (Groom et al., 2019).
В данном исследовании была поставлена задача идентификации ЦБ (сине-зеленых водорослей) по данным спутникового дистанционного зондирования. Сине-зеленые водоросли привлекают внимание исследователей в виду их токсичности и способности создавать обширные интенсивные цветения. Это может приводить к негативному воздействию на рыболовную и рекреационною отрасли. Немаловажным также является учащение таких цветений в различных частях света вследствие увеличивающейся антропогенной нагрузки на водные объекты, изменения климата и, возможно, и других факторов. Исследование данной проблемы требует методов, не только позволяющих регистрировать аномальные цветения, но и идентифицировать присутствие этих водорослей в небольших количествах. Цель – исследования проложить путь к определению содержания ЦБ, содержащихся в воде в любом количестве, создать работоспособные алгоритмы обработки гиперспектральных данных.
Пигмент фикоцианин является уникальным пигментом для ЦБ в пресноводных акваториях (Randolph et al., 2008; Castenholz, 2001). Этот факт неоднократно использовался различными авторами для разработки алгоритмов идентификации экстремальных цветений ЦБ в озерах. В морских водах фикоцианин может встречаться не только в цианобактериях, но и в классах Rhodophyta и Cryptophyta (Wright et al., 2005). Несмотря на то, что в морских акваториях этот пигмент не уникален для ЦБ, количество видов фитопланктона, которые его содержат весьма ограничено.
Существует множество методов идентификации и картирования ЦБ (Stumpf et al., 2016). Для целей нашего исследования можно условно разделить их на методы, которые идентифицируют ЦБ с использованием максимума поглощения фикоцианина на 620 нм (Schalles & Yacobi, 2000; Randolph et al., 2008) и те, которые ориентируются на косвенные спектральные признаки. Например, на основе общей формы спектра Rrs (Sun et al., 2015; Карабашев и Евдошенко, 2015) или максимума коэффициента отражения в области 680 нм (Wynne et al., 2008; Wynne et al., 2013). Обе группы содержат алгоритмы как для определения концентрации, так и для качественной идентификации.
В данном исследовании была поставлена задача идентификации (обнаружения) ЦБ Черном и Азовском морях. Данный выбор обусловлен следующими причинами:
1) ЦБ уже исследовались коллективом МГИ по данным мультиспектральных датчиков MODIS и Landsat (Алескерова и др., 2018; Kubriakov et al., 2021). с использованием статистической взаимосвязи между формой спектра в сине-зеленой области и вероятностью присутствия сине-зеленых водорослей в северо-западе ЧМ и АМ. Таким образом, данное исследование является развитием существующего направления и попыткой осветить вопрос цветения ЦБ в Черном и Азовском морях с использованием нового типа данных.
2) Использование максимума поглощения фикоцианина в качестве спектрального признака присутствия сине-зеленых водорослей предпочтительно с точки зрения интерпретации результатов. Именно фикоцианин является однозначным индикатором присутствия ЦБ. Такой подход позволит однозначно определить наличие сине-зеленых водорослей без привлечения дополнительной информации (например, данных о частоте или интенсивности цветений сине-зеленых водорослей в исследуемой области). Использование же особенностей других областей спектра требует набора in situ и спутниковых данных, с количеством измерений, достаточным, чтобы статистически достоверно определить взаимосвязь между областями спектра не подверженными интенсивному влиянию фикоцианина и присутствием ЦБ.
3) Использование гиперспектральные данных о цвете моря позволяет исследовать содержание отдельных пигментов в воде с недоступной для мультиспектральных датчиков точностью, благодаря детальному спектральному разрешению. Также для исследовательской работы полезно иметь более одного канала в области 620 нм, для изучения интенсивности и ширины спектральных особенностей, обусловленных присутствием фикоцианина. На данный момент гиперспектральные данные, особенно спутниковые используются не очень интенсивно. С учетом планируемого в 2022 запуска гиперспектрального датчика цвета вод Ocean Color Imager (Groom et al., 2019) следует ожидать увеличения доступности такого типа данных и разрабатывать алгоритмы их обработки.
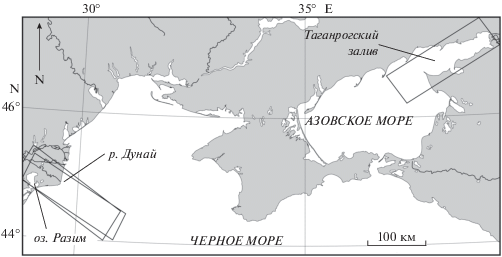
ИССЛЕДУЕМЫЙ РЕГИОН
Регион исследования определялся исходя из сочетания двух факторов: наиболее вероятного района присутствия ЦБ в Азовском море (АМ) и Черном море (ЧМ) и расположения областей с наилучшим покрытием спутниковыми данными гиперспектрального датчика HICO. Исследуемыми районами являются северо-запад ЧМ (СЗЧМ), район устья Дуная и Таганрогский залив (ТЗ) с близлежащими акваториями в АМ.
АМ является полузамкнутым внутренним водоемом, сообщающимся в своей южной части с ЧМ через неглубокий Керченский пролив. Средняя соленость вод АМ изменяется (1988–2000 гг.) от 10.4 до 11.8 и Таганрогского залива от 5.7 до 7.8. АМ и Таганрогский залив являются водоемами, соответственно, прибрежного и эстуарийного типа (Студеникина и др., 1999; Ларионов и Макаревич, 2001). Эвтрофикация и пониженные значения солености способствуют цветению ЦБ в бассейне, – таких как Microcystis aeruginosa, Aphanisomenon flosaquae, Anabaena spp. и др. (Макаревич, Ларионов, 2006; Матишов и др., 2009; Лужняк, 2011; Lomakin, 2018).
Фитопланктонное сообщество СЗЧМ характеризуется тем, что уже в 2008–2010 г. наблюдается тенденция появления 3–4 пиков развития фитопланктона шельфовых водах: весна, лето, осень и зима, ранее исследователи отмечали, только 2–3: весенне–летний, осенний (Попов и др., 2010). Весенний максимум фитопланктона в значительной степени определяется влиянием весенних половодий на реках СЗЧМ (Попов и др., 2010). Наибольшее влияние оказывают Днепр и Южный Буг. Вклад различных видов в структуру фитопланктонного сообщества в открытых водах СЗЧМ изменился. Наиболее значительный рост числа видов наблюдался у Chlorophyta и Cyanobacteria. Увеличение видового разнообразия часто происходит за счет представителей пресноводных и солоноватоводных комплексов, среди которых много зеленых и сине-зеленых водорослей. Снижение солености создало благоприятные условия для развития этих представителей фитопланктона. (Попов и др., 2010, Теренько и Нестерова, 2015; Дорофеев и Сухих, 2020).
Учитывая распресненность прибрежных вод исследуемых акваторий, сообщения о присутствии ЦБ в них и тот факт, что Фц в морских акваториях встречается преимущественно в цианобактериях, мы можем принять Фц в качестве однозначного индикатора присутствия ЦБ.
АЛГОРИТМЫ ОБРАБОТКИ СПУТНИКОВЫХ ДАННЫХ
В данном исследовании было применено три алгоритма обработки спутниковых данных с целью идентификации ЦБ и фикоцианина. Первый алгоритм разработан Вэйном с соавторами (Wynne et al., 2008; Wynne et al., 2013) и использует параметр, описывающий форму спектра на длине волны 681 нм. Второй алгоритм разработан автором статьи и использует параметр описывающий форму спектра в области 620 нм. Третий алгоритм разработан Рандольфом с соавторами (Randolph et al., 2008) и является полуаналитическим алгоритмом для определения концентрации Хл-а и фикоцианина.
В качестве исходных данных для алгоритмов были использованы данные уровня L2 гиперспектрального спутникового датчика HICO космического агентства NASA (https://oceancolor.gsfc.nasa.gov/data/hico/). На данный момент это единственный гиперспектральный спутниковый датчик, разработанный специально для исследования цвета прибрежных вод. HICO получил более 10 000 изображений по всему миру за период пока он был установлен на Международной Космической Станции с сентября 2009 по сентябрь 2014. Недолгий период существования HICO делают затруднительным сбор синхронной базыx in situ и спутниковых данных, но несомненной выгодой является возможность использования специализированных гиперспектральных спутниковых данных видимого диапазона пока не доступны данные перспективных гиперспектральных спутниковых данных видимого диапазона. Длины волн (каналы) для данных L2 датчика HICO распределены в диапазоне 353–719 нм с шагом 5.7 нм. Данные представляют собой значения спектрального коэффициента рассеяния (коэффициента отражения для дистанционного зондирования), определяемого как Rrs (0, λ) = = Lu(0, λ)/Ed (0, λ), где Lu(0, λ) и Ed(0, λ) – соответственно, спектральная яркость восходящего света над поверхностью воды в надирном направлении и освещенность водной поверхности; λ – длина волны.
Алгоритм идентификации цветений ЦБ
Это эмпирический алгоритм, использующий статистическую связь формы спектра исходящего из воды излучения с наличием цветения ЦБ. Существуют и используются для мониторинга цветений ЦБ модификации алгоритма для датчиков MERIS (Wynne et al., 2008) и MODIS (Wynne et al., 2013). В данном исследовании использованы данные гиперспектрального датчика HICO. В алгоритме использованы три канала датчика HICO и выбраны длины волн близкие к длинам волн использованным в наиболее разработанной версии этого алгоритма, а именно версии для датчика MERIS. Таким образом, для HICO λ = 679 нм, λ‒ = 662 нм и λ+ = 708 нм и расхождение с оригинальными длинами волн MERIS не превышает 3 нм.
Форма спектра при 681 нм используется для того, чтобы отличить цветение ЦБ от цветения другого фитопланктона с помощью изображений спутникового датчика MERIS. Во время интенсивного цветения ЦБ спектр около 681 нм имеет понижение, поскольку рассеяние, вызванное ЦБ, подавляет сигнал флуоресценции, создавая тем самым понижение значений nLw или Rrs относительно значений в соседних каналах. Эта взаимосвязь согласуется как с данными дистанционного зондирования, так и с данными in situ (Wynne и др., 2008, Wynne и Stumpf, 2015).
Несколько биофизических факторов могут приводить к провалу в спектре в области 681 нм по сравнению с каналами 667 (или 665 нм). Поглощение Хл-а более интенсивно в области спектра около 680 нм, чем около 665 нм (Bricaud et al. 1995), что понижает коэффициент отражения или яркость в области 680 нм относительно соседних областей спектра. Существует также свидетельство того, что ЦБ не флуоресцируют так же сильно, как другие типы водорослей (Seppälä et al. 2007). Существуют и другие интерпретации и исследования, посвященные механизмам образования максимума или минимума в области 680 нм, например, (Huot, Brown и Cullen, 2005; Gower и др., 1999; Wynne и др., 2013).
Алгоритм идентификации фикоцианина
Алгоритм идентификации фикоцианина в воде по спутниковым данным был разработан автором статьи по аналогии с предыдущим алгоритмом. Был выбран подход на основе анализа максимума поглощения фикоцианина на 620 нм с использованием гиперспектральных спутниковых данных. Теоретическим обоснованием является тот факт, что фикоцианин, содержащийся в ЦБ, имеет единственный максимум поглощения на длине волны 620 нм. Величина провала (минимума) в этой области должна коррелировать с содержанием фикоцианина. Подход на основе определения высоты линии флуоресценции, аналогичный Wynne и др. (2008) может быть использован для идентификации и количественной оценки этого провала. Использование гиперспектральных данных позволяет экспериментировать с выбором λ, λ– и λ+ и подобрать сочетание каналов, для выделения минимума, обусловленного фикоцианином в спектре Rrs на фоне влияния других оптически активных компонентов воды.
Было протестировано несколько вариантов алгоритма (с использованием различных комбинаций каналов в области 620 нм). Результаты получались качественно и количественно сходными. В статье приводятся данные для двух наиболее сильно различающихся вариантов исполнения, с λ–, λ и λ+ равными, соответственно, 605, 622 и 633 нм, и 610, 622 и 627 нм. Наиболее оптимальными можно считать длины волн 610, 622 и 633 нм, позволяющие получить один из наиболее строгих критериев идентификации. На рис. 2 и рис. 3 видно, что провал в спектрах в диапазоне 600–645 нм присутствует практически всегда, поэтому использование каналов, наиболее близких к длине волны 620 нм, позволяет минимизировать влияние пигментов с максимумом поглощения, близким к 620 нм. Алгоритм рассматривает область спектра вблизи 620 нм – это максимум поглощения света фикоцианином. Недостатком может являться то, что вблизи поглощают другие пигменты (Хл-с, Хл-b, фикоэритрин), которые могут снижать точность определения этим методом, особенно если набор пигментов в воде будет сильно меняться во времени или в пространстве. Эту особенность нужно учитывать, сравнивая значения ФЦИ полученные в разные сезоны и в различных акваториях, так как одни и те же значения индекса могут соответствовать различным концентрациям Фц.
Рис. 2.
Иллюстрация каналов, используемых для расчета ИЦ (λ– = 662 нм, λ = 679 нм и λ+ = 708 нм) и ФЦИ (Вариант № 1: λ– = 605 нм, λ = 622 нм и λ+ = 633 нм; Вариант № 2: λ– = 610 нм, λ = 622 нм и λ+ = 627 нм). Цифры 622 и 679 обозначают λ для ФЦИ и ИЦ, соответственно. Врезка в левой нижней стороне иллюстрирует базовый уровень для ФЦИ в варианте № 1 – черная линия, и варианте № 2 – красная линия. Спектры Rrs уровня L2, HICO, 2 сентября 2014 г.
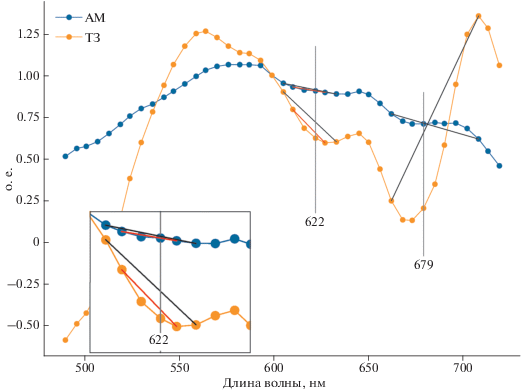
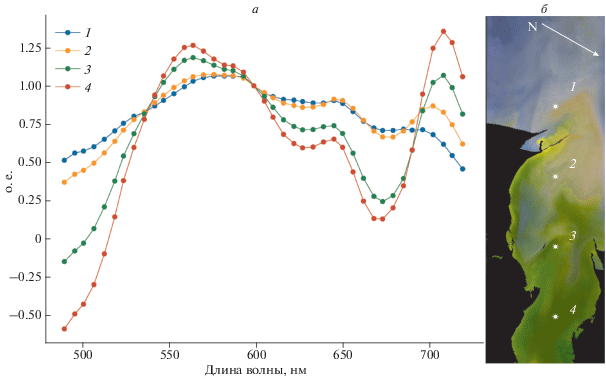
Рис. 3.
Изменение спектров от мористой к устьевой части ТЗ (а). Спектры нормированы на длину волны 599 нм и представлены в относительных единицах. Расположение точек, для которых построены спектры на фоне RGB изображения ТЗ (б). Белая стрелка указывает направление на север. Изображение HICO за 2 сентября 2014 г.
Коэффициент корреляции между результатами, получаемыми по вышеприведенным вариантам исполнения алгоритма определения ФЦИ составил 0.76 для 9-ти изображений в СЗЧМ и 0.87 для 4 изображений в ТЗ.
Информационный продукт, получаемый в результате применения данного алгоритма, был назван фикоцианиновым индексом (ФЦИ). Преимуществом ФЦИ перед ИЦ является то, что он реагирует непосредственно на максимум поглощения фикоцианина, а не на результат сложного взаимодействия флуоресценции Хл-а любого фитопланктона с поглощением и рассеянием различными оптически активными компонентами в воде. Тем не менее, область максимума поглощения фикоцианина подвержена влиянию Хл-а, хлорофилла-б (Хл-б) и поглощения желтым веществом. Это может являться причиной неустойчивости ФЦИ при различных гидрооптических условиях, например, возможны различные типы ошибок при повышенных прибрежных стоках и цветениях фитопланктона, не являющегося ЦБ. Приближение λ– и λ+ к 620 нм является попыткой, избежать ложных положительных идентификаций. В качестве примера можно взять также спектры 1 и 4 на рис. 3, приведенном ниже. Спектр 1, соответствующий водам с меньшим содержанием фитопланктона содержит два минимума в интересующей нас области: 620 нм (Фц) и 630–640 нм (предположительно, Хл-б). В более эвтрофных водах, которым соответствует спектр 4 эти два минимума сливаются. В такой ситуации разнесение λ– и λ+ дальше от 620 нм повышает вероятность что алгоритм будет срабатывать не на Фц, а на какой-то другой пигмент или их комбинацию.
Полуэмпирический алгоритм
Алгоритм был разработан для определения количественных значений концентрации фикоцианина (Simis et al., 2005).
Алгоритм был применен Randolph (2008) с соавторами к гиперспектральным судовым данным в двух водохранилищах. Детальное описание применение алгоритма к гиперспектральным данным Rrs дано в (Randolph et al., 2005). Авторы определяют поглощение Хл-а и Фц на 665 и 620 нм, соответственно. Для оценки концентрации Фц был применен коэффициент удельного поглощения Фц на 620 нм, равный 0.0070 м2 мг Фц–1. В результате проверки по контактным данным точность алгоритма характеризовалась коэффициентом детерминации r2 = 0.85 (при числе проверочных измерений in situ, n = 48, и уровне значимости p < < 0.0001), и среднеквадратической ошибке СКО = = 19.54 мг/м3.
С целью успешного применения данного алгоритма к спутниковым данным датчика HICO уровня L2, в алгоритме были произведены модификации. Была произведена интерполяция каналов, чтобы соответствовать длинам волн, использованным Randolph et al., 2008. Rrs620 был получен интерполяцией данных Rrs622 и Rrs616. Использовалось поглощение водой на 719 нм (Pope and Fry, 1997) вместо 778 нм, так как 719 нм – это максимальная длина волны содержащаяся в стандартных данных HICO уровня L2.
В процессе расчетов определяется концентрация Хл-а которая используется для введения поправки на поглощение Хл-а в области поглощения фикоцианина. Затем рассчитывается концентрация фикоцианина в мг/м3. Несмотря на то, что алгоритм определяет количественные значения, в нашем случае их стоит использовать как ориентировочные, так как алгоритм был разработан для другого региона и нами были внесены упомянутые выше изменения, а данные in situ для проведения проверки и настройки алгоритма на момент проведения исследования не доступны.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Исследование, описываемое в данной статье, состоит из двух частей. В первой визуально анализируются спектры прибрежных вод с целью идентификации признаков присутствия ЦБ и фикоцианина. Во второй части производится применение трех вышеописанных алгоритмов к данным HICO в исследуемых регионах.
Анализ спектров
На рис. 3 представлено изменение спектров коэффициента отражения для дистанционного зондирования от мористой части ТЗ к кутовой его части. На спектре № 1 просматриваются отдельные небольшие минимумы в каналах 616 и 633–639 нм, являющиеся следствием поглощения Фц и Хл-б, соответственно. Максимум поглощения Фц и Хл-б располагается на 620 и 650 нм, соответственно (Ruiz-Verdú и др., 2008). Эти минимумы смещены в синюю сторону спектра вследствие влияния поглощения желтым веществом. На спектре № 2, соответствующем устьевой части залива, слияние этих максимумов обуславливает общий минимум Rrs. На спектрах № 3 и 4 (рис. 3, а), расположенных в более биопродуктивных водах в ТЗ, эти минимумы полностью объединяются в один большой минимум, который в литературе часто интерпретируется как поглощение Фц и может использоваться для идентификации и определения концентрации Фц (Gitelson et al., 2011; Sun et al., 2015).
Обработка изображений
В данном разделе рассматриваются три изображения обработанные каждое тремя алгоритмами. На рис. 4 представлен ТЗ. Все три алгоритма показывают очень схожие пространственные распределения. Это подтверждается коэффициентами корреляции в табл. 1, колонка “27 июня 2013, ТЗ”. Столь хорошая согласованность между тремя алгоритмами, скорее всего, связана с сильным влиянием стока реки Дон, что приводит к взаимосвязанным изменениям распределения фитопланктона, минеральной взвеси, желтого вещества и других параметров. Три алгоритма используют различные области спектра подверженные влиянию различного набора факторов, но мощный речной сток приводит к практически одновременному изменению этих факторов.
Рис. 4.
Сравнение распределения ИЦ (а), ИФЦ (б) и концентрации Фц (в) в прибрежных водах СЗЧМ. Черные стрелки указывают направление на север. Изображение за 27 июня 2013 г.
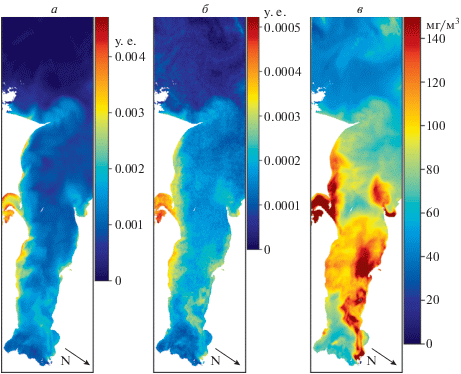
Таблица 1.
Коэффициенты корреляции (r) между ИЦ, ФЦИ и концентрацией Фц для данных, проиллюстрированных на рис. 4, 5 и 6.
| Параметр | 27 июня 2013, ТЗ | 10 июля 2014, СЗЧМ | 9 сентября 2014, СЗЧМ | |||
|---|---|---|---|---|---|---|
| ФЦИ | концентрация Фц | ФЦИ | концентрация Фц | ФЦИ | концентрация Фц | |
| ИЦ | 0.937 | 0.798 | 0.707 | 0.465 | 0.808 | –0.036 |
| ФЦИ | – | 0.890 | – | 0.669 | – | 0.072 |
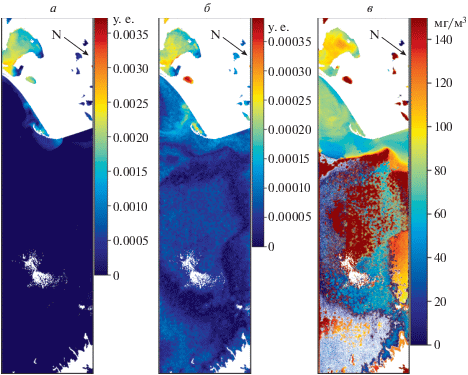
Рис. 5 и 6 иллюстрируют применение трех алгоритмов к данным в СЗЧМ в середине лета и начале осени, соответственно. Для изображения от 10 июля 2014 (рис. 5) важно отметить, что ИФЦ (рис. 5, б) в плане категоричности оценки ситуации занимает промежуточное положение между результатами, получаемыми по ИЦ (рис. 5, а) и распределением концентрации Фц (рис. 5, в). ИЦ идентифицирует ЦБ только в акватории озера Разим и в части плюма реки Дунай. ИФЦ и концентрация Фц (рис 5, б и 5, в, соответственно) показывают более плавное пространственное распределение. Это может быть связано с тем, что ИЦ является продуктом, разработанным для идентификации интенсивных и опасных цветений ЦБ и нечувствителен к низким концентрациям ЦБ. Полуаналитический алгоритм, напротив, определяет концентрацию Фц, которая по сути своей не может изменяться в пространстве резко, и показывает присутствие ЦБ, а не только их цветение. ИФЦ был разработан в данном исследовании, именно для того, чтобы иметь возможность качественно отслеживать пространственные вариации Фц, даже если он присутствует в концентрации ниже, чем при цветении (интенсивном развитии) ЦБ. Окончательный ответ на вопрос какое пространственное распределение является более соответствующим действительности может дать только привлечение данных in situ. Тем не менее, можно предположить, что алгоритмы построенные на основе использования канала максимума поглощения Фц в области 620 нм (ИФЦ и Фц) должны давать физически более обоснованный результат.
Рис. 5.
Сравнение распределения ИЦ (а), ИФЦ (б) и концентрации ФЦ (в) в прибрежных водах СЗЧМ. Черные стрелки указывают направление на север. Изображение HICO за 10 июля 2014 г.
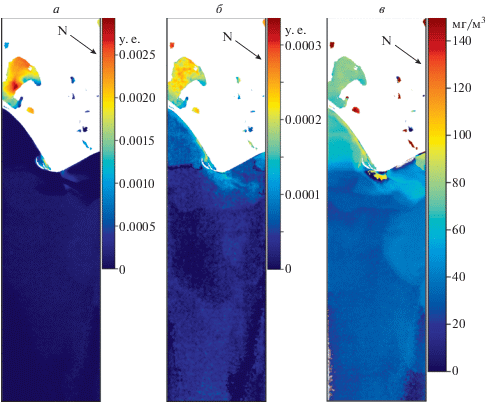
Рис. 6.
Сравнение распределения ИЦ (а), ИФЦ (б) и концентрации ФЦ (в) в прибрежных водах СЗЧМ. Черные стрелки указывают направление на север. Изображение HICO за 9 сентября 2014.
На рис. 6 представлено распределение ИЦ (рис. 6, а), ИФЦ (рис. 6, б) и концентрации Фц (рис. 6, в) 9 сентября 2014 г. Комментарии к распределению совпадают с таковыми для ситуации 10 июля 2014 г. Особенность же состоит в наличии в изображении большой области с ошибками атмосферной коррекции (рис. 6, в) в большей части изображения с мористой стороны. Эти области специально не были замаскированы, чтобы продемонстрировать устойчивость алгоритмов на основе анализа формы спектра (рис. 6, а, 6, б) к ошибкам атмосферной коррекции. Дополнительной причиной расхождений между ИФЦ и ФЦ может быть использование в алгоритме определения концентрации ФЦ коррекции учитывающей влияние поглощения Хл-а на область максимума спектра поглощения фикоцианина.
И ИЦ, и ФЦИ являются эмпирическими алгоритмами при картировании результатов обработки спутникового изображения они дают качественную картину распределения ЦБ. При наличии достаточного количества измерений in situ, есть возможность также эмпирически (статистически) связать значения ИЦ и ФЦИ в ограниченном регионе c концентрацией ЦБ. В литературных источниках можно встретить оптические данные с гиперспектральным разрешением, полученные в областях цветения ЦБ (см., например, Gitelson et al., 2011; Sun et al., 2015; Ruiz-Verdu et al., 2008), но эти примеры относятся в большинстве своем, к пресноводным акваториям и интенсивным цветениям ЦБ. В данном исследовании освещается вопрос как изменяется спектр (с гиперспектральным разрешением) Rrs при изменении содержания ЦБ в прибрежных морских водах и производится качественное картирование этого процесса с использованием различных алгоритмов обработки спутниковых данных.
Эмпирические алгоритмы ИЦ и ИФЦ показывают схожие результаты, что подтверждается результатами анализа пространственных распределений и коэффициентами корреляции. Это согласуется с литературными данными об устойчивости такого типа к ошибкам атмосферной коррекции (Stumps et al., 2016). Результаты применения полуэмпирического алгоритма в общем, согласуются со значениями ИЦ и ФЦИ (см. рис. 4 и 5, табл. 1, колонки “27 июня 2013, ТЗ” и “10 июля 2014, СЗЧМ”). Явным исключением являются случаи, когда полуэмпирический алгоритм определения концентрации Фц применяется к областям с ошибками в атмосферной коррекции (рис. 6 и табл. 1, колонка “9 сентября 2014, СЗЧМ”). Это является серьезной проблемой для практического применения, но не является внутренним недостатком алгоритма.
ЗАКЛЮЧЕНИЕ
1) К отдельным акваториям ЧМ и АМ применены три различных алгоритма: два для идентификации ЦБ и один для качественной оценки концентрации Фц.
2) Разработан и протестирован эмпирический алгоритм оценки присутствия Фц в водах природных объектов по гиперспектральным оптическим данным. Разработанный алгоритм может быть применен к гиперспектральным данным и прошедшим атмосферную коррекцию и позволяет на качественном уровне определять присутствие фикоцианина в природных водах. В таком виде он может считаться глобальным. Алгоритм может использоваться в автоматическом режиме. Данный алгоритм может являться основой для регионального эмпирического алгоритма количественного определения Фц или ЦБ в воде, в таком случае потребуется привлечение региональных данных in situ.
3) На основе гиперспектральных спутниковых данных идентифицировано присутствие Фц в прибрежных акваториях ЧМ и АМ. Сделано обоснованное предположение, что Фц является следствием присутствия ЦБ.
Для пикселей с качественной атмосферной коррекцией все три алгоритма показывают схожие результаты с корреляцией 0.47–0.94. Для пикселей, у которых в результате ошибок атмосферной коррекции наблюдались отрицательные значения Rrs в красной области спектра полуаналитический алгоритм не применим. Алгоритмы определения ИЦ (Wynne et al., 2005) и ИФЦ (данное исследование) даже в пикселях с некачественной атмосферной коррекцией срабатывают и дают схожие результаты (r = 0.71–0.94).
Список литературы
Алескерова А.А., Кубряков А.А., Станичный С.В., Лишаев П.Н., Мизюк А.И. Цветение цианобактерий в Азовском море по данным сенсоров спутников серии Landsat // Исслед. Земли из космоса. 2018. № 6. С. 52–64.
Дорофеев В.Л., Сухих Л.И. Исследование потоков компонентов морской экосистемы с северо-западного шельфа в глубоководную часть Черного моря // Морской гидрофизический журн. 2020. Т. 36. № 5. С. 501–513. https://doi.org/10.22449/0233-7584-2020-5-501-513
Карабашев Г.С., Евдошенко М.А. Спектральные признаки цветения цианобактерий в Балтийском море по данным сканера MODIS // Современные проблемы дистанционного зондирования Земли из космоса. 2015. Т. 12. № 3. С. 158–170.
Ларионов В.В., Макаревич П.Р. Общие закономерности развития фитопланктонных сообществ эстуарных областей Азовского и северных морей России // Среда, биота и моделирование экологических процессов в Азовском море. Апатиты: Изд-во Кольск. НЦ РАН. 2001. С. 88–97.
Лужняк О.Л. Развитие фитопланктона Таганрогского залива в весенне-летний период после исключительно теплой зимы 2006/2007 гг. // Изв. высш. уч. зав. Северо-Кавказский регион. Естественные науки. 2011. № 1. С. 67–71.
Макаревич П.Р., Ларионов В.В. Особенности строения фитопланктонных сообществ в зонах градиентов солености бассейна Азовского моря // Альгология. 2006. Т. 16. № 2. С. 216–226.
Матишов Г.Г., Матишов Д.Г., Степанян О.В., Аксенов Д.С. Комплексные исследования Азовского, Черного и Каспийского морей на научно-исследовательском судне “Денеб” в 2007 г. // Океанолония. 2009. Т. 49. № 2. С. 313–318.
Попов Ю.И., Ковалишина С.П., Грандова М.А., Матыгин А.С. Многолетние изменения характеристик морских вод и фитопланктона северо-западного шельфа Черного моря // Современные рыбохозяйственные и экологические проблемы Азово-Черноморского региона. 20–23 июня 2012. Керчь, Крым, УДК 551.465. С. 171–178.
Теренько Д.А., Нестерова Л.М. Cyanoprocariota планктона северо-западной части Черного моря (Украина) // Аlgologia. 2015. Т. 25. № 3. С. 278–296.
Bracher A., Bouman H.A., Brewin R.J.W., Bricaud A., Brotas V., Ciotti A.M., Clementson L., Devred E., Di Cicco A., Dutkiewicz S., Hardman-Mountford N.J., Hickman A.E., Hieronymi M., Hirata T., Losa S.N., Mouw C.B., Organelli E., Raitsos D.E., Uitz J., Vogt M., Wolanin A. Obtaining phytoplankton diversity from ocean color: a scientific roadmap for future development // Front. Mar. Sci. 2017. 4. 55. P. 1–15. https://doi.org/10.3389/fmars.2017.00055.
Castenholz R.W., Phylum BX. Cyanobacteria // Bergey’s Manual of Determinative Bacteriology. 2001. V. 2. P. 473–599.
Gitelson A.A., Gao B.-C., Li R.-R., Berdnikov S., Saprygin V. Estimation of chlorophyll-a concentration in productive turbid waters using a Hyperspectral Imager for the Coastal Ocean – the Azov Sea case study // Environ. Res. Lett. 2011. V. 6. P. 1–6.
Gower J.F.R., Doerffer R., Borstad G.A. Interpretation of the 685 nm peak in water-leaving radiance spectra in terms of ̄ uorescence, absorption and scattering, and its observation by M ERIS // Int. J. Remote Sensing. 1999. V. 20 № 9. P. 1771–1786.
Groom S., Sathyendranath S., Ban Y., Bernard S., Brewin R., Brotas V., Brockmann C., Chauhan P., Choi J., Chuprin A., Ciavatta S., Cipollini P., Donlon C., Franz B., He X., Hirata T., Jackson T., Kampel M., Krasemann H., Lavender S., Pardo-Martinez S., Mélin F., Platt T., Santoleri R., Skakala J., Schaeffer B., Smith M., Steinmetz F., Valente A., Wang M. Satellite Ocean Colour: Current Status and Future Perspective // Frontiers in Marine Science. 2019. V. 6. P. 485.
Huot Y., Brown C.A., Cullen J.J. New Algorithm for MODIS Sun-Induced Chlorophyll Fluorescence and a Comparison with Present Data Products // Limnology and Oceanography Methods. 2005. V. 3. P. 108–130.
IOCCG, 2014. Phytoplankton functional types from space. In: Sathyendranath, S. (Ed.), Reports of the International Ocean-Colour Coordinating Group, No. 15, IOCCG, Dartmouth, Canada.
Kubryakov A.A., Lishaev P.N., Aleskerova A.A., Stanichny S.V. Spatio-temporal variability of cyanobacteria blooms in the North-Western shelf of the Black Sea from MODIS and Landsat satellite data // Harmful algae. 2021. (в пeчaти)
Lomakin P.D. Field of the Dissolved Organic Matter Content in the Taganrog Bay (the Sea of Azov) // Physical Oceanography. 2018. V. 25. № 6. P. 459–471. https://doi.org/10.22449/1573-160X-2018-6-4 59-471
Pope R.M., Fry E.S. Absorption spectrum ~380 –700 nm of pure water. II. Integrating cavity measurements // Applied Optics. 1997. V. 36. № 33. P. 8710–8723.
Randolph K., Wilson J., Tedesco L., Li L., Pascual D.L., Soyeux E. Hyperspectral remote sensing of cyanobacteria in turbid productive water using optically active pigments, chlorophyll-a and phycocyanin // Remote Sensing of Environment. 2008. V. 112. P. 4009–4019.
Ruiz-Verdú A., Simis S.G.H., de Hoyos C., Gons H.J., Peña-Martínez R. An evaluation of algorithms for the remote sensing of cyanobacterial biomass // Remote Sensing of Environment. 2008. V. 112 P. 3996–4008.
Schalles J.F., Yacobi Y.Z. Remote detection and seasonal patterns of phycocyanin, carotenoid and chlorophyll pigments in eutrophic waters // Archiv fur Hydrobiologie Special Issues Advances in Limnology. 2000. V. 55. P. 153−168.
Simis S., Peters S., Gons H. Remote sensing of the cyanobacterial pigment phycocyanin in turbid inland water // American Society of Limnology and Oceanography. 2005. V. 50. № 11. P. 237–245.
Stumpf R.P., Davis, T.W., Wynne T.T., Graham J.L., Loftin K.A., Johengen T.H., Gossiaux D., Palladino D., Burtner A. Challenges for mapping cyanotoxin patterns from remote sensing of cyanobacteria // Harmful Algae. 2016. V. 54. P. 160–173.
Sun D., Hu C., Qiu Z., Shi K. Estimating phycocyanin pigment concentration in productive inland waters using Landsat measurements: A case study in Lake Dianchi // Optics Express. 2015. V. 23. № 3. P. 3055.
Uitz, J., Huot, Y., Bruyant, F., Babin, M., Claustre, H. Relating phytoplankton photophysiological properties to community structure on large scales // Limnol. Oceanogr. 53(2), 2008. 614–630.
Wright S.W., Jeffrey S.W., Mantoura R.F.C. Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods. UNESCO Pub. 2005. 667 p.
Wynne T.T., Stumpf R.P., Tomlinson M.C., Warner R.A., Tester P.A., Dyble J., Fahnenstiel G.L. Relating spectral shape to cyanobacterial blooms in the Laurentian Great Lakes // International J. Remote Sensing. 2008. V. 29. № 12. P. 3665–3672.
Wynne T.T., Stumpf R.P., Briggs T.O. Comparing MODIS and MERIS spectral shapes for cyanobacterial bloom detection // International J. Remote Sensing. 2013. P. 1–11.
Wynne T.T., Stumpf R.P. Spatial and Temporal Patterns in the Seasonal Distribution of Toxic Cyanobacteria in Western Lake Erie from 2002–2014 // Toxins. 2015. V. 7. P. 1649–1663. https://doi.org/10.3390/toxins7051649
Дополнительные материалы отсутствуют.
Инструменты
Исследование Земли из Космоса