

Известия РАН. Серия биологическая, 2019, № 2, стр. 215-224
Запасы фитомассы и органического углерода среднетаежных ельников при восстановлении после сплошнолесосечной рубки
А. Ф. Осипов *, В. В. Тужилкина, А. А. Дымов, К. С. Бобкова
Институт биологии Коми научного центра УрО РАН
167982 г. Сыктывкар, ул. Коммунистическая, 28, Россия
* E-mail: osipov@ib.komisc.ru
Поступила в редакцию 18.04.2017
После доработки 01.09.2017
Принята к публикации 07.08.2017
Аннотация
Проанализированы материалы, характеризующие лесовосстановительный процесс среднетаежного ельника черничного на автоморфных почвах после сплошнолесосечной рубки. Приведены данные по биоразнообразию растений и запасам углерода растительного органического вещества (ОВ) в коренном ельнике, на вырубке и приспевающем березняке, возникшем после рубки. Установлено, что в фитомассе коренного ельника аккумулируется 99.01, на вырубке – 18.28, а в 36-летнем березняке разнотравном – 56.48 т С/га. Отмечен вклад отдельных компонентов фитоценоза − древостоя, подроста и крупных древесных остатков − в концентрацию углерода ОВ.
Бореальные леса, выполняя важные средообразующие функции, играют огромную роль в регуляции климата и обмене веществ биосферы Северного полушария. Изучение динамики углерода органического вещества (ОВ) требует анализа влияния различных нарушений на функционирование лесных экосистем (Howard et al., 2004). Это позволит дать оценку связывания парниковых газов лесами России в случае ратификации Парижского соглашения по климату (United…, 2015). Особую роль в этих оценках играют формирующиеся молодняки и искусственно высаженные насаждения, которые рассматриваются как основные поглотители диоксида углерода (Филипчук и др., 2017).
Промышленные рубки − наиболее значимый антропогенный фактор, приводящий к нарушению структурной организации таежных биогеоценозов и естественных потоков вещества и энергии в них (Панов и др., 2012; Holtsmark, 2012). Рубка леса приводит к глубокой трансформации лесных экосистем и играет главную роль в сукцессионном процессе, в результате которого формируются специфические комплексы антропогенных фитоценозов (Walker, Willig, 1999).
Восстановление хвойных экосистем на вырубках еловых лесов таежной зоны происходит в результате смены производных лиственных и лиственно-хвойных насаждений. Это обусловлено обильным плодоношением лиственных (береза, осина, рябина) пород, большой дальностью распространения их семян, способностью восстанавливаться вегетативным путем, быстрым ростом, устойчивостью к конкуренции трав. К настоящему времени этот процесс на вырубках хвойных лесов европейского Севера достаточно изучен, разработаны схемы формирования типов вырубок в связи с исходными типами еловых лесов, выявлены зональные особенности естественного лесовозобновления (Мелехов, 1968; Побединский, 1973), динамика изменения биоразнообразия (Ильчуков, 2003 и др.), а также влияние рубок на почвы (Дымов, 2017). Биологическая продуктивность производных лиственно-хвойных молодняков на европейском Севере была рассмотрена ранее (Казимиров и др., 1978).
Однако для севера европейской территории России, где ведется интенсивная лесозаготовка, отмечается недостаток данных о динамике пулов ОВ и углерода на вырубках и в производных сообществах в процессе их формирования. Единичны данные о запасах фитомассы и углерода в подросте, которые характеризуют биологическую устойчивость экосистем. Не уделяется должного внимания изучению растений подлеска, оказывающего влияние на развитие травянистого яруса и возобновление древесных растений (Побединский, 1973). Численность подлеска, его состав и структура по высоте зависят от типа леса, класса бонитета, стадии развития, состава, сомкнутости крон деревьев и т.д. Подобные сведения о нарушенных территорих необходимы для объективной оценки экологических последствий хозяйственной деятельности в длительных временных масштабах (Ведрова и др., 2010).
Еловые леса на территории Республики Коми занимают 16.2 млн. га, а площадь ежегодных промышленных рубок в них составляет 40–60 тыс. га (Государственный…, 2016). В связи с этим теоретический и практический интерес представляют работы, объединяющие сведения о динамике растительности и пулов углерода ОВ в процессе восстановительной сукцессии ельников на вырубках.
Цель работы – оценка динамики запасов ОВ и углерода при восстановлении среднетаежного ельника черничного после сплошнолесосечной рубки.
МАТЕРИАЛЫ И МЕТОДЫ
Для исследования процесса восстановления лесных экосистем после вырубок можно использовать нескольких подходов. Первый − метод пространственно-временных аналогий, при котором в схожих лесорастительных условиях закладываются участки, находящиеся на разных стадиях развития (Yanai et al., 2003). Для применения этого подхода нужен тщательный подбор участков со сходными геологической историей и условиями почвообразования. Второй подход − наблюдение за развитием лесной экосистемы на постоянных пробных площадях (ППП). Первый подход позволяет получить данные о состоянии изучаемого признака на разных этапах развития экосистемы за короткий период наблюдений, второй подход более точен, хотя требует много временных затрат и преемственности поколений исследователей. В третьем подходе для характеристики динамики пулов ОВ применяют имитационное моделирование. Примеры таких моделей − РОБУЛ (Замолодчиков и др., 2011), EFIMOD (Shanin et al., 2011), CBM-CFS3 (Kurz et al., 2009)
Мы использовали метод пространственно-временных аналогий и провели исследования на территории Республики Коми в подзоне средней тайги в южной части Тиманского кряжа на возвышенности Джежимпарма. В ходе рекогносцировочных работ были подобраны три участка, находящихся на расстоянии 5–10 км один от другого. Объектами исследования были коренной ельник черничный, березняк разнотравный, возникший после сплошнолесосечной вырубки 1969–1970 гг., и 4-летняя вырубка ельника черничного, которые произрастают на типичных подзолистых почвах, сформированных на покровных суглинках одинакового генезиса (Дымов, Милановский, 2014). Даты рубок и исходные характеристики насаждений установлены по данным Усть-Куломского лесничества.
Коренной ельник черничный (ППП-1, 61°45′47.4″ с.ш., 54°17′48.1″ в.д.). Сложный по составу (6Е4Пх + Б) древесный ярус состоит из Picea obovata Ledeb., Abies sibirica Ledeb., Betula pubescens Ehrh. Древостой с умеренной численностью деревьев (825 экз./га) разновысотный. Он характеризуются средней полнотой, IV классом бонитета, запасом древесины растущих деревьев 236 м3/га (табл. 1). Сухостой в древостое 66 экз./га с запасом древесины 15 м3/га представлен елью и пихтой. Подрост, произрастающий группами, имеет состав 71Е22Пх7Б. Его густота 2.6 тыс. экз./га, из них 2.3 тыс. экз./га − растущий подрост (рис. 1). Подлесок состоит из Sorbus aucuparia L., единично Rosa acicularis Lindl., Lonicera pallustris Ledeb. Травяно-кустарничковый ярус с проективным покрытием ~50% имеет простое синузиальное строение. Моховой покров с проективным покрытием 80−85% образован главным образом зелеными мхами, в микропонижениях встречаются сфагновые мхи. Видовая насыщенность растений нижних ярусов небогатая (табл. 2).
Таблица 1.
Таксационная характеристика исследуемых древостоев
| Тип леса (№ ППП) | Состав древостоя |
Порода | Возраст, лет |
Число деревьев, экз./га |
Сумма площадей сечений, м2/га |
Запас древесины, м3/га |
Средние | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| растущих | сухих | растущих | сухих | растущих | сухих | высота, м | диаметр, см | ||||
| Ельник черничный (1) | 6Е4Пх + Б | Ель | 60–230 | 408 | 33 | 15.6 | 1.4 | 132 | 16 | 14 | 18 |
| Пихта | 70–120 | 392 | 25 | 11.7 | 1.1 | 107 | 1 | 14 | 18 | ||
| Береза | не опр. | 25 | –* | 1 | – | 7 | – | 13 | 15 | ||
| Березняк разнотравный (3) | 7Б2Е1Пх ед.Ос |
Ель | 60–70 | 418 | – | 5.4 | – | 34 | – | 10 | 12 |
| Пихта | 50–70 | 143 | 11 | 4 | 0.1 | 9 | 0.2 | 8 | 10 | ||
| Береза | 25–34 | 1601 | 11 | 19.5 | 0.1 | 143 | 0.1 | 15 | 9.9 | ||
| Осина | 25–34 | 10 | – | 0.1 | – | 1 | – | 16 | 13 | ||
Рис. 1.
Распределение подроста по категориям крупности в насаждениях восстановительной сукцессии ельника. 1 – здоровый, 2 – сомнительный, 3 – усыхающий, 4 – сухой.
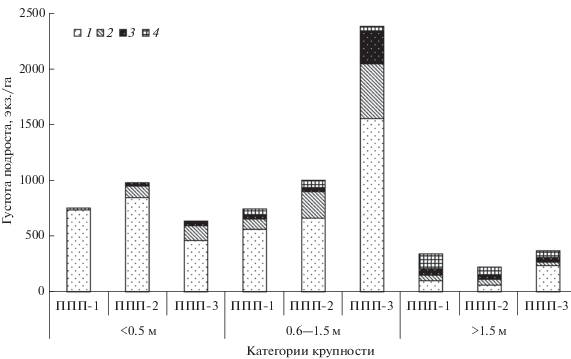
Таблица 2.
Состав и обилие (баллы) растений напочвенного покрова
| Жизненная форма, вид | ППП-1 | ППП-2 | ППП-3 |
|---|---|---|---|
| Кустарнички | |||
| Vaccinium myrtillus L. | 4 | 1 | 2 |
| V.vitis idaea L. | + | 2 | – |
| Травы | |||
| Aconitum excelsum Reichb. | – | – | + |
| Angelica sylvestris. L. | – | – | + |
| Bacethryon caespitosum (L.) A.Diet. | – | 1 | – |
| Calamagrosis sp. | – | 2 | – |
| Carex( sp.) | 1 | 1 | 2 |
| Chamaenerion angustifolium (I.) Scop. | – | 2 | – |
| Dryopteris (sp.) | 3 | – | 2 |
| Equisetum sylvaticum L. | 2 | 3 | 2 |
| Gymnocarpium Linnea (L.) Newm. | – | 1 | 4 |
| Linnaea borealis L. | 4 | 1 | + |
| Luzula pilosa (L.) Wild. | 2 | 2 | 2 |
| Lycopodium annotinum L. | 2 | + | 1 |
| Maintheum bifolium (L.) F.W.Schmidt. | 4 | + | 3 |
| Melampyrum sylvaticum L. | – | – | + |
| Oxalis acetosella L. | 4 | + | 5 |
| Pyrola rotundifolia L. | – | – | 1 |
| Rubus arcticus L. | – | 2 | – |
| Solidago virgaurea L. | – | – | 1 |
| Stellaria bungeana Fenz | 2 | – | 2 |
| Trientalis europaea L. | 2 | + | 4 |
| Мхи | |||
| Dicranum polysetum (Mich.) Sw. | 2 | 2 | 2 |
| Hylocomium splendens (Hedw.) Br.,Sch.et Gmb. | 2 | 2 | 1 |
| Pleurozium schreberi (Brid) Mitt. | 5 | 3 | 1 |
| Polytrichum commune Hedw. | 2 | 2 | 1 |
| Sphagnum girgensohnii Russ. | 1 | + | – |
| Sphagnum russowii.Warnst. | + | + | – |
Вырубка ельника черничного (ППП-2 61°59′′3.1´´ с. ш., 54°05′50.6″ в.д.). Фитоценоз развивается после сплошнолесосечной рубки, которая была проведена зимой 2001/2002 г. До рубки древостой был смешанный по составу 7Е2Б1Пх, разновозрастнoй с запасом древесины 258 м3/га, IV класса бонитета, полнотой 0.7. Под пологом в достаточном количестве развивается подрост. Сортиментная заготовка древесины осуществляется по скандинавской технологии с использованием на пасеках лесозаготовительной техники (пара харвестер–форвардер). На вырубке происходит интенсивное восстановление лесной растительности, основа которой – подрост ели предварительной генерации. Состав формирующегося молодняка 45Е45Б10Пх. Численность елового подроста 2.2 тыс. экз./га, из них здоровых перспективных 71, сомнительных 18, усыхающих 5 и сухих 6% (рис. 1). Отмечается также появление большого количества подлесочного древесного растения – рябины (3.5 тыс. экз./га) вегетативного происхождения, развивающейся от сохранившихся в процессе рубки деревьев. Средняя высота ее равна 1.2 м. Также из подлесочных пород единично встречаются ива и малина.
Сохранились тонкомерные деревья ели (33 экз./га) и пихты (67 экз./га) с объемом древесины 6.2 м3/га. У ели средний диаметр оставленных деревьев 10 см, средняя высота 8 м, у пихты – 10 см и 13 м соответственно. Разнообразие растений живого напочвенного покрова и их встречаемость представлены в табл. 2.
Березняк разнотравный (ППП-3, 61°48′25.6″ с.ш., 54°04′0.6″ в.д.). Он развивается после рубки ельника черничного. Для древостоя до рубки характерны состав 7Е2Б1Пх с запасом древесины 260 м3/га, полнота 0.7, IV класс бонитета. Сплошнолесосечная рубка была проведена зимой 1969/1970 г. Древесину вывозили хлыстами на гусеничном тракторе. В настоящее время древостой березняка формируют B. pubescens, P. obovata, A. sibirica, Populus tremula (7Б2Е1Пх ед. Ос., запасы древесины древостоя 187 м3/га, его полнота 29 м2/га, класс бонитета II, береза послерубочного происхождения). Редкие сухостойные деревья (22 экз./га) представлены пихтой и березой (табл. 1). Подлесок образуют рябина (2.1 тыс. экз./га) и ива (20 экз./га). Подрост состоит из ели (3.4 тыс. экз./га), пихты (0.5 тыс. экз./га), березы (1.1 тыс. экз./га), состав его 68Е9Пх23Б. Ель и пихта (в основном здоровые, последующего (послерубочного) происхождения) представлены деревьями мелкой и средней крупности (рис. 1). Значительная часть их произрастает на трелевочных волоках. Травяно-кустарничковый ярус с проективным покрытием ~40% состоит в основном из травянистых растений. Мохово-лишайниковый покров имеет проективное покрытие 30% и образован зелеными мхами (табл. 2).
Закладка круговых ППП площадью 300 м2, перечет деревьев, характеристика их состояния проводились согласно методам международной программы ICP Forest (Manual…, 1994). В коренном ельнике чернично-зеленомошном были заложены четыре круговые ППП, в березняке разнотравном и на вырубке – по три. Был проведен сплошной перечет деревьев и подроста. Определено их жизненное состояние. У каждого дерева определялся диаметр на высоте 1.3 м, а у растений подроста – высота. У 25 деревьев доминирующей породы и по 10 у каждой сопутствующей породы измеряли высоты с помощью высотомера Haglof (Швеция). Затем для каждой породы выводили регрессионное уравнение зависимости высоты от диаметра. Таксационная обработка материала выполнялась стандартными методами. Определение запасов древесины осуществляли по региональным таблицам зависимости объема ствола от его высоты и диаметра (Лесотаксационный…, 2012). Состав древостоя рассчитывали по вкладу отдельных видов древесных растений в общие запасы древесины древостоя, состав подроста – по доле особей каждой породы. Подрост распределяли по категориям крупности: мелкий (<0.5 м), средний (0.6–1.5 м) и крупный (>1.5 м).
Описание растений напочвенного покрова проводилось на пяти учетных площадках на каждом участке исследования. Обилие растений определяли в цифровом выражении по шкале Друде. Для характеристики сходства растений между ППП применяли коэффициент Жаккара (Методы…, 2002).
Под крупными древесными остатками (КДО) мы понимали сухостойные и упавшие деревья диаметром 6 см и более, деревья со сломанными верхушками, пни (Russel et al., 2015). На вырубке также были учтены порубочные остатки. При перечете учитывали диаметр, высоту дерева и стадию гниения древесины. Выделяли пять стадий гниения древесины по морфологическим признакам, характеризующим покрытие корой, наличие крупных и мелких ветвей, твердость древесины (Стороженко, 2007). Массу стволов и пней вычисляли по их объему и базисной плотности (БП) древесины в зависимости от стадии гниения (Sandström et al., 2007). Результаты, характеризующие БП гниющей древесины в регионе исследований, были опубликованы нами ранее (Бобкова и др., 2015).
Запасы ОВ в древесном ярусе определяли с помощью регрессионных уравнений зависимости массы отдельных фракций деревьев от диаметра ствола на высоте 1.3 м., а растений подроста − от высоты особи. Характеристики уравнений и их статистические показатели были приведены ранее (Бобкова и др., 2014). Уравнения выведены на основе анализа 5−10 модельных деревьев. Для определения массы растений напочвенного покрова применяли метод укосов на площади 0.25 × 0.25 см в 40-кратной повторности на каждой ППП. Данные по фитомассе приведены в абсолютно сухой массе.
Пересчет запасов ОВ фитомассы древостоя, подлеска и напочвенного покрова на углерод осуществляли с помощью коэффициентов для каждой фракции (Бобкова, Тужилкина, 2001). Так, концентрация углерода в хвое и листьях древесных пород составила 46–47, в древесине ~47, в ветвях 46–48, в коре 48–50, в кустарничках 48–53, травах ~42, в мхах 42–45%. Для КДО использовали коэффициент 0.5.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Запасы углерода фитомассы древостоев. В древостое коренного старовозрастного ельника черничного сконцентрировано 165.42 т/га ОВ или 79.33 т /га углерода, большая часть которого приходится на древесину ствола (58%) и корни (23%) (табл. 3). Доли хвои и ветвей составляют по 7, коры – 5%. Чуть больше половины фитомассы древостоя сформировано елью. Значимо (43%) участие деревьев пихты, а роль березы в общих запасах фитомассы и углерода несущественна.
Таблица 3.
Запасы фитомассы (над чертой) и углерода (под чертой) в древостоях, т/га
| Фитоценоз | Порода | Хвоя (листья) | Древесина ствола | Кора ствола | Ветви | Корни |
|---|---|---|---|---|---|---|
| Ельник черничный | Ель | $\frac{{6.21 \pm 1.49}}{{2.86 \pm 0.69}}$ | $\frac{{55 \pm 6.05}}{{26.12 \pm 2.87}}$ | $\frac{{5.04 \pm 0.76}}{{2.51 \pm 0.38}}$ | $\frac{{6.21 \pm 1.43}}{{3.04 \pm 0.7}}$ | $\frac{{21.3 \pm 2.34}}{{10.29 \pm 1.13}}$ |
| Пихта | $\frac{{5.4 \pm 1.3}}{{2.49 \pm 0.6}}$ | $\frac{{40.13 \pm 4.41}}{{19.06 \pm 2.1}}$ | $\frac{{3.98 \pm 0.6}}{{1.98 \pm 0.3}}$ | $\frac{{}}{{}}$$\frac{{5.36 \pm 1.23}}{{2.62 \pm 0.6}}$ | $\frac{{15.53 \pm 1.71}}{{7.77 \pm 0.83}}$ | |
| Береза | $\frac{{0.01 \pm 0.005}}{{0.004 \pm 0.002}}$ | $\frac{{0.78 \pm 0.26}}{{0.36 \pm 0.12}}$ | $\frac{{0.12 \pm 0.03}}{{0.06 \pm 0.02}}$ | $\frac{{0.11 \pm 0.02}}{{0.05 \pm 0.01}}$ | $\frac{{0.24 \pm 0.05}}{{0.12 \pm 0.02}}$ | |
| Березняк разнотравный | Ель | $\frac{{3.13 \pm 0.75}}{{1.44 \pm 0.35}}$ | $\frac{{15.55 \pm 1.71}}{{7.38 \pm 0.81}}$ | $\frac{{1.82 \pm 0.27}}{{0.91 \pm 0.14}}$ | $\frac{{3.05 \pm 0.7}}{{1.49 \pm 0.34}}$ | $\frac{{6.01 \pm 0.66}}{{2.9 \pm 0.32}}$ |
| Пихта | $\frac{{1.45 \pm 0.35}}{{0.67 \pm 0.16}}$ | $\frac{{8.84 \pm 0.97}}{{4.2 \pm 0.46}}$ | $\frac{{0.94 \pm 0.14}}{{0.47 \pm 0.07}}$ | $\frac{{1.42 \pm 0.33}}{{0.69 \pm 0.16}}$ | $\frac{{3.42 \pm 0.38}}{{1.65 \pm 0.18}}$ | |
| Береза | $\frac{{0.51 \pm 0.18}}{{0.24 \pm 0.08}}$ | $\frac{{39.75 \pm 13.52}}{{18.67 \pm 6.35}}$ | $\frac{{6.94 \pm 1.81}}{{3.49 \pm 0.91}}$ | $\frac{{5.59 \pm 0.84}}{{2.58 \pm 0.39}}$ | $\frac{{9.96 \pm 1.99}}{{4.64 \pm 0.93}}$ | |
| Вырубка ельника (недоруб) | Ель | $\frac{{0.16 \pm 0.04}}{{0.07 \pm 0.02}}$ | $\frac{{0.58 \pm 0.06}}{{0.28 \pm 0.03}}$ | $\frac{{0.08 \pm 0.01}}{{0.04 \pm 0.01}}$ | $\frac{{0.15 \pm 0.04}}{{0.08 \pm 0.02}}$ | $\frac{{0.23 \pm 0.02}}{{0.11 \pm 0.01}}$ |
| Пихта | $\frac{{0.47 \pm 0.11}}{{0.22 \pm 0.05}}$ | $\frac{{2.04 \pm 0.22}}{{0.97 \pm 0.11}}$ | $\frac{{0.26 \pm 0.04}}{{0.13 \pm 0.02}}$ | $\frac{{0.46 \pm 0.11}}{{0.23 \pm 0.05}}$ | $\frac{{0.79 \pm 0.09}}{{0.79 \pm 0.09}}$ |
В древесном ярусе 36-летнего березняка разнотравного запасы фитомассы составляют 108.38 т/га или 51.42 т С/га, большую часть которой (58%) аккумулируют деревья березы. В формировании древостоя участие ели и пихты составляет 27 и 15% соответственно. Древесина ствола − основное депо ОВ фитоценоза. Она концентрирует 59% общих его запасов, на кору и ветви приходится по 9, на листья – 5, корни – 19%.
На вырубке ельника черничного запасы ОВ в тонкомерных деревьях пихты и ели, оставшихся после рубки, незначительны и составляют 5.22 т/га или 2.51 т С/га. Распределение по фракциям следующее: древесина ствола (50%), корни (20%), хвоя и ветви (по 12%), кора (6%).
Массы ОВ и углерода древостоя березняка разнотравного также сопоставимы с ранее опубликованными. В условиях средней тайги Карелии фитомасса надземной части древостоев березняков сходного возраста варьирует от 55 до 97 т/га (Казимиров и др., 1978). Участие отдельных органов древесных растений в общей массе древостоя характерно для лесных насаждений таежной зоны.
Фитомасса и углерод подроста. В коренном ельнике черничном, произрастающем на автоморфных почвах, происходит возобновление ели и пихты. После рубки древостоя в формировании фитоценоза на вырубках значительную роль играет сохранившийся подрост хвойных растений, выражен восстановительный процесс березы и подлесочной породы – рябины (рис. 1). К 36 годам на вырубке ельника сформировался смешанный лиственно-хвойный древостой, при доминировании березы с участием ели и пихты. Под его пологом достаточно хорошо развит еловый подрост, который по мере отпада березы в последующем займет господствующее положение в составе древостоя. Такая смена древесной растительности на вырубках характерна для ельников средней тайги.
Биомасса и запасы углерода в надземных органах растений подроста незначительны. Так, в ельнике черничном аккумулировано 1.01 т/га ОВ или 0.49 т С /га, на четырехлетней вырубке ельника черничного – 0.62 или 0.3, в березняке разнотравном – 2.06 или 0.99 соответственно. В зависимости от стадии сукцессии растительного покрова вклад отдельных пород в общую массу различается. Доля ели во всех сообществах примерно одинакова и составляет 52–55%. В общей фитомассе подроста отмечена значительная вариация в участии березы, запасы которой незначительны в коренном ельнике зеленомошном и существенны (31%) в березняке разнотравном. Несмотря на небольшое количество, важную роль в массе ОВ играют крупные особи подроста, в которых сосредоточено 64–85% общих запасов ОВ. Увеличение доли участия в фитомассе подроста средней крупности до 34% в общей биомассе этого элемента экосистемы отмечено на свежей вырубке, что связано с его густотой (2.08 тыс. экз./га) в этом сообществе.
Выявление закономерностей распределения фитомассы по фракциям в молодом возрасте интересно с точки зрения уточнения их биометрических характеристик. В общих запасах надземной фитомассы и углерода 57−59% концентрируют стволы растений подроста. Участие ветвей составляет 19–22%, ассимиляционного аппарата − 20–24%. Корни подроста не учитывали в связи с большими неопределенностями при их оценке. Это связано с тем, что подрост лиственных пород часто имеет вегетативное происхождение, когда из одного корня произрастает несколько особей. В связи с этим дать адекватную оценку биомассе подземных органов не представляется возможным без дополнительных исследований.
Фитомасса и углерод подлеска. В коренном ельнике черничном подлесок сформирован деревьями рябины разного размера плотностью 1892 экз./га, у 142 из них диаметры 6–12 см, высоты <10 м. Они сосредоточены на нижних и средних ступенях толщины. Запасы углерода в этом ярусе елового фитоценоза составляют 5.55 ± 1.72 т/га. Концентрация фитомассы и углерода в подлеске на вырубке существенно ниже (0.52 ± 0.16 и 0.26 ± 0.08 т/га соответственно), чем в ельнике черничном. Число деревьев рябины, также принадлежащих к низким (6 и 8 см) ступеням толщины, в березняке разнотравном (122 экз./га) сопоставимо с коренным ельником черничным. Вместе с тем в сообществе достаточно много (2133 экз./га) подроста рябины. Всего в растениях рябины березняка разнотравного сосредоточено 3.26 ± 1.01 т С/га, образованных преимущественно крупными особями.
Запасы углерода в растениях напочвенного покрова. При рубке и в процессе формирования фитоценоза на вырубке происходят существенные изменения в проективном покрытии и биоразнообразии растений нижних ярусов (табл. 2). Так, коэффициент видового сходства растений между старовозрастным ельником и вырубкой равен 0.69, между вырубкой и березняком – 0.52, ельником и березняком – 0.58. На вырубке по сравнению с ельником заметно увеличение видов злаковых растений, а в березняке – травянистых растений. В березняке разнотравном отсутствуют сфагновые мхи.
В коренном ельнике черничном масса растений напочвенного покрова составляет 0.75 т/га, большая часть которого образована мхами (49%) и корнями трав и кустарничков (41%). На долю надземных органов кустарничков и трав приходится ~10% (табл. 4). На вырубке ельника черничного в результате резкого улучшения светового режима происходит увеличение запасов углерода (до 1.42 т С/га) в растениях этого яруса в первую очередь за счет трав, надземные органы которых формируют 11% общей массы. Ухудшение гидрологического режима почвы в результате рубки ведет за собой увеличение массы мхов (главным образом сфагновых). Более низкие концентрации (0.26 т С/га) запасов углерода отмечены в березняке разнотравном. Вероятно, это связано с развитым древесным пологом, пропускающим к поверхности почвы небольшое количество солнечной радиации.
Таблица 4.
Запасы углерода в растениях напочвенного покрова, кг/га
| Фитоценоз | Кустарнички | Травянистые растения | Корни трав и кустарничков | Мхи | Итого |
|---|---|---|---|---|---|
| Ельник черничный | 43 ± 14 | 38 ± 14 | 303 ± 29 | 362 ± 66 | 746 ± 123 |
| Березняк разнотравный | 14 ± 7 | 24 ± 1 | 142 ± 14 | 76 ± 28 | 256 ± 50 |
| Вырубка ельника черничного | 0.4 ± 0.3 | 157 ± 39 | 588 ± 56 | 670 ± 134 | 1415 ± 229 |
Запасы углерода в КДО. В коренном ельнике черничном в сухостое ели и пихты (58 экз./га) концентрируется в древесине 2.93 ± 0.21 т С/га (табл. 5). Довольно много валежа различных стадий гниения с запасом углерода 9.96 ± 0.66 т/га. Распределение КДО по классам деструкции свидетельствует о постепенном выпадении деревьев. Так, значительная (44%) его доля приходится на четвертую, 32% − на третью, 21% − на вторую, 3% − на первую стадии разложения. Доля валежа, сформированного пихтой, составляет 52%, однако ее участие в массе КДО составляет 27%. Доли березы и ели в общей численности деревьев составляют 26 и 22% соответственно, а их запасы − 32 и 41% общего объема. В КДО ельника черничного (316 экз./га) сосредоточено 104.3 м3/га древесины с аккумуляцией углерода 12.89 ± 1.05 т/га. Запасы древесины и углерода в КДО составляют 38 и 44% массы и запасов древесины в живых деревьях.
Таблица 5.
Запасы углерода в крупных древесных остатках, т/га
| Порода | Сухостой | Валеж по классам деструкции | Порубочные остатки | Всего | ||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | Итого | ||||
| Коренной ельник черничный | ||||||||
| Ель | 2.78 | −1 | 0.98 | 2.05 | 1.01 | 4.04 | − | 6.82 |
| Пихта | 0.15 | 0.32 | 0.27 | 0.99 | 1.14 | 2.72 | − | 2.87 |
| Береза | − | 0.02 | 0.84 | 0.09 | 2.25 | 3.2 | − | 3.2 |
| Вырубка ельника черничного | ||||||||
| Ель | 0.04 | 2.24 | 2.26 | 0.38 | 1.98 | 6.86 | 5.162 | 6.9 |
| Пихта | − | 0.49 | − | − | − | 0.49 | − | 0.49 |
| Береза | − | − | 1.24 | − | − | 1.24 | − | 1.24 |
| Березняк разнотравный | ||||||||
| Пихта | 0.04 | − | − | − | 0.44 | 0.44 | − | 0.48 |
| Береза | 0.01 | − | − | − | − | − | − | 0.02 |
Запасы КДО на четырехлетней вырубке образуют как отмершие деревья, унаследованные от материнского насаждения, так и остатки, появившиеся в результате рубки. Следует отметить, что КДО сформированы в основном валежом густотой 278 экз./га с запасом древесины 61.5 м3/га, в котором аккумулируется 8.63 ± 0.67 т С/га, и порубочными остатками. Ранее (Bobkova et al., 2007) для этого сообщества было установлено, что запасы древесины в порубочных остатках, включающих в себе обломки и вершины деревьев диаметром <6 см, оставленные при сортиментной заготовке древесины, составляют 11% запаса исходного насаждения, что составляет 25.18 м3/га или 5.16 ± 0.20 т С/га. Сухостой единичен и образован тонкомерными деревьями ели. Всего в КДО вырубки сконцентрировано 13.79 ± 0.87 т С/га, что вполне сопоставимо с ненарушенным коренным ельником.
36-летний березняк разнотравный, развивающийся после сплошнолесосечной рубки ельника черничного, характеризуется небольшими запасами КДО. Начало интенсивной дифференциации деревьев по состоянию не наблюдается. Признак этого − небольшая численность сухостоя, состоящего из тонкомерных деревьев пихты и березы (густота 22 экз./га), где сконцентрировано 0.05 т С/га. Вследствие слабого отпада валеж единичен и представлен сильно фрагментированными остатками на четвертой стадии разложения. Древесина валежа в таежной зоне разлагается до четвертой стадии в подзоне южной и северной тайги за 35–45 лет соответственно (Стороженко, 2007). Следовательно, в березняке спустя 36 лет после рубки в насаждении отмечается практически полная деструкция КДО, унаследованных от материнского древостоя. Всего масса углерода КДО березняка разнотравного составляет ~0.5 т С/га.
Полученные нами данные по запасам КДО подтверждают имеющиеся в литературе сведения об U-образной динамике массы КДО насаждений, возникших после разного рода катастрофических нарушений (Замолодчиков, 2009). Согласно данным автора, сразу после нарушения запасы КДО, которые включают как унаследованный КДО, так и новообразованный, сопоставимы с ненарушенным сообществом. Затем вследствие деструкции масса углерода в КДО снижается. Отмирание особей подроста и тонкомера с небольшой массой ОВ не способствует накоплению КДО на начальных этапах развития ельников после сплошнолесосечных рубок. Поэтому более высокие концентрации их отмечены в старовозрастном ельнике зеленомошном и на вырубке ельника. Следует также отметить, что запасы КДО в пересчете на углерод в перестойных ельниках исследуемого региона варьируют от 9 до 17 т С/га (Бобкова и др., 2015). Полученные нами данные по концентрации углерода в КДО вполне вписываются в эти пределы.
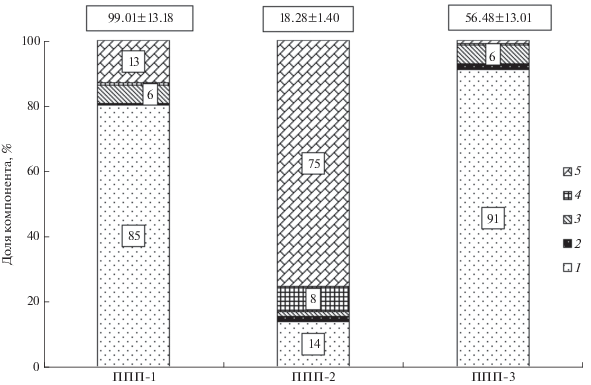
Общие запасы углерода. В процессе послерубочной восстановительной сукцессии ельника черничного наблюдается постепенное увеличение запасов ОВ и углерода в насаждениях (рис. 2). Одновременно с этим происходит изменение роли отдельных компонентов. Так, в старовозрастном ельнике и 36-летнем березняке основной пул ОВ и углерода − древостой. В фитоценозах этих сообществ участие подлеска составляет 6%. Вклад подроста на начальных стадиях восстановительной сукцессии 2% и снижается практически до нуля в коренном ельнике черничном. Доля растений напочвенного покрова выше в сообществе на четырехлетней вырубке. По мере усиления средообразующей роли древостоя, перехватывающего большую часть солнечной радиации, в процессе сукцессии вклад напочвенного покрова в общую массу ОВ и углерода уменьшается. Древесина КДО формирует 75% запасов ОВ и углерода на четырехлетней вырубке ельника черничного. По мере деструкции и восстановления древостоя ее роль постепенно снижается и достигает 1% в 36-летнем березняке разнотравном. С развитием насаждения происходит постепенное накопление КДО, участие которых в старовозрастном ельнике составляет 13%.
ЗАКЛЮЧЕНИЕ
Получены новые сведения, сочетающие данные о динамике биоразнообразия, таксационных характеристик древостоев и запасов углерода ОВ в процессе восстановительной сукцессии после сплошнолесосечной рубки среднетаежного ельника на типичных подзолистых почвах. Установлено, что по мере развития в насаждениях происходит накопление запасов углерода. Одновременно происходит изменение роли отдельных компонентов в его общей массе. Так, в коренном ельнике черничном на типичных подзолистых почвах запасы углерода составляют 99.01 т/га. На вырубке ельника черничного сконцентрировано 18.28 т С/га (образованы в основном КДО). В 36-летнем разнотравном березняке в пуле углерода (56.48 т/га) исключительную роль играет древостой. Приведенные данные подтверждают теорию процесса естественного лесовосстановления еловых экосистем после промышленных рубок. Они являются основой для дальнейших исследований, сочетая наблюдение на ППП и метод пространственно-временных аналогий.
Работа выполнена в рамках темы НИР “Пространственно-временная динамика структуры и продуктивности фитоценозов лесных и болотных экосистем на европейском Северо-Востоке России” (номер государственного учета НИОКТР АААА-А17-117122090014-8), при частичной финансовой поддержке проекта Комплексной программы УрО РАН 18-4-4-29 “Зональные закономерности бюджета углерода в лиственно-хвойных экосистемах европейского Северо-Востока” и гранта Президента Российской Федерации для молодых кандидатов наук МК-6670.2016.5.
Список литературы
Бобкова К.С., Тужилкина В.В. Содержание углерода и калорийность органического вещества в лесных экосистемах Севера // Экология. 2001. № 1. С. 69–71.
Бобкова К.С., Кузнецов М.А., Осипов А.Ф. Запасы крупных древесных остатков в ельниках средней тайги европейского Северо-Востока // Изв. вузов. Лес. журн. 2015. № 2. С. 9–20.
Бобкова К.С., Машика А.В., Смагин А.В. Динамика содержания органического вещества в среднетаежных ельниках на автоморфных почвах. СПб.: Наука, 2014. 270 с.
Ведрова Э.Ф., Мухортова Л.В., Иванов В.В., Кривобоков Л.В., Болонева М.В. Восстановление запасов органического вещества после рубок в лесных экосистемах восточного Прибайкалья // Изв. РАН. Сер. биол. 2010. № 1. С. 83−94.
Государственный доклад “О состоянии окружающей среды Республики Коми в 2015 году”: Министерство природных ресурсов и охраны окружающей среды Республики Коми. Сыктывкар: ГБУ РК ТФИ РК, 2016. 173 с.
Дымов А.А. Влияние сплошных рубок в бореальных лесах России на почвы (обзор) // Почвоведение. 2017. № 7. С. 787–798.
Дымов А.А., Милановский Е.Ю. Изменение органического вещества таежных почв в процессе естественного лесовозобновления растительности после рубок (средняя тайга Республики Коми) // Почвоведение. 2014. № 1. С. 39−47.
Замолодчиков Д.Г. Оценка пула углерода крупных древесных остатков в лесах России с учетом влияния пожаров и рубок // Лесоведение. 2009. № 4. С. 3–15.
Замолодчиков Д.Г., Грабовский В.Г., Краев Г.Н. Динамика бюджета углерода лесов России за два последних десятилетия // Лесоведение. 2011. № 6. С. 16–28.
Ильчуков С.В. Динамика структуры лесного покрова на сплошных вырубках (подзона средней тайги, Республика Коми). Екатеринбург: УрО РАН, 2003. 120 с.
Казимиров Н.И., Морозова Р.М., Куликова В.К. Органическая масса и потоки веществ в березняках средней тайги. Л.: Наука, 1978. 216 с.
Лесотаксационный справочник по Северо-Востоку европейской части Российской Федерации. Архангельск: Правда Севера, 2012. 672 с.
Мелехов И.С. Динамическая типология леса // Лес. хоз-во. 1968. № 3. С. 15–21.
Методы изучения лесных сообществ / Под ред. Ярмишко В.Т., Лянгузова Н.В. СПб.: НИИХимии СПбГУ, 2002. 240 с.
Панов А.В., Онучин А.А., Зражевская Г.К., Шибистова О.Б. Структура и динамика пулов органического вещества на вырубках в сосняках лишайниковых среднетаежной подзоны Приенисейской Сибири // Изв. РАН. Сер. биол. 2012. № 6. С. 658–666.
Побединский А.В. Рубки и возобновление в таежных лесах СССР. М.: Лес. пром-сть, 1973. 200 с.
Стороженко В.Г. Устойчивые лесные сообщества. Теория и эксперимент. Тула: Гриф и К, 2007. 192 с.
Филипчук А.Н., Моисеев Б.Н., Малышева Н.В. Методика учета поглощения СО2 лесами Российской Федерации // Леса России: политика, промышленность, наука, образование: Матер. Второй междунар. науч.-техн. конф. Т. 2. СПб.: СПбГЛТУ, 2017. С. 155–158.
Bobkova K.S., Tuzhilkina V.V., Simonov G.A. Removal of nutrients and carbon at logging of spruce for industrial needs in the Russian European North-East / Forest soils: research results, problems and futureoutlook: Abstr. Intern. Conf. (Syktyvkar, Russia, 4–11 September, 2007). Syktyvkar, 2007. P. 193.
Holtsmark B. Harvesting in boreal forests and the biofuel carbon debt // Clim. Change. 2012. V. 112(2). P. 415–428.
Howard E.A., Gower S.T., Foley J.A., Kucharik C.J. Effects of logging on carbon dynamics of a jack pine forest in Saskatchewan, Canada // Glob. Change Biol. 2004. V. 10. P. 1267–1284.
Kurz W.A., Dymond C.C., White T.M., Stinson G., Shaw C.H., Rampley G.J., Smyth C., Simpson B.N., Neilson E.T., Trofymow J.A., Metsaranta J., Apps M.J. CBM-CFS3: A model of carbon-dynamics in forestry and land-use change implementing IPCC standards // Ecol. Modell. 2009. V. 220. P. 480–504.
Manual on Methodologies and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests. Hamburg; Prague: United Nations Environment Programme and Economic Commission for Europe, 1994. 477 p.
Russell M.B., Fraver S., Aakala T., Gove J.H., Woodall C.W., D’Amatof A.W., Ducey M.J. Quantifying carbon stores and decomposition in dead wood: A review // Forest Ecol. Manag. 2015. V. 350. P. 107–128.
Sandström F., Petersson H., Kruys N., Ståhl G. Biomass conversion factors (density and carbon concentration) by decay classes for dead wood of Pinus sylvestris, Picea abies and Betula spp. in boreal forests of Sweden // Forest Ecol. Manag. 2007 V. 243. P. 19–27.
Shanin V.N., Komarov A.S., Mikhailov A.V., Bykhovets S.S. Modelling carbon and nitrogen dynamics in forest ecosystems of Central Russia under different climate change scenarios and forest management regimes // Ecol. Modell. 2011. V. 222. P. 2262–2275.
United Nations, United Nations Framework Convention on Climate Change (UNFCC), Conference of the parties, Report of the conference of the parties on its twenty-first session, held in Paris from 30 November to 13 December 2015. Available at: (http://unfccc.int/resource/docs/2015/cop21/eng/10a01.pdf).
Walker L.R., Willig M.R. An introduction to terrestrial disturbances // Ecosystems of disturbed ground / Ed. Walker L.R. Amsterdam: Elsevier Sci, 1999. P. 1–16.
Yanai R.D., Currie W.S., Goodale C.L. Soil Carbon dynamics after forest harvest: an ecosystem paradigm reconsidered // Ecosystems. 2003. V. 6. P. 197–212.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая