

Известия РАН. Серия биологическая, 2019, № 5, стр. 466-477
Дождевые черви (Oligochaeta, Lumbricidae) в тундрах Восточной Европы
О. Л. Макарова 1, *, А. А. Колесникова 2
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
2 Институт биологии Коми научного центра УрО РАН
167982 Сыктывкар, ул. Коммунистическая, 28, Россия
* E-mail: ol_makarova@mail.ru
Поступила в редакцию 04.03.2019
После доработки 15.04.2019
Принята к публикации 15.04.2019
Аннотация
Установлено, что в тундровых ландшафтах Восточной Европы встречаются восточноевропейско-сибирский Eisenia nordenskioldi nordenskioldi и космополитные Lumbricus rubellus (единично вблизи границы леса) и Dendrobaena octaedra. В равнинных тундрах Палеарктики D. octaedra не найден восточнее Тазовского п-ова, а E. n. nordenskioldi – западнее Печорской губы, причем оба вида совместно обитают в Арктике только на территории Ненецкого и Ямало-Ненецкого автономных округов. Приведены новые отметки дождевых червей на севере Восточной Европы. Показана значительная биотопическая пластичность D. octaedra и E. n. nordenskioldi в тундровом ландшафте и отмечено, что холодоустойчивый подстилочно-почвенный E. n. nordenskioldi предпочитает открытые местообитания (кустарничковые тундры, луга, марши), а менее холодоустойчивый в фазе червя подстилочный D. octaedra чаще встречается под покровом кустарников или в криволесье. Предположено, что E. n. nordenskioldi был более широко распространен в Восточной Европе во время последнего оледенения.
В российской части Арктики дождевые черви встречаются повсеместно, кроме самых северных архипелагов (Земля Франца-Иосифа, Северная Земля, о-ва Де-Лонга). Их численность в зональных тундрах обычно низка, 2–19 экз./м2 (Чернов, 1978; Перель, 1979; Боескоров, 2004; Таскаева и др., 2015; Конакова и др., 2017), но локально может достигать 40–150 экз./м2 в хорошо дренированной черноземовидной почве под луговой разнотравно-злаковой растительностью на речных склонах (Чернов, 1961, 1962, 1978). Видовой состав дождевых червей в зональных тундрах беден (как правило, 1–2 вида), в некоторых исследованиях на территории подзоны южных тундр они вообще не были найдены (Козловская, 1955; Перель, 1979) или встречались очень локально в биотопах, наиболее прогреваемых летом и/или хорошо защищенных снегом от зимних морозов (Тихомиров, 1937; Стебаев, 1959; Перель, 1979; Боескоров, 2004). Именно малочисленность червей в тундрах затрудняет изучение биотопической приуроченности видов.
Первые сообщения о дождевых червях в восточно-европейских тундрах связаны со шведскими арктическими экспедициями 1870–1876 гг., когда был описан вид Eisenia nordenskioldi (Eisen, 1879) (в том числе с о. Вайгач), а Dendrobaena octaedra (Savigny, 1826) указан для о. Южный Новой Земли. Позднее был отмечен D. octaedra с Мурманского берега и о. Колгуев (материалы Зоологического музея, СПб.) (Michaelsen, 1910), что подтвердилось его повторным обнаружением в двух районах острова (Тихомиров, 1937) и на берегах Дальнезеленецкой губы (Стриганова, 1973). В сводке 1979 г. Т.С. Перель указала D. octaedra с п-ова Канин, а затем добавила отметки D. octaedra из Большеземельской тундры и с Югорского п-ова, а E. nordenskioldi – из Амдермы (Всеволодова-Перель, 1988). В последние десятилетия находки дождевых червей в восточно-европейских тундрах были единичными (Таскаева, Мелехина, 2011; Таскаева и др., 2015; Конакова и др., 2017).
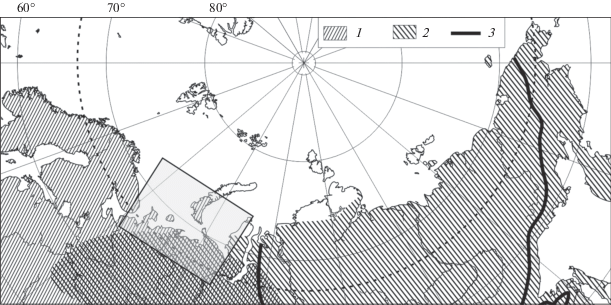
В Арктике сосуществование этих наиболее холодоустойчивых, морозостойких видов Lumbricidae наблюдается только на территории Большеземельской тундры, Ямала и Гыдана (Перель, 1979; Всеволодова-Перель, 1988; Стриганова, Порядина, 2005). Находки в равнинных тундрах Старого Света космополитного подстилочного D. octaedra ограничены на востоке Тазовским п-овом, а восточноевропейско-сибирский подстилочно-почвенный E. n. nordenskioldi не отмечен в тундрах западнее р. Печоры (рис. 1).
Рис. 1.
Области распространения дождевых червей Dendrobaena octaedra (1) и Eisenia nordenskioldi nordenskioldi (2) в Северной Евразии (Всеволодова-Перель, 1988; Берман и др., 2002; Стриганова, Порядина, 2005; отметки в табл. 1); 3 – изотерма средней температуры воздуха в январе –28°С.
Цель работы – анализ распространения видов дождевых червей на территории восточно-европейских тундр и их биотопических предпочтений, а также проверка рабочей гипотезы: в тундровых районах, где D. octaedra и E. nordenskioldi сосуществуют, их биотопические преферендумы не совпадают.
МАТЕРИАЛЫ И МЕТОДЫ
Мы располагаем материалами дождевых червей с о. Долгий, Печорское море (сборы О.Л. Макаровой, 2004 г.), где их собирали вручную, выгоняли в эклекторах (155 проб 5 × 5 см на всю глубину подстилки и слоя почвы 0–5 см) и учитывали в почвенных ловушках (диаметром 65 мм с 0.5%‑ным формалином), установленных в 23 биотопах (3050 лов.-сут.). Всего было найдено 38 экз. (24 взрослых).
В коллекции Института биологии Республики Коми имеются сборы А.А. Колесниковой с коллегами (2001–2017 гг.) из восьми районов Ненецкого АО и четырех районов Республики Коми (табл. 1). В каждом районе в 2–5 типичных биотопах разбирали по 10 почвенных проб с площади 25 × 25 см (подстилка + слой почвы 0–5 см). Всего в 38 биотопах было обнаружено 86 экз. червей.
Таблица 1.
Новые находки дождевых червей на территории восточно-европейских тундр (по материалам ИБ Коми НЦ УрО РАН, ИПЭЭ РАН, ФИЦКИА РАН, ИБПС ДВО РАН)
| Пункт | Район | Координаты | Сборщики | Время сбора | Отмечены | |||
|---|---|---|---|---|---|---|---|---|
| с.ш. | в.д. | D. o. | E. n. n. | L. r. | ||||
| 1. Шойна | п-ов Канин | 67°87′ | 44°19′ | А.Б. Бабенко, А.А. Нехаева | VII/2017 | + | – | – |
| 2. Гольцовое | о. Колгуев | 69°21′ | 49°17′ | А.Б. Бабенко | VIII/1994 | + | – | – |
| 3. Верхняя Песчанка | То же | 69°24′ | 48°84′ | С.Б. Розенфельд | VII–VIII/2011 | + | – | – |
| 4. Средняя Песчанка | » | 69°15′ | 49°27′ | То же | VIII/2011 | + | – | – |
| 5. Малые Кармакулы | Новая Земля | 72°38′ | 52°71′ | В.М. Спицин | VII/2015 | + | – | – |
| 6. Безымянная | То же | 72°85′ | 53°71′ | То же | VII/2017 | + | – | – |
| 7. Еренной Нос | Дельта р. Печоры | 68°33′ | 53°83′ | А.А. Колесникова | VIII/2001 | + | + | – |
| 8. Костяной Нос | То же | 68°33′ | 53°63′ | То же | VIII/2001 | + | + | – |
| 9. Ловецкий | » | 68°34′ | 53°94′ | » | VIII/2001 | + | + | – |
| 10. Кашин | » | 68°24′ | 53°88′ | » | VIII/2001 | + | + | – |
| 11. Ячей | Большеземельская тундра | 68°09′ | 54°78′ | О.Л. Макарова, М.С. Бизин | VII/2015 | + | + | – |
| 12. Шапкина | То же | 67°17′ | 53°98′ | А.А. Колесникова | VIII/2005 | + | + | – |
| 13. Матюй-Саля | » | 68°49′ | 57°23′ | О.Л. Макарова, М.С. Бизин | VIII/2015 | + | + | – |
| 14. Колва | » | 67°70′ | 58°43′ | А.А. Колесникова | VII–VIII/1999 | – | + | – |
| 15. Море-Ю | » | 68°29′ | 59°94′ | О.Л. Макарова, М.С. Бизин | VIII/2015 | + | + | – |
| 16. Большая Роговая | » | 67°02′ | 61°63′ | А.А. Колесникова | VIII/2010 | + | + | – |
| 17. Воркута | » | 67°51′ | 64°02′ | То же | VII/2017 | + | + | + |
| 18. Хребтовый | Полярный Урал | 67°19′ | 65°03′ | » | VII/2008 | – | + | – |
| 19. Пага-Ты | То же | 66°21′ | 62°45′ | » | VI–VII/2007 | + | + | + |
| 20. Лохорта | » | 66°59′ | 61°39′ | » | VIII/2010 | + | + | + |
| 21. Долгий вост. | о. Долгий | 69°26′ | 59°19′ | О.Л. Макарова | VIII/2004 | + | + | – |
| 22. Спрудже | о. Вайгач | 69°84′ | 59°61′ | А.А. Колесникова | VII–VIII/2004 | – | + | – |
| 23. Варнек | То же | 69°71′ | 60°11′ | А.Б. Бабенко | VII/2015 | – | + | – |
В трех центральных районах о. Колгуев дождевые черви были собраны вручную (2 экз.) и с помощью эклекторов (19 экз., 36 проб) (А.Б. Бабенко, 1994 г.; С.Б. Розенфельд, 2011 г.).
В июле 2017 г. на п-ове Канин были собраны 14 экз. червей (ручная разборка А.Б. Бабенко и А.А. Нехаевой); в 64 эклекторных пробах черви не были обнаружены. В июле 2018 г. в окрестностях пос. Амдерма было собрано 11 половозрелых червей (М.С. Бизин и Б.Д. Ефейкин).
В ходе экспедиции Арктического плавучего университета в 2015 г. были собраны 3 экз. на о. Вайгач и 2 экз. на о. Южный Новой Земли; 1 неполовозрелая особь была найдена в 50 эклекторных пробах 2017 г. на этом же острове в бух. Безымянной (табл. 1).
В июле–августе 2015 г. в ходе комплексной экспедиции ИПЭЭ РАН на приморских маршах Печорского моря (НАО-2015) были проведены учеты (табл. 2) методами почвенных ловушек, прикопок, сифтования подстилки (по 5 проб с площади 25 × 25 см в биотопе), почвенных (по 5 проб 25 × 25 × 5 см в биотопе) и эклекторных (по 5–8 проб в биотопе) проб. В низовьях р. Ячей (Болванская губа) были обследованы основные сообщества маршевого комплекса (три уровня), торфяник, болото, ерник, ивняк и приречные луга. На м. Матюй-Саля (Паханческая губа) были взяты пробы на всех пяти выделяемых по растительности уровнях приморских маршей и в девяти основных типах биотопов коренного берега. В низовьях р. Море-Ю (Хайпудырская губа) обследовали приморские марши на пяти уровнях и 11 иных биотопов. Всего было обнаружено 84 экз. дождевых червей, из которых 44 экз. взрослых.
Таблица 2.
Cборы дождевых червей в Большеземельской тундре (экспедиция НАО-2015, июль–август 2015 г.)
| Район работ | Число биотопов | Число проб | ПЛ | Число собранных червей | ||
|---|---|---|---|---|---|---|
| ЭВ | Сиф | РР | ||||
| Болванская губа, низовье р. Ячей | 9 | 46 | 40 | 20 | 350 | 16 |
| Паханческая губа, м. Матюй-Саля | 14 | 65 | 40 | 10 | 700 | 51 |
| Хайпудырская губа, низовье р. Море-Ю | 16 | 25 | 20 | 18 | 800 | 17 |
| Всего | 39 | 136 | 100 | 48 | 1850 | 84 |
Определение материала проведено по книге Всеволодовой-Перель (1997), автор которой проверила определения червей с о. Долгий (сборы 2004 г.) и из Большеземельской тундры (2015 г.).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Дождевые черви были впервые обнаружены в 23 пунктах севера Восточной Европы (табл. 1). Все известные на сегодня местонахождения (40) в этом регионе приведены на рис. 2. Обнаружены три вида Lumbricidae – D. octaedra, E. nordenskioldi nordenskioldi и Lumbricus rubellus Hoffmeister, 1843. Последний вид отмечен только в горных тундрах Полярного Урала и в окрестностях г. Воркуты (южный предел тундровой зоны). Находка L. rubellus на севере Большеземельской тундры, в бассейне р. Черной (Конакова и др., 2017) требует подтверждения. Во всех значительных выборках дождевых червей присутствуют как взрослые, так и неполовозрелые особи.
Рис. 2.
Находки видов дождевых червей на севере Восточной Европы: 23 из 40 точек – новые отметки (табл. 1), остальные – Eisen, 1879; Козловская, 1957; Малевич, 1976; Всеволодова-Перель, 1988; Берман и др., 2002; Таскаева, Мелехина, 2011; Мещерякова, Берман, 2014; Таскаева и др., 2015; Конакова и др., 2017. 1 – Dendrobaena octaedra, 2 – Eisenia nordenskioldi nordenskioldi, 3 – оба вида.
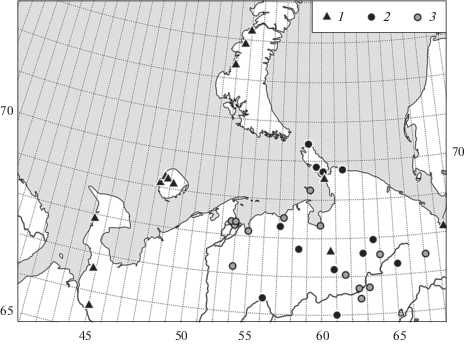
Встречаемость D. octaedra и E. n. nordenskioldi в регионе приблизительно одинакова (28 и 27 пунктов соответственно, но лишь в 15 пунктах (38%) виды обитают совместно). На западе региона отмечен только D. octaedra (Новая Земля, о. Колгуев, п-ов Канин), на северо-востоке – только E. n. nordenskioldi (о. Вайгач, Амдерма).
На о. Долгий дождевые черви обнаружены в 19 из 23 изученных биотопов, в 5 биотопах – только E. n. nordenskioldi или D. octaedra, в 14 биотопах – оба вида. Оба вида встречались вдоль всего гидрологического профиля, в том числе в дренированных щебнистых тундрах, ивняках, торфяниках, на лугах и в различных низовых болотах.
На о. Колгуев в трех районах его центральной части был обнаружен только D. octaedra. В эклекторных учетах обилие его ювенильных особей в тундровых местообитаниях составило 160–240 экз./м2.
В коллекции Института биологии Коми НЦ УрО РАН, в материале из 12 районов (табл. 1, пункты 7–10, 12, 14, 16–20, 22), были обнаружены D. octaedra, E. n. nordenskioldi и L. rubellus. С учетом опубликованных данных (Таскаева и др., 2015; Конакова и др., 2017) диапазон численности дождевых червей в 38 изученных биотопах составил 2–11 экз./м2. D. octaedra и E. n. nordenskioldi были по-разному распределены в биотопах разного типа (табл. 3). В открытых биотопах (тундры, луга) были значительно выше встречаемость и численность E. n. nordenskioldi. В зарослях кустарников и деревьев чаще встречался D. octaedra, который был здесь достоверно обильнее, чем в тундровых биотопах.
Таблица 3.
Встречаемость (Вс, %) и диапазон значений численности (ДЧ, экз./м2, в скобках – средние значения) дождевых червей в сообществах разных типов на территории восточно-европейских тундр (равнинных и горных) и лесотундры (по материалам коллекции Института биологии Коми НЦ УрО РАН, 1999–2017)
| Вид | Тундры*, луга (20 биотопов) | Заросли кустарников и деревьев** (18 биотопов) | ||
|---|---|---|---|---|
| Вс | ДЧ | Вс | ДЧ | |
| Dendrobaena octaedra | 25.0 | 0–4.8 (0.6)а | 66.7 | 0–9.6 (2.3)в |
| Eisenia nordenskioldi | 65.0 | 0–11.6 (2.2)б | 44.4 | 0–3.2 (0.9)г |
На побережье Большеземельской тундры в Болванской губе черви были найдены в 4 из 9 изученных биотопов, в Паханческой – в 9 из 14, в Хайпудырской – в 7 из 16 (в целом в 20 из 39 биотопов). Во всех районах на приморских маршах дождевые черви заселяют только верхние уровни, где их численность, включая молодь, по эклекторным учетам варьирует от 80 до 400 экз./м2 (по результатам сифтования – от 3 до 10 экз./м2). Сходный диапазон численности ювенильных особей отмечен в наиболее прогреваемых, хорошо дренированных склоновых биотопах: на разнотравном лугу – 320, в дриаднике на сопке – 240 экз./м2 (соответствующие результаты сифтования – 29 и 10 экз./м2). В почве низких маршей, болот и торфяников черви не обнаружены.
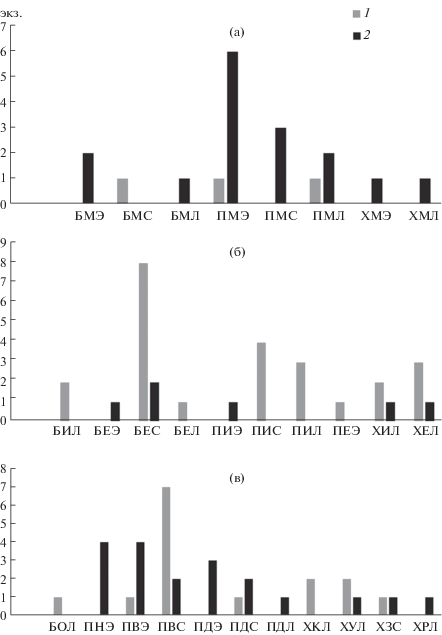
Только в 11 из 20 биотопов обитают оба вида червей. Для анализа биотопических предпочтений видов мы сгруппировали местообитания, где были обнаружены черви, в три комплекса (рис. 3): приморские марши верхних уровней, заросли крупных кустарников и открытые сообщества с разнотравьем (главным образом различные луга или кустарничково-разнотравные ассоциации). На маршах многочисленнее был E. n. nordenskioldi, а под покровом кустарников преобладал D. octaedra (тест Манна–Уитни: P < 0.05). В открытых сообществах с разнотравьем и/или кустарничками соотношение видов варьировало незакономерно.
Рис. 3.
Число особей Dendrobaena octaedra (1) и Eisenia nordenskioldi nordenskioldi (2), полученных разными методами в различных районах и биотопах Большеземельской тундры (табл. 2). а – морские марши, б – кустарниковые заросли, в – луга и разнотравно-кустарничковые сообщества. Первая буква кода обозначает район: Б – Болванская губа, П – Паханческая губа, Х – Хайпудырская губа. Вторая буква кода обозначает биотоп: В – разнотравный луг на восточном склоне, Д – дриадник, Е – ерник, З – влажный луг с преобладанием злаков, И – ивняк, К – кустарничково-разнотравное сообщество на песчаном гребне, М – марш, Н – нивальное разнотравно-кустарничковое сообщество, О – разнотравный луг на речном обрыве, Р – разнотравный пойменный луг, У – сухой разнотравный луг на террасе. Третья буква кода обозначает метод сбора: Э – эклекторная выгонка, С – сифтование, Л – почвенные ловушки и прикопки.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Разнообразие и обилие люмбрицид. С учетом новых данных в Арктике обитают четыре вида дождевых червей. В полярных пустынях они не найдены (Чернов, 1978; наши неопубликованные сведения о Земле Франца-Иосифа и Северной Земле), а в тундровой зоне отмечены космополитные D. octaedra и L. rubellus, восточноевропейско-сибирский E. n. nordenskioldi и сибирский E. atlavinyteae Perel et Graphodatsky, 1984 (Всеволодова-Перель, 1988, 1997; Всеволодова-Перель, Лейрих, 2014; наши неопубликованные данные). Находки в тундрах червей L. rubellus, погибающих уже при –2…–3°С (Мещерякова, Берман, 2014), фрагментарны (Ude, 1902; Конакова и др., 2017; новые данные), а некоторые требуют подтверждения. Остальные три вида, способные к обитанию на вечной мерзлоте, характеризуются высокой холодоустойчивостью на всех стадиях жизненного цикла (Берман, Мещерякова, 2013; Мещерякова, Берман, 2014), чем определяется возможность их развития в течение двух и более лет.
Обращает на себя внимание отсутствие эндогейных видов. D. octaedra обитает преимущественно в подстилке, три остальных вида принадлежат к почвенно-подстилочной жизненной форме (Перель, 1979). Вероятно, это определяется в первую очередь малой толщиной и низкими температурами деятельного слоя почвы в тундрах даже при островном характере вечной мерзлоты.
В арктических ландшафтах Старого Света с запада на восток наблюдается постепенное снижение разнообразия дождевых червей. Так, в естественных тундрах на севере Скандинавии обитают 4 вида (Terhivuo, 1988), в тундрах Восточной Европы и Западной Сибири – по 3 (редко 4) вида (Всеволодова-Перель, 1997; Стриганова, Порядина, 2005; новые данные), в среднесибирском секторе Арктики – 2 вида (Всеволодова-Перель, 1997; Рыбалов, 2002), в восточно-сибирском секторе – 1 вид (Всеволодова-Перель, 1988; Боескоров, 2004), что, по-видимому, определяется условиями зимовки (Мещерякова, Берман, 2014). Находки Lumbricus terrestris Linnaeus, 1758 в Большеземельской тундре (Конакова и др., 2017) и Aporrectodea caliginosa (Savigny, 1826) в тундрах Гыдана (Стриганова, Порядина, 2005) требуют подтверждения. На всей территории Якутии (с экстремально холодными зимами) обитает только E. n. nordenskioldi (Белимов и др., 1981; Убугунова и др., 2007; Всеволодова-Перель, Лейрих, 2014), обладающий рекордными показателями холодоустойчивости коконов и червей (Берман, Лейрих, 1985; Мещерякова, Берман, 2014). В американском секторе Арктики Lumbricidae отсутствуют (Berman, Marusik, 1994; Reynolds, 1995; Берман и др., 2002б).
В результате ревизии всех литературных данных и новых материалов (40 пунктов, рис. 2) в тундрах Восточной Европы обнаружено только три вида дождевых червей – E. nordenskioldi, D. octaedra и L. rubellus (редок). Все виды чрезвычайно политопны и часто доминируют в различных местообитаниях средней тайги Республики Коми, где общее разнообразие червей значительно выше – до 10 видов (Крылова и др., 2011; Акулова и др., 2017). Локальные фауны червей в тайге западной части Палеарктики также значительно богаче, чем в тундре – по 5–10 видов (Гаврилов, Перель, 1958; Коробейников, 1978; Кудряшева, 1987; Стриганова, Порядина, 2005; Крылова и др., 2011; Гераськина, 2016; Акулова и др., 2017).
Численность дождевых червей при выгонке в эклекторах на порядок превышает значения, полученные при ручном разборе проб. Так, обилие ювенильных особей D. octaedra в тундрах о. Колгуев колебалось от 160 до 240 экз./м2. В некоторых сырых биотопах побережья Большеземельской тундры черви отсутствовали, но в большинстве местообитаний была установлена численность 80–400 экз./м2. Это очень высокие показатели по сравнению с данными ручной разборки (4–19 экз./м2), обычно приводимыми для равнинных тундр (Стебаев, 1962; Стриганова, 1973; Чернов, 1978; Боескоров, 2004; Таскаева и др., 2015; Конакова и др., 2017). Редко (для хорошо дренированных почв и залуженных склонов) указывалась плотность червей 40–150 экз./м2 (Чернов, 1961, 1962, 1978), что близко к значениям в некоторых районах горных тундр (140–160 экз./м2) при плотности коконов до 500 экз./м2 (Берман, 1970; Ермаков, Голованова, 2010).
В окрестностях Падимейских озер (Таскаева и др., 2015) и бассейне р. Черной (Конакова и др., 2017), а также в пунктах 16, 17 и 20 (табл. 1), где численность червей составила при ручной разборке 2–11 экз./м2, эклекторная выгонка также показала на 1–2 порядка бóльшие значения (наши неопубликованные данные).
По-видимому, бóльшая часть особей дождевых червей не доживает до размеров, позволяющих распознать их при ручной разборке проб. Тем не менее именно этим методом, как правило, обнаруживаются половозрелые особи, пригодные для определения. Таким образом, для полноты учетов численности помимо ручной разборки крупных образцов должна применяться экстракция червей в эклекторах или водных воронках (Abrahamsen, 1972).
Особенности обитающих в Арктике видов люмбрицид. Способность E. n. nordenskioldi и D. оctaedra выдерживать экстремальные условия разного рода (Берман и др., 2002а, б; Берман, Мещерякова, 2013; Мещерякова, Берман, 2014) служит хорошим объяснением успешности их экспансии на север.
D. octaedra – относительно мелкий (до 4 см), полиплоидный, партеногенетический, эврибионтный, подстилочный вид (Muldal, 1952; Викторов, 1993; Всеволодова-Перель, 1997; Всеволодова-Перель, Булатова, 2008) – распространен на всех континентах, кроме Антарктиды. Коконы выдерживают охлаждение до –45°С (Мещерякова, Берман, 2014), черви – до –12…–14°С (Берман и др., 2002б). Распространение вида на севере Палеарктики в целом согласуется с показателями холодоустойчивости червей – восточная граница D. octaedra в Восточной Европе и Западной Сибири находится в пределах изотерм –12…–14°С минимальной температуры почвы на глубине 3 см (Берман и др., 2002б). Расчеты, основанные на результатах лабораторного содержания, указывают на то, что развитие этого вида даже в условиях средней полосы продолжается более года (Мазанцева, 1987; Берман и др., 2002б).
E. n. nordenskioldi – среднего размера (до 15 см), подстилочно-почвенный, эврибионтный червь; наиболее распространенная полиплоидная амфимиктическая форма вида E. nordenskioldi (Викторов, 1993; Всеволодова-Перель, 1997; Всеволодова-Перель, Булатова, 2008). Обитает он преимущественно в восточных районах европейской части России и Украины (до Крыма и Северного Кавказа на юге), а также в азиатской части России (Всеволодова-Перель, 1997; Проконова, 2005). В мерзлотных почвах на огромной территории Восточной Сибири и Дальнего Востока это единственный вид дождевых червей (Всеволодова-Перель, 1988; Боескоров, 2004). E. n. nordenskioldi обнаружен на арктических островах Вайгач, Большой Ляховский, Врангеля (Всеволодова-Перель, 1988). В подзоне типичных тундр его биомасса может составлять 80–91% (до 60 г/м2) общей почвенной зоомассы (Чернов, 1962, 1978; Стриганова, 1985). Пищевая активность тундровых E. n. nordenskioldi наблюдается даже при 0°С, что не отмечено для южных популяций (Стриганова, 1985). При этом объем поглощаемой пищи и скорость ее прохождения через кишечник относительно невысоки (Стриганова, 1984). Помимо высокой холодоустойчивости (черви и коконы выдерживают охлаждение до –33 и –40°С соответственно) и морозостойкости (Берман, Лейрих, 1985; Мещерякова, Берман, 2014) этот вид может переживать длительную засуху (Берман и др., 2002а), значительную (рН до 4.5) кислотность почв (Берман, Мещерякова, 2013; Колесникова и др., 2013), а в условиях горных тундр способен к факультативной фитофагии (Берман, 1974). Все эти факторы, по-видимому, и определяют его эврибионтность и обширный ареал. Низкая интенсивность питания и относительно крупные размеры указывают на то, что цикл развития вида, вероятно, занимает несколько лет. В тундровых сборах, как правило, преобладают ювенильные особи. Исследование генетической структуры вида на основании изменчивости митохондриальной и ядерной ДНК обнаружило существование ряда филогенетических линий, разнообразие которых отмечено в горах Алтае-Саянской системы (линии 1–6); но тундры Азии, до о. Вайгач на западе, заселены преимущественно девятой линией (Шеховцов и др., 2017; Shekhovtsov et al., 2018).
L. rubellus – среднего размера (до 15 см), подстилочно-почвенный, диплодный вид с перекрестным оплодотворением, распространен всесветно (Muldal, 1952; Перель, 1979; Викторов, 1993; Всеволодова-Перель, 1997; Всеволодова-Перель, Булатова, 2008). Наши находки L. rubellus в равнинных тундрах Восточной Европы единичны (на южной границе зоны – в окрестностях Воркуты и в двух районах горных тундр на Полярном Урале) и чаще связаны с ложбинами стока или кустарниковыми зарослями (новые данные), хотя в тундрах на севере Норвегии вид обычен (Terhivuo, 1988). Он часто доминирует в средней тайге Коми, но на севере таежной зоны отмечается редко (Крылова и др., 2011). У этого вида устойчивы к замерзанию (до –35°С) только коконы (Мещерякова, Берман, 2014). Даже в северной тайге в разные сезоны года популяции L. rubellus состояли из очень разных по размеру особей (наши неопубликованные данные), что свидетельствует о развитии, длящемся более года (Крылова и др., 2011). Таким образом, в тундре для обитания этого вида могут подойти только незамерзающие почвы в местах скопления снега. Именно с этим, вероятно, связано массовое развитие этого вида в пойменных местообитаниях даже в условиях тайги (Перель, 1979; Матвеева, 1982; Колесникова и др., 2013; Акулова и др., 2017; новые данные по Архангельской обл.). Это косвенно подтверждается его неоднократными находками на северном морском побережье Норвегии (69–70° с.ш.), но отсутствием в прилегающих с юга районах Швеции и Финляндии (Terhivuo, 1988). Поэтому отметка L. rubellus в кустарничково-лишайниковой тундре на севере Большеземельской тундры (Конакова и др., 2017) при среднеянварских температурах воздуха –16°С и почвы на глубине 3 см менее –12°С (Берман и др., 2002б) требует подтверждения. Предполагалось, что и указание L. rubellus в устье р. Лены (Ude, 1902) ошибочно (Перель, 1979).
Распространение видов в восточно-европейском секторе Арктики. E. nordenskioldi и D. octaedra – самые северные представители сем. Lumbricidae в Старом Свете (Перель, 1979; Всеволодова-Перель, 1988; Tiunov et al., 2006). D. octaedra встречается преимущественно в европейском секторе Арктики, а E. n. nordenskioldi – в азиатском (рис. 1), и только на территории Ненецкого и Ямало-Ненецкого автономных округов обитают оба этих вида (Перель, 1979). В наших материалах, как и полтора века назад, на Новой Земле и о. Колгуев (всего три и четыре пункта соответственно) отмечен только D. octaedra, а на о. Вайгач (четыре пункта) – только E. n. nordenskioldi (рис. 2). Новые находки E. n. nordenskioldi подтвердили установленный прежде рубеж – его отметки в равнинных тундрах Старого Света ограничены на западе долиной р. Печоры. D. octaedra, напротив, не найден восточнее Тазовского п-ова (Всеволодова-Перель, 1988; Берман и др., 2002б; Стриганова, Порядина, 2005). В наших неопубликованных материалах с Западного Таймыра – с о. Диксон (2001), из бух. Медуза (2014, сборы С.П. Харитонова), района устья р. Тарея (2010), окрестностей оз. Ладанах (1999), оз. Собачье на западе плато Путорана (1995), оз. Таймыр (1994) – D. octaedra также не был обнаружен. В северных районах Евразии с запада на восток значительно усиливается суровость зим, что препятствует проникновению D. octaedra на большую часть территории Сибири. Восточный предел распространения вида практически совпадает с изолиниями средней температуры воздуха в январе –28°С (рис. 1) и среднегодовой температуры –10°С.
D. octaedra принадлежит к группе космополитных, “перегринных” (Michaelsen, 1903) видов, способных быстро осваивать освободившиеся после схода ледников пространства. “Эндемичный” (Michaelsen, 1903) E. n. nordenskioldi имеет обширный, но ограниченный ареал – заселяет практически целиком Восточную Европу и Сибирь. Северо-западная граница его ареала почти повторяет контуры последнего Скандинавского ледового щита (Svendsen et al., 2004; Величко и др., 2017), по периферии которого располагалась зона “тундростепи” с многолетнемерзлыми почвами, простирающаяся на бóльшую часть Сибири (Динамика…, 2002). Спорадичность западных находок этого вида и их связь с интразональными местообитаниями (поймами, балками) позволяют сделать предположение об остаточном характере этих популяций.
Отсутствие эврибионтного E. n. nordenskioldi на Новой Земле и о. Колгуев могло бы быть связано с трудностью достижения островов этим амфимиктическим видом, но его никогда не находили в тундрах Кольского п-ова, Канинской и Малоземельской тундрах, что в настоящее время не имеет объяснения. Отметка в Хибинах E. n. nordenskioldi по одному ювенильному экземпляру (Zenkova, Rapoport, 2013) требует подтверждения.
Биотопические предпочтения. Количественные исследования распределения дождевых червей в восточно-европейских тундрах не проводились. Их численность в тундрах, обычно определяемая ручной разборкой проб, очень низка. Поскольку в Арктике сосуществование E. n. nordenskioldi и D. octaedra наблюдается только на территории Большеземельской тундры, Ямала и Гыдана, важно рассмотреть возможность их биотопического размежевания. Мы сравнили распределение этих видов в 38 биотопах 12 районов по материалам Института биологии Коми (табл. 1) и в 39 биотопах трех прибрежных районов Большеземельской тундры (табл. 2).
В таежной зоне Республики Коми явные биотопические предпочтения у этих видов не прослеживаются (Акулова, Долгин, 2005; Колесникова и др., 2013; Гераськина, 2016; Акулова и др., 2017). Результаты анализа коллекций Института биологии Коми, собранных в тундре и лесотундре, с учетом опубликованных данных (Таскаева и др., 2015; Конакова и др., 2017) представлены в табл. 3. Все показатели (встречаемость, диапазон и средние значения численности) указывают на явное предпочтение подстилочным D. octaedra биотопов, где зимой может скапливаться снег (ерники, заросли кустарников, колки), тогда как в открытых местообитаниях (тундры, луга) чаще встречается и достоверно более многочислен подстилочно-почвенный E. n. nordenskioldi.
В трех районах побережья Большеземельской тундры у этих видов обнаружены те же предпочтения (рис. 3). В специфических условиях приморских маршей доминирует E. n. nordenskioldi, а под покровом кустарников (в ерниках, околоводных ивняках и ивовых колках) – D. octaedra. В открытых сообществах с разнотравьем и/или кустарничками явных тенденций в распределении видов не прослеживается.
Нельзя исключить, что наличие опада и “удержание” зимой снега в кустарниковых и древесных зарослях благоприятствуют развитию менее холодостойкого вида D. octaedra. В других северных регионах в условиях предельно низких зимних температур D. octaedra также обнаруживается именно в местах скопления снега – долинах ручьев, зарослях кустарников и лесных колках (Стебаев, 1962; Берман и др., 2002б).
ЗАКЛЮЧЕНИЕ
На большей части территории восточно-европейских тундр встречаются только два наиболее холодоустойчивых вида дождевых червей – космополит D. octaedra и преимущественно сибирский E. n. nordenskioldi. Новые находки подтвердили, что в европейском секторе Арктики они сосуществуют только в Большеземельской тундре и на Полярном Урале, а восточная граница ареала D. octaedra в тундрах Палеарктики хорошо описывается пороговыми значениями зимних температур. Причины отсутствия эврибионтного E. n. nordenskioldi в Малоземельской и Канинской тундрах неясны. В районах совместного обитания подстилочно-почвенный E. n. nordenskioldi предпочитает открытые местообитания (различные тундры, луга), тогда как менее холодоустойчивый подстилочный D. octaedra достоверно многочисленнее под покровом кустарников (в ивняках, ерниках, березовом криволесье) со смягченными условиями зимовки. При количественных учетах дождевых червей в тундрах важно сочетать ручной разбор почвенных образцов и выгонку в эклекторах.
Работы Б.Р. Стригановой по дождевым червям в течение многих лет поддерживали интерес отечественных исследователей к этой группе почвенных животных. При обработке материала много полезных советов мы получили от Т.С. Перель, Д.И. Бермана, А.Б. Бабенко, А.Н. Лейрих, А.П. Гераськиной, И.Б. Рапопорт, А.В. Тиунова, А.Г. Викторова. В организации полевых исследований участвовали И.А. Лавриненко, О.В. Лавриненко, М.С. Бизин, С.А. Золотой, С.А. Уваров, Т.М. Романенко. А.А. Кудрин, Т.Н. Конакова, А.А. Нехаева, С.Б. Розенфельд, В.М. Спицин, С.П. Харитонов, З.А. Янченко передали нам свои сборы червей. Экспедиция НАО-2015 была проведена в рамках проекта ПРООН/ГЭФ–Минприроды России “Задачи сохранения биоразнообразия в политике и программах развития энергетического сектора России”. Иллюстрации подготовлены К.В. Макаровым. Мы благодарны всем названным организациям и коллегам, а также двум анонимным рецензентам.
Работа выполена при финансовой поддержке РФФИ (грант 17-04-01603), в рамках Программы Президиума РАН “Биоразнообразие и природные ресурсы России” и госбюджетной темы НИР отдела экологии животных ИБ Коми НЦ УрО РАН АААА-А17-117112850235-2.
Список литературы
Акулова Л.И., Долгин М.М. Животное население (мезофауна) почв среднетаежных луговых экосистем европейского Северо-Востока России // Вестн. Северного (Арктического) федерального ун-та. Сер. естеств. науки. 2005. № 2. С. 64–69.
Акулова Л.И., Долгин М.М., Колесникова А.А. Распространение и численность дождевых червей (Lumbricidae) в подзоне средней тайги Республики Коми // Вестн. Ин-та биологии Коми НЦ УрО РАН. 2017. № 1. С. 4–16.
Белимов Г.Т., Ларионов П.Д., Седалищев В.Т. О распространении и условиях зимовки дождевых червей в долине Средней Лены // Биол. науки. 1981. № 11. С. 46–50.
Берман Д.И. Численность дождевых червей в вертикальных поясах гор Южной Сибири // Бюл. МОИП, отд. биологии. 1970. Т. 75. № 2. С. 153–155.
Берман Д.И. О соотношении трофических групп в биоценозе разнотравно-лишайниковой горной тундры юго-восточного Алтая // Бюл. МОИП, отд. биологии. 1974. Т. 79. № 3. С. 52–63.
Берман Д.И., Лейрих А.Н. О способности дождевого червя Eisenia nordenskioldi (Eisen) (Lumbricidae, Oligochaeta) переносить отрицательные температуры // Докл. АН СССР. 1985. Т. 285. № 5. С. 1258–1261.
Берман Д.И., Мещерякова Е.Н. Ареалы и холодоустойчивость двух подвидов дождевого червя (Eisenia nordenskioldi, Lumbricidae, Oligochaeta) // Зоол. журн. 2013. Т. 92. № 7. С. 771–780.
Берман Д.И., Лейрих А.Н., Алфимов А.В. Об устойчивости дождевого червя, Eisenia nordenskioldi (Oligochaeta, Lumbricidae), к экстремально низкой влажности почвы на северо-востоке Азии // Зоол. журн. 2002а. Т. 81. № 11. С. 1308–1318.
Берман Д.И., Мещерякова Е.Н., Алфимов А.В., Лейрих А.Н. Распространение дождевого червя, Dendrobaena octaedra (Lumbricidae, Oligochaeta), на севере Голарктики ограничено недостаточной морозостойкостью // Зоол. журн. 2002б. Т. 81. № 10. С. 1210–1221.
Боескоров В.С. Экологические условия обитания дождевого червя Eisenia nordenskioldi, Eisen в мерзлотных почвах Якутии: Дис. канд. биол. наук. Улан-Удэ: Институт прикладной экологии Севера Республики Саха, 2004. 211 с.
Величко А.А., Фаустова М.А., Писарева В.В., Карпухина Н.В. История Cкандинавского ледникового покрова и окружающих ландшафтов в валдайскую ледниковую эпоху и начале голоцена // Лед и снег. 2017. Т. 57. № 3. С. 391–416.
Викторов А.Г. Разнообразие полиплоидных рас в семействе дождевых червей Lumbricidae // Успехи соврем. биологии. 1993. Т. 113. Вып. 3. С. 304–312.
Всеволодова-Перель Т.С. Распространение дождевых червей на севере Палеарктики (в пределах СССР) // Биология почв Северной Европы. М.: Наука, 1988. С. 84–99.
Всеволодова-Перель Т.С. Дождевые черви: Кадастр и определитель. М.: Наука, 1997. 102 с.
Всеволодова-Перель Т.С., Булатова Н.Ш. Полиплоидные расы дождевых червей (Lumbricidae, Oligochaeta), распространенные в пределах Восточно-Европейской равнины и в Сибири // Изв. РАН. Сер. биол. 2008. № 4. С. 448–452.
Всеволодова-Перель Т.С., Лейрих А.Н. Распространение и экология дождевого червя Eisenia nordenskioldi pallida (Oligochaeta, Lumbricidae), массового на юге Сибири и Дальнего Востока // Зоол. журн. 2014. Т. 93. № 1. С. 45–52.
Гаврилов К.А., Перель Т.С. Дождевые черви и другие беспозвоночные в почвах лесов Вологодской области // Почвоведение. 1958. № 8. С. 133–140.
Гераськина А.П. Население дождевых червей в основных типах темнохвойных лесов Печоро-Илычского заповедника // Зоол. журн. 2016. Т. 95. № 4. С. 394–405.
Динамика ландшафтных компонентов и внутренних морских бассейнов Северной Евразии за последние 130 000 лет (общая палеогеография) / Ред. Величко А.А. М.: GEOS, 2002. 240 с.
Ермаков А.И., Голованова Е.В. Видовой состав и численность дождевых червей в тундровых биоценозах горного массива Денежкин Камень (Северный Урал) // Сиб. экол. журн. 2010. № 1. С. 15–20.
Козловская Л.С. К характеристике почвенной фауны Большеземельской тундры // Докл. АН СССР, нов. сер. 1955. Т. 104. № 3. С. 485–486.
Козловская Л.С. Сравнительная характеристика почвенной фауны в заполярной части бассейна Усы // Тр. Ин-та леса. 1957. Т. 36. С. 165–177.
Колесникова А.А., Таскаева А.А., Лаптева Е.М., Дёгтева С.В. Вертикальное распределение Collembola, Lumbricidae и Elateridae в аллювиальных почвах пойменных лесов // Сиб. экол. журн. 2013. № 1. С. 45–55.
Конакова Т.Н., Колесникова А.А., Таскаева А.А., Накул Г.Л. Разнообразие почвенных беспозвоночных бассейна р. Чёрная (Большеземельская тундра, Ненецкий автономный округ) // Евраз. энтомол. журн. 2017. Т. 16. № 1. С 88–98.
Коробейников Ю.И. Дождевые черви в почвах лесов Ильменского заповедника // Проблемы почвенной зоологии. Минск: Наука и техника, 1978. С. 118–119.
Крылова Л.П., Акулова Л.И., Долгин М.М. Дождевые черви (Oligohaeta, Lumbricidae) таежной зоны Республики Коми. Сыктывкар: Коми гос. пед. ин-т, 2011. 104 с.
Кудряшева И.В. Почвенные беспозвоночные (мезофауна) лесов низовьев р. Онеги // Почвенная фауна Северной Европы. М.: Наука, 1987. С. 39–50.
Мазанцева Г.П. Рост, развитие и размножение дождевого червя Dendrobaena octaedra (Sav.) (Lumbricidae, Oligochaeta) // Проблемы почвенной зоологии. Тбилиси: Мецниереба, 1987. С. 175–176.
Малевич И.И. Распространение семейства Lumbricidae в европейской части СССР (по картографическим материалам) // Фауна и экология беспозвоночных животных. М.: МГПИ, 1976. Ч. 2. С. 3–11.
Матвеева В.Г. Комплексы дождевых червей пойменных лугов в ландшафтных зонах Европейской территории СССР // Антропогенное воздействие на фауну почв. М.: МГПИ, 1982. С. 54–67.
Мещерякова Е.Н., Берман Д.И. Устойчивость к отрицательным температурам и географическое распространение дождевых червей (Oligochaeta, Lumbricidae, Moniligastridae) // Зоол. журн. 2014. Т. 93. № 1. С. 53–64.
Перель Т.С. Распространение и закономерности распределения дождевых червей фауны СССР (с определительными таблицами Lumbricidae и других Megadrili). М.: Наука, 1979. 272 с.
Проконова Т.В. Основные черты люмбрикофауны (Oligochaeta, Lumbricidae) интразональных ландшафтов Центрального Предкавказья // Фауна Ставрополья. Ставрополь: СГУ, 2005. № 13. С. 84–92.
Рыбалов Л.Б. Зонально-ландшафтная смена населения почвенных беспозвоночных в Приенисейском районе средней Сибири и роль температурных адаптаций в меридиональном (зональном) распределении беспозвоночных // Russ. Entomol. J. 2002. V. 11. № 1. P. 77–86.
Стебаев И.В. Почвенные беспозвоночные салехардских тундр и изменение их группировок под влиянием земледелия // Зоол. журн. 1959. Т. 38. № 10. С. 1550–1572.
Стебаев И.В. Зоологическая характеристика тундровых почв // Зоол. журн. 1962. Т. 41. № 6. С. 816–825.
Стриганова Б.Р. Почвенная фауна северного побережья Кольского полуострова // Ред. Гиляров М.С. Экология почвенных беспозвоночных. М.: Наука, 1973. С. 75–84.
Стриганова Б.Р. Различия пищевой активности разных форм дождевых червей Eisenia nordenskioldi // Зоол. журн. 1984. Т. 63. № 11. С. 1610–1615.
Стриганова Б.Р. Питание дождевых червей Eisenia nordenskioldi в Субарктике // Докл. АН СССР. 1985. Т. 284. № 1. С. 253–256.
Стриганова Б.Р., Порядина Н.М. Животное население почв бореальных лесов Западно-Сибирской равнины. М.: КМК, 2005. 234 с.
Таскаева А.А., Мелехина Е.Н. Активность почвообитающих и наземных беспозвоночных животных в зимний период // Функционирование субарктической гидротермальной экосистемы в зимний период / Ред. Боголицын К.Г., Болотов И.Н. Екатеринбург: УрО РАН, 2011. С. 183–193.
Таскаева А.А., Кудрин А.А., Конакова Т.Н., Колесникова А.А. Разнообразие почвенных беспозвоночных окрестностей Падимейских озер (Большеземельская тундра) // Евраз. энтомол. журн. 2015. Т. 14. № 5. С. 480–488.
Тихомиров Б.А. Об условиях обитания дождевых червей в тундровых почвах // Природа. 1937. № 5. С. 52–58.
Убугунова В.И., Боескоров В.С., Саввинов Г.Н. Распространение дождевых червей в почвах Якутии // Почвоведение. 2007. № 1. С. 69–73.
Чернов Ю.И. К изучению животного населения почв арктических тундр Якутии // Зоол. журн. 1961. Т. 40. № 3. С. 326–333.
Чернов Ю.И. Особенности животного населения почв и дернины некоторых типов тундр западного Таймыра // Уч. зап. МОИП им. Н.К. Крупской. Биогеография. 1962. Т. 109. № 1. С. 183–196.
Чернов Ю.И. Структура животного населения Субарктики. М.: Наука, 1978. 167 с.
Шеховцов С.В., Берман Д.И., Голованова Е.В., Пельтек С.Е. Генетическое разнообразие дождевого червя Eisenia nordenskioldi (Lumbricidae, Annelida) // Вавиловский журн. генетики и селекции. 2017. 8 с.
Abrahamsen G. Ecological study of Lumbricidae (Oligochaeta) in Norwegian coniferous forest soils // Pedobiologia. 1972. Bd 12. F. 4. S. 267–281.
Berman D.I., Marusik Yu.M. On Bimastos parvus (Oligochaeta: Lumbricidae) from Yukon Territory (Canada), with discussion of distribution of the earthworms in Northeastern North America and Northeastern Siberia // Megadrilogica. 1994. V. 5. № 10. P. 113–116.
Eisen G. The Oligochaeta collected during the Swedish expeditions to the Arctic regions in the years 1870, 1875 and 1876 // Kongl. Svenska Vetenskaps-Akademiens Handlingar. 1879. Bd 17. № 7. P. 1–49.
Michaelsen W. Die geographische Verbreitung der Oligochaeten. Berlin, R. Friedländer, 1903. 186 S.
Michaelsen W. Zur Kenntnis der Lumbriciden und ihrer Verbreitung // Ежегодник Зоол. музея Импер. Акад. Наук. 1910. Т. 15. С. 1–74.
Muldal S. The chromosome of the earthworms. I. The evolution of polyploidy // Heredity. 1952. V. 6. P. 55–76.
Reynolds J.W. The distribution of earthworms (Annelida, Oligochaeta) in North America // Advances in Ecology and Environmental Sciences / Eds Mishra P.C., Behera N., Senapati B.K., Guru B.C. New Delhi: Ashish Publ. House, 1995. P. 133–153.
Shekhovtsov S.V., Berman D.I., Bulakhova N.A., Vinokurov N.N., Peltek S.E. Phylogeography of Eisenia nordenskioldi nordenskioldi (Lumbricidae, Oligochaeta) from the north of Asia // Polar Biol. 2018. V. 41. № 2. P. 237–247. https://doi.org/10.1007/s00300-017-2184-2
Svendsen J.I., Alexanderson H., Astakhov V.I., Demidov I., Dowdeswell J.A., Funder S., Gataullin V., Henriksen M., Hjort C., Houmark-Nielsen M., Hubberten H.W., Ingólfson O., Jakobsson M., Kjær K., Larsen E., Lokrantz H., Lunkka J.P., Lyså A., Mangerud J., Matiouchkov A., Murray A., Möller P., Niessen F., Nikolskaya O., Polyak P., Saarnisto M., Siegert C., Siegert M., Spielhagen R., Stein R. Late Quaternary ice sheet history of Northern Eurasia // Quat. Sci. Rev. 2004. V. 23. P. 1229–1271.
Terhivuo J. The Finnish Lumbricidae (Oligochaeta) fauna and its formation // Ann. Zool. Fenn. 1988. V. 25. P. 529–547.
Tiunov A.V., Hale C.M., Holdsworth A.R., Vsevolodova-Perel T.S. Invasion patterns of Lumbricidae into the previously earthworm-free areas of northeastern Europe and the western Great Lakes region of North America // Biol. Invas. 2006. V. 8. P. 1223–1234.
Ude H. Die arktischen Enchytraeiden und Lumbriciden, sowie die geographische Verbreitung dieser Familien // Fauna Arctica. 1902. Bd 2. S. 1–34.
Zenkova I.V., Rapoport I.B. Species richness and high altitude distribution of earthworms in the Khibiny Massive (Murmansk Region) (Oligochaeta) // Advances in Earthworm Taxonomy VI (Annelida: Oligochaeta) / Eds Pavlíček T., Cardet P., Almeida M.T., Pascoal C., Cassio F. Proc. 6th Intl. Oligochaeta Taxonomy Meeting, Palmeira de Faro, Portugal, 22–25 April, 2013. Kasparek Verl.: Heidelberg, 2013. P. 141–151.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая