

Известия РАН. Серия биологическая, 2019, № 5, стр. 478-484
Раковинные амебы гондвано-тропической группы и юго-восточная граница Палеарктики
А. А. Бобров 1, *, С. Шимано 2, Я. Цинь 3, Ц. Ян 4
1 Московский государственный университет им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, стр. 1, Россия
2 Университет Хосэй
102-8160 Фуджими, Хийода, Токио, Япония
3 Китайский Университет Геонаук
430074 Ухань, Китай
4 Институт городской среды, Китайская Академия Наук
361021 Сямынь, Китай
* E-mail: anatoly-bobrov@yandex.ru
Поступила в редакцию 04.03.2019
После доработки 15.04.2019
Принята к публикации 15.04.2019
Аннотация
Проанализировано географическое распространение раковинных амеб родов Hoogenraadia и Planhoogenraadia в Ориентальной биогеографической области и в Приморском крае Дальнего Востока. Установлено, что на территории Сихотэ-Алинского заповедника и островов Хонсю и Хоккайдо, входящих в юго-восточную часть Палеарктики, встречаются раковинные амебы из гондвано-тропической группы. С учетом географического распространения этой группы простейших предложено уточнить положение юго-восточной границы Палеарктики в дальневосточном регионе.
Современная биогеографическая картина мира опирается в основном на данные о распространении нескольких групп растений (покрытосеменные, голосеменные, папоротникообразные) и животных (амфибии, рептилии, птицы, млекопитающие) (Воронов и др., 2002). Почти полтора века назад Уоллес (Wallace, 1876) разделил сушу на шесть зоогеографических регионов (рис. 1а), в значительной степени совпадающих с границами тектонических континентальных плит. Многие современные исследователи согласны с мнением Уоллеса (Ruesta, 2013; Whittaker et al., 2013). Фитогеографическая картина мира (Тахтаджян, 1978) также во многом совпадает с представлениями Уоллеса.
Рис. 1.
Зоогеографические области Уоллеса (Wallece, 1876) (а); биогеографическая карта мира, основанная на нумерической классификации (Huggett, 2004) (б).
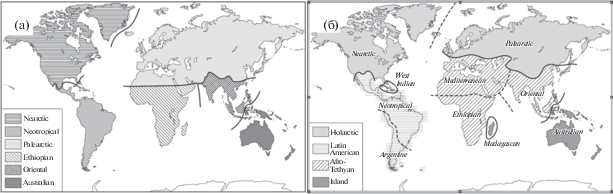
Однако немало публикаций, основанных на филогенетических связях животных и растений (Smith, 1983; Huggett, 2004; Holt et al., 2012), вносят значимые изменения в привычную биогеографическую картину мира (рис. 1б). Как правило, в них представлены результаты статистического анализа распределения фаун и флор разных регионов Земли. В частности, для количественного определения изменений филогенетического состава амфибий, птиц и млекопитающих были использованы показатели бета-разнообразия (Holt et al., 2012). Анализ их комбинированных значений выявил в общей сложности 20 зоогеографических регионов, вошедших в 11 крупных областей. Одним из ключевых несоответствий классификаций Холта и Уоллеса стали существенные различия в обозначении южной границы Палеарктики. Еще бóльшие изменения предложены Смитом (Smith, 1983), который выделил Средиземноморскую область и значительно перенес на север широту юго-восточной границы Палеарктики. Проблемы и преимущества количественного подхода к биогеографическим построениям подробно обсуждены Кентом (Kent, 2006). Синтетический подход к проблеме вернул южную границу Палеарктики практически к границам Уоллеса, к Северной Африке (Cox et al., 2001). К близкому выводу пришли и другие авторы (Procheş, Ramdhani, 2012). На сегодня существуют две точки зрения на мировую биогеографию, соответствующие представлениям Уоллеса и Смита.
Частичные несоответствия между пространственными моделями распространения разных видов и филогенетическими отношениями таксонов некоторые авторы предлагают устранить с помощью метода дихотомического анализа (Ebach, Parenti, 2015) с учетом климатических и тектонических факторов (Ficetola et al., 2017). Но зоогеографические регионы Уоллеса – результат синтетического анализа распределения флоры и фауны – кажутся не менее обоснованными, чем в его время, несмотря на новые сведения о распределении биоты, изменения теоретических представлений и использование сложных аналитических методов.
Климат как фактор выделения биогеографических областей наиболее важен для проведения субрегиональных границ, а тектонические особенности территорий – для выделения границ между регионами (Ficetola et al., 2017). Проблему границ и переходных зон между регионами как наиболее сложную некоторые исследователи предлагают решать методами сетевой теории (Vilhena, Antonelli, 2014). Первичный этап решения этой проблемы – поиск индикаторных с точки зрения биогеографии видов. Один из проблемных и интереснейших биогеографических регионов Палеарктической области – ее юго-восточная часть или западная прибрежно-континентальная тихоокеанская зона (Приморский край). На юге эта область граничит с Китаем в зоне маньчжуро-японских смешанных лесов, на севере – с восточно-сибирской тайгой; на востоке (через Японское море) находятся маньчжуро-японские смешанные леса (о. Хоккайдо) и ориентальные листопадные леса (Udvardi, 1975). Флора Приморья считается в значительной мере производной от тропической флоры, часть ее имеет третичный возраст 65 млн. лет (Алехин и др., 1961; Колесов, 1980) и по схеме геоботанического районирования (Колесников, 1955) относится к восточно-азиатской хвойно-широколиственной области или к манчьжуро-японским смешанным лесам (Udvardi, 1975). Более подробно флора этого региона и ее история были проанализированы ранее (Коломыц, 2015).
Биогеографическая картина мира значительно меняется в зависимости от того, какие организмы используются в качестве модельных групп (Holt et al., 2012). Раковинные амебы – свободноживущие простейшие с преимущественно анемохорным распространением, а значительная часть их видов – космополиты. Однако еще Эренберг (Ehrenberg, 1871, цит. по Bonnet, 1983) показал, что в составе раковинных амеб крупных географических областей существуют фаунистические различия. Позднее эта точка зрения получила подтверждение в работах Хейниса (Heinis, 1914) и Дефляндра (Deflandre, 1928, 1936) о распространении некоторых видов из родов Nebela и Arcella. Практически во всех более поздних публикациях (Jung, 1942; Бассин, 1945; Hoogenraad, De Groot, 1979; Bonnet, 1983, 1984; Корганова, 1987, 1994; Smith, Wilkinson, 1987; Bobrov, 2001; Mitchell, Meisterfeld, 2005; Foissner, 2006; Smith et al., 2008; Heger et al., 2011) были отмечены различия в южной и северной фауне раковинных амеб и подтверждено существование видов с локальным географическим распространением. Значительный вклад в анализ географического распространения раковинных амеб Ориентальной области внес Боннэ (Bonnet, 1977, 1980а, б, 1981, 1982, 1985, 1987).
Большинство почвообитающих видов раковинных амеб Палеарктики относятся к эврихорной группе простейших. Особое положение занимают виды из гондвано-тропической группы раковинных амеб родов Hoogenraadia и Planhoogenraadia, обнаруженные в почвах Приморья (Бобров, 2000; Bobrov, 2001). Эти два рода представляют особый интерес, поскольку их распространение, по-видимому, ограничено относительно низкими широтами. Они мало изучены и не фигурируют в основных руководствах по идентификации раковинных амеб. Этим обусловлена актуальность обзорных публикаций о находках этих видов для экологов и биогеографов, работающих с пробами из различных географических регионов. Традиционно считается, что виды родов Hoogenraadia и Planhoogenradia эндемичны для тропической и субтропической зон Гондваны (Smith et al., 2008).
Цель данного исследования – анализ распространения в юго-восточной части Палеарктики раковинных амеб родов Hoogenraadia и Planhoogenraadia.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование основано на анализе собственных и опубликованных данных по распространению родов Hoogenraadia и Planhoogenraadia на территории, включающей в себя восточное Тихоокеанское побережье Палеарктики (от Приморья до Командорских о-вов и тундр Магаданской обл.) (Bobrov, 2012), а также Индомалайскую (далее в тексте Ориентальную) и частично Океаническую биогеографические области.
Биогеографическое районирование в данной статье дано согласно Классификации биогеографических провинций, принятой ЮНЕСКО в 1975 г. (Udvardi, 1975).
Были исследованы почвенные пробы: во-первых, из Палеарктической области – Россия, Приморье (Сихотэ-Алинский биосферный заповедник, провинция маньчжуро-японских смешанных лесов) (Bobrov, 2001); Япония – о. Хонсю (японские вечнозеленые или субтропические леса) (Shimano et al., 2014); Китай – провинции Фуцзянь (китайские субтропические леса) и Хубэй (ориентальные листопадные леса); во-вторых, из Ориентальной области – Вьетнам, заповедник Катьен (муссонные полулистопадные леса); Зондские о-ва, о. Флорес, острова Вознесения и Святой Елены (тропические леса).
В ранее опубликованных работах (табл. 1) встречаемость представителей родов Hoogenraadia и Planhoogenraadia рассматривается во-первых, в Ориентальной области – Таиланд (индонезийские дождевые леса, тропические лиственные леса); Непал (индо-гангские муссонные леса, субтропический горный пояс); Индонезия (Целебесская провинция, влажные вечнозеленые леса); Филиппины (Филиппинская провинция, влажные тропические леса); во-вторых в Океанической области – о. Аполима, Полинезия; Папуа Новая Гвинея (Папуасская провинция, влажные тропические леса).
Таблица 1.
Географическое распространение и основные типы местообитаний видов Hoogenraadia и Planhoogenraadia в районе восточного Тихоокеанского побережья
| Вид | Регион | Местообитания |
|---|---|---|
| H. africana | Китай (Qin et al., 2011) | Сфагновые и водные |
| H. asiatica | Китай (Wang, Min, 1987; Yang et al., 2005) | Четвертичные отложения, водные местообитания |
| H. cryptostoma | Япония, острова Хонсю и Якушима* | Моховые и сфагновые |
| H. humicola | Филиппины, Индонезия, Непал (Bonnet, 1977, 1980a, b, 1981, 1992); о. Аполима (Корганова, 1994); Вьетнам (Bobrov et al., 2010); Россия, Приморье (Bobrov, 2001); Зондские о-ва, о. Флорес* | Богатые органикой почвы, подстилки субтропических почв, влажные место- обитания (глеевые почвы, мюллевый тип гумуса) |
| P. acuta | Китай (Фуцзянь)* | КП |
| P. alta | Индонезия, Таиланд (Bonnet, 1987, 1992) | КП |
| P. bonneti | Таиланд (Todorov, Golemansky, 1999); Китай (Хубэй)* | Эпифитные мхи на деревьях, КП |
| P. cantabrica | Индонезия, Таиланд (Bonnet, 1985, 1987, 1992) | КП |
| P. daurica | Россия, Приморье (Bobrov, 2001); Япония, Токио (Shimano et al., 2014); Китай (Фуцзянь, Хубэй)* | КП, бурые почвы |
| P. elegans | Индонезия, Новая Гвинея, Непал, Таиланд, Филиппины (Bonnet, 1977, 1980a, b; 1981; 1982, 1987, 1992); Китай (Фуцзянь, Хубэй)* | КП, вулканические почвы, почвы тропических и субтропических лесов |
| P. gracilis | Таиланд (Bonnet, 1987) | КП |
| P. media | Индонезия, Филиппины, Непал, (Bonnet, 1977, 1980a, b, 1992); Россия, Приморье (Bobrov, 2001) | КП, почвы тропических и субтропических лесов |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализ распространения видов Hoogenraadia и Planhoogenraadia (табл. 1) показал, что они характерны для почв Ориентальной и Океанической областей и также встречаются в почвах влажных тропических широколиственных лесов. По нашим исследованиям (в Японии, Вьетнаме, Китае, на о. Флорес), местообитания этих видов входят в субтропическую и тропическую зоны, включая острова Хонсю и Хоккайдо. В Голарктической области они были найдены в маньчжуро-японских смешанных лесах (рис. 2) – это самая северная точка обнаружения этих гондвано-тропических видов (Bobrov, 2001; Bobrov et al., 2014).
Рис. 2.
Биогеографическая карта Палеарктики и Ориентальной области (Huggett, 2004). 1 – граница Голарктики по Уоллесу (Wallece, 1876); 2 – граница настоящего исследования, севернее которой виды гондвано-тропической группы не найдены; 3 – находки видов рода Planhoogenraadia; 4 – находки видов рода Hoogenraadia; 5 – районы, где виды гондвано-тропической группы не найдены.
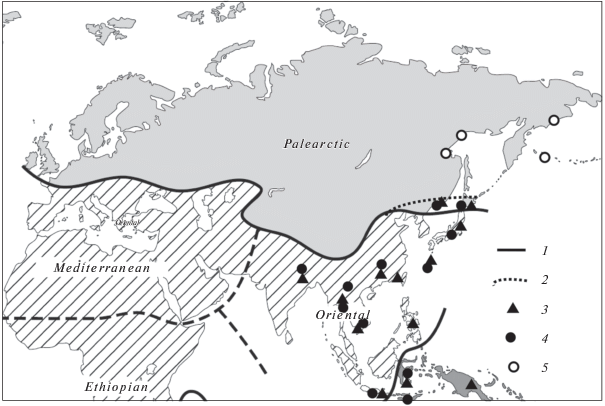
В состав сообществ Ориентальной биогеографической области входят виды из тропической группировки Hoogenraadia africana, H. asiatica, H. cryptostoma, H. humicola, Planhoogenraadia acuta, P. alta, P. bonneti, P. cantabrica, P. daurica, P. elegans, P. gracilis, P. media (табл. 1).
Такие виды из гондвано-тропической группы, как H. humicola, P. daurica и P. media, были найдены в поверхностных пробах темноцветной и бурой лесных почв с околонейтральной рН (Bobrov, 2001). Все виды рода Planhoogenraadia заселяют рендзины и рендзиноподобные почвы или их дериваты, такие как бурые лесные почвы, развивающиеся в условиях тропического и субтропического климатов и богатые органическим веществом (табл. 1). В равной мере это относится и к H. humicola. В этом его отличие от других видов рода Hoogenraadia, например H. africana и H. cryptostoma, которые были найдены в сфагновых африканских топях (Gauthier-Lievre, Thomas, 1958).
Ряд по частоте встречаемости видов в регионах Ориентальной области, включая Приморье, следующий: P. elegans (9 находок), H. humicola (8), P. media (5), P. cantabrica (3), P. daurica (3), P. alta (2), P. bonneti (2), H. cryptostoma (2), P. acuta, P. gracilis, H. africana, H. asiatica (по 1 находке). Виды найдены чаще всего в карбонатных почвах широколиственных лесов (Bonnet, 1984). Исключение составляют H. cryptostoma, H. africana и H. asiatica, населяющие сфагновые, моховые и водные местообитания (табл. 1).
Находки наиболее часто встречающегося вида P. elegans сосредоточены в южной части Ориентальной области и субтропиках Центрального Китая; по-видимому, это теплолюбивый вид. Это относится и к другим видам обоих родов, за исключением H. humicola, P. daurica и P. media, которые найдены в Приморье (Bobrov, 2001) и выдерживают относительно низкие зимние температуры (до –10°С). Типы местообитаний этих видов – карбонатные почвы и влажные почвы широколиственных лесов, богатые органикой.
Карта распространения видов Hoogenraadia и Planhoogenraadia (рис. 2) дает лишь общее представление о том, какие регионы Ориентальной области они населяют. Существующие лакуны – результат недостаточной изученности мировой почвенной протистофауны в целом, удаленность, а нередко и труднодоступность отдельных районов (Foissner, 2006; Smith et al., 2008). Полученные данные дополняют результаты более ранних исследований раковинных амеб Дальнего Востока (Bobrov, 2001; Bobrov et al., 2014).
На территории Дальнего Востока и сопредельных регионов границы основных фаунистических подобластей Палеарктики сходятся и образуют переходную зону между Палеарктикой и Ориентальной областью. Уникальность этого региона отмечали уже давно, и о придании ему самостоятельного биогеографического статуса писали Н.А. Северцов, П.П. Сушкин, П.П. Семенов-Тян-Шанский (Колесов, 1980). По своему происхождению часть фауны этого региона связана с северо-восточным Китаем и северной частью Корейского п-ова, а также с островами Японского архипелага. Поэтому фауна, в первую очередь фауна о. Хонсю, обнаруживает филогенетические связи с фауной Приморья и южных провинций Китая. Эти связи подтверждаются и находками видов Hoogenraadia и Planhoogenraadia в Приморье, на островах Хонсю, Хоккайдо, Якушима и в Китае (рис. 2).
Выше северной границы Приморья, или “линии Арсеньева”, фауна представлена широко распространенными видами таежной Сибири. Та же закономерность характерна и для протистофауны (Bobrov, 2001). В исследованных пробах почв с юго-западного и северного Приохотья, с островов Командорского архипелага, с Камчатки и Сахалина раковинные амебы из гондвано-тропической группы не были обнаружены (Бобров, 1999; Bobrov, 2012). Это можно объяснить отсутствием смешанных широколиственных лесов маньчжурского типа в Приохотье, Хабаровском крае, на Камчатке, северном Сахалине (Bobrov, 2012). Единственные практически не исследованные районы Ангарской обл. – четвертичные рефугиумы и широколиственные леса южного Сахалина. Находки в них видов из Ориентальной области могли бы еще раз подтвердить важность протозоологических исследований для детальных биогеографических исследований.
В синтетической схеме биогеографических областей (Huggett, 2004) юго-восточная граница Палеарктики проходит несколько выше Корейского п-ова и разделяет острова Хоккайдо и Хонсю Японского архипелага. На уровне субрегионов о. Хонсю разделен на две почти равные части – северную и южную. Северная часть занята ориентальными листопадными лесами, занимающими также и значительную часть континентального Китая, на которой отмечены виды гондвано-тропической группы. Южная часть занята маньчжуро-японскими смешанными лесами, которые на континенте граничат с Приморским краем; в ней также найдены виды родов Hoogenraadia и Planhoogenraadia. В последние годы появляются публикации, которые предлагают уточнение биогеографических регионов Китая, включая риентированную по направлению юг–север палеоарктико-восточную границу (He et al., 2017). Процесс ревизии и уточнения границ биогеографических регионов и субрегионов неизбежен и связан с накоплением новых данных о пространственном распределении флоры и фауны.
По данным протозоологического анализа в связи с находками гондвано-тропических видов раковинных амеб граница между Палеарктической и Ориентальной областями в континентальной и островной частях может быть поднята в более северные широты, включая Сихотэ-Алинский заповедник и острова Хонсю и Хоккайдо (рис. 2). Этот анализ может быть также учтен при региональном биогеографическом картировании.
Исследование проведено при финансовой поддержке РФФИ (гранты 15-29-02518 и 16-04-00451/17), Японского общества содействия науке JSPS KAKENHI (грант № 15H02858), Национального природного научного фонда Китая (гранты № 41502167, № 31672312) и китайского проекта The Belt & Road (№ DL2017ZDGZ (WH) 031).
Список литературы
Алехин В.В., Кудряшов Л.В., Говорухин В.С. География растений с основами ботаники. М.: Гос. уч.-пед. изд. Мин. прос. РСФСР, 1961. 532 с.
Бассин Ф.Н. Географическое распространение раковинных корненожек: Дис. докт. биол. наук. М.: МГУ,1944. 449 с.
Бобров А.А. Эколого-географические закономерности распространения и структуры сообществ раковинных амеб (Protozoa, Testacea): Автореф. дис. докт. биол. наук. М.: МГУ, 1999. 47 с.
Бобров А.А. Planhoogenraadia daurica Bobrov, новый вид раковинной амебы (Protozoa: Testacea) из Приморья (Дальний Восток) // Зоол. журн. 2000. Т. 79. № 1. С. 108–110.
Воронов А.Г., Дроздов Н.Н., Криволуцкий Д.А., Мяло Е.Г. Биогеография с основами экологии. М.: Высш. шк., 2002. 391 с.
Колесников Б.П. Очерк растительности Дальнего Востока. Хабаровск: Хабар. кн. изд-во, 1955. 104 с.
Колесов А.М. Зоогеография Дальнего Востока. М.: Мысль, 1980. 254 с.
Коломыц Э.Г. Эволюционная экология бореальных лесов на Тихоокеанском мегаэкотоне Северной Евразии // Самарская Лука: проблемы региональной и глобальной экологии. 2015. Т. 24. № 3. С. 5–139.
Корганова Г.А. Apolimia – новый род почвенных раковинных амеб с островов Западное Самоа и замечания по составу семейства Hyalospheniidae (Arcellinidae) // Зоол. журн. 1987. Т. 66. Вып. 2. С. 165–169.
Корганова Г.А. Раковинные амебы (Protozoa) в островных экосистемах // Животное население островов Юго-Западной Океании. М.: Наука, 1994. С. 184–209.
Тахтаджян А.Л. Флористические области Земли. Л.: Наука, 1978. 247 с.
Bobrov A., Qin Y., Wilkinson D.M. Latitudinal diversity gradients in free-living microorganisms – Hoogenraadia, a key genus in testate amoebae biogeography // Acta Protozool. 2014. V. 54. P. 1–8.
Bobrov A.A. Finding of the Tropical Group Testate Amoebae (Protozoa: Testacea) at the Far East (Sikhote Alin Reserve) // Biol. Bull. 2001. V. 28. № 4. P. 401–407.
Bobrov A.A. Pacific transect: Testate amoebae of the Commander Islands to Papua New Guinea // 6th Int. Symp. on Testate Amoebae, Xiamen, China, 15–18 Oct. 2012. Program & Abstracts. P. 53.
Bobrov A.A., Mazei Y.A., Tiunov A.V. Testate Amoebae of a Monsoon Tropical Forest of South Vietnam // Acta Protozool. 2010. V. 49. P. 311–325.
Bonnet L. Le peuplement Thécamoébiens des sols du Nepal et son intérêt biogéographique // Bull. Soc. Hist. Nat. Toulouse. 1977. V. 113. F. 3–4. P. 331–348.
Bonnet L. Quelques aspects de la faune thécamoébienne des sols de la Nouvelle-Guinée (Papouasie) // Bull. Soc. Hist. Nat. Toulouse. 1980a. V. 116. F. 3–4. P. 266–276.
Bonnet L. Faunistique et biogéographie des Thécamoébiens. VI. Thécamoébiens de quelques sols des Phillippines // Bull. Soc. Hist. Nat. Toulouse. 1980b. V. 116. F. 3–4. P. 277–282.
Bonnet L. L’intérêt biogéographique du peuplement Thécamoébien des sols du Népal // Paléogéographie et biogéographie de l’Himalaya et du Sous-Continant Indien. Paris: C.N.R.S., 1981. P. 53–60.
Bonnet L. Faunistique et biogéographie des Thécamoébiens. VII. Thécamoébiens de quelques sols forestiers de Thaïlande // Bull. Soc. Hist. Nat. Toulouse. 1982. V. 118. P. 245–262.
Bonnet L. Intérêt biogéographie et paleogéographie des Thécamoébiens des sols // Extrait des Annales de la station biol. de Besse-en-Chandesse. 1983. P. 298–334.
Bonnet L. Nouvelles données sur le genre Planhoogenraadia (Thécamoebiens) // Bull. Soc. Hist. Nat. Toulouse. 1984. V. 120. P. 117–122.
Bonnet L. Faunistique et biogéographie des Thécamoébiens. VIII. Thécamoébiens de quelques sols d’Indonésie // Bull. Soc. Hist. Nat. Toulouse. 1985. V. 121. P. 13–15.
Bonnet L. Faunistique et biogéographie des Thécamoébiens. IX. Thécamoébiens de quelques sols forestiers de Thaïlande (2ème partie) // Bull. Soc. Hist. Nat. Toulouse. 1987. V. 123. P. 115–121.
Bonnet L. Faunistique et biogéographie des Thécamoébiens. X. Thécamoébiens de quelques sols d’Indonésie (2ème partie) // Bull. Soc. Hist. Nat. Toulouse. 1992. V. 128. P. 17–21.
Cox C.B., Cottage F., Close B., Surrey A. The biogeographic regions reconsidered // J. Biogeogr. 2001. V. 28. P. 511–523.
Deflandre G. Le genre Arcella Ehrenberg // Arch. Protist. 1928. Bd 64. H. 1–2. S. 152–287.
Deflandre G. Etude monographie sur le genre Nebela Leidy // Ann. Protistol. 1936. V. 5. P. 201–286.
Ebach M.C., Parenti L.R. The dichotomy of the modern bioregionalization revival // J. Biogeogr. 2015. V. 42. № 10. P. 1801–1808.
Ficetola G.F., Mazel F., Thuiller W. Global determinants of zoogeographical boundaries Gentile Francesco. Suppl. information // Nat. Ecol. Evol. Macmillan Publ. Ltd. 2017. V. 1. № 0089. P. 1–12.
Foissner W. Biogeography and dispersal of micro-organisms: a review emphasising protists // Acta Protozool. 2006. V. 45. P. 111–136.
Gauthier-Lievre L., Thomas R. Les genres Difflugia, Pentagonia, Magrebia et Hoogenraadia (Rhizopodes Testacea) en Afrique // Arch. Protist. 1958. Bd 103. H. 1–2. S. 241–370.
He J., Kreft H., Gao E., Wang Z., Jiang H. Patterns and drivers of zoogeographical regions of terrestrial vertebrates in China // J. Biogeogr. 2017. V. 44. P. 1172–1184.
Heger T.J., Lara E., Mitchell E.A.D. Arcellinida testate amoebae (Amoebozoa: Arcellinida): model of organisms for assessing microbial biogeography // Ed. Fontaneto D. Biogeography of Microscopic Organisms: Everything Small Everywhere? Cambridge: Cambr. Univ. Press., 2011. P. 111–129.
Heinis Fr. Die Moosfauna Columbiens / Eds Fuhrmann O., Mayor E. Voyage d’Exploration scientifique en Colombie // Mem. Soc. Neuchateloise Sci. Nat. 1914. V. 5. P. 675–730.
Holt B.G., Lessard J.-P., Borregaard M.K., Fritz S.A., Araújo M.B., Dimitrov D., Fabre P.-H., Graham C.H., Graves G.R., Jønsson K.A., Nogués-Bravo D., Wang Z., Whittaker R.J., Fjeldså J., Rahbek C. An Update of Wallace’s Zoogeographic Regions of the World // Sciencexpress. 2012. P. 1–10. http://www.sciencemag.org/content/early/recent//1126/science. 1228282.
Hoogenraad H.R., De Groot A.A. Die geographische Verbreitung der süsswasser Rhizopoden // Hydrobiol. Bull. (Amsterdam). 1979. V. 13. № 2–3. P. 152–171.
Huggett R.J. Fundamentals of Biogeography Front Cover Psychology Press. Routledge: Science, 2004. 439 p.
Jung W. Südchilenische Thecamoben // Arch. Protist. 1942. Bd 95. P. 253–365.
Kent M. Numerical classification and ordination methods in biogeography // Progr. Phys. Geogr. 2006. V. 30. № 3. P. 399–408.
Mitchell E.A.D., Meisterfeld R. Taxonomic confusion blurs the debate on cosmopolitanism versus local endemism of free-living protists // Protist. 2005. V. 156. P. 263–267.
Procheş S., Ramdhani S. The World’s Zoogeographical Regions Confirmed by Cross-Taxon Analyses // BioSci. 2012. V. 62. № 3. P. 260–270.
Qin Y., Xie S., Smith H.G., Swindles G.T., Gu Y. Diversity, distribution and biogeography of testate amoebae in China: Implications for ecological studies in Asia // Eur. J. Protistol. 2011. V. 47. P. 1–9.
Ruesta M., Rodriguez M., Hawkins B.A. Identifying global zoogeographical regions: lessons from Wallace // J. Biogeogr. 2013. V. 40. № 12. P. 2215–2225.
Shimano S., Bobrov A., Mazei Yu. Testate Amoebae of the Imperial Palace, Tokyo // Mem. Nat. Mus. Nat. Sci. Tokyo. 2014. V. 50. P. 21–28.
Smith Ch.H. A system of world mammal faunal regions. I. Logical and statistical derivation of the regions // J. Biogeogr. 1983. V. 10. P. 455–466.
Smith G., Wilkinson D.M. Biogeography of testate rhizopods in the southern temperate and Antarctic zones // Coll. sur les ecosystemes terrestres subantarctiques. Paimont., CNFRA, 1987. № 58. P. 83–96.
Smith H.G., Bobrov A., Lara E. Diversity and biogeography of testate amoebae // Biodiv. Conserv. 2008. V. 17. № 2. P. 345–363.
Todorov M., Golemansky V. Planhoogenraadia bonneti sp.n. and Centropyxis thailandica sp.n. (Rhizopoda: Testacea), two new testaceans from Thailand // Acta Protozool. 1999. V. 38. P. 255–261.
Udvardi M.D.F. A Classification of the Biogeographical Provinces of the World // Intl. Union Conserv. Nat. and Natural Res. (Occasional Paper 18). Morges, Switzerland, 1975. 49 p.
Vilhena D.A., Antonelli A. Beyond similarity: A network approach for identifying and delimiting biogeographical regions // Nat. Comm. 2014. P. 1–9. www.nature.com/naturecommunications.
Wallace A.R. The Geographical Distribution of Animals. Cambridge: Cambr. Univ. Press, 1876. https://doi.org/10.1017/CBO9781139097109
Wang P., Min Q. On the paleoenvironmental significance of thecamoebians in the Quaternary studies of China // Micropalaeontol. Sinica. 1987. V. 4. P. 345–349.
Whittaker R., Riddle B.R., Hawkins B.A., Ladle R.J. The geographical distribution of life and the problem of regionalization: 100 years after Alfred Russel Wallace // J. Biogeogr. 2013. V. 40. P. 2209–2214.
Yang J., Zhang W., Feng W., Shen Y. Testate Amoebae (Protozoa: Rhizopoda) from Northwest Yannan, China // J. Freshwater Ecol. 2005. V. 20. № 3. P. 583–590.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая