

Известия РАН. Серия биологическая, 2019, № 1, стр. 52-61
Концепция поливариантности онтогенеза и современная эволюционная морфология
А. А. Нотов a, *, Л. А. Жукова b
a Тверской государственный университет,
Россия, 170100 Тверь, ул. Желябова, 33
b Марийский государственный университет,
Россия, 424000 Йошкар-Ола, пл. Ленина, 1
* E-mail: anotov@mail.ru
Поступила в редакцию 07.05.2018
После доработки 31.07.2018
Принята к публикации 31.07.2018
Аннотация
Рассмотрена возможность использования концепции поливариантности онтогенеза в эволюционной морфологии. Отмечено, что концепция ориентирована на выявление всех вариантов, сопряженных с разными аспектами организации живых существ. Установлено, что в сочетании с анализом корреляционной структуры признаков и методами популяционной мерономии предложенный подход может способствовать выяснению модусов эволюции структурного разнообразия.
Интерес к морфологии способствовал прогрессивному развитию эволюционной биологии в ХХ в. (Серебрякова, 1972, 1983; Meyen, 1973, 1984; Воробьева, Мейен, 1988; Воробьева, 1991; Мамкаев, 1991, 1996 и др.). Однако активный рост молекулярной филогенетики на рубеже ХХ–ХХI вв. привел к падению престижа эволюционной морфологии (Тимонин, 2001 и др.). В настоящее время ситуация стала критической (Расницын, 2014; Ivanova-Kazas, 2016; Notov, 2016 и др.). Тем не менее некоторые работы все же позволяют надеяться на возобновление внимания к изучению структурной эволюции (Тимонин, 2011; Mamkaev, 2011; Пожидаев, 2015; Kosevich, 2015; Rasnitsyn et al., 2015; Notov, 2016, 2017; Rutishauser, 2016; Кузнецова, Тимонин, 2017; Матюхин, 2017; Нуралиев и др., 2017; Pardo et al., 2017; Simpson et al., 2017; Harrison, Morriss, 2018 и др.). В этой связи актуален анализ концепций, которые могли бы способствовать укреплению позиций эволюционной морфологии. На наш взгляд, заслуживает внимания концепция поливариантности (ПВ) онтогенеза, или в более общем виде ПВ развития (Жукова, 1986, 1995, 2008; Поливариантность…, 2006; Нотов, Жукова, 2013).
Возникновение представлений о ПВ индивидуального развития организма (Сабинин, 1963; Воронцова, Заугольнова, 1978; Жукова, 1986 и др.) было связано в большей степени с изучением онтогенеза растений. В зоологии они появились позднее (Макаров, 1991; Оленев, Григоркина, 1998; Оленев, 2002 и др.). Концепция ПВ сформировалась как подход к анализу гетерогенности популяций (Жукова, Комаров, 1990; Жукова, 1995). Сейчас она приобретает статус общебиологической парадигмы, которая рассматривает ПВ как универсальное свойство живых объектов разного уровня организации (Поливариантность…, 2006; Нотов, Жукова, 2013, 2016).
Цель работы – оценка возможности использования концепции ПВ в эволюционных исследованиях, анализ ее методических основ и связей с другими подходами.
ОБЩАЯ ХАРАКТЕРИСТИКА КОНЦЕПЦИИ ПВ
В рамках концепции ПВ онтогенез понимается как весь процесс индивидуального развития – от зиготы или иного зачатка (диаспоры) до естественной смерти на завершающих этапах вследствие старения (Жукова, 1995). Принята универсальная схема периодизации онтогенеза с высоким уровнем детализации (Уранов, 1975; Жукова, 1995; Жукова, Зубкова, 2016 и др.). Для семенных растений отмечают, как правило, 4–5 периодов и 11–12 онтогенетических состояний (Жукова, 1995; Жукова, Зубкова, 2016).
Концепция ориентирована на выявление всех возможных различий между особями в популяции. Они рассматриваются как результат проявления ПВ онтогенеза. Учтена специфика онтогенеза растений. В связи с активным морфогенезом в течение всей жизни его пластичность достигает высокого уровня. Различные пути (траектории) развития весьма многообразны (Жукова, 1995; Нотов, 1999). Обычны варианты с “выпадением” отдельных онтогенетических состояний, может нарушаться их последовательность, варьирует степень полноты онтогенеза. Нередко в ходе онтогенеза изменяется жизненная форма, утрачивается целостность организма, происходит его разделение на части (партикуляция) с образованием дочерних особей (Жукова, 1995, 2008 и др.).
Систематизация вариантов проводится в соответствии с общими принципами системного анализа организма и онтогенеза (Жукова, 1995, 2008; Нотов, Жукова, 2013 и др.). При классификации учитываются связь с универсальными аспектами организации, отношение к определенному структурному уровню, масштаб индивидуальных различий хода онтогенеза, характер взаимосвязей с другими вариантами. С основными аспектами организации сопряжены соответствующие надтипы ПВ – структурная, функциональная, динамическая и путей онтогенеза. В качестве надтипов выделены ПВ размножения и циклов воспроизведения. Специфика внешних связей объекта рассмотрена в пределах надтипа экологическая ПВ. Связь различий с определенным структурным уровнем (молекулярный, клеточный, организменный) отражена в названии некоторых типов. Среди них, например, биохимическая, анатомическая и морфологическая ПВ. В пределах морфологического типа организменному уровню соответствует ПВ жизненных форм (Поливариантность…, 2006; Жукова, 2008 и др.). Масштабами изменения хода онтогенеза отличаются ПВ темпов индивидуального развития и ПВ путей онтогенеза. В первом случае варьирует динамика формирования определенных онтогенетических состояний, в последнем существенно трансформируется типичный ход онтогенеза. Варианты могут быть независимыми, взаимосвязанными или иерархически соподчиненными, что существенно осложняет создание классификации (Нотов, Жукова, 2016). Характер связей выявляется при анализе спектра ПВ конкретного объекта. В одной из последних версий классификации ПВ онтогенеза выделены 7 надтипов и 11 типов (Нотов, Жукова, 2013, 2016).
Предложенный подход отражает специфику организации и индивидуального развития не только растений, но и других модульных живых существ (Нотов, 1999; Notov, 2015, 2016). Он может быть применен также и к унитарным организмам. Начата разработка общей классификации типов ПВ (Нотов, Жукова, 2013, 2016).
Таким образом, концепция дает целостное представление о многообразии форм ПВ, ориентирована на детальный анализ всех возможных проявлений вариабельности онтогенеза. При этом учитываются варианты различного масштаба и степени сложности, сопряженные с разными аспектами организации и структурными уровнями. Концепция используется в качестве одного из способов оценки гетерогенности популяций (Поливариантность…, 2006). Актуально сопоставление его с другими подходами к изучению вариабельности и разнообразия.
ДРУГИЕ ПОДХОДЫ К АНАЛИЗУ ВАРИАБЕЛЬНОСТИ
В рамках нашей задачи заслуживают внимания направления, связанные с изучением вариабельности индивидуального развития и эволюционной проблематикой. Особый интерес представляют подходы, которые можно использовать для выяснения модусов эволюции структурного разнообразия.
Основной метод эволюционной морфологии сравнительно-морфологический. В рамках конструктивно-морфологических исследований в 1970–1980-е гг. сформировался мерономический подход (Мейен, 1978; Воробьева, Мейен, 1988 и др.). Он был ориентирован на анализ номотетического аспекта морфологической эволюции (Meyen, 1973; Мейен, 1990 и др.). Развитие этого направления позволило переосмыслить широкий круг мировоззренческих и методологических проблем (Чайковский, 1990; Чебанов, 2017). Был сделан фронтальный анализ обширного материала по современным и ископаемым организмам (Meyen, 1973, 1984; Мейен, 1987 и др.). Он помог выявить нетривиальные модусы морфологической эволюции, феномен транзитивного полиморфизма (Мейен, 1987, 1988 и др.). В рамках конструктивно-морфологического подхода были выяснены общие модусы структурных преобразований, разработаны представления о ступенях эволюции морфофункциональных систем, метод морфологических спектров, принцип исходного архетипического многообразия (Мамкаев, 1991, 1996, 2004; Воробьева, 2006; Mamkaev, 2011 и др.). В связи с активным развитием evo-devo в центре внимания сейчас оказались механизмы трансформации и сопряженные с ними пути преобразования онтогенеза (Воробьева, 2006, 2007, 2010; Шишкин, 2010; Озернюк, Исаева, 2016 и др.).
В рамках традиционных для синтетитческой теории эволюции исследований изменчивости и полиморфизма популяций сформировалась фенетика (Яблоков, 1980 и др.). Групповой анализ изменчивости был направлен на выяснение специфики микроэволюционных процессов. Изучали также закономерности становления гомологичных признаков в онтогенезе (Воробьева, Медведева, 1980). В настоящее время вызывает интерес связь с макроэволюционной проблематикой. Активно развивается популяционная мерономия (Васильев, 2005; Васильев, Васильева, 2009 и др.). Она синтезировала представления эпигенетической теории эволюции, номогенеза (в понимании Мейена) и эволюционной экологии. Стали проводить анализ эпигенетической дивергенции таксонов разного ранга, филогенетических связей и исследования транзитивного полиморфизма по большому числу структур. Изучение эпигенетических аспектов транзитивного полиморфизма позволило соотнести механизмы микро- и макроэволюционных трансформаций морфогенеза (Васильев, Васильева, 2009). С позиции эволюционной морфологии более значимы предлагаемые способы выявления гомологии, эволюционной роли параллелизмов и морфологических новаций.
Актуально сопоставление разных подходов. Особое значение имеет выяснение степени сходства их целевых установок и методологической специфики. Такой анализ позволит оценить возможность синтеза подходов.
О ВОЗМОЖНОСТИ СИНТЕЗА ПОДХОДОВ
Отмечены три подхода к анализу разнообразия и вариабельности: концепция ПВ онтогенеза, популяционная мерономия, конструктивно-морфологический подход. Из них пока лишь последний нашел широкое применение в эволюционной морфологии (Воробьева, Мейен, 1988; Мамкаев, 1991 и др.). При сравнении подходов (табл. 1) он рассмотрен в варианте, который используется в эволюционных исследованиях – в контексте мерономии Мейена (Meyen, 1973; Мейен, 1978 и др.), в сочетании с концепцией эволюции онтогенеза (Шмальгаузен, 1982; Шишкин, 1987, 2010 и др.) и морфофункциональным анализом (Воробьева, 2007). В рамках всех трех подходов реализуется системная методология (Воробьева, 2006; Васильев, Васильева, 2009; Нотов, Жукова, 2013, 2016 и др.).
Таблица 1.
Сравнительная характеристика подходов к анализу вариабельности
| Характеристика | Концепция ПВ онтогенеза | Популяционная мерономия | Конструктивно-морфологический подход |
|---|---|---|---|
| Объекты | Организм, популяция, вид | Популяция, вид, таксоны надвидового ранга |
Виды, таксоны надвидового ранга, их архетипы |
| Предмет исследования |
Вариабельность онтогенеза, разнообразие его вариантов |
Вариабельность фенов и фенокомпозиций, разнообразие фенотипов | Вариабельность меронов, архетипов, разнообразие рефренов |
| Задачи | Выявление и анализ полного спектра вариантов онтогенеза, которые сопряжены со всеми аспектами организации биосистем |
Выяснение закономерностей гомологической изменчивости морфологических структур | Комплексный анализ
структурного разнообразия, выяснение закономерностей и модусов его ЭВ-преобразования |
| Связь с ПВ онтогенеза |
Вариабельность онтогенеза – ключевой аспект анализа; ПВ – главная идея и основное содержание концепции |
Изучение изменчивости как одного из результатов вариабельности развития косвенно связано с анализом ПВ онтогенеза; выявляется популяционная специфика программ развития | Среди компонентов методологической базы концепция эволюции онтогенеза, теория филэмбриогенеза; представления о многообразии креодов согласуются с идеей ПВ онтогенеза |
| Система корреляций | Не анализируются | Обязательный элемент исследований | Ключевой объект морфофункционального и исторического анализа |
| МИЭВ-проблематика | Не рассматривается, но результаты значимы для оценки мобилизационного резерва изменчивости |
Основной метод анализа МИЭВ-процессов | Интересен как компонент общей методологии типологического анализа |
| МАЭВ-проблематика | Не рассматривается, но результаты могут представлять интерес для ее анализа |
Позволяет выявлять связи МИЭВ- и МАЭВ-процессов, выяснять филогенетические отношения | Методическая основа выявления общих закономерностей МАЭВ |
| Значение для ЭВ-морфологии |
В сочетании с анализом корреляций и архетипов таксонов может способствовать выявлению модусов морфологической ЭВ | Помогает оценивать ЭВ-роль параллелизмов и морфологических новаций, решать проблемы гомологизации | Основной подход к анализу результатов и модусов морфологической ЭВ, ее механизмов и закономерностей |
Специфика определяется прежде всего особенностями объектов, предметов и задач исследований, характером связей с ПВ онтогенеза, микро- и макроэволюционной проблематикой и эволюционной морфологией (табл. 1). В отличие от других подходов в концепции ПВ рассматривается не только вариабельность структур, но и многообразие вариантов, связанных со всеми аспектами изучения организмов (табл. 1). Полный спектр вариантов создает базу для взаимодействия с генетикой и биологией развития. Такой синтез даст возможность выявлять характер связи каждого варианта с процессами развития. Результаты генетиков подтвердили значительное разнообразие механизмов, лежащих в основе любого типа изменчивости (Тиходеев, 2012, 2013). Особенно неоднородны варианты онтогенетической изменчивости (Тиходеев, 2013). Она не связана жестко ни с конкретной формой наследования, ни с определенными молекулярными механизмами, может сочетаться с разными элементами наследственной и ненаследственной изменчивости.
Основой для выявления механизмов эволюционных преобразований стала концепция эволюции онтогенеза (Шмальгаузен, 1982; Шишкин, 1987, 2010; Озернюк, Исаева, 2016 и др.). Общие представления о популяции, онтогенезе и многообразии его вариантов согласуются с базовыми положениями концепции ПВ и популяционной мерономии. В эволюционных исследованиях популяция исходно рассматривалась как саморегулирующаяся полиморфная система (Шмальгаузен, 1968). Предполагалось, что “надындивидуальные биологические системы в пределах вида” имеют определенные уровни “генетического и модификационного полиморфизма, … соотношений между разными полами и между возрастными формами, длительности жизни и ее этапов, плодовитости и скорости смены поколений и многие другие черты организации, характеризующие эволюционную пластичность” (Шмальгаузен, 1968, с. 174). Фактически в неявном виде наряду со структурной была отмечена ПВ способов размножения и воспроизведения, темпов онтогенеза, но без использования терминологии концепции ПВ. Обращалось также внимание на то, что модификации могут проявляться “в течение всего онтогенеза каждой особи, начиная от зиготы и до конца ее жизни” (Шмальгаузен, 1968, с. 33). Сходство ключевых положений определяет возможность взаимодействия подходов. Представления о механизмах вариабельности (ПВ) онтогенеза и ее эволюционной роли могут способствовать развитию теоретической базы каждого подхода. Пока они в должной степени сформированы только в рамках концепции эволюции онтогенеза и учитываются при выявлении модусов морфологической эволюции (табл. 1).
Типологические исследования на основе конструктивно-морфологического подхода характеризуются наиболее высоким уровнем абстрагирования. Их объекты – мероны, архетипы; выясняется мерономическое разнообразие таксонов высокого ранга (табл. 1). Ключевое значение имеет анализ вариабельности этих объектов, фактически рассматриваются спектры ПВ в пределах крупной систематической группы. При изучении морфологической эволюции подчеркивается значимость представлений о возможных путях трансформации морфогенеза и онтогенеза (Воробьева, Мейен, 1988; Тимонин, 2011 и др.). С точки зрения взаимодействия конструктивно-морфологического направления с другими подходами интересны получаемые при их использовании спектры изменчивости структур и вариантов путей онтогенеза. Такие спектры облегчают конструирование архетипа, оценку изменчивости радикала таксона, уровня и характера вариабельности меронов. Направленная изменчивость архетипических признаков – основа для дивергенции таксонов (Любарский, 1996).
Возможности каждого из трех подходов существенно расширяет изучение корреляционных связей. Однако в рамках концепции ПВ они пока не рассматриваются (табл. 1). Анализ системы корреляций достаточно полно раскрывает специфику взаимоотношений между разными характеристиками и вариантами. Он создает основу для конструирования архетипа (Любарский, 1996). Данные о корреляциях помогают соотносить устойчивость и изменчивость, рассматривать проблему целостности (Ростова, 2002).
Концепция ПВ открыта для взаимодействия с другими подходами. Оно позволит в определенной степени расширить область ее применения. Полные спектры вариантов могут быть полезны для оценки мобилизационного резерва изменчивости (табл. 1), выявления архетипов, примеров транзитивного полиморфизма. Решение этих задач значимо в контексте микро- и макроэволюционной проблематики.
СПЕКТРЫ ПВ И ЭВОЛЮЦИОННАЯ МОРФОЛОГИЯ
“Создание целостной теории онтогенеза” – одна из ключевых задач развития эволюционной морфологии “на ее пути к новому синтезу” (Воробьева, 1991, с. 256). Каким же образом можно реализовать потенциал междисциплинарных связей концепции ПВ?
При выяснении модусов морфологической эволюции в равной степени интересны проявления устойчивости и вариабельности. Применение конструкционно-морфологического подхода предполагает выявление архетипов таксонов высокого ранга и выполнение масштабного мерономического анализа. При использовании концепции ПВ необходимого для этого уровня абстрактности можно достигнуть путем упорядочивания индивидуальных спектров ПВ с учетом специфики корреляционной системы и получения интегрального спектра с помощью их поэтапного обобщения.
Для разработки методических основ учета данных о спектрах ПВ в эволюционной морфологии актуален специальный анализ крупных таксонов со значительным структурным многообразием. Особенно интересны группы, в которых выявлены модусы эволюционных преобразований, изучена система корреляций, оценена гетерогенность популяций с позиции концепции ПВ. В этом случае привлечение данных о разных типах ПВ облегчает уточнение и детализацию обнаруженных модусов, выяснение механизмов структурных преобразований.
К числу таких модельных таксонов можно отнести, например, злаки (Poaceae) и подтрибу Alchemillinae Rothm. (Rosaceae). Каждая из этих групп была объектом комплексного анализа (Серебрякова, 1968, 1971, 1974; Жукова, 1986, 1995; Notov, Kusnetzova, 2004; Курченко, 2010 и др.). Они интересны с точки зрения структуры корреляционных систем.
Многие структурные и ритмологические особенности злаков коррелируют со строением побега (Серебрякова, 1968, 1971, 1974). Оно определяет специфику структуры и ритма роста на всех этапах морфогенеза побега и сопряжено с основными программами формирования системы побегов, включая общий алгоритм ее развития в онтогенезе (табл. 2). Возникновение розеткообразующих форм обусловлено особенностями экологической дифференциации видов (Серебрякова, 1968, 1971). Все это позволило предложить оригинальную систему жизненных форм злаков, которая базируется на представлениях об их биоморфологической эволюции. В качестве основополагающего признака выбран тип побега, а не традиционное разделение на древесные и травянистые формы (Серебрякова, 1971). Соотнесение данных о корреляционных связях и изменчивости признаков помогло выявить модусы структурной эволюции и общие тенденции (Серебрякова, 1968, 1971, 1974).
Таблица 2.
Спектр поливариантности (ПВ) и характер корреляции признаков у злаков
| Характеристики | Тип побега (ПБ) | Типы ПВ |
|
|---|---|---|---|
| безрозеточный | розеткообразующий | ||
| Число чешуевидных листьев | Много | Мало | 1 |
| Емкость почки | Больше | Меньше | 1, 2 |
| Продолжительность развития ПБ | Моноциклические | Ди-, полициклические | 1, 4, 8 |
| Дифференциация ПБ | Однотипные | Разнотипнотипные | 1, 3, 4, 6, 8 |
| Ветвление ПБ | Нередко рассеянное | Концентрированное, кущение | 1, 6 |
| Побегообразование | Экстравагинальное | Экстра- и интравагинальное | 1, 4 |
| Внутренний ритм развития ПБ | Крупноквантовость | Мелкоквантовость | 8, 9 |
| Развертывание почек возобновления | Однократное | Пролонгированное, многократное | 4, 8 |
| Ритм кущения | Постгенеративное | Прегенеративное | 1, 4, 6, 8 |
| Темпы онтогенеза | Медленный, с постепенным усилением ПБ | Нередко быстрый, с цветущим первичным ПБ |
5, 6, 8, 11 |
| Жизненные формы | Многолетние травы, однолетники, древесные, кустарниковые | Многолетние травы, однолетники | 1, 3–6, 8, 10, 11 |
| Экотопы | Чаще лесные | Луговые, открытые биотопы | 1, 4, 6, 8, 10, 11 |
Примечание. Надтипы и типы ПВ соответствуют опубликованным ранее (Нотов, Жукова, 2013, с изменениями): структурная – морфологическая (1), анатомическая (2), размерная (3); динамическая – феноритмологическая (4), темпов онтогенеза (5); репродукции – способов размножения (6); воспроизведения – жизненных циклов (7); функциональная – физиологическая (8), биохимическая (9); экологическая – экологических позиций (10); путей онтогенеза – полноты и типа онтогенеза (11). Характер корреляции признаков соответствует приведенному ранее (Серебрякова, 1968, 1971).
Не менее значимо строение побега для представителей подтрибы Alchemillinae. С его структурой коррелируют основные особенности листа и почки (табл. 3). С преобразованием типов побегов была сопряжена эволюция архитектурных моделей и жизненных форм (Notov, Kusnetzova, 2004).
Таблица 3.
Спектр поливариантности (ПВ) и характер корреляции признаков в подтрибе Alchemillinae
| Характеристики | Тип побега (ПБ) | Типы ПВ |
|||
|---|---|---|---|---|---|
| безрозеточный | розеточный | ||||
| Продолжительность развития ПБ | Моноциклические | Полициклические | 1, 3, 8 | ||
| Дифференциация ПБ | Однотипные | Разнотипные | 1, 3, 4, 6, 8, 9 | ||
| Черешок и пластинка листа (ПЛ) |
Черешок короткий, основание ПЛ усеченное или клиновидное | Черешок длинный, основание ПЛ сердцевидное |
1, 8 | ||
| Прилистники | Чаще в незначительной степени приросшие к черешку | Почти полностью приросшие к черешку | 1 | ||
| Влагалище | Замкнутое | Часто окрытое, реже в разной степени замкнутое | 1 | ||
| Листовые примордии в почке | Зачатки прилистников охватывают ПЛ | Зачатки прилистников не охватывают ПЛ | 1, 2, 8 | ||
| Ритм развития листового примордия |  |
Есть фаза опережающего роста зачатков прилистников | 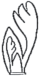 |
Нет фазы опережающего роста зачатков прилистников | 1, 2, 8 |
| Жизненные формы | Кустарники, полукустарники, однолетники | Многолетние травы, реже кустарнички |
1, 3, 4, 5, 6, 7, 8, 10, 11 | ||
Примечание. Обозначения типов ПВ такие же, как в табл. 2. Характер корреляции признаков соответствует описанному ранее (Notov, Kusnetzova, 2004).
В результате поэтапного обобщения данных о ПВ представителей этих систематических групп (Жукова, Комаров, 1990; Жукова, 1995, 2008; Notov, Kusnetzova, 2004; Нотов, Андреева, 2013; Жукова и др., 2015 и др.) и материалов специальных исследований нами получены интегральные спектры ПВ. Они соотнесены с информацией о корреляции признаков (табл. 2, 3). В рамках этой статьи оказалось возможным представить сведения только по выявленным типам ПВ без детализации по вариантам. Анализ спектров ПВ позволил оценить степень целостности корреляционной системы. Появилась возможность определить специфику связей между устойчивостью и вариабельностью структур и процессов, которые сопряжены с различными аспектами организации и проявляются в соответствующих типах ПВ. Для каждого признака корреляционной плеяды установлена связь с большим или меньшим числом других признаков и определенных типов ПВ (табл. 2, 3). Сложные признаки и характеристики сочетались практически со всеми типами ПВ. Например, дифференциация побегов сопряжена с проявлением комплекса взаимосвязанных признаков, затрагивающих разные аспекты организации. С этими же аспектами связаны и выявляемые типы ПВ (морфологическая, ритмологическая, способов размножения, физиологическая и др.) (табл. 2, 3).
Полученные результаты интересны с точки зрения возможных направлений взаимодействия разных подходов. Сочетание корреляционного анализа с оценкой спектров ПВ дает возможность отразить многомерность и соподчиненность структурно-функциональных связей, включая пути онтогенеза. Этот способ может стать средством детализации представлений о мобилизационном резерве изменчивости. Комплексные исследования ПВ с использованием методов популяционной мерономии могут усилить прогностическую ценность каждого подхода. Статистический анализ данных позволяет получить частотные характеристики спектров ПВ.
Для эволюционной морфологии специальный интерес могут представлять данные о ПВ биоморф. Жизненная форма координирует максимальное число различных связей (табл. 2, 3). Ее вариабельность можно рассматривать в качестве самостоятельного типа ПВ (Поливариантность…, 2006; Жукова, 2008 и др.). С биоморфологическими особенностями сопряжены многие другие типы ПВ. Однако такие наиболее значимые с точки зрения разных траекторий онтогенеза типы, как ПВ жизненных циклов, путей онтогенеза, экологических позиций, главным образом связаны именно со спецификой жизненной формы. Хотя ПВ жизненных циклов наиболее полно проявляется у низших растений, ее можно обнаружить и у семенных растений. Благодаря встречаемости регулярного апомиксиса в разных группах реализуются различные варианты сокращенных жизненных циклов – примеры этого типа ПВ. Такая ПВ выявлена у представителей секции Brevicaulon Rothm. рода Alchemilla L. (Глазунова, 2000). При этом все виды секции совершенно идентичны с точки зрения архитектурной модели и жизненной формы, имеют однотипное строение системы побегов, листа и почки (Notov, Kusnetzova, 2004). В связи с регулярным апомиксисом у них сформировались общие особенности в корреляционной структуре и изменчивости признаков цветка (Глазунова, Мятлев, 1990).
Биоморфологические характеристики в большей степени коррелируют с наиболее масштабными перестройками онтогенеза и общей программы развития. Данные об их вариабельности особенно важны при изучении модусов трансформации жизненных форм и архитектурных моделей (Серебрякова, 1971, 1972 и др.). С позиций эволюционной морфологии актуален специальный анализ у злаков и представителей Alchemillinae биоморф, рассматриваемых в рамках демографической классификации. Уже собраны разноплановые данные о ПВ этих групп жизненных форм. При преобладании у злаков с розеткообразующими побегами моноцентрических форм полицентрические варианты могут быть отмечены даже у плотнодерновинных злаков (Жукова, 1995, 2008 и др.). Выявление модусов трансформации данных типов биоморф позволит связать структурную эволюцию с формированием разных демографических стратегий.
Рассмотренные примеры свидетельствуют о возможности использования данных о спектрах ПВ в эволюционной морфологии и целесообразности взаимодействия концепции ПВ с другими подходами. Материалы о ПВ помогают оценивать уровень пластичности структурно-функциональной организации. Они дополняют характеристику мобилизационного резерва изменчивости. Соотнесение спектров ПВ и особенностей корреляционных систем актуально при выявлении архетипов таксонов и модусов преобразования структур. Информация о ПВ имеет большое значение при исследовании морфологической эволюции модульных организмов, обладающих открытым ростом и высокой пластичностью онтогенеза. Она позволит расширить возможности анализа эволюции жизненных форм и архитектурных моделей, так как их морфологическая специфика сопряжена с общими программами онтогенеза. Изучение структурного разнообразия с позиций концепции ПВ поможет детализировать принцип исходного морфофункционального архетипического многообразия (Мамкаев, 2004) на примере ботанических объектов и выявлять механизмы, связывающие онто- и филогенезы.
ЗАКЛЮЧЕНИЕ
Таким образом, актуально взаимодействие концепции ПВ онтогенеза с другими подходами к оценке вариабельности и структурного разнообразия. Оно позволит объединить усилия разных специалистов и станет основой для междисциплинарного синтеза знаний, который расширит возможности каждого подхода. Анализ интегральных спектров ПВ в сочетании с данными о структуре корреляционных систем, методами типологии и популяционной мерономии может облегчить выяснение модусов морфологической эволюции. Такие комплексные исследования будут способствовать использованию концепции ПВ в эволюционной биологии. Они важны и с точки зрения укрепления позиций современной эволюционной морфологии.
Список литературы
Васильев А.Г. Эпигенетические основы фенетики: на пути к популяционной мерономии. Екатеринбург: Академкнига, 2005. 639 с.
Васильев А.Г., Васильева И.А. Гомологическая изменчивость морфологических структур и эпигенетическая дивергенция таксонов: основы популяционной мерономии. М.: КМК, 2009. 511 с.
Воробьева Э.И. Эволюционный синтез и эволюционная морфология // Современная эволюционная морфология. Киев: Наук. думка, 1991. С. 244–261.
Воробьева Э.И. Морфологическая эволюция: принципы оценки, закономерности, механизмы // Палеонтол. журн. 2006. № 6. С. 18–33.
Воробьева Э.И. Морфофункциональный подход в палеонтологии // Палеонтол. журн. 2007. № 4. С. 3–15.
Воробьева Э.И. Современная эволюционная биология развития: механический и молекулярно-генетический или фенотипический подходы? // Онтогенез. 2010. Т. 41. № 5. С. 332–339.
Воробьева Э.И., Медведева И.М. К вопросу об эволюции онтогенеза и роли изменчивости // Внутривидовая изменчивость в онтогенезе животных. М.: Наука, 1980. С. 5–18.
Воробьева Э.И., Мейен С.В. Морфологические исследования в палеонтологии // Соврем. палеонтология. Т. 1. М.: Недра, 1988. С. 80–123.
Воронцова Л.И., Заугольнова Л.Б. Мультивариантность развития особей в течение онтогенеза и ее значение в регуляции численности и состава ценопопуляций растений // Журн. общ. биологии. 1978. Т. 39. № 4. С. 555–562.
Глазунова К.П. Род Alchemilla L. (Rosaceae) – классический объект для изучения факультативного апомиксиса // Эмбриология цветковых растений: терминология и концепции. Т. 3. СПб.: Мир и семья, 2000. С. 206–214.
Глазунова К.П., Мятлев В.Д. Корреляционная структура и изменчивость признаков при регулярном апомиксисе (на примере агамного вида манжетка горная) // Бюл. МОИП. Отд. биол. 1990. Т. 95. № 6. С. 96–110.
Жукова Л.А. Поливариантность онтогенезов луговых растений // Жизненные формы в экологии и систематике растений. М.: Изд-во МГПИ, 1986. С. 104–114.
Жукова Л.А. Популяционная жизнь луговых растений. М.: Ланар, 1995. 224 с.
Жукова Л.А. Поливариантность развития организмов в разных царствах биосферы // Современные подходы к описанию структуры растения. Киров: Лобань, 2008. Разд. 6.3. С. 240–260.
Жукова Л.А., Зубкова Е.В. Демографический подход, принципы выделения онтогенетических состояний и жизненности, поливариантность развития растений // Вестн. ТвГУ. Сер. Биология и экология. 2016. № 4. С. 169–183.
Жукова Л.А., Комаров А.С. Поливариантность онтогенеза и динамика ценопопуляций растений // Журн. общ. биологии. 1990. Т. 51. № 4. С. 450–461.
Жукова Л.А., Ведерникова О.П., Быченко Т.М., Османова Г.О. Лекарственные растения. Разнообразие жизненных форм. Йошкар-Ола: Изд-во МарГУ, 2015. 167 с.
Кузнецова Т.В., Тимонин А.К. Соцветие: морфология, эволюция, таксономическое значение (применение комплементарных подходов). М.: КМК, 2017. 183 с.
Курченко Е.И. Род полевица (Agrostis L., сем. Poaceae) России и сопредельных стран: Морфология, систематика и эволюционные отношения. М.: Прометей, 2010. 516 с.
Любарский Г.Ю. Архетип, стиль и ранг в биологической систематике. М.: КМК Scientific Press, 1996. 432 с.
Макаров К.В. Поливариантность жизненного цикла жужелиц (Coleoptera, Carabidae) // Проблемы почвенной зоологии. Новосибирск: Зап.-Сиб. лесоустроительное предприятие, 1991. С. 132.
Мамкаев Ю.В. Методы и закономерности эволюционной морфологии // Современная эволюционная морфология. Киев: Наук. думка, 1991. С. 33–55.
Мамкаев Ю.В. Морфологические принципы систематизациии биоразнообразия // Журн. общ. биологии. 1996. Т. 57. № 2. С. 40–51.
Мамкаев Ю.В. Дарвинизм и номогенез // Фундаментальные зоологические исследования. M.; СПб.: КМК, 2004. С. 114–143.
Матюхин Д.Л. Псевдоциклы и псевдоциклическое сходство побеговых систем у хвойных // Журн. общ. биологии. 2017. Т. 78. № 3. С. 43–53.
Мейен С.В. Основные аспекты типологии организмов // Журн. общ. биологии. 1978. Т. 39. № 4. С. 495–508.
Мейен С.В. Основы палеоботаники. М.: Недра, 1987. 404 с.
Мейен С.В. Нетривиальные модусы морфологической эволюции высших растений // Современные проблемы эволюционной морфологии. М.: Наука, 1988. С. 91–103.
Мейен С.В. Нетривиальная биология (заметки о …) // Журн. общ. биологии. 1990. Т. 51. № 1. С. 4–14.
Нотов А.А. О специфике функциональной организации и индивидуального развития модульных объектов // Журн. общ. биологии. 1999. Т. 60. № 1. С. 60–79.
Нотов А.А., Андреева Е.А. Аномалии генеративных структур у моноподиально-розеточных розоцветных (Rosaceae: Rosoideae). Тверь: Изд-во ТвГУ, 2013. 198 с.
Нотов А.А., Жукова Л.А. О роли популяционно-онтогенетического подхода в развитии современной биологии и экологии // Вестн. ТвГУ. Сер. Биология и экология. 2013. Вып. 32. № 31. С. 293–330.
Нотов А.А., Жукова Л.А. Поливариантность развития биосистем: основные задачи и направления исследований // Современные концепции экологии биосистем и их роль в решении проблем сохранения природы и природопользования. Пенза: Изд-во ПГУ, 2016. С. 148–150.
Нуралиев М.С., Соколов Д.Д., Оскольский А.А. Эволюционная морфология цветка Araliaceae (на примере азиатских представителей рода Schefflera). М.: МАКС Пресс, 2017. 456 с.
Озернюк Н.Д., Исаева В.В. Эволюция онтогенеза. М.: КМК, 2016. 407 с.
Оленев Г.В. Альтернативные типы онтогенеза цикломорфных грызунов и их роль в популяционной динамике (экологический анализ) // Экология. 2002. № 5. С. 341–350.
Оленев Г.В., Григоркина Е.Б. Функциональная структурированность популяций мелких млекопитающих (радиобиологический аспект) // Экология. 1998. № 6. С. 447–451.
Пожидаев А.Е. Рефренная структура биологического многообразия и теория филогенеза // Палеоботан. временник. 2015. № 2. С. 115–127.
Поливариантность развития организмов, популяций и сообществ. Йошкар-Ола: Изд-во МарГУ, 2006. 326 с.
Расницын А.П. Эволюционная теория: современный этап // Палеонтол. журн. 2014. № 1. С. 3–8.
Ростова Н.С. Корреляции: структура и изменчивость. СПб.: Изд-во СПбГУ, 2002. 308 с.
Сабинин Д.А. Физиология развития растений. М.: Изд-во АН СССР, 1963. 196 с.
Серебрякова Т.И. Побегообразование и жизненные формы некоторых овсяниц (Festuca L.) в связи с их эволюцией // Вопросы морфогенеза цветковых растений и строение их популяций. М.: Наука, 1968. С. 7–51.
Серебрякова Т.И. Морфогенез побегов и эволюция жизненных форм злаков. М.: Наука, 1971. 359 с.
Серебрякова Т.И. Учение о жизненных формах растений на современном этапе // Итоги науки и техники. Ботаника. Т. 1. М.: Изд-во ВИНИТИ, 1972. С. 84–169.
Серебрякова Т.И. Эволюционные отношения жизненных форм в некоторых секциях рода Poa L. // Проблемы филогении высших растений. М.: Наука, 1974. С. 116–152.
Серебрякова Т.И. О некоторых модусах морфологической эволюции цветковых растений // Журн. общ. биологии. 1983. Т. 44. № 5. С. 579–593.
Тимонин А.К. Роль морфологии в ботанике // Гомологии в ботанике: опыт и рефлексия. СПб.: СПб-союз ученых, 2001. С. 10–17.
Тимонин А.К. Аномальное вторичное утолщение центросеменных: специфика морфофункциональной эволюции растений. М.: КМК, 2011. 355 с.
Тиходеев О.Н. Кризис традиционных представлений об изменчивости: на пути к новой парадигме // Экол. генетика. 2012. Т. 10. № 4. С. 56–65.
Тиходеев О.Н. Классификация изменчивости по факторам, определяющим фенотип: традиционные взгляды и их современная ревизия // Экол. генетика. 2013. Т. 11. № 3. С. 79–92.
Уранов А.А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов // Биол. науки. 1975. № 2. С. 7–34.
Чайковский Ю.В. Элементы эволюционной диатропики. М.: Наука, 1990. 272 с.
Чебанов С.В. Мерономия С.В. Мейена: К 40-летию формулирования // Lethaea rossica. Рос. палеоботан. журн. 2017. Т. 14. С. 64–92.
Шишкин М.А. Индивидуальное развитие и эволюционная теория // Эволюция и биоценотические кризисы. М.: Наука, 1987. С. 76–124.
Шишкин М.А. Эволюционная теория и научное мышление // Палеонтол. журн. 2010. № 6. С. 3–17.
Шмальгаузен И.И. Кибернетические вопросы биологии. Новосибирск: Наука, 1968. 225 с.
Шмальгаузен И.И. Организм как целое в индивидуальном и историческом развитии. М.: Наука, 1982. 383 с.
Яблоков А.В. Фенетика. Эволюция, популяция, признак. М.: Наука, 1980. 135 с.
Harrison C.J., Morris J.L. The origin and early evolution of vascular plant shoots and leaves // Philos. Trans. Roy. Soc. B: Biol. Sci. 2018. V. 373(1739). Art. 20160496.
Ivanova-Kazas O.M. Molecules, morphology, and phylogeny // Paleontol. J. 2016. V. 50. № 13. P. 1474–1476.
Kosevich I.A. Basis of ontogenetic and evolutionary transformations of thecate hydroids // Paleontol. J. 2015. V. 49. № 14. P. 1561–1571.
Mamkaev Yu.V. Morphological interpretation of Darwinism // Paleontol. J. 2011. V. 44. № 12. P. 1509–1517.
Meyen S.V. Plant morphology in its nomothetical aspects // Bot. Rev. 1973. V. 39. Iss. 3. P. 205–260.
Meyen S.V. Basic features of Gymnosperm systematics and phylogeny as evidenced by the fossil record // Bot. Rev. 1984. V. 50. № 1. P. 1–111.
Notov A.A. Homeosis and evolution of modular organisms // Paleontol. J. 2015. V. 49. № 14. P. 1681–1690.
Notov A.A. Pseudocyclical similarities and structural evolution of modular organisms // Biol. Bull. 2016. V. 43. № 3. P. 226–234.
Notov A.A. How did modular organisms appear? Functional and evolutionary aspects // Wulfenia. 2017. V. 24. P. 75–91.
Notov A.A., Kusnetzova T.V. Architectural units, axiality and their taxonomic implications in Alchemillinae // Wulfenia. 2004. V. 11. P. 85–130.
Pardo J.D., Szostakiwskyj M., Ahlberg P.E., Anderson J.S. Hidden morphological diversity among early tetrapods // Nature. 2017. V. 546(7660). P. 642–645.
Rasnitsyn A.P., Aristov D.S., Rasnitsyn D.A. Dynamics of insect diversity during the early and middle permian // Paleontol. J. 2015. V. 49. № 12. P. 1282–1309.
Rutishauser R. Evolution of unusual morphologies in Lentibulariaceae (bladderworts and allies) and Podostemaceae (river-weeds): a pictorial report at the interface of developmental biology and morphological diversification (review) // An. Bot. 2016. V. 117. № 5. P. 811–832.
Simpson C., Jackson J.B.C., Herrera-Cubilla A. Evolutionary determinants of morphological polymorphism in colonial animals // Am. Natur. 2017. V. 190. № 1. P. 17–28.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая