

Известия РАН. Серия биологическая, 2019, № 5, стр. 524-532
Внутрирасовое разнообразие карпатской расы медоносной пчелы Apis mellifera carpatica
А. Ф. Сафонкин 1, *, Т. А. Триселева 1, Т. О. Быкова 2
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
117071 Москва, Ленинский просп., 33, Россия
2 Крымский федеральный университет им. В.И. Вернадского
295007 Симферополь, просп. Академика Вернадского, 4, Россия
* E-mail: andrej-safonkin@yandex.ru
Поступила в редакцию 04.03.2019
После доработки 15.04.2019
Принята к публикации 15.04.2019
Аннотация
Проведено морфометрическое и молекулярно-генетическое исследование внутрирасовых различий карпатской расы медоносной пчелы из России и Таджикистана по сравнению со стандартом расы (Вучковская линия из Закарпатской обл. Украины). Отмечено, что наиболее информативные признаки “кубитальный индекс” и “дискоидальное смещение” значительно отклоняются от стандарта, хотя расчет генетической дистанции между выборками из разных регионов свидетельствует о единстве карпатской расы пчелы по ареалу. По нуклеотидному составу в гене СОI мтДНК длиной 1535 п.н. выделено две группы гаплотипов, связанных со временем формирования пасеки и различающихся по аминокислотному составу белка СОI (группа I – аспарагин, группа II – серин). Обнаружено, что группа I помимо образцов пчел из разных регионов включает в себя пчел изолированной пасеки из горно-лесной зоны Крыма, сформированной на основе пчел линии 77 из Закарпатья, а группа II – образцы пчел современной линии стандарта “Вучковский”.
Медоносная пчела Apis mellifera L. характеризуется внутривидовой дифференциацией, позволяющей выделить от 25 (Ruttner, 1992) до 28 (Engel, 1999) географических рас медоносной пчелы, адаптированных к местным условиям (Ruttner, 1988). На территории Восточной Европы в последнее время широко распространена карпатская раса медоносной пчелы Apis mellifera carpatica (Avetisyan, Gubin, Davidenco, 1966) (Гайдар, Левченко, 2003). Еще в начале XX в. некоторые исследователи обращали внимание на то, что по комплексу биологических и морфологических признаков карпатские пчелы составляют особую группу, существенно отличающуюся от Apis mellifera carnica из Австрии (Гайни, 1923). Однако до настоящего времени существуют разные точки зрения на степень отличия карпатской расы пчел от ветви A. m. carnica, вплоть до самостоятельного подвида (Зиновьева и др., 2013). Губин (Губин, 1972) выделял A. m. carpatica как особую популяцию A. m. carnica, которая сформировалась в результате распространения пчел по долинам р. Дуная, а затем р. Тисы, с проникновением в Восточные Бескиды. С этой точки зрения карпатская раса – экотип A. m. carnica (Маннапов и др., 2015). Некоторые авторы считали карпатских пчел промежуточной формой между расами пчел A. m. carnica и Apis mellifera sossimai (украинской) (Сэндуляк, 1965) или восточной популяцией украинской суперпороды А. m. carnica Pollm. (Уфимцева, Осинцева, 2009). Эти гипотезы основаны на морфологическом и экологическом подходах с учетом генетических методов сравнения.
Первоначально генетический метод изучения рас пчел был основан на сравнительном анализе длины и нуклеотидной изменчивости межгенного участка СОI–СОII мтДНК и анализе некоторых микросателлитов хромосом ядра. Например, было показано, что популяция пчел A. m. carpatica Румынии по локусу СОI–COII однородна (Cauia et al., 2008). В других исследованиях популяций A. m. carpatica, также основанных на анализе межгенного локуса и микросателлитов, были отмечены различия по популяциям, обитающим в разных регионах (Адыгея–Закарпатье), и показано, что карпатская раса далека от A. m. carnica (Калашников, 2013). В то же время (Зиновьева и др., 2013) был проведен сравнительный анализ популяций по микросателлитам, показавший, что степень внутрипопуляционной дифференциации у A. m. carpatica и A. m. carnica иногда выше, чем между популяциями A. m. carpatica и A. m. carnica. Эти данные, по мнению авторов, могут указывать на их общее происхождение и существование A. m. carpatica в качестве популяции A. m. carnica. Таким образом, выбор межгенного участка СОI–COII и микросателлитов для анализа межпопуляционных различий дает противоречивые результаты. В последнее время появились работы, свидетельствующие о ценности гена COI мтДНК для анализа расового и популяционного разнообразия A. mellifera (Martimianakis et al., 2011; Ozdil, Ilhan, 2012; Быкова и др., 2016).
При морфологических исследованиях пчел предполагается, что экстерьерные признаки (длина хоботка, длина и ширина переднего крыла, длина третьего и длина четвертого тергитов, кубитальный индекс, дискоидальное смещение, окраска тергитов, форма задней границы воскового зеркальца пятого стернита) наиболее устойчивы и по ним можно определить расовую принадлежность (Маннапов и др., 2015).
В настоящее время превалирует мнение, что только комплексный подход позволяет полностью оценить ту или иную породу пчел. Актуальность исследования морфогенетических различий связана с большим хозяйственным значением пчел. “Карпатские пчелы за относительно короткий период … стали массово использоваться на пасеках почти всех республик бывшего Советского Союза. Наряду с этим в последние 15 лет с выгодой начали разводить карпаток пчеловоды Венгрии, Чехии, Словакии и особенно Польши, где в основном распространены краинские пчелы” (Гайдар, Левченко, 2003). Предполагается, что превосходство A. m. carpatica над A. m. carnica определяется особенностями их этологических характеристик (Морев и др., 2013). Однако в современной литературе накапливаются данные о том, что в результате деятельности человека происходит интенсивная гибридизация пород пчел. При этом при метизации с другими расами происходит изменение экологической пластичности помеси с потерей полезных признаков исходных рас пчел (Морев и др., 2013). “Потеря породного разнообразия оказывается не только утратой уникального бесценного генетического разнообразия, но и сужением генетического потенциала, принципиально ограничивающим возможности селекционной работы, породообразовательного процесса в настоящем и будущем” (Столповский, 2010). Таким образом, сохранение генофондов различных пород животных, в том числе и медоносных пчел, оптимизация их использования непосредственно зависят от степени разработки методов контроля уровня их биоразнообразия и популяционно-генетической структуры. До настоящего времени в литературе нет достаточного массива данных, оценивающего пределы внутрирасовой изменчивости пчел по комплексу признаков.
Цель работы – изучение внутрирасового разнообразия пчел A. m. carpatica на основе анализа морфологических и молекулярных данных.
МАТЕРИАЛЫ И МЕТОДЫ
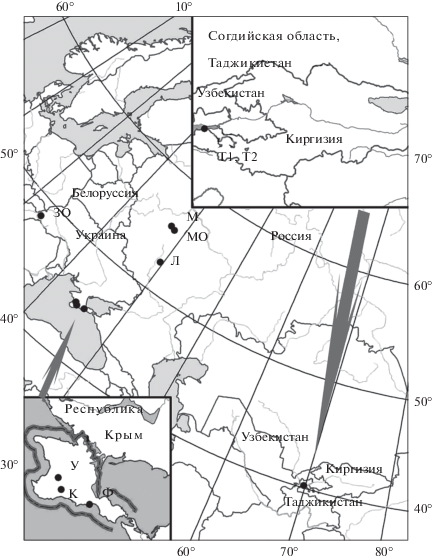
Материалом для анализа послужили особи рабочих пчел с пасек, расположенных в Республике Крым – “Красные пещеры” (К) и с. Укромное (У) Симферопольского р-на, г. Феодосия (Ф); в Республике Таджикистан – с. Хистеварз Бободжон Гафуровского р-на Согдийской обл. (Т1, Т2); в Московской обл. – г. Гжель (МО); в Липецкой обл. (Л); на территории Московской государственной академии ветеринарной медицины и биотехнологии им. К.И. Скрябина, г. Москва (М); в Закарпатской обл. Украины (ЗО) (рис. 1). Пчелиные семьи на пасеку К были привезены из Средней Азии и в течение 30 лет разводились в горно-лесной зоне Крыма без завоза маток известных рас. Образцы из Закарпатья были представлены Вучковской линией карпатской расы.
Рис. 1.
Места сбора образцов карпатской расы медоносной пчелы Apis mellifera carpatica. Буквенные обозначения на рис. 1–3 – места сбора: К – “Красные пещеры”, У – с. Укромное, Ф – г. Феодосия (Республика Крым); Т1, Т2 – с. Хистеварз Бободжон Гафуровского р-на Согдийской обл. (Республика Таджикистан); МО – г. Гжель (Московская обл.); Л – Липецкая обл.; М – территория Московской государственной академии ветеринарной медицины и биотехнологии им. К.И. Скрябина, г. Москва; ЗО – Закарпатская обл. Украины.
Анализ морфометрических показателей. Рабочая гипотеза о возможной изменчивости признаков основана на факте длительной изоляции или продолжительного разведения без завоза маток известных рас. Поэтому для данного анализа взяты выборки пчел c пасек южной части ареала карпатки – из Крыма и Таджикистана. Контролем была выборка пчел Вучковской линии. Были проанализированы следующие морфометрические признаки определения расы пчел (Маннапов и др., 2015): длина хоботка, кубитальный индекс, дискоидальное смещение, длина и ширина переднего правого крыла. Табл. 1 построена по принципу сравнения собственных и известных для пчел карпатской расы литературных данных, поэтому признак “кубитальный индекс” отражен в ней как в процентах (отношение длины меньшей жилки к большей ×100), так и в долях (обратное соотношение). Измерения были проведены с помощью микроскопов МБС (ЛОМО, Россия) и VHX-1000E (Keyence, Япония). Статистический анализ проводили с помощью программ Statistica 10.0. Мы использовали однофакторный анализ с неравным числом повторностей (ANOVA) для выявления различий между популяциями по значениям морфометрических параметров. Для множественного сравнения значений был использован Fisher’s LSD post hoc тест при 95%-ном уровне значимости.
Таблица 1.
Морфометрические показатели карпатской расы медоносной пчелы Apis mellifera carpatica (среднее значение ± стандартная ошибка)
| Регион, пасека, раса, стандарт | n | Длина хоботка |
Верхнее крыло | Источник | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| дискоидальное смещение, % | кубитальный индекс | длина | ширина | |||||||
| + | 0 | – | % | доля | ||||||
| ЗО (стандарт “Вучковский”) | 68 | 6.41 ± 0.07a | 89.5 | 10.5 | 0 | 37.55 ± 0.85a | 2.73 ± 0.05a | 9.54 ± 0.03a | 3.35 ± 0.02a | Собственные данные |
| Т1 | 20 | 5.95 ± 0.12b | 55.0 | 35.0 | 10.0 | 43.62 ± 1.35b | 2.34 ± 0.08b | 8.97 ± 0.07b | 3.15 ± 0.03b | То же |
| Т2 | 57 | 5.92 ± 0.11b | 100 | 0 | 0 | 45.21 ± 0.76b | 2.24 ± 0.03b | 9.35 ± 0.04a | 3.03 ± 0.02c | » |
| К | 292 | 6.42 ± 0.02a | 40.75 | 47.94 | 11.30 | 42.13 ± 0.52b | 2.48 ± 0.03b | 9.06 ± 0.01b | 3.12 ± 0.01b | » |
| Ф | 27 | 6.30 ± 0.06a | 66.67 | 33.33 | 0 | 40.75 ± 1.26a, b | 2.51 ± 0.07a, b | 9.04 ± 0.04b | 3.15 ± 0.02b | » |
| У | 38 | 6.42 ± 0.04a | 63.16 | 34.21 | 2.63 | 39.42 ± 0.80a, b | 2.57 ± 0.05a | 8.99 ± 0.04b | 3.13 ± 0.02b | » |
| ANOVA | F = 18.03; р < 0.001 |
F = 7.12;
р < 0.001; T1/ЗО: p = 0.03; У/T2: р = 0.006 |
F = 8.04;
р < 0.001; У/T2, K/T2: р = 0.006 |
F = 60.9; р < 0.001 | F = 49.8;
р < 0.001; T1/T2: р = 0.03; T2/У: р < 0.02 |
|||||
| Карпатская раса | – | 6.5–6.6 | > 80 | – | – | – | 2.4–2.6 | 9.3–9.6 | – | (Губин, 1976) |
| Карпатская раса | – | 6.6–7.0 | 95–100 (≥85)* | – | – | 33–43 (38)* |
2.3–3 (2.6)* |
– | – | (Гайдар, 2004) |
| Стандарт “Майкопский” | – | 6.7±0.2 | 93 | 5 | 2 | 47.9 ± 0.2 | 2.4–2.6 | 9.27–9.63 | 3.21 | (Маннапов и др., 2015) |
| Карпатская раса | – | – | 99.74 | – | – | – | 2.781±0.016 | – | – | (Папп и др., 2013) |
Молекулярно-генетический анализ. Общая ДНК была выделена из лапок имаго пчел с использованием реагентов “Диатом 200” (Изоген, Москва). Амплификацию проводили с использованием набора реактивов для ПЦР 5хMasterMix (Диалат, Москва). Анализируемый фрагмент гена COI мтДНК составлял 1535 пар нуклеотидов. Для амплификации данного участка были использованы три пары праймеров. ПЦР проводили с праймерами LCO1490 и HCO2198 для начального участка гена COI согласно протоколу (Folmer et al., 1994). Для амплификации остального участка использовали экспериментально подобранные нами две пары праймеров:
F2029 (TTCTTCACCTTCAGTAGATTTTG) и R3248 (TGAATTAAGTGGGG AAATTTTA); Fint2587 (AGCAACTTATCATGGTTCAAAA) и Rint2734 (GA ACATAATGAAAATGTCCAACA).
Реакцию проводили при следующих условиях: 95° – 3 мин (1 цикл); 95° – 45 с, 50° – 1 мин, 72° – 2 мин (35 циклов); 72° – 5 мин (1 цикл). Продукт амплификации очищали методом осаждения раствором этилового спирта с добавлением 5 М ацетата натрия. Электрофорез и чтение нуклеотидных последовательностей продукта амплификации выполняли на автоматическом секвенаторе ABI PRISM 3130 (Applied Biosystems, США) с использованием набора реактивов BigDye Terminatot kit 3.1 (Applied Biosystems) в кабинете методов молекулярной диагностики ИПЭЭ РАН.
Обработку данных проводили с использованием пакетов программ MEGA 5 (Tamura et al., 2011), Network ver. 4.6.1.1 (Bandelt et al., 1999), ARLEQUIN ver. 3.5 (Excoffier, Lischer, 2011), MrBayes ver. 3.0 (Ronquist, Huelsenbeck, 2003). Всего было проанализировано 26 последовательностей. Полученные последовательности гена COI мтДНК образцов A. m. carpatica, представленные отдельными гаплотипами или образцами одних и тех же гаплотипов, но из разных мест сбора, депонированы в GenBank (htpp://www.ncbi.nlm.nih.gov/) под номерами MF100910-MF100926. Всего в GenBank зарегистрировано 17 образцов. Для сравнительного анализа дополнительно из GenBank были взяты последовательности гомологичного участка русской Apis mellifera mellifera (KJ396188.1) и итальянской Apis mellifera ligustica (LO6178.1) рас.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Морфометрический анализ. Результаты анализа представлены в табл. 1. Длина верхнего крыла колеблется от 8.97 до 9.54 мм. Не отличаются от контроля (Вучковская линия, ЗО) по признаку длины крыла только пчелы пасеки Т2. Ширина крыла во всех выборках меньше контрольной, особенно узкие крылья у пчел пасеки Т2. Признак “дискоидальное смещение” имеет в основном положительное и нейтральное значение. Отдельные особи из У, Т1 и особенно К имеют отрицательное значение этого признака. Признак “кубитальный индекс” меняется от 2.25 (45.2%) у пчел из Т2 до 2.71 (38.8%) у Вучковкой линии (ЗО). В процентном и в долевом выражении показателя отличается пасека Т2. Длина хоботка варьирует от 5.53 до 6.42 мм. Значимые различия обнаружены только между пчелами Вучковской линии и семьями с пасек Таджикистана.
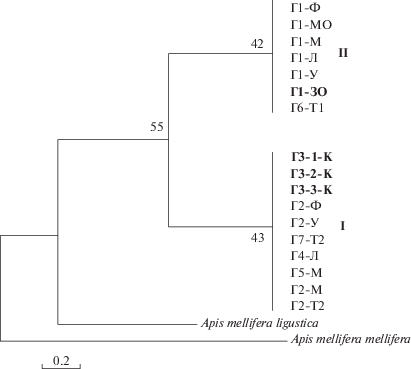
Молекулярно-генетический анализ. При выравнивании полученных нами нуклеотидных последовательностей гена COI мтДНК образцов медоносной пчелы обнаружены девять вариабельных сайтов, пять из которых – парсимониально-информативные. Общее число замен – восемь транзиций, одна трансверсия. Нуклеотидный состав представлен 13.1% цитозина, 41.45% тимина, 34.42% аденина, 11.03% гуанина. Нуклеотидная изменчивость всей выборки составляет 0.1% (S.E. = = 0). Описано девять гаплотипов, из которых четыре уникальны, пять встречаются в нескольких образцах с частотой 2–9. Дендрограммы, построенные методами Байеса (MrBayes), ближайшего связывания (Neighbor-Joining) и максимальной экономии (Maximim Parsimony) по нуклеотидным последовательностям и аминокислотам, совпадают в ключевых узлах с выделением двух групп.
В обеих группах присутствуют образцы, собранные со всех анализируемых пасек. Однако нуклеотидные последовательности образцов пчел, входящих в группу I, имеют семь вариабельных сайтов, из которых три парсимониально-информативные. В ней присутствуют образцы с пасеки К. Группа II состоит из образцов, нуклеотидные последовательности которых имеют всего один вариабельный сайт. В этой группе присутствуют образцы Вучковской линии.
Дендрограмма (рис. 2), построенная на основе аминокислот, несмотря на небольшую поддержку, подтверждает принципиальное разделение образцов на две группы. Замена гуанозина на аденозин во второй позиции кодона (сайт 110) приводит к замещению серина на аспарагин. Для группы I, в которую входят образцы с пасеки К, характерен аспарагин, для группы II, в которую входят образцы с пасеки ЗО (стандарт “Вучковский”), – серин.
Рис. 2.
Дендрограмма отношений образцов карпатской расы медоносной пчелы Apis mellifera carpatica из разных регионов, построенная на основании аминокислотной последовательности белка СОI мтДНК методом ближайшего связывания (Neighbor-Joining) с использованием модели p-distance. Шкала – генетическая дистанция между гаплотипами. Г1–Г7 – отдельные гаплотипы; для рис. 2 и 3. Аминокислоты: I – аспарагин, II – серин. Жирным шрифтом выделены особи Вучковской линии и с пасеки К.
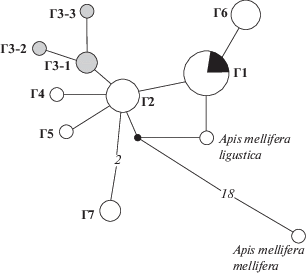
Отношения между гаплотипами иллюстрирует также медианная сеть (рис. 3). Центральное место на ней занимает гаплотип 2 (Г2), от которого отходят все другие гаплотипы. Один из гаплотипов этой группы дает начало Г1, куда входят Вучковская линия, а также часть образцов из других популяций. Г2 берет свое начало через гипотетического предка от среднерусской темно-серой пчелы, а Г1 ближе к итальянской породе.
Рис. 3.
Медианная сеть гаплотипов образцов карпатской расы медоносной пчелы Apis mellifera carpatica из разных регионов, построенная на основании нуклеотидной последовательности гена СОI мтДНК. Круги – отдельные гаплотипы, диаметр круга соответствует числу образцов. Черной точкой отмечен предковый гаплотип. Числа на ветвях – количество нуклеотидных замен, отсутствие числа между гаплотипами – одна замена. Черным цветом в Г1 выделены особи Вучковской линии, серым – с пасеки К.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Известно, что карпатская раса пчел характеризуется достаточно стабильным комплексом морфоэкологических признаков (Гайдар, 2004; Папп и др., 2013; Маннапов и др., 2015). В.А. Губин, обобщив работы по изучению морфологических признаков местных пчел Украинских Карпат, предложил морфоэтологический стандарт для карпатских пчел (Губин, 1976). При этом он обращал внимание на то, что предложенный стандарт не средний показатель для пчел Закарпатья, а эталон чистопородной пчелы карпатской популяции. Однако в литературе мало данных, свидетельствующих о том, насколько эти признаки позволяют соотнести пчел отдельной пасеки с исследуемой расой. Актуальность вопроса связана с тем, что начиная с 1970-х гг. пчелы карпатской расы получили широкое распространение не только в Восточной Европе, но и в Средней Азии, в том числе и в Таджикистане (Гайдар, Левченко, 2003).
Отклонения от стандарта длины и ширины крыла и длины хоботка, обнаруженные при анализе материала, могут определяться не только вкладом трутней местных рас при свободном скрещивании на пасеках, но и адаптациями пчел к местным условиям и флоре конкретного региона. Это подтверждается морфометрическим анализом пчел из Таджикистана и Крыма, особенно пчел с пасеки К. Так, длина верхних крыльев стандарта “Вучковский” соответствует только показателям исследованной Вучковской линии ЗО (табл. 1). Однако ширина крыла пчел ЗО больше, чем стандарт Вучковской линии, а в остальных исследованных выборках – меньше (табл. 1). Отметим укороченные хоботки пчел с таджикских пасек.
Наиболее информативными морфологическими признаками для определения чистоты породы пчел, по нашим данным, можно принять признаки “дискоидальное смещение” и “кубитальный индекс”.
Известно, что карпатская раса имеет от 95 до 100% положительного смещения, до 5% – нейтрального (Гайдар, 2004; Папп и др., 2013; Маннапов и др., 2015). Показатели, близкие к стандарту “Вучковский”, имеют пчелы из ЗО. В то же время семьи с пасек Таджикистана и Крыма, особенно К, содержат особей не только с положительным, но и с нейтральным или отрицательным дискоидальным смещением крыла. По-видимому, этот признак может быть использован для определения степени чистопородности расы конкретного улья.
Признак “кубитальный индекс”, варьирующий у пчел не только с разных пасек, но и из разных ульев, не связан с наличием в выборках признака “отрицательное дискоидальное смещение” (табл. 1). Известно, что при стандарте 33–43% (Гайдар, 2004) показатель данного признака может увеличиваться у пчел из других регионов до 45% (Морев и др., 2013). Также известно, что среднерусская и ряд южных пород пчел обладают высоким кубитальным индексом (от 55 до 65%), в то время как A. m. sossimai и A. m. carnica – низким (30–45%) (Быкова и др., 2016). Высокий кубитальный индекс у пчел из наших выборок (табл. 1, Т1, Т2, К) также может свидетельствовать о вкладе местных трутней других рас в снижение чистопородности пасеки.
Таблица 2.
Молекулярное разнообразие карпатской расы медоносной пчелы Apis mellifera carpatica с пасек разных регионов
| Регион, место сбора | n | Число гаплотипов/ уникальных гаплотипов |
Pi | H | π | Число вариабельных сайтов |
|---|---|---|---|---|---|---|
| К, Ф, У | 9 | 5/2 | 1.5 ± 0.99 | 0.86 ± 0.09 | 0.0009 ± 0.0007 | 4 транзиции |
| Т1, Т2 | 8 | 3/2 | 2 ± 1.26 | 0.71 ± 0.12 | 0.0013 ± 0.0009 | 3 транзиции, 1 трансверсия |
| М, МО, Л | 7 | 4/2 | 1.14 ± 0.84 | 0.71 ± 0.18 | 0.0007 ± 0.0006 | 3 транзиции |
Примечание. Материал обработан с помощью программы ARLEQUIN. n – число образцов, Pi – среднее число попарных различий между гаплотипами, H – гаплотипическое разнообразие, π – нуклеотидная изменчивость. Исследованные пасеки: К, Ф, У – Республика Крым; Т1, Т2 – Республика Таджикистан; М, МО, Л – Центральный федеральный округ России.
Впервые проведенный анализ изменчивости нуклеотидной последовательности полного гена СОI мтДНК в нашем исследовании позволил охарактеризовать другое направление в разнообразии пчел карпатской расы – разнообразие пчелиных маток.
Минимальная генетическая дистанция (0.001) между выборками из разных регионов свидетельствует о единстве карпатской породы по ареалу. В то же время образцы медоносной пчелы из разных регионов характеризуются высоким гаплотипическим и низким нуклеотидным разнообразием (табл. 2). Это может свидетельствовать о быстром росте численности семей пчел карпатской расы от изначально небольшого числа основателей в конкретной пасеке, в случае когда прошло достаточное время для восстановления уровня гаплотипической изменчивости за счет мутаций, но недостаточное для накопления значительных различий между нуклеотидными последовательностями. Подавляющее большинство мутаций затрагивают третье основание в кодонах и не приводят к изменению аминокислотной последовательности, за исключением замены гуанозина на аденозин во второй позиции кодона, что приводит к замещению серина на аспарагин.
Разделение образцов пчел на две группы на филогенетическом дереве может быть связано с временем формирования пасеки. Известно, что тип карпатской пчелы был во многом сформирован на основе линии 77 пчел Закарпаья, которая впоследствии была утеряна (Папп и др., 2013). Мы предполагаем, что пчелы с изолированной пасеки из горно-лесной зоны Крыма (К), привезенные из Таджикистана и входящие в I группу, являются потомками пчел линии 77. Длительное время разведения пчел пасеки этой линии подтверждается большей вариабельностью гена СОI. Стандарт “Вучковский” был создан уже на основе восстановленной пасеки. Его можно отнести к аналогам, но не к прямым потомкам первоначальной линии массового разведения пчел этой расы. Недавнее разведение пчелиных маток от этой линии подтверждается низкой нуклеотидной изменчивостью. Различия в нуклеотидных последовательностях СОI мтДНК позволяют получить информацию о пчелах – основательницах пасек. Предполагается, что чистые линии и любительские пасеки различаются по ряду морфологических признаков (Калашников, 2013). Различия выведенных линий карпатской расы могут быть связаны как с адаптационными возможностями пчелиных семей, так и с искусственным или неуправляемым скрещиванием пчелиных маток (Папп, 2013; Маннапов и др., 2015). Наиболее достоверно выявить линию пчел, которая лежит в основе данной пасеки, по нашим исследованиям, может аминокислотный состав белка СОI мтДНК.
Комплексный подход к изучению разнообразия пчел карпатской расы по регионам позволяет оценить как длительность разведения, т.е. относительное время получения пчелиных маток – основательниц пасеки, так и степень чистопородности пчел пасеки при длительном разведении, по отклонению от стандарта выбранных морфологических признаков.
Авторы выражают благодарность А.В. Ивашову (Крымский федеральный университет) за ценные идеи по проведению исследований, И.Г. Мещерскому (ИПЭЭ РАН) за помощь в подборе праймеров, А.А. Варшавскому (ИПЭЭ РАН) за оформление карты сбора материала, А.Б. Махмудову (с. Хистеварз), Е.А. Пивоварову (г. Гжель) и А.В. Королеву (г. Москва) за представленный для анализа материал.
Работа выполнена при финансовой поддержке программы Президиума РАН № 41 “Биоразнообразие природных систем и биологические ресурсы России”.
Список литературы
Быкова Т.О., Триселева Т.А., Ивашов А.В., Сафонкин А.Ф. К оценке морфогенетического разнообразия медоносной пчелы Apis mellifera L. из горно-лесной зоны Крыма // Изв. РАН. Сер. биол. 2016. № 6. С. 625−630.
Гайдар В.А. Морфоэтологический стандарт карпатских пчел // Пчеловодство. 2004. № 6. С. 14−15.
Гайдар В.А., Левченко И.А. Сравнительная оценка карпатских и краинских пчел // Пчеловодство. 2003. № 8. С. 18−20.
Гайни М. Пчеловодство Подкарпатской Руси. Ужгород. 1923. 28 с. http://bdjilka.in.ua/index.php/ru/ component/content/article/79-pchely/90-istoriya-vozniknoveniya-karpatskoj-porody-na-zakarpate
Губин В.А. Карпатская популяция карники // Пчеловодство. 1972. № 5. С. 28−31.
Губин В.А. О морфологическом породном стандарте // Пчеловодство. 1976. № 2. С. 11−12.
Зиновьева Н.А., Форнара М.С., Бородачев А.В., Гладырь Е.А., Лебедев В.И., Акимова С.Н., Кривцов Н.И., Эрнст Л.К. Дифференциация карпатских и краинских пчел с использованием микросателлитов // Пчеловодство. 2013. № 1. С. 14−17.
Калашников А.Е. Изучение дифференциации отечественных популяций медоносной пчелы Apis mellifera и их инфицированности РНК-содержащими вирусами с помощью молекулярно-генетических методов: Автореф. дис. канд. биол. наук. Дубровицы: ГНУ ВИЖ Россельхозакадемии, 2013. 22 с.
Маннапов А.Г., Хоружий Л.И., Симоганов Н.А., Редькова Л.А. Технология производства продукции пчеловодства по законам природного стандарта. М.: Проспект, 2015. С. 24−30.
Морев И.А., Морева Л.Я., Козуб М.А. Экологическая пластичность популяций пчел юга России // Вестн. Тамбов. ун-та. 2013. Т. 18. Вып. 3. С. 890−895.
Папп В.В. Динамика некоторых признаков автохтонных карпатских пчел под действием направленного отбора // Актуальные проблемы интенсивного животноводства. 2013. Вып. 16. Ч. 2. С. 201−206.
Папп В.В., Керек С.С., Кейль Е.I. Поглиблене вивчення деяких морфологічних ознак карпатських бджіл, їх породна характеристика та диференціація за допомогою програмного забезпечення “Beemorph”// Сільський господар. 2013. № 11−12. С. 25−31.
Столповский Ю.А. Популяционно-генетические основы сохранения ресурсов генофондов доместифицированных видов животных: Автореф. дис. докт. биол. наук. М.: ИОГен, 2010. 50 с.
Cэндyляк E. Contributii la studiul genofenotipic al albinelor Apis mellifera carpatica // Cercet. genet. Lucr. 1 simpos. nat. genet. Bucuresti. 1965.
Уфимцева Н.С., Осинцева Л.А. Породы и методы разведения медоносной пчелы, Apis mellifera L. Новосибирск: Биол.-техн. ин-т, 2009. 47 с.
Bandelt H.J., Foster P., Röhl A. Median-joining networks for inferring intraspecific phylogenies // Mol. Biol. Evol. 1999. V. 16. № 1. P. 37−48.
Cauia E., Usurelu D., Magdalena L.M., Cimponeriu D., Apostol P., Siceanu A., Holban A., Gavrila L. Preliminary researches regarding the genetic and morphometric characterization of honeybees (A. mellifera L.) from Romania // Lucrări ştiinţifice Zootehnie şi Biotehnologii, Timişoara. 2008. V. 41. № 2. P. 278−286.
Engel M.S. The taxonomy of recent and fossil honey bees (Hymenoptera: Apidae, Apis) // J. Hym. Res. 1999. V. 8. P. 165−196.
Excoffier L., Lischer H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows // Mol. Ecol. Res. 2011. V. 10. P. 564–567.
Folmer O., Black M., Hoeh W., Lutz R., Vrijenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates // Mol. Mar. Biol. Biotech. 1994. V. 3. № 5. P. 294–299.
Martimianakis S., Klossa-Kilia E., Bouga M., Kilias G. Phylogenetic relationships of Greek Apis mellifera subspecies based on sequencing of mtDNA segments (COI and ND5) // J. Agric. Res. 2011. V. 50. № 1. P. 42–50.
Ozdil F., Ilhan F. Phylogenetic relationship of Turkish Apis mellifera subspecies based on sequencing of mitochondrial cytochrome C oxidase I region // Genet. Mol. Res. 2012. V. 11. № 2. P. 1130–1141.
Ronquist F., Huelsenbeck J.P. MRBAYES 3: Bayesian phylogenetic inference under mixed models // Bioinformatics. 2003. V. 19. P. 1572–1574.
Ruttner F. Biogeography and taxonomy of honeybees. Berlin: Springer Verlag, 1988. 284 p.
Ruttner F. Naturgeschichte der Honigbienen // Ehrenwirth Verlag, Munich. Grenoble; Bukarest: Apimondia, 1992. S. 380−383.
Tamura K., Peterson D., Peterson N., Stecher G., Nei M., Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods // Mol. Biol. Evol. 2011. V. 28. P. 2731–2739.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая