

Известия РАН. Серия биологическая, 2019, № 5, стр. 533-543
Жуки-жужелицы (Coleoptera, Carabidae) и зоодиагностика экологической сукцессии на техногенных катенах буроугольных отвалов КАТЭКа (Красноярский край)
В. Г. Мордкович 1, И. И. Любечанский 1, *
1 Институт систематики и экологии животных СО РАН
630091 Новосибирск, ул. Фрунзе, 11, Россия
* E-mail: lubech@gmail.com
Поступила в редакцию 04.03.2019
После доработки 15.04.2019
Принята к публикации 15.04.2019
Аннотация
Исследовано население жужелиц на искусственных катенах отвалов буроугольных карьеров в Красноярском крае (возраст 1 мес., 7 и 25 лет). Материал собран на элювиальной, транзитной и аккумулятивной позициях каждой катены. Установлено, что видовое разнообразие и динамическая плотность жужелиц после 25 лет сукцессии на всех позициях техногенных катен не достигают идентичности с природными сообществами. Отмечено, что скорость и направление сукцессии таксоцена жужелиц существенно различаются в зависимости от положения ценозов в рельефе, а отдельные виды, свойственные зональным экосистемам лесостепи, появляются к 7 годам после начала сукцессии. Обнаружено, что на аккумулятивной позиции сукцессия развивается медленнее, чем на элювиальной и транзитной позициях, где рудеральные виды жужелиц постепенно сменяются луговыми мезофилами, а затем мезоксерофилами. Показано, что таксоцен жужелиц на аккумулятивной позиции к 25 годам обретает черты сходства не с травяными, а с древесными сообществами лесостепи в отличие от фитоценоза и почвы.
Важнейшая причина сокращения разнообразия биоты – изменение “пространственной матрицы среды обитания” (Маргалеф, 1992; Хански, 2010). Этому способствует вкрапление в ландшафтный фон биосферы техногенных местообитаний, заселяемых рудеральной биотой обедненного состава, со случайной ценотической организацией и неупорядоченной динамикой (Parmenter, Macmahon, 1987; Dunger et al., 2001; Hendrychová et al., 2012). Весомый вклад в искажение эволюционно сложившейся системы местообитаний вносит угледобыча, которая в России по данным Росстата с 2002 по 2013 г. выросла на 37% (http://www.gks.ru/ bgd/regl/B08_14p/Main.htm). Наиболее негативные экологические последствия имеет добыча угля открытым способом, которая сопровождается уничтожением естественного ландшафта и заменой его обширными карьерами и отвалами с погребением естественных экосистем под многометровой толщей вскрышных пород. Технологией угледобычи предусмотрена рекультивация земель: техническая, т.е. выравнивание или террасирование холмисто-грядового рельефа отвалов, и биологическая, т.е. искусственный засев техноземов травяными или древесными растениями. Однако на практике мероприятия по рекультивации земель обычно либо проводятся с многолетним опозданием, либо не проводятся совсем. В результате происходят хаотическое изменение рельефа и формирование суррогатных экосистем.
Интерес к изучению сукцессий биоты на отвалах, в частности, угольных предприятий возник в 1960-е гг. (Dunger, 1968). Данные, собранные за полвека в разных регионах (в основном на буроугольных отвалах в Восточной Германии, Чехии, СССР, а также в США), выявили ряд закономерностей, имеющих универсальное экологическое значение. На территориях, занятых отвалами, формируются экосистемы, лишь отчасти напоминающие естественные, но сохраняющие зональную направленность сукцессионного развития. Однако они практически никогда не достигают состояния климаксовых биогеоценозов соответствующего биома (Бурыкин, 1985; Титлянова и др., 1993).
Трансформация основных свойств экосистем, возникающих на отвалах, обусловлена прежде всего состоянием и активностью живой фазы, сукцессионными сменами растительного покрова и беспозвоночных животных, интенсивностью биологического круговорота. Медленнее, чем биота, формируется почва. Качественный состав обменных катионов техноземов определяется в первую очередь свойствами подстилающей породы, т.е. вскрыши. Процесс гумусонакопления доминирует над минерализацией, но развивается очень медленно – 0.5 мг С/г. на 1 г субстрата в первые 30 лет. Позднее интенсивность гумусонакопления снижается в 5 раз. Формируется примитивная двучленная почва с маломощным гумусово-аккумулятивным горизонтом, залегающим непосредственно на материнской породе. Искусственное нанесение плодородного гумусового слоя (10–20 см) на поверхность отвала не приводит к восстановлению основных физико-химических свойств, типичных для естественных почв. Техноземы остаются средой обитания с предельно коротким временем формирования и стирания почвенных свойств (Титлянова и др., 1993; Topp et al., 2001; Шугалей, 2009). Тем не менее нанесение плодородного слоя на рекультивируемую поверхность ускоряет процесс формирования растительности и положительно сказывается на фауне почв (Frouz et al., 2007).
Число видов в фитоценозах отвалов угледобычи, предельно низкое изначально (2–4), со временем растет. Однако характерные для климаксовых стадий сукцессии виды, которые появляются, хотя бы единично, с первого года сукцессии, редко становятся доминантами сообщества даже через 20–30 лет. Зато рудеральные виды сохраняют доминантные позиции в сообществе не только на пионерных этапах, но и через 10 и более лет. Видового богатства типичных лугов или лесов эти фитоценозы не достигают и за 50 лет (Brändle et al., 2000). В ходе сукцессии растительности биологический круговорот становится все более закрытым, а скорость обмена общего запаса углерода и азота в экосистеме замедляется (Титлянова, Миронычева-Токарева, 1988).
Из всех живых организмов быстрее других включаются в биологический круговорот отвалов гетеротрофы (Dunger, 2008). Микроартроподы (ногохвостки, панцирные клещи) появляются в субстрате отвалов в первые дни после отсыпки. Их численность и разнообразие с увеличением возраста отвалов увеличиваются во много раз (Бабенко, 1982; Стебаева, Андриевский, 1997). Сведений о структуре и динамике населения макроартропод (мезофауны) значительно меньше. В отличие от простейших, микроартропод и ювенильных особей пауков, попадающих в отвалы путем пассивного переноса ветром, макроартроподы проникают в отвалы активно. Наиболее разнообразно и обильно представлены на отвалах жуки, главным образом стафилиниды и жужелицы (Мордкович, Кулагин, 1986; Дмитриенко, Шаймуратова, 1986; Мордкович, 1994). Предполагается, что наиболее успешными колонистами оказываются всеядные виды (Parmenter, Macmachon, 1987). Высокое видовое разнообразие жуков на рекультивируемых землях может быть связано с разнообразием местообитаний, возникающим из-за холмисто-грядового рельефа отвала. Эти микроместообитания могут служить убежищем редких видов, порой достигающих там высокой численности (Brändle et al., 2000). По мере зарастания отвалов общая численность жесткокрылых может испытывать значительные флуктуации на фоне постепенного снижения степени доминирования видов-рудералов (Frouz et al., 2008; Лузянин и др., 2015). Однако общие закономерности динамики таксоцена жесткокрылых на рекультивируемых землях остаются плохо исследованными, а особенности сообществ жуков в первые дни и недели, прошедшие после появления отвалов, – совсем неизученными.
Жужелицы (Carabidae) были выбраны модельной группой, поскольку эта группа жуков выделяется высоким уровнем видового богатства и численности и широким диапазоном экологических предпочтений. Кроме того, это самый многочисленный компонент зооэдафона, приуроченный к поверхности почвы и ее верхнему горизонту, а поэтому наиболее доступный для изучения.
Полигоном для исследования послужили отвалы буроугольных карьеров КАТЭКа, находящегося в пределах Ачинско-Боготольской, Чулымо-Енисейской и Канской котловин Среднесибирского плоскогорья. Рекультивация земель на КАТЭКе началась с 1973 г. Этот процесс включает в себя несколько последовательных этапов. Первый этап – снятие “вскрыши”, т.е. пород, перекрывающих угольные пласты, отгрузка и вывал породы с формированием отвала. Отвалы – искусственные холмы из легких и средних суглинков, перемешанных с алевролитовым щебнем, высотой 15–25 м, чередующиеся с депрессиями рельефа между ними. Физиономически местность напоминает естественный грядово-лощинный рельеф лесостепной широтной зоны Сибири. Второй этап (рекультивация) предусматривает выравнивание рельефа и формирование приподнятого над остальной местностью плакора. На последующих этапах рекультивации поверх материнской породы насыпают слоем 25–50 см естественную почву. Конечный этап – биологическая рекультивация, т.е. естественное или ускоренное возобновление почвенно-растительного покрова на отвалах.
В 1980-е–начале 1990-х гг. усилиями большого коллектива специалистов-биологов и почвоведов под руководством А.А. Титляновой на территории КАТЭКа проводились комплексные исследования биоразнообразия и биотического круговорота, участником которых был один из авторов данного сообщения. Итоги этих работ изложены в коллективной монографии “Сукцессии и биологический круговорот” (Титлянова и др., 1993). В нее по разным причинам не попали материалы по макроартроподам. Этот пробел, хотя и с большим опозданием, отчасти восполняется данной статьей.
Цель работы – исследование сукцессии таксоцена жуков-жужелиц и оценка влияния холмисто-грядового рельефа и катенной организации местообитаний на параметры α- и β-разнообразия на отвалах угледобычи, а также получение ответов на следующие вопросы:
насколько отличаются состав и разнообразие жужелиц в техногенных экосистемах Канско-Ачинского топливно-энергетического комплекса (КАТЭКа) от таковых в естественных экосистемах “материнского” лесостепного биома?
имеет ли организация сообществ жужелиц на техногенных отвалах признаки классической экологической сукцессии или представляет собой произвольные флуктуации разнообразия и численности случайно занесенных видов?
достигает ли развитие таксоцена жужелиц состояния, характерного для климаксовых стадий экологических сукцессий в лесостепи? Достаточно ли для этого 20-летнего периода, предписанного технологическими стандартами плановой рекультивации (Моторина, 1986)?
как различаются направление, скорость, порядок сукцессионных смен на разных элементах техногенного рельефа и на отвалах разного возраста?
МАТЕРИАЛЫ И МЕТОДЫ
Исследования почвенного населения проводились летом 1986 г. на отвалах естественного зарастания возрастом 1 мес., 7 лет и 25 лет, расположенных внутри карьера на территории Назаровского угольного разреза, в 10 км юго-западнее г. Назарово Красноярского края. В пределах каждого отвала рассматривалась цепочка местообитаний: элювиальные на вершине каждого холма, транзитные на склоне и аккумулятивные в депрессии между холмами. Всего было исследовано девять местообитаний на отвалах. Избранные возрасты отвалов соответствуют условным этапам выравнивания искусственного рельефа и зарастания субстрата сначала рудеральной, а затем характерной растительностью естественных лугов окружающего зонального лесостепного ландшафта.
Атмосферные осадки размывают рыхлые суглинки отвалов, перемещают тонкие фракции субстрата вниз по профилю и обогащают ими пониженные элементы рельефа. Одновременно происходит увеличение щебнистости субстрата вершины и особенно склонов холма. Высота холма к 7-летнему возрасту снижается с 25 до 22 м, а к 25-летнему – до 20 м. Крутизна склона уменьшается с первоначальных 40° до 35° к 7 годам и до 30° к 25 годам. Вынос мелкозема с вершины и склона холма вызывает уменьшение емкости поглощения в начальном периоде почвообразования и тормозит его развитие, а в депрессиях рельефа наоборот (Титлянова и др., 1993). Профиль отвала можно рассматривать как техногенную катену, находящуюся в процессе активного формирования, но уже с дифференциацией позиций на элювиальные, транзитные и аккумулятивные.
В период учетов мезофауны измерялись температура верхних 5 см почвы с помощью почвенных термометров Савинова и содержание влаги в почвенных образцах с помощью взвешивания и последующего высушивания. Разница температур на разных позициях катены была минимальной (2–4°С) на отвале 7-летнего возраста, а на 25-летнем возрастала до 10°С. Наиболее прохладными оказались местообитания аккумулятивных позиций отвалов месячного возраста, наиболее теплыми – местообитания элювиальных позиций на отвалах любого возраста, но особенно 25-летних (табл. 1). Влажность почвы увеличивалась сверху вниз по склону отвалов любого возраста (в 1.5 раза на 7‑летнем и в 4 раза на 25-летнем отвалах). Различия гидротермического режима почв на элювиальных и транзитных позициях были минимальными в первые 7 лет сукцессии и возрастали вдвое на катене 25-летнего возраста (табл. 1).
Таблица 1.
Экологические характеристики техногенных экосистем отвалов Назаровского угольного разреза КАТЭКа (Титлянова и др., 1993)
| Параметры | Возраст отвалов* | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 мес. (0) | 7 лет (66) | 25 лет (79) | |||||||
| EL | TR | AC | EL | TR | AC | EL | TR | AC | |
| Сообщества | Единичные растения | Бурьянное | Бурьянное | Злаково- камышовое | Злаковое | Разнотравно-бобово- злаковое | Бобово- злаково- осоковое | ||
| Виды-доминанты | Tussilago farfara, Artemisia vulgaris, Cirsium setosum |
T. farfara, A. vulgaris, C. setosum, Elytrigia repens |
Typha angustifolia, E. repens, T. farfara, C. setosum |
E. repens, Poa pratensis, Calamagrostis epygeios |
E. repens, Equisetum pratense, Trifolium pratense, Achillea millefolium |
Carex capitate, P. pratensis, C. epygeios, T. pratense |
|||
| Фитомасса, г/м2 | 0 | 0 | 0 | 1337 | 1006 | 1098 | 2339 | 1848 | 2860 |
| Температура почвы (0–10 см), °C | 23.5 | 20.5 | 19 | 27.5 | 26 | 25 | 30.7 | 28 | 20.6 |
| Влажность почвы (0–10 см), % | 14.1 | 13.3 | 25.2 | 18.7 | 18.4 | 28.1 | 8.9 | 18.8 | 36.4 |
| Содержание Сорг, % | 0.11 | – | 0.33 | 0.75 | 0.38 | 1.38 | 3.58 | 5.50 | 8.02 |
| Сорг, г/м2 | 71 | – | 213 | 435 | 225 | 825 | 1790 | 2200 | 3208 |
Тенденция образования и накопления гумуса соответствовала направлению изменения растительного покрова. Концентрация органического углерода (Сорг) и его запас в субстрате отвалов на два порядка меньше, чем в нативных почвах лесостепи. Накопление гумуса увеличивается с возрастом отвала на всех позициях, а на аккумулятивной позиции 25-летнего отвала достигает значений, близких к таковым в естественных луговых почвах (табл. 1) (Титлянова и др., 1993). Запас фитомассы также увеличивается с возрастом отвалов. Общее число видов высших растений на 7-летнем отвале составляет 66, на 25-летнем – 79. На его элювиальной и транзитной позициях складываются ассоциации высокорослых сорняков с огромной биомассой и высоким проективным покрытием. На переувлажненных аккумулятивных позициях 7-летнего отвала формируется примитивное злаково-камышовое сообщество. На 25-летнем отвале все три позиции рельефа различаются по составу доминантов. Более богатое луговым разнотравьем сообщество формируется на аккумулятивной позиции 25-летнего отвала.
В качестве контроля для сравнения с отвалами, в их окрестностях были выбраны два типичных естественных ландшафта среднесибирской лесостепи: остепненный луг на элювиально-транзитной позиции естественной катены с лугово-черноземной почвой и злаково-разнотравной мезоксерофитной растительностью (Stipa pennata, Poa pratensis, Onobrychis arenaria, Potentilla longifolia, Artemisia commutata) и глинисто-галечная отмель в пойме р. Чулым практически без высшей растительности.
В изученных местообитаниях жужелицы составляли 80–90% общего видового богатства и обилия макроартропод. Жужелиц учитывали методом почвенных ловушек (пластиковые стаканчики диаметром 6.5 см). Учеты проводили 10 ловушками в каждом биотопе в течение 5 сут в период летнего максимума активности жужелиц (21–27.07.1986 г.).
Для каждого биотопа рассчитывали индекс видового разнообразия Менхиника. Его значение <1, соответствующее низкому разнообразию видов при высокой численности ярко выраженного доминанта, может служить признаком примитивной организации сообщества, находящегося на пионерной стадии сукцессии. Для сообщества лесостепного биома на средней стадии развития характерны значения индекса Менхиника 1–2, для климаксовых сообществ – еще более высокие значения (Мордкович, 2017). С помощью кластерного анализа было исследовано сходство населения жужелиц изученных биотопов. Вычисляли бинарный (коэффициент Симпсона) и количественный коэффициенты сходства (индекс Дайса). Кластеры строили методом средней связи (UPGMA) с расчетом бутстрэп-коэффициентов (n = 1000). Расчеты проводили в программе PAST V.2.17 (Hammer et al., 2001).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Во всех исследованных искусственных и естественных местообитаниях было выявлено 56 видов жужелиц, суммарная динамическая плотность которых колебалась в разных биотопах от единиц до >100 экз./50 ловушко-суток (л.-с.). Всего через месяц после отсыпки породы видовое разнообразие жужелиц на элювиальной и транзитной позициях катены в отсутствие высшей растительности достигало 5–8 видов, а динамическая плотность – 63–90 экз./50 л.-с. (рис. 1). На аккумулятивной позиции катены, на границе с временным водоемом, разнообразие жужелиц достигало 11 видов, а динамическая плотность – 72 экз./50 л.-с. Единственным доминантом на элювиальной и транзитной позициях был Amara fulva. На аккумулятивной позиции доминировал тот же вид, но с добавлением субдоминантов Bembidion semipunctatum и Nebria livida.
Рис. 1.
Изменение видового богатства и динамической плотности жужелиц по ходу сукцессии на разных позициях искусственных катен и в естественных местообитаниях. а – элювиальная, б – транзитная, в – аккумулятивная позиции катены, Л – остепненный луг; О – отмель. 1 – число видов, 2 – динамическая плотность, экз./50 л.-с., 3 – индекс Менхиника, умноженный на 10.
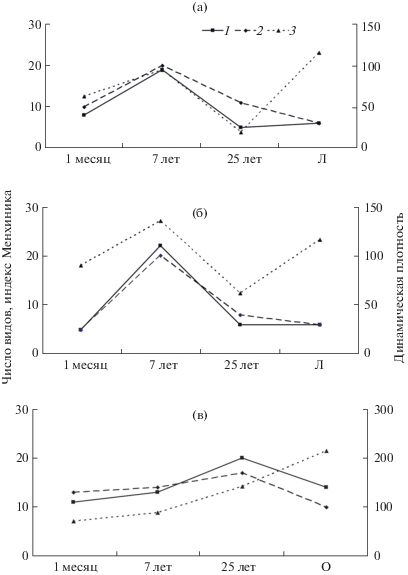
На отвале 7-летнего возраста, заросшем к этому времени густым высокорослым бурьяном, видовое разнообразие жужелиц слабо возрастало на аккумулятивной позиции, на элювиальной возрастало в 2.4, на транзитной – в 4.4 раза по сравнению со свежим отвалом (рис. 1). Произошла смена доминантов. На элювиальной и транзитной позициях катены наряду с A. fulva, продолжавшим играть ведущую роль, и Bembidion femoratum равное с ними положение занимали B. quadrimaculatum, Curtonotus convexiusculus, C. aulicus, Acupalpus meridianus. В сообществе аккумулятивной позиции доминирующее положение сохранял Bembidion semipunctatum, но вместе с ним появлялось и значительное число B. quadrimaculatum, B. lunatum, Asaphidion pallipes.
На 25-летнем этапе сукцессии, когда на элювиальной и транзитной позициях катены закрепилась злаково-разнотравная луговая растительность, наблюдались более низкое разнообразие видов (5–6) и динамическая плотность (до 19 экз./50 л.-с.) жужелиц (рис. 1). Доминантами стали Calathus erratus, Amara equestris, Poecilus fortipes. На аккумулятивной позиции искусственной катены к 25-летнему возрасту сформировался гигрофитный луг с осокой и вейником. Видовое разнообразие жужелиц было выше в 1.5 раза, а динамическая плотность – в 1.6 раза по сравнению с отвалом 7-летнего возраста. Доминантами стали Pterostichus niger, Trechus secalis, Carabus violaceus, C. henningi, C. regalis.
Ход сукцессии до 25 лет на элювиальной и транзитной позициях отвалов был очень похож. Значения индекса Менхиника уже на месячном этапе сукцессии колебались от 0.5 до 1, а к 7 годам достигли значения 2, что свидетельствует о быстрых темпах развития сукцессии в местообитаниях КАТЭКа. Однако к 25 годам сложность таксоцена жужелиц вновь уменьшилась (индекс Менхиника составил 0.8–1).
Более последовательно происходила сукцессия населения жужелиц аккумулятивной позиции катены. Здесь индекс Менхиника медленно, но непрерывно увеличивался от 1.3 до 1.7 от месячного этапа к 7-летнему и далее к 25-летнему.
В контрольных биотопах (остепненный луг для элювиальной и транзитной позиций и речная отмель для аккумулятивной) число видов и динамическая плотность жужелиц были ниже, а индекс Менхиника – выше, чем на промежуточных стадиях сукцессии отвалов (рис. 1).
Кластерный анализ населения жужелиц изученных позиций катен показал, что все биотопы разделяются на две части – “старые” биотопы (25 лет) вместе с естественным биотопом остепненного луга и “молодые” биотопы (1 мес., 7 лет) вместе с постоянно обновляющимся биотопом речной отмели. Сходство отвалов с “эталонными” биотопами было невелико, что может свидетельствовать о четырех независимых кластерах (рис. 2). Исследованные биотопы группируются в основном по степени близости по сукцессионному возрасту. Элювиальные и транзитные позиции оказываются более сходными между собой (причем с высокими бутстрэп-значениями), чем с аккумулятивными. Дерево, построенное по значениям коэффициента Симпсона, учитывающего только присутствие или отсутствие вида, отличается высоким уровнем сходства по позициям в “молодых” отвалах. Элювиальные позиции между собой и транзитные между собой имеют сходство 100% при низких бутстрэп-коэффициентах (51–60).
Рис. 2.
Сходство населения жужелиц отвалов Назаровского угольного разреза КАТЭКа по индексу Дайса. Около узлов графа указаны бутстрэп-коэффициенты (n = 1000). 1 – отвал возраста 1 мес., 7 – отвал возраста 7 лет, 25 – отвал возраста 25 лет; Л – остепненный луг, О – отмель; EL – элювиальная, TR – транзитная, AC – аккумулятивная позиции катены.
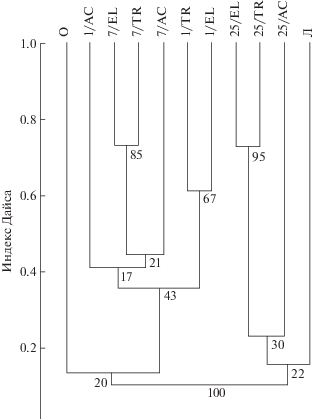
При изучении распределения доминирующих видов жужелиц по позициям разновозрастных катен мы смогли выделить виды, численность которых была значимо выше в некоторых биотопах. Динамическая плотность A. fulva достигала 78 экз./50 л.-с. на транзитной позиции свежего отвала, B. semipunctatum был обилен на аккумулятивной позиции свежего и семилетнего отвала (19 и 40 экз./50 л.-с. соответственно), а также на отмели (38 экз./50 л.-с.). C. convexiusculus был характерен для транзитной позиции 7-летнего отвала, а C. erratus – 25-летнего (62 и 24 экз./50 л.-с. соответственно). Эти виды могут рассматриваться как индикаторы соответствующих стадий сукцессии и геоморфологического положения биотопа.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Сравнение полученных результатов с имеющимися данными по ногохвосткам и панцирным клещам обнаруживает принципиальное различие тенденций изменения разнообразия микро- и макроартропод по мере старения отвалов.
Заселение орибатидами и коллемболами отвалов возрастом 1 мес. первоначально происходит за счет немногих видов, численность которых сначала невелика. Однако далее численность некоторых из них (панцирные клещи Opiella nova, Tectocepheus velatus) скачкообразно растет, делая их доминантами сообщества (Бабенко, 1982; Дмитриенко, 1990; Титлянова и др., 1993; Стебаева, Андриевский, 1997; Богородская и др., 2010). На 4-летнем отвале восстановление комплекса беспозвоночных происходит медленнее. В сообществе доминируют ногохвостки и хищные клещи (Богородская и др., 2010). Третья стадия сукцессии микроартропод отмечается в отвалах 7–10-летнего возраста. Она характеризуется переходом к многовидовому сообществу при сохранении доминирования все тех же рудералов, что и на предыдущих стадиях сукцессии. В еще более старых по возрасту отвалах (20–30 лет) видовое богатство возрастает в несколько раз, но численность видов нестабильна; по-прежнему доминируют рудеральные виды (Титлянова и др., 1993; Wanner, Dunger, 2002).
Таким образом, для микроартропод с их пассивной форезией и необходимостью развиваться непосредственно в субстрате свежеотсыпанного отвала характерно последовательное увеличение разнообразия видов и численности в течение 25–30 лет. Однако полного тождества с составом и структурой населения какого-либо из зональных сообществ лесостепи (лугового, лугово-степного или парково-лесного) не наблюдается (Бабенко, 1982; Титлянова и др., 1993; Frouz et al., 2008; Hendriková et al., 2012).
Сукцессии населения жужелиц, представляющих категорию макроартропод, происходят по-другому. Заселяя отвал в первые часы после отсыпки, имаго жуков массово десантируются из окружающих экосистем и без промедления включаются в биологический круговорот свежеотсыпанного отвала. Этому способствуют нацеленная в отличие от таковой у микроартропод миграция, реализуемая с помощью специальных адаптаций (развитых крыльев, хеморецепции на сырую землю, конечностей, приспособленных к передвижению не только по плоской поверхности, но и в рыхлом субстрате), а также полифагия. На средневозрастном отвале наблюдается высокая численность жужелиц, а на 25-летнем происходит ее снижение. Если у микроартропод даже через 25 лет в составе населения отвалов доминируют, как и на молодых отвалах, виды-эврибионты, то у жужелиц среди доминантов оказываются виды-стенобионты естественной лесостепи.
Порядок, скорость и направление дифференциации населения жужелиц зависят не только от возраста отвала, но и от его положения в рельефе (рис. 2). На элювиальных и транзитных позициях, которые год от года становятся теплее и суше, происходит замещение рудералов (B. femoratum, A. fulva), безраздельно господствующих на месячном отвале, луговыми мезофилами на 7-летнем отвале (B. quadrimaculatum, C. convexiusculus, C. aulicus, A. meridianus). На 25-летнем отвале, где влажность почвы на элювиальной и транзитной позициях уменьшается вдвое по сравнению с таковой на месячном отвале (табл. 1), доминантные позиции в населении жужелиц захватывают мезоксерофильные виды, характерные в естественных ландшафтах сибирской лесостепи для лугов (Мордкович, Любечанский, 2010): C. erratus, A. equestris, P. fortipes, Harpalus nigritarsis, H. modestus. На аккумулятивных позициях, где почва с возрастом становится влажнее (табл. 1), видов-амфибионтов (Nebria livida, B. semipunctatum), которые встречались по кромке временного водоема отвала месячного возраста, на 7-летнем отвале сменяют луговые гигрофилы A. pallipes, B. lunatum и луговые мезофилы C. aulicus, C. convexiusculus, Clivina fossor (табл. 2).
Таблица 2.
Распределение жужелиц по обследованным биотопам КАТЭКа (экз./50 л.-с.)
| Вид | Позиция катены | Биотопы сравнения | N | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1/EL | 1/TR | 1/AC | 7/EL | 7/TR | 7/AC | 25/EL | 25/TR | 25/AC | Л | О | ||
| Acupalpus meridianus | 0 | 0 | 0 | 14 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 22 |
| Agonum ericeti | 0 | 0 | 0 | 0 | 1 | 3 | 0 | 0 | 5 | 0 | 0 | 9 |
| A. gracilipes | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| A. livens | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 |
| Amara apricaria | 7 | 6 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 16 |
| A. bifrons | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| A. communis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 5 |
| A. equestris | 0 | 0 | 0 | 1 | 3 | 0 | 1 | 13 | 0 | 0 | 0 | 18 |
| A. fulva | 40 | 78 | 31 | 10 | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 176 |
| A. ovata | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| A. parvicollis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 3 |
| A. spreta | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 3 |
| A. tibialis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 3 |
| Anisodactylus signatus | 0 | 0 | 0 | 1 | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 4 |
| Asaphidion pallipes | 0 | 0 | 1 | 0 | 1 | 6 | 0 | 0 | 0 | 0 | 0 | 8 |
| Bembidion articulatum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 4 |
| B. femoratum | 10 | 2 | 0 | 5 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 28 |
| B. litorale | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 55 | 55 |
| B. lunatum | 0 | 0 | 3 | 0 | 2 | 4 | 0 | 0 | 0 | 0 | 0 | 9 |
| B. properans | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 0 | 8 |
| B. quadrimaculatum | 2 | 0 | 2 | 17 | 8 | 10 | 0 | 0 | 0 | 3 | 1 | 43 |
| B. semipunctatum | 0 | 0 | 19 | 0 | 0 | 40 | 0 | 0 | 0 | 0 | 38 | 97 |
| B. striatum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 46 | 46 |
| Calathus erratus | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 24 | 6 | 0 | 0 | 32 |
| Cal. melanocephalus | 1 | 0 | 0 | 2 | 2 | 4 | 0 | 0 | 5 | 0 | 0 | 14 |
| Cal. micropterus | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 2 |
| Carabus aeruginosus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 3 |
| C. henningi | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 5 |
| C. regalis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 7 |
| C. violaceus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 0 | 0 | 8 |
| Chlaenius nigricornis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 3 |
| Clivina fossor | 0 | 0 | 1 | 3 | 0 | 2 | 0 | 0 | 3 | 0 | 8 | 17 |
| Curtonotus aulicus | 0 | 0 | 0 | 9 | 4 | 0 | 0 | 0 | 1 | 0 | 0 | 14 |
| Cur. convexinsculus | 1 | 2 | 0 | 15 | 62 | 4 | 0 | 0 | 0 | 0 | 0 | 84 |
| Dyschirius obscurus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 24 | 24 |
| Dyschirius sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 5 |
| Elaphrus riparius | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 6 |
| Harpalus affinis | 0 | 0 | 1 | 4 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 9 |
| H. distinquendus | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| H.modestus | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| H. nigrans | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 1 | 0 | 0 | 0 | 8 |
| H. rubripes | 0 | 0 | 0 | 2 | 1 | 4 | 0 | 1 | 3 | 0 | 0 | 11 |
| H. rufipes | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 1 | 6 | 0 | 0 | 9 |
| Loricera pilicornis | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 2 |
| Microlestes minutulus | 1 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| Nebria gyllenhali | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 5 |
| N. livida | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 13 | 24 |
| Poecilus cupreus | 0 | 2 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 4 |
| P. fortipes | 0 | 0 | 0 | 2 | 1 | 0 | 8 | 22 | 6 | 48 | 0 | 87 |
| P. versicolor | 0 | 0 | 0 | 1 | 2 | 0 | 0 | 0 | 9 | 0 | 0 | 12 |
| Pterostichus eximus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 53 | 0 | 53 |
| Pt. melanarius | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 1 | 0 | 7 |
| Pt. minor | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 |
| Pt. niger | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 44 | 0 | 6 | 50 |
| Synuchus vivalis | 1 | 0 | 0 | 4 | 0 | 4 | 0 | 0 | 3 | 0 | 0 | 12 |
| Trechus secalis | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 15 | 0 | 0 | 15 |
| Сумма особей в позиции катены | 63 | 90 | 72 | 95 | 135 | 87 | 19 | 62 | 142 | 119 | 217 | 1101 |
Появление на аккумулятивной позиции 7-летнего отвала единичных экземпляров лесных по топическим предпочтениям видов, кажущееся случайностью, на 25-летнем отвале превращается в тенденцию, когда в доминанты населения жужелиц выбиваются виды лесов и опушек (T. secalis, C. violaceus, C. henningi, C. regalis) (Мордкович, Любечанский, 2010), хотя никаких признаков облесения в почве или в растительном покрове не отмечено (Титлянова и др., 1993). Таким образом, лесной таксоцен жужелиц развивается раньше, чем появляется соответствующая растительность, и в этом можно видеть отличие сукцессии на отвалах от идущей в естественных биоценозах лесостепи. По-видимому, следует признать жужелиц предвестниками каких-то, пока не выявленных, процессов и свойств, прогнозирующих в ближайшей перспективе оформление в характеризуемом местообитании экосистемы лесного типа.
За сменой видов жужелиц с разными топическими предпочтениями кроются кардинальные перестройки структурно-функциональной организации формирующихся на отвалах сообществ. Наибольший вклад в этот процесс вносят виды-ценофилы, обладающие высокой численностью и четко дифференцирующие геоморфологические позиции и стадии сукцессии.
ЗАКЛЮЧЕНИЕ
Со временем дифференциация экологических условий на разных позициях рельефа отвалов, ничтожная в месячном возрасте, последовательно увеличивается. Катенный тип организации отвалов и дробная дифференциация среды в пространстве и во времени формируют множество местообитаний макроартропод.
Скорость и направление сукцессии таксоцена жужелиц существенно различаются в зависимости от положения ценозов в рельефе. На элювиальной и транзитной позициях техногенных катен отдельные виды, свойственные зональным экосистемам лесостепи, появляются к 7 годам после начала сукцессии. Наиболее быстрыми темпами и в наиболее комфортном для почвенных беспозвоночных направлении меняются условия на аккумулятивных позициях катен. Здесь сукцессия развивается последовательно, но медленнее, чем на элювиальной и транзитной позициях. Видовое разнообразие и динамическая плотность жужелиц после 25 лет сукцессии на всех позициях техногенных катен не достигают идентичности с природными сообществами. Таксоцен жужелиц на аккумулятивной позиции к 25 годам обретает черты сходства не с травяными, а с древесными сообществами лесостепи в отличие от фитоценоза и почвы.
Охарактеризованные закономерности изменения населения таксоцена жужелиц, последовательные во времени и пространстве, могут в совокупности квалифицироваться как экологические сукцессии, поскольку приближаются к состоянию, близкому к климаксовым состояниям сообществ естественной лесостепи. Однако этот процесс требует не 12–18 лет, принятых сегодня стандартом времени рекультивации, а гораздо больше. По нашим данным, для этого недостаточно и 25 лет.
Авторы благодарны А.А. Титляновой, Н.П. Миронычевой-Токаревой, В.С. Андриевскому, Н.П. Косых, Г.Д. Мордкович, Н.А. Афанасьеву (Институт почвоведения и агрохимии СО РАН, Новосибирск) за сотрудничество в совместных исследованиях, Б.М Катаеву (ЗИН РАН, Санкт-Петербург) и И.А. Белоусову (ВИЗР, Санкт-Петербург) за помощь в определении жужелиц.
Исследование выполнено при финансовой поддержке Программы фундаментальных научных исследований (ФНИ) государственных академий наук на 2013–2020 гг. (проект № VI.51.1.7. (0311-2016-0007) (АААА-А16-116121410123-1)).
Список литературы
Бабенко А.Б. Динамика почвенных микроартропод при зарастании техногенных отвалов // Зоол. журн. 1982. Т. 61. № 12. С. 1815–1825.
Богородская А.В., Краснощёкова Е.Н., Трефилова О.В., Шишикин А.С. Сезонная динамика развития микробиоценозов и комплексов беспозвоночных на отвалах вскрышных пород Бородинского буроугольного разреза (КАТЭК) // География и природ. ресурсы. 2010. № 4. С. 36–45.
Бурыкин A.M. Некоторые теоретические вопросы рекультивации техногенных ландшафтов // Рекультивация земель, нарушенных горными работами на КМА. Воронеж: ВСХИ, 1985. С. 3–11.
Дмитриенко В.К. Динамика сообществ беспозвоночных в грунтах промышленных отвалов западной зоны КАТЭКа, используемых для лесовыращивания // Современное состояние биоценозов зоны КАТЭКа. Л.: Гидрометеоиздат, 1990. С. 186–197.
Дмитриенко В.К., Шаймуратова Н.С. Структура комплексов педобионтов рекультивируемых земель западной зоны КАТЭКа // География и природ. ресурсы. 1986. № 4. С. 81–85.
Лузянин С.Л., Дудко Р.Ю., Беспалов А.Н., Еремеева Н.И. Биоразнообразие жужелиц (Coleoptera, Carabidae) на отвалах угольных разрезов Кузбасса // Евраз. энтомол. журн. 2015. Т. 14. № 5. С. 455–467.
Маргалеф Р. Облик биосферы. М.: Наука, 1992. 214 с.
Мордкович В.Г. Зоологические сукцессии на молодых техногенных катенах и принцип пространственно-временных аналогов // Изв. РАН. Сер. биол. 1994. № 3. С. 446–452.
Мордкович В.Г. Сопряженность экологических сукцессий с катенной организацией пространства // Журн. общ. биологии. 2017. Т. 78. № 4. С. 32–46.
Мордкович В.Г., Кулагин О.Е. Состав жужелиц и диагностика направления техногенных экосистем Кузбасса // Изв. СО АН СССР. Сер. биол. 1986. № 2. С. 86–92.
Мордкович В.Г., Любечанский И.И. Экологические группы видов жуков-жужелиц (Coleoptera, Carabidae): сущность, принципы выделения, состав и востребованность // Евраз. энтомол. журн. 2010. Т. 9. С. 195–202.
Моторина Л.В. Естественное восстановление биогеоценозов и рекультивация земель, нарушенных при открытой добыче полезных ископаемых // Общ. проблемы биогеоценологии. 1986. Т. 2. С. 44–46.
Стебаева С.К., Андриевский В.С. Ногохвостки (Collembola) и панцирные клещи (Oribatei) на буроугольных отвалах Сибири // Зоол. журн. 1997. Т. 76. № 9. С. 1004–1015.
Титлянова А.А., Миронычева-Токарева Н.П., Наумов Н.Б. Круговорот углерода в травяных экосистемах при зарастании отвалов // Почвоведение. 1988. № 7. С. 164–174.
Титлянова А.А., Афанасьев Н.А., Наумова Н.Б., Андриевский В.С., Артамонова В.С., Булавко Г.И., Гантимурова Н.И., Косинова Л.Ю., Косых Н.П., Миронычева-Токарева Н.П., Мордкович Г.Д., Наумов А.В., Напрасникова Е.В., Половинко Г.П., Стебаева С.К., Якутин М.В. Сукцессии и биологический круговорот / Ред. Курачев В.М. Новосибирск: Наука, 1993. 157 с.
Хански И. Ускользающий мир. Экологические последствия утраты местообитаний. М.: КМК, 2010. 344 с.
Шугалей Л.С. Устойчивость почв лесостепи и южной тайги Средней Сибири к экзогенным воздействиям // Вестн. КрасГАУ. 2009. № 9. С. 66–77.
Brändle M., Durka W., Altmoos M. Diversity of surface dwelling beetle assemblages in open-cast lignite mines in Central Germany // Biodiv. Conserv. 2000. V. 9. P. 1297–1311.
Dunger W. Die Entwicklung der Bodenfauna aus rekultivieren Kippen und Halden des Braunkoblengelbaues // Abhandl. Berichte Naturkundemuseum, Gorlitz. 1968. Bd 43. № 2. 256 S.
Dunger W. Tiere im Boden // 4 unveränderte Auflage Nachdruck der 3 Auflage von 1983. Westarp Wissenschaften-Verlagsgesellschaft GmbH, Hohenwarsleben, 2008. 280 S.
Dunger W., Wanner M., Hauser H., Hohberg K., Schulz H.-J., Schwalbe T., Seifert B., Vogel J., Voigtländer K., Zimdars B., Zulka K.P. Development of soil fauna at mine sites during 46 years after afforestation // Pedobiologia. 2001. V. 45. P. 243–271.
Frouz J., Pižl V., Tajovský K. The effect of earthworms and other saprophagous macrofauna on soil microstructure in reclaimed and un-reclaimed post-mining sites in Central Europe // Eur. J. Soil Biol. 2007. V. 43. Suppl. 1. P. S184–S189.
Frouz J., Prach K., Pižl V., Háněl L., Starý J., Tajovský K., Materna J., Balík V., Kalčík J., Řehounková K. Interactions between soil development, vegetation and soil fauna during spontaneous succession in post mining sites // Eur. J. Soil Biol. 2008. V. 44. P. 109–121.
Hammer Ø., Harper D.A.T., Ryan P.D. PAST: Paleontological statistics software package for education and data analysis // Palaeontol. Electron. 2001. V. 4. № 1. 9 p. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Hendrychová M., Šálek M., Tajovský K., Řehoř M. Soil properties and species richness of invertebrates on afforested sites after brown coal // Restor. Ecol. 2012. V. 20. P. 561–567.
Parmenter R.R., Macmahon J.A. Early Successional Patterns of Arthropod Recolonization on Reclaimed Strip Mines in Southwestern Wyoming: The Ground-dwelling Beetle Fauna (Coleoptera) // Environ. Entomol. 1987. V. 16. P. 168–177.
Topp W., Simon M., Kautz G., Dworschak U., Nicolini F., Prückner S. Soil fauna of a reclaimed lignite open-cast mine of the Rhineland: improvement of soil quality by surface pattern // Ecol. Eng. 2001. V. 17. P. 307–322.
Wanner M., Dunger W. Primary immigration and succession of soil organisms on reclaimed opencast coal mining areas in eastern Germany // Eur. J. Soil Biol. 2002. V. 38. P. 137–143.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая