

Известия РАН. Серия биологическая, 2019, № 5, стр. 457-465
Холодоустойчивость и распространение филогенетических линий дождевого червя Eisenia nordenskioldi (Oligochaeta, Lumbricidae)
Д. И. Берман 1, Н. А. Булахова 1, 2, *, Е. Н. Мещерякова 1, С. В. Шеховцов 1, 3
1 Институт биологических проблем Севера ДВО РАН
685000 Магадан, ул. Портовая, 18, Россия
2 Научно-исследовательский институт биологии и биофизики, Томский государственный университет
634050 Томск, просп. Ленина, 36, Россия
3 Федеральный исследовательский центр, Институт цитологии и генетики СО РАН
630090 Новосибирск, просп. Академика Лаврентьева, 10, Россия
* E-mail: sigma44@mail.ru
Поступила в редакцию 04.03.2019
После доработки 15.04.2019
Принята к публикации 15.04.2019
Аннотация
Отмечено, что таксономический статус филогенетических линий, выделенных в пределах широко распространенного вида дождевых червей Eisenia nordenskioldi, не ясен; в пользу продвинутости внутривидовой дифференциации свидетельствуют различия их холодоустойчивости. Обнаружено, что среди четырех исследованных линий подвида E. n. nordenskioldi и двух линий подвида E. n. pallida есть как умеренно устойчивые (переносящие охлаждение –10…–12°C), так и выживающие при значительно более низких температурах (–28…–34°C). Установлено, что характер ареалов некоторых генетических линий может быть объяснен резистентностью червей к отрицательным температурам, а в ряде других случаев ареалы генетических линий не совпадают с ожидаемыми пространственными пределами.
Широкое применение молекулярных методов для изучения структуры таксонов с громадными ареалами и развитие филогеографии как мощного направления – знаки современной биологии. Недавно появились публикации и о генетической неоднородности ряда видов дождевых червей (King et al., 2008; Novo et al., 2009; Porco et al., 2013; и др.), в том числе Eisenia nordenskioldi Eisen, 1879, распространенного в Евразии от Восточной Европы до Тихого океана (Shekhovtsov et al., 2016, 2018b; Шеховцов и др., 2017) (рис. 1). Этот вид ключевой для Сибири и, как считалось (Перель, 1979), единственный среди дождевых червей, встречающийся на большей части ее территории, в том числе в районах с экстремально низкими зимними температурами.
Рис. 1.
Распространение генетических линий дождевых червей двух подвидов Eisenia nordenskioldi (Шеховцов и др., 2017 с дополнениями). Линии E. n. nordenskioldi: 1 – 1-я, 2 – 3-я, 3 – 7-я, 4 – 9-я; линии E. n. pallida: 5 – 1-я, 6 – 3-я. Заштрихованные области – места сбора червей для экспериментов.
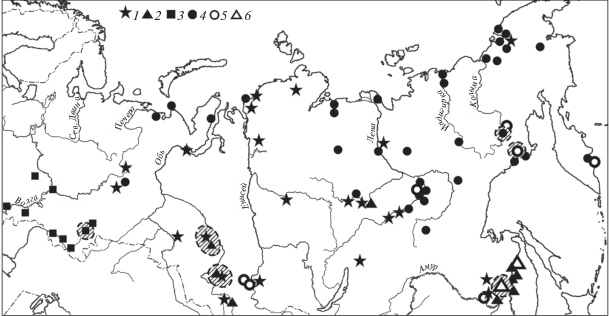
Известно, что E. nordenskioldi подразделялся на два подвида: E. n. nordenskioldi (Eisen, 1879) и Eisenia n. pallida Malevič, 1956. Молекулярно-генетические исследования выявили в пределах номинативного подвида не менее девяти генетических линий, а в пределах E. n. pallida – не менее пяти (Shekhovtsov et al., 2016; Шеховцов и др., 2017). Cудя по большому числу попарных нуклеотидных замен между линиями, достигающему в мтДНК 20%, а в ядерных генах 1.2–3.5%, по крайней мере часть из них может претендовать на видовой статус. Предпринятое нами исследование генетической структуры E. nordenskioldi на огромной территории, от Кольского п-ова до Чукотки, позволило наметить ареалы генетических линий пока только схематично (рис. 1). Но уже сейчас можно утверждать, что ряд линий аллопатричен (Шеховцов и др., 2017).
Аллопатрию могут определять различия адаптивных потенциалов, в частности разная холодоустойчивость (она же криорезистентность), т.е. способность переносить низкие отрицательные температуры, необходимая для существования в холодных регионах. По-видимому, ареал E. nordenskioldi полностью расположен в пределах территорий с низкими зимними температурами, однако в его разных географических частях условия в почве существенно различаются. Минимальные температуры почвы зависят не столько от положения регионов в системе географических координат, сколько от сочетания степени зимнего выхолаживания воздуха и мощности снежного покрова. В качестве подобного примера напомним, что на юге Сибири, в Забайкалье, выделен второй “полюс почвенного холода”, не уступающий по суровости первому “полюсу” в Оймяконе (Алфимов, Берман, 2006).
На ряде групп беспозвоночных показано, что таксономическая близость не гарантирует тождественности криорезистентности (Берман, Лейрих, 2017). Устойчивость к холоду червей разных линий одного подвида может оказаться неодинаковой, что будет свидетельствовать в пользу продвинутости внутривидовой дифференциации и может позволить рассматривать границы ареалов филогенетических линий как функцию зимней температуры почвы.
Для проверки этих предположений проведено определение холодоустойчивости червей четырех линий E. n. nordenskioldi (1, 3, 7 и 9-й) и двух линий E. n. pallida (1-й и 3-й) и сопоставление ее с зимними температурными условиями в почве в пределах предварительно пока выявленных границ ареалов (Шеховцов и др., 2017) (рис. 1).
МАТЕРИАЛЫ И МЕТОДЫ
Пункты сбора червей для экспериментов. E. n. nordenskioldi 1-й и 3-й линий собраны у с. Подгорное Томской обл. (57° с.ш., 82° в.д.) и в новосибирском Академгородке (55° с.ш., 82° в.д.), 7-й линии – в окрестностях д. Файзуллино Республики Башкортостан (52° с.ш., 58° в.д.), 9-й – в Магаданской обл. (стационар “Абориген”, 62° с.ш., 159° в.д.). E. n. pallida 1-й линии собран в пригороде Магадана (59° с.ш., 150° в.д.), 3-й – в 5 км от с. Казакевичево, в 30 км к югу от Хабаровска (48° с.ш., 1342° в.д.).
Черви 1-й и 3-й линий E. n. nordenskioldi в названных выше пунктах обитают совместно и нечетко различаются по окраске; генетическая принадлежность выживших при пороговых температурах особей была подтверждена после эксперимента.
Местообитания червей. Сбор животных проведен незадолго до начала зимовки (в конце августа–сентябре). В окрестностях с. Подгорное черви 1-й и 3-й линий номинативного подвида обычны (до 10 особей/м2) в лесах различных типов на коренном берегу и в поймах (кроме длительно заливаемых участков). Численность червей 1-й линии выше в лесах, численность червей 3-й линии – на опушках и участках с луговой растительностью. Весной и летом черви либо обитают под подстилкой на поверхности почвы, либо опускаются в горизонт 15–20 см, но после выпадения первого снега на указанных глубинах не найдены, т.е. места их зимовки, вероятно, находятся глубже 20 см.
В окрестностях новосибирского Академгородка черви обеих линий номинативного подвида встречаются в лесной подстилке и верхних слоях почвы в сосново-березово-осиновом лесу. Перед зимовкой (сентябрь–октябрь) черви 1-й линии в поверхностных горизонтах почвы отсутствуют – вероятное свидетельство того, что и на юге региона они уходят вглубь. Особи 3-й линии, напротив, встречаются вместе с Dendrobaena octaedra (Savigny, 1826) и Lumbricus rubellus Hoffmeister, 1843 в подстилке и верхних сантиметрах минерального горизонта.
На Южном Урале черви 7-й линии собраны в сухом березовом колке на границе со степью. В последних числах сентября они были сконцентрированы на глубине 10–15 см.
Черви 9-й линии занимают широкий набор местообитаний: окраины степных участков, всевозможные растительные группировки на водоупорной мерзлоте и без нее, от долин до высокогорий в лесной и тундровой зонах. Они не заселяют и стабильно сухие, и почвы с pH <5 (Берман, Мещерякова, 2013).
1-я линия E. n. pallida была собрана в окрестностях Магадана, где она встречалась исключительно на огородах и никогда не была обнаружена на невозделываемых землях (Берман, Мещерякова, 2013).
Под Хабаровском черви 3-й линии E. n. pallida были найдены на высокой пойме Амурской протоки р. Уссури, занятой вторичным широколиственным лесом. Они здесь обычны, иногда чрезвычайно многочисленны (>100 особей/м2). В конце сентября черви находятся в почве не глубже 10 см.
Акклимация, определение стратегии холодоустойчивости и порогов переносимых температур (ППТ) проведены в лаборатории биоценологии ИБПС ДВО РАН (Магадан) стандартными методами, разработанными для других видов дождевых червей (Берман, Лейрих, 1985; Лейрих, Мещерякова, 2015).
При подготовке к экспериментам (использовали только половозрелых особей) проводили акклимацию животных в вентилируемых пластиковых емкостях (200 мл) с почвой влажностью ~35% при следующих температурах: по 14 сут при 5 и 3°C, затем по 7 сут при 1, 0 и –1°С. Уже при температуре почвы ниже 5°C черви прекращают питаться; будучи потревожены, они вяло двигаются. Их покровы остаются влажными до 0°C и становятся сухими и жесткими при отрицательных температурах, а тело – почти плоским.
Для выяснения стратегии холодоустойчивости (перенесение отрицательных температур в переохлажденном или замороженном состоянии) измеряли температуру максимального переохлаждения (Тп) и соотносили ее с ППТ. Если ППТ оказывался ниже Тп, то делался вывод о способности червя выдерживать замерзание (Block, 1995).
Предельно переносимую температуру определяли путем оценки выживания особей при минимальной из тестируемых температур (табл. 1). Количество и значения температурных ступеней для разных линий различаются, так как перед началом экспериментов холодоустойчивость червей даже приблизительно не была известна. Ее приходилось “нащупывать”, начиная с небольших значений. Размер выборок при исследовании Тп составлял 10–12 особей, ППТ – от 6 особей на промежуточных ступенях до 67 особей на близких к предельно переносимым температурам (табл. 1). Выжившие в экспериментах черви после нагревания их до 10°C становились активными примерно через 2 сут.
Таблица 1.
Ступени определения предельно переносимых температур (ППТ, °C) дождевыми червями разных генетических линий двух подвидов Eisenia nordenskioldi
| Подвид | Линия | Температурные ступени***** |
|---|---|---|
| Eisenia nordenskioldi nordenskioldi | 1П* 1Н** 3П* 3Н** 7 9*** |
–5 (30), –8 (42), –10 (13), –12 (28), –13 (15) –5 (30), –8 (45), –10 (15), –12 (67), –13 (16) –5 (8), –8 (16), –10 (6), –12 (18), –13 (21) –5 (0), –8 (0), –10 (0), –12 (19), –13 (15) –8 (12), –12 (12), –20 (14), –23 (22), –24 (40), –26 (45) –12 (25), –14 (25), –16 (25), –17 (25), –19 (25), –21 (25), –22 (25), –23 (25), –25 (25), –28 (25), –30 (25), –34 (25) |
| Eisenia nordenskioldi pallida | 1**** 3 |
–10 (25), –12 (25), –15 (25), –18 (25), –20 (42), –25 (44),
28 (40), –30 (64) –5 (18), –7 (15), –10 (30), –12 (30) |
Примечание. * – 1П и 3П – с. Подгорное; ** – 1Н и 3Н – Новосибирск; *** – (Берман, Лейрих, 1985); **** – (Берман, Мещерякова, 2013); ***** – в скобках указан размер выборок (экз.); для табл. 1 и 2.
Материалы по холодоустойчивости 9-й линии номинативного подвида и 1-й линии подвида E. n. pallida были опубликованы ранее (Берман, Лейрих, 1985; Берман, Мещерякова, 2013).
Температурные условия в местах зимовки червей были определены в окрестностях с. Подгорное и вблизи с. Казакевичево с помощью автономных электронных регистраторов (модель DS1922L, точность ±0.5°С) на глубинах 2 и 10 см 2 раза в сутки. Регистраторы перед установкой были откалиброваны; отклонения от истинных значений не превышали 0.5°С.
Характеристика температурных условий обитания 9-й линии E. n. nordenskioldi в верховьях р. Колымы была опубликована ранее (Берман, Лейрих, 1985). Многолетние средние температуры воздуха и почвы на глубинах 20 и 40 см, а также максимальная высота снежного покрова приведены по Справочникам по климату СССР (1955, 1965, 1966, 1968).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Температуры максимального переохлаждения червей всех изученных линий близки, их средние значения –4.6…–3.1°C (табл. 2). ППТ, вопреки близости значений Тп, находятся в диапазоне ‒34…–10°C, т.е. весьма существенно различаются у разных линий (рис. 2). Судя по соотношению Тп и ППТ, черви всех рассмотренных нами линий обоих подвидов E. nordenskioldi могут длительно находиться при температурах, которые намного ниже точки переохлаждения, что свидетельствует о способности выдерживать замораживание. Напомним, что помимо E. nordenskioldi в фауне России (возможно, и в мировой) известны лишь четыре морозоустойчивых вида дождевых червей: Dendrobaena octaedra, Drawida ghilarovi Gates, 1969, Aporrectodea caliginosa (Savigny, 1826) и Eisenia sibirica Perel et Graphodatsky, 1984 (Берман и др., 2016).
Таблица 2.
Температура максимального переохлаждения (Тп, °C) червей разных генетических линий двух подвидов Eisenia nordenskioldi
| Подвид | Линия | n | Тп± SEM |
|---|---|---|---|
| E. nordenskioldi nordenskioldi | 1П* 1Н** |
10 | –3.6 ± 0.1 |
| 10 | –3.5 ± 0.3 | ||
| 3П* 3Н** |
10 | –3.4 ± 0.1 | |
| 10 | –3.1 ± 0.4 | ||
| 7 | 10 | –4.4 ± 0.4 | |
| 9*** | 48 | –3.5 ± 0.2 | |
| E. nordenskioldi pallida | 1**** | 11 | –4.6 ± 0.3 |
| 3 | 12 | –3.6 ± 0.2 |
Рис. 2.
Выживание червей четырех генетических линий Eisenia nordenskioldi nordenskioldi после трехсуточной экспозиции при разных температурах: 1–4 – 1-я, 3-я, 7-я, 9-я (Берман, Лейрих, 1985) линии соответственно.
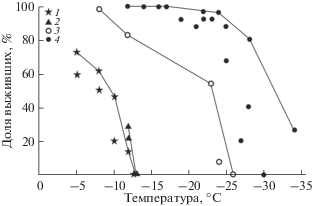
ППТ и распространение 9-й линии E. n. nordenskioldi. Эти черви среди исследованных наиболее устойчивы к отрицательным температурам (рис. 2): переносят до –34°C (Берман, Лейрих, 1985) имеют один из самых обширных и наиболее северных ареалов (Шеховцов и др., 2017) (рис. 1). Значительная его часть охватывает регионы с крайне жесткими зимними условиями: Чукотку, бассейны рек Колымы, Индигирки, Лены, Оленека, Анабара, п-ов Ямал и др. Судя по выявленной холодоустойчивости, даже в чрезвычайно холодном регионе, в верховьях р. Колымы, единственный тип непригодных местообитаний для зимовки этих червей – полностью бесснежные участки, на которых температура в слое 0–5 см может опускаться ниже –40°C при температуре воздуха –58°C (Алфимов, 1985). За пределами континентальных регионов Северо-Востока и Якутии E. n. nordenskioldi могут зимовать и на бесснежных местах (Тихомиров, 1937).
Обладая среди люмбрицид не только выдающейся криорезистентностью, но и устойчивостью к сухости и высокой кислотности почвы (Берман и др., 2002), черви 9-й линии между тем не обнаружены в наших обширных сборах (рис. 1) ни в бассейне Амура, ни в Средней Сибири, ни на большей части Западной Сибири. Однако очевидно, что отсутствие этих животных в относительно хорошо обследованных регионах юга Сибири и Дальнего Востока никак не может быть результатом недостаточной холодоустойчивости червей и их зимующих коконов.
ППТ и распространение 7-й линии E. n. nordenskioldi. Эта линия номинативного подвида – вторая по устойчивости к отрицательным температурам среди протестированных. Большая часть исследованной выборки выдержала охлаждение до –23°C (рис. 2). Черви 7-й линии найдены на юго-востоке европейской части России и на Южном Урале (рис. 1) – в республиках Татарстан и Башкортостан, Нижегородской, Оренбургской, Саратовской и Челябинской областях (Shekhovtsov et al., 2016) – и пока не известны в азиатской части России. В пределах их ареала черви других линий не встречены.
Названная территория находится в области изотерм средних температур воздуха в январе ‒12…–16°C (Алисов, 1956). Температура почвы на глубине 3 см под снежным покровом средней мощности не опускается в названных регионах ниже –17°C (Алфимов, 2005), а средние месячные температуры даже в самый холодный месяц составляют на глубине 20 см –6.4°C, на глубине 40 см –3.5°C (Справочник…, 1955, 1965).
Таким образом, холодоустойчивость червей 7‑й линии намного превышает зимний температурный фон почв региона. Имеющаяся криорезистентность могла бы позволить им распространиться как на запад и север, так и далеко на восток. Здесь непригодными для нее были бы лишь континентальные районы Якутии и северо-востока Азии. Однако этого не происходит, так как холодоустойчивость не единственный фактор, определяющий распространение генетических линий E. nordenskioldi. Отметим, что по мтДНК 7‑я линия родственна 9-й и объединяется с ней в один кластер (Shekhovtsov et al., 2016), что может обусловливать сродство высокой холодоустойчивости.
ППТ и распространение 1-й и 3-й линий E. n. nordenskioldi. Черви этих линий почти в 2 и 3 раза менее устойчивы к отрицательным температурам, чем черви 7-й и 9-й линий соответственно, но близки по этому признаку между собой. От четверти до пятой части животных из окрестностей с. Подгорное и г. Новосибирска перенесли охлаждение до –12°C, но –13°C не выдержали. Таким образом, –12°C следует считать ППТ (табл. 1, рис. 2).
Черви 1-й линии на исследованной нами территории (рис. 1) обнаружены в 25 точках, преимущественно в естественных местообитаниях. По молекулярно-генетическим данным можно выделить три заметно различающиеся группы: западную, куда входят черви со Среднего Урала, из Заполярья (о. Вайгач, пос. Лабытнанги, п-ов Таймыр), Новосибирской и Томской областей, и две восточные, найденные в Хакасии и Якутии (Shekhovtsov et al., 2018a). Западная группа, несмотря на значительный ареал, характеризуется небольшой генетической изменчивостью (~2% вариабельных позиций), что указывает, вероятно, на ее недавнее расселение; изменчивость восточных групп не ясна, так как исследованных точек пока мало.
Эта картина сильно отличается от таковой для 7-й и 9-й линий, имеющих значительное “внутрилинейное” разнообразие (Shekhovtsov et al., 2018b; Шеховцов и др., 2017). 1-я линия, по-видимому, расширила свой ареал до громадных размеров сравнительно недавно, в голоцене.
Черви 3-й линии распространены менее широко. Они найдены на юге Западной Сибири (Томская и Новосибирская области, Республика Алтай – пос. Артыбаш), на р. Марха (приток Вилюя) близ устья, в Еврейской автономной области, в нижнем течении Амура (Шеховцов и др., 2017).
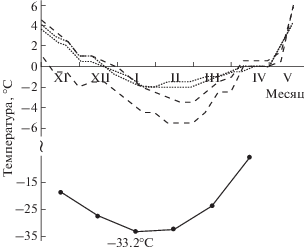
ППТ червей –12°C достаточна для благополучной зимовки почти повсеместно на юго-востоке Западной Сибири и в лесных среднегорьях Алтая с учетом относительно высокого фона минимальных температур поверхностного горизонта почв (Алфимов и др., 2012; Берман и др., 2016). Вблизи с. Подгорное и г. Томска в подавляющей части 12 обследованных местообитаний, где были найдены черви, минимумы температуры в почве на глубине 10 см не опускались ниже –6°C, лишь в двух местообитаниях они составляли –12…–13°C (Алфимов и др., 2012). В окрестностях Хабаровска зимняя температура в первых 10 см почвы также была значительно выше –12°C (рис. 3).
Рис. 3.
Ход средних месячных температур воздуха (сплошная кривая) и минимальных за декаду температур почвы на глубине 2 см (штриховые кривые) и 10 см (точечные кривые) с осени 2013 г. по весну 2014 г. в двух местообитаниях червей 3-й генетической линии Eisenia nordenskioldi pallida в Большехехцирском заповеднике. Температура воздуха приведена по данным метеостанции Хабаровск (удаление 27 км).
Черви 1-й и 3-й линий номинативного подвида достигают крупных размеров, что обычно служит признаком люмбрицид-норников. Вероятно, представителей какой-то из этих линий (или обеих) имела в виду Перель (Перель, 1982) и вслед за ней ряд других авторов, упоминая особенно крупных октаплоидных особей E. n. nordenskioldi, встречающихся в тундрах Таймыра, на Салаирском кряже, в Московской и Курской областях (Стриганова, Тиунов, 1994; Всеволодова-Перель, Булатова, 2008). “Гигантскую для червей морфу, превышающую обычную втрое…” отмечает Ганин (Ганин, 1997, с. 25) для Дальнего Востока.
Весьма вероятно, что именно характерной для норных червей зимовкой в глубине почвы можно объяснить обитание червей 1-й линии в таких суровых местностях, как бассейн верхнего течения р. Бурея (окрестности пос. Ургал), где в январе средняя температура воздуха –31.1°C при максимальной высоте снежного покрова лишь 29 см. Но вечной мерзлоты в этом регионе нет, что, вероятно, позволяет червям уходить на зиму в глубокие горизонты.
Обнаружение 1-й и 3-й линий номинативного подвида на территориях с холодной зимой и вечной мерзлотой, таких как Юго-Западная Якутия или Центральная Якутия, на первый взгляд, кажется странным. Действительно, в окрестностях г. Олекминск, например, средняя температура воздуха самого холодного месяца –33.5°C. Но минимальные температуры почвы на этой территории, как и в междуречье Лены и Вилюя, могут быть существенно повышены в так называемых таликовых зонах (Геокриология…, 1989) – участках с незамерзающими и находящимися близко к поверхности подземными водами (прирусловый сток рек, берега непромерзающих озер, выходы глубинных вод и т.д.). В пределах таликовых зон отмечены наиболее высокие зимние температуры почвы на глубине 20 см (–11…–5.5°C). Даже на глубине 2–3 см они были не ниже –12°C (Berman et al., 2016), а средние многолетние на этой глубине не опускаются в регионе ниже –10.4°C (Справочник…, 1966). Таким образом, черви на исследованных территориях Юго-Западной Якутии могут зимовать в комфортных для них условиях.
Однако черви 1-й линии обнаружены в низовьях Оби, в районе пос. Жиганск (на Лене), устья р. Марха (притока Вилюя), оз. Собачьего у Норильска, на п-ове Таймыр (рис. 1). Климат этих регионов крайне жесткий, мощность сезонно-талого слоя порой едва превышает первые десятки сантиметров. К примеру, на метеостанции Диксон в 2002–2010 гг. минимальная температура на глубине 20 см опускалась до –21.8°C (вытяжные термометры) при высоте снега в феврале–марте 20–25 см (сайт ВНИИГМИ-МЦД http://meteo.ru/data).
Если исходить из того, что холодоустойчивость одних и тех же видов дождевых червей по ареалу одинакова (Мещерякова, Берман, 2014), то объяснить присутствие 1-й линии, обладающей невысокой холодоустойчивостью, в названных в предыдущем абзаце суровых местностях трудно. Однако отсутствие географической изменчивости холодоустойчивости показано на небольшом материале и нуждается в более тщательных доказательствах.
В настоящее время биология и экология червей 1-й линии совершенно не изучены. Представления о ней без каких-либо оснований перенесены с 9-й линии или даже с E. n. nordenskioldi неопределенных линий. Для объективных суждений необходимо выяснить как минимум стандартные экологические характеристики червей 1-й линии, включающие в себя биотопическую приуроченность, температурные условия зимовки, холодоустойчивость коконов и червей северных популяций и пр. Напомним, что у некоторых видов дождевых червей разница холодоустойчивости червей и коконов достигает 30–40°C (Мещерякова, Берман, 2014). Важны и другие биологические показатели этих червей (например, возможность партеногенеза, скорости развития), способствующие формированию малочисленных локальных популяций. Такие микропопуляции могут быть приурочены к особо теплым зимой местам (микрорельеф, положение относительно направления преобладающих ветров и др.) и сохраняться в них длительное время без обмена генами с другими популяциями.
ППТ и распространение 1-й и 3-й линий E. n. pallida. Черви 1-й линии из пригородов Магадана обладают поразительно высокой криорезистентностью –28°C (Берман, Мещерякова, 2013) (рис. 4). Они обнаружены здесь только на огородах и не расселяются в природные местообитания, тогда как черви 9-й линии номинативного подвида в антропогенных биотопах никогда не были встречены. На зимовку животные уходят в глубь обрабатываемой почвы до 60 см (Берман, Мещерякова, 2013), где температуры выше переносимых ими. Так, минимальные температуры почвы на глубинах 2, 20 и 40 см в трех точках огорода в окрестностях г. Магадана осенью 2011–зимой 2012 гг. составляли –10…–13.5°С, –8…–11.5°С и –6.5…–8.5°С соответственно. Черви 1-й линии E. n. pallida заселяют также огороды в пос. Сеймчан, находящемся в континентальной части региона, где температуры января примерно на 10°C ниже, чем в Магадане. Только в антропогенно измененных биотопах черви 1-й линии обнаружены на Камчатке, в Амурской обл. и Республике Хакасия. Удаленность мест находок (рис. 1) этих червей при незначительной генетической изменчивости и приуроченность к антропогенным местообитаниям (в отличие от 1-й линии E. n. nordenskioldi) свидетельствуют о недавнем расселении при участии человека.
Рис. 4.
Выживание червей 1-й (1) (Берман, Мещерякова, 2013) и 3-й (2) генетических линий Eisenia nordenskioldi pallida после трехсуточной экспозиции при разных температурах.
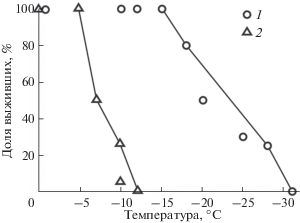
Черви 3-й линии E. n. pallida, найденные пока только в Хабаровском крае (Шеховцов и др., 2017) (рис. 1), переносят температуру не ниже –10°C. При названной температуре в выборке из 12 особей выжило три (25%), в выборке из 18 особей только одна (<6%) (рис. 4). Таким образом, эти черви по значениям ППТ близки к животным 1-й и 3-й линий E. n. nordenskioldi.
Под Хабаровском непосредственно в местах сбора животных температура за зиму в горизонте 0–2 см не опускалась ниже –5.5°C, на глубине 10 см – ниже –2°C, т.е. не достигала пределов их криорезистентности. Соотношение почвенных температур и холодоустойчивости червей свидетельствует о том, что на исследованной пойменной террасе Уссури популяция благополучна. Но температурные условия и вне пойменных пространств не хуже. Так, на междуречье Амура и его притока Тунгуски у пос. Николаевка минимумы за зиму 2007/2008 г. на глубине 2 см не были ниже –11.8°C, а на глубине 25 см – ниже –8.2°C (наши неопубликованные данные).
ППТ и ареалы генетических линий червей – синтез. Исходя из изложенного выше, казалось бы, прослеживается зависимость положения и величины ареалов от холодоустойчивости червей. Действительно, самая устойчивая 9-я линия (–34°С) чрезвычайно широко распространена в самых холодных регионах Севера и Якутии. Очень низкие значения ППТ имеют также 1-я линия E. n. pallida (–28°С) и 7-я линия номинативного подвида (–23°C). Первая из названных обитает на окультуренных землях под Магаданом, но могла бы с такой холодоустойчивостью быть повсеместной не только на побережье, но и в континентальных районах, чего не происходит. Удивительно, что 7-я линия, обладая явно избыточной и не адаптивной по отношению к температурному режиму местообитаний на юго-востоке Восточной Европы холодоустойчивостью, в Азии не обнаружена. Криорезистентность этих червей настолько велика, что свидетельствует о полной независимости их географического распространения от температурных условий зимовки в пределах всей территории, заселенной E. nordenskioldi.
Напротив, у червей 3-й линии E. n. pallida холодоустойчивость минимальная из исследованных (–10°С). Она лишь незначительно выше (–12°С) у 1-й и 3-й линий номинативного подвида. Казалось бы, распространение этих трех линий должно быть ограничено зимними почвенными температурами, однако полученная картина связи положения ареалов и холодоустойчивости червей значительно сложнее.
Отсутствие 9-й линии на юге, как и 7-й линии на севере Европы и где бы то ни было в Азии и, напротив, крайне широкое распространение 1-й линии номинативного подвида с ее незначительной холодоустойчивостью в Сибири, в том числе в Субарктике, – все это примеры несоответствия экофизиологических возможностей червей и занимаемых ими ареалов. Таким образом, характер ареала только части линий может быть объяснен их резистентностью к отрицательным температурам.
ЗАКЛЮЧЕНИЕ
Способность E. n. nordenskioldi переносить температуры –34°C, ранее определенная у червей из бассейна верховий Колымы, как выяснилось, относится не к виду в целом, а исключительно к его 9-й генетической линии. Среди четырех исследованных линий подвида E. n. nordenskioldi и двух линий E. n. pallida есть как умеренно устойчивые (переносящие температуры –10…–12°C), так и выживающие при значительно более низких температурах (–28…–34°C). У трех линий (1-й и 3-й номинативного подвида и 3-й подвида pallida) из шести значения холодоустойчивости близки, у трех других резко различаются. Полученный результат свидетельствует о внутривидовой экофизиологической дифференциации при сохранении общей для E. nordenskioldi черты – способности переносить длительное замораживание.
Характер ареала 7-й и 9-й линий номинативного подвида определяется, вероятно, палеогеографическими причинами или иными, не связанными с температурой факторами, поскольку их холодоустойчивость не может лимитировать современное распространение. Черви трех линий обоих подвидов умеренно устойчивы к отрицательным температурам, что находится в соответствии с положением ареала на юге Сибири.
Наиболее сложно интерпретировать несоответствие между небольшой криорезистентностью червей 1-й линии номинативного подвида (–12°С), измеренной у животных на юге Западной Сибири, и распространением их на севере в регионах с вечной мерзлотой. Уточнить это можно путем проведения анализа, прежде всего биотопической приуроченности, температурных условий зимовки и холодоустойчивости червей и коконов из северных популяций.
Авторы признательны ${\text{Т }}{\text{.С }}{\text{.}}\,{\text{В с е в о л о д о й - П е р е л ь }}$ за уточнение видовой принадлежности червей, А.Б. Бабенко, И.Н. Корниенко, А.А. Котову, О.Л. Макаровой, Н.А. Петруниной, Н.Н. Тридриху, С.П. Харитонову, З.А. Янченко и С.А. Ярошевой за сбор и доставку червей из труднодоступных районов; заместителю директора Большехехцирского заповедника Р.С. Андроновой за организационную помощь.
Работа выполнена при финансовой поддержке РФФИ (грант 16–04–00082).
Список литературы
Алисов Б.П. Климат СССР. М.: Изд-во МГУ, 1956. 129 с.
Алфимов А.В. Термический режим верхних слоев почвы в основных экосистемах пояса редколесий бассейна Верхней Колымы // Пояс редколесий верховий Колымы (район строительства Колымской ГЭС). Владивосток: ДВНЦ АН СССР, 1985. С. 9–29.
Алфимов А.В. Распределение минимальных температур в поверхностном слое почвы под снегом в Северной Евразии // Почвоведение. 2005. № 4. С. 438–445.
Алфимов А.В., Берман Д.И. Два полюса холода под снежным покровом Евразии // Природа. 2006. № 6. С. 27–33.
Алфимов А.В., Берман Д.И., Булахова Н.А. Зимние температурные условия в корнеобитаемом слое почв в Сибири и на северо-востоке Азии // Вестн. СВНЦ ДВО РАН. 2012. № 3. С. 10–18.
Берман Д.И., Лейрих А.Н. О способности дождевого червя Eisenia nordenskioldi (Eisen) (Lumbricidae, Oligochaeta) переносить отрицательные температуры // Докл. АН СССР. 1985. Т. 285. № 5. С. 1258–1261.
Берман Д.И., Лейрих А.Н. Холодоустойчивость массовых почвообитающих беспозвоночных животных северо-востока Азии. 2. Холодоустойчивость как адаптация к климату // Зоол. журн. 2017. Т. 96. № 10. С. 1119–1131.
Берман Д.И., Мещерякова Е.Н. Ареалы и холодоустойчивость двух подвидов дождевого червя (Eisenia nordenskioldi, Lumbricidae, Oligochaeta) // Зоол. журн. 2013. Т. 92. № 7. С. 771–780.
Берман Д.И., Булахова Н.А., Мещерякова Е.Н. Холодоустойчивость и ареал дождевого червя Eisenia sibirica (Oligochaeta, Lumbricidae) // Сиб. экол. журн. 2016. № 1. С. 56–64
Берман Д.И., Лейрих А.Н., Алфимов А.В. Об устойчивости дождевого червя, Eisenia nordenskioldi (Oligochaeta, Lumbricidae), к экстремально низкой влажности почвы на северо-востоке Азии // Зоол. журн. 2002. Т. 81. № 11. С. 1308–1318.
Всеволодова-Перель Т.С., Булатова Н.Ш. Полиплоидные расы дождевых червей (Lumbricidae, Oligochaeta), распространенные в пределах Восточно-Европейской равнины и в Сибири // Изв. РАН. Сер. биол. 2008. № 4. С. 448–452.
Ганин Г.Н. Почвенные животные Уссурийского края. Владивосток; Хабаровск: Дальнаука, 1997. 160 с.
Геокриология СССР. М.: Недра, 1989. 516 с.
Лейрих А.Н., Мещерякова Е.Н. К методам исследования холодоустойчивости беспозвоночных животных // Зоол. журн. 2015. Т. 94. № 8. С. 972–984.
Мещерякова Е.Н., Берман Д.И. Устойчивость к отрицательным температурам и географическое распространение дождевых червей (Oligochaeta, Lumbricidae, Moniligastridae) // Зоол. журн. 2014. Т. 93. № 1. С. 53–64.
Перель Т.С. Распространение и закономерности распределения дождевых червей фауны СССР. М.: Наука, 1979. 272 с.
Перель Т.С. Географические особенности размножения дождевых червей сем. Lumbricidae (Oligochaeta) // Журн. общ. биологии. 1982. Т. 43. № 5. С. 649–658.
Справочник по климату СССР. Вып. 9, 12, 24, 25, 33. Ч. II, IV Л.: Гидрометеоиздат, 1955, 1965, 1966, 1968.
Стриганова Б.Р., Тиунов А.В. Пищевая стратегия дождевых червей в горных почвах на многолетней мерзлоте // Изв. РАН. Сер. биол. 1994. № 3. С. 435–445.
Тихомиров Б.А. Об условиях обитания дождевых червей в тундровых почвах // Природа. 1937. № 5. С. 52–58.
Шеховцов С.В., Берман Д.И., Голованова Е.В., Пельтек С.Е. Генетическая структура дождевого червя Eisenia nordenskioldi (Lumbricidae) // Вавиловский журн. генетики и селекции. 2017. Т. 21. № 5. С. 588–595.
Berman D.I., Bulakhova N.A., Alfimov A.V., Meshcheryakova E.N. How the most northern lizard, Zootoca vivipara, overwinters in Siberia // Polar Biol. 2016. V. 39. P. 2411–2425.
Block W. Insects and freezing // Sci. Progress. 1995. V. 78. № 4. P. 349–372.
King R.A., Tibble A.L., Symondson W.O.C. Opening a can of worms: unprecedented sympatric cryptic diversity within British lumbricid earthworms // Mol. Ecol. 2008. V. 17. P. 4684–4698.
Novo M., Almodóvar A., Díaz-Cosín D.J. High genetic divergence of hormogastrid earthworms (Annelida, Oligochaeta) in the central Iberian Peninsula: evolutionary and demographic implications // Zool. Scripta. 2009. V. 38. P. 537–552.
Porco D., Decaëns T., Deharveng L., James S.W., Skarżyński D., Erséus C., Butt K.R., Richard B., Hebert P.D.N. Biological invasions in soil: DNA barcoding as a monitoring tool in a multiple taxa survey targeting European earthworms and springtails in North America // Biol. Invasions. 2013. V. 15. P. 899–910.
Shekhovtsov S.V., Golovanova E.V., Peltek S.E. Mitochondrial DNA variation in Eisenia n. nordenskioldi (Lumbricidae) in Europe and Southern Urals // Mitochondrial DNA. 2016. V. 27. P. 4643–4645.
Shekhovtsov S.V., Berman D.I., Bulakhova N.A., Makarova O.L., Peltek S.E. Phylogeography of earthworms from high latitudes of Eurasia // Acta Zool. Acad. Sci. Hung. 2018a. V. 64. P. 369–382.
Shekhovtsov S.V., Berman D.I., Bulakhova N.A., Vinokurov N.N., Peltek S.E. Phylogeography of Eisenia nordenskioldi nordenskioldi (Lumbricidae, Oligochaeta) from the north of Asia // Polar Biol. 2018b. V. 41. P. 237–247.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая