

Известия РАН. Серия биологическая, 2019, № 6, стр. 625-639
Сравнительные особенности питания лошади Пржевальского Equus рrzewalskii, двугорбого верблюда Camelus bactrianus и сайгака Saiga tatarica на степном изолированном пастбище
Б. Д. Абатуров 1, *, Р. Р. Джапова 2, В. Д. Казьмин 3, Е. Ч. Аюшева 2, В. В. Джапова 2
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
2 Калмыцкий государственный университет им. Б.Б. Городовикова
358000 Элиста, ул. Пушкина, 11, Россия
3 Государственный биосфeрный заповедник “Ростовский”
347510 Ростовская обл., пос. Орловский, пер. Чапаевский, 102, Россия
* E-mail: abaturovbd@mail.ru
Поступила в редакцию 22.03.2018
После доработки 27.05.2019
Принята к публикации 03.06.2019
Аннотация
Исследованы кормовые ресурсы, питание и обеспеченность пищей свободнопасущихся двугорбых верблюдов, лошадей Пржевальского и сайгаков на степном огороженном разнотравно-злаковом пастбище. Посезонно оценены состав потребляемых растений, избирательность питания, переваримость и количество потребляемого корма. Отмечено, что все три вида различаются по всем показателям питания.
Кормовая растительность на природных пастбищах неоднородна по питательной ценности. Травянистые растения делят на две различающиеся кормовыми свойствами группы: злаки и разнотравье. Злаки в нашем случае объединяют однодольные растения – злаки и осоки, разнотравье включает в себя двудольные травы, полукустарнички и кустарнички. В зарубежных работах с этой целью чаще используются сходные англоязычные термины: Grasses и Forbs (Holechek, 1984) или Grass и Browse (The Ecology…, 2008).
Известно, что разнотравье превосходит злаки по питательной ценности, в частности листья большинства видов пастбищного разнотравья богаче злаковых по содержанию белков и в целом по многообразию синтезированных в растении соединений (вторичных метаболитов), многие из которых токсичны. Большинство злаков характеризуется более низкой переваримостью, что вызвано повышенным содержанием в их клеточных стенках лигнина и кремния, увеличивающих механическую прочность растительных тканей, ограничивающих пищеварение и выполняющих у пастбищных растений защитные функции по отношению к их потребителям (Van Soest, Jones, 1968; Holechek, 1984; McNаughton et al., 1985; Колесников, Абатуров, 1997; Скопин, 2003; Massey et al., 2007). С кормовыми особенностями пастбищной растительности и различной способностью животных ее утилизировать связано деление растительноядных млекопитающих по типам питания и пищевой специализации. Среди копытных выделяются три группы: высокоизбирательные потребители разнотравья, т.е. легкоусвояемых растительных кормов; менее избирательные потребители преимущественно злаковых растений, отличающихся более низкой питательной ценностью; большая группа животных с промежуточным питанием (Holechek, 1984; Hofmann, 1989, 1991). Для такой специализации характерна адаптивная природа, связанная со спецификой строения и физиологии пищеварительного аппарата различных по типам питания животных. Известно, что млекопитающие с разной морфологией пищеварительного аппарата неодинаково реагируют на разные по кормовому качеству растения. Лошади, для которых характерна заднекишечная ферментация потребленного корма, успешно поедают и даже предпочитают грубые кормовые растения, в том числе злаки. В то же время для многих видов жвачных копытных со сложным четырехкамерным желудком более привлекательно разнотравье (Salter et al., 1979; Hofmann,1991; Абатуров и др., 1996; Menard et al., 2002; Позднякова и др., 2011). Давно замечено, что, например, ковыли – одни из самых распространенных степных злаков – поедаются лучше лошадьми, хуже овцами и крупным рогатым скотом, а хуже всего верблюдами (Ларин и др., 1950). К последней категории относятся и сайгаки, которые в опытах по кормлению набором разных видов растений, а также в полевых экспериментах со свободной пастьбой всегда избегали поедание не только ковылей, но и злаков вообще (Абатуров и др., 1982, 1998, 2005). Очевидно, что и требования к качеству кормовых ресурсов у животных с разной спецификой питания неодинаковы. Это служит причиной различий в предпочтении определенных кормов и выраженной у большинства травоядных избирательности питания.
Особую важность избирательность питания приобретает в многовидовых сообществах пастбищных животных при совместной пастьбе видов с разной спецификой питания. Мы сравнили питание трех видов млекопитающих: двугорбого верблюда Camelus bactrianus, лошади Пржевальского Equus przewalskii и сайгака Saiga tatarica, различающихся типами пищеварения и пасущихся на одинаковом пастбище.
Цель работы – оценить различия в выборе и утилизации поедаемых растений при пастьбе этих видов млекопитающих на естественном степном пастбище, установить требования животных к среде обитания и их обеспеченности кормом с учетом конкуренции между ними за кормовые ресурсы.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования были проведены в 2015 и 2016 гг. на степном пастбище вольерного комплекса Центра редких животных европейских степей Ассоциации “Живая природа степей”, расположенного в Кумо-Манычской впадине в подзоне сухих дерновинно-злаковых степей (46°24′ с.ш., 42°37′ в.д.). Вольерный комплекс занимает изолированную забором и водоемами территорию площадью ~15 км2 с абсолютными высотами ~30 м над уровнем моря. Территория комплекса слабохолмистая, расположена на водоразделе между озерными котловинами Маныч и Грузское, имеет сложную геоморфологическую структуру, включает в себя солончаковые речные русла, неоднородна по составу растительных сообществ.
Проведено сравнение состава, соотношения в рационе и предпочтения потребляемых видов растений тремя видами копытных млекопитающих: представителем моногастричных нежвачных копытных (лошадью Пржевальского) и двумя видами жвачных (сайгаком и двугорбым верблюдом), различающихся строением желудка. Сайгаки и лошади Пржевальского выпасались на отдельном степном участке (~80 га), отгороженном забором от остальной территории. Верблюды паслись за пределами этого участка.
Геоботанические исследования проводили общепринятыми методами геоботанического обследования в апреле, июне–июле и сентябре–октябре 2015 и 2016 гг. и в конце зимнего (снежного) периода (март) 2017 г. Растительность описывали на стандартной площади размером не менее 100 м2. Русские названия видов растений приведены в соответствии с принятыми в работе Станкова и Талиева (Станков, Талиев, 1957), латинские – в сводке Черепанова (Черепанов, 1995). Учеты надземной растительной массы проводили на укосных площадках размером 50 × 50 см. Материал укосов разобран по видам растений, высушен до постоянной массы при 90°С. На огороженном для лошадей Пржевальского и сайгаков участке наблюдения и учеты проведены во всех представленных фитоценозах: однолетниково-житняковом, полынно-злаковом, однолетниковом и однолетниково-безостокостровом на каштановых почвах, полынковом на лугово-каштановых почвах. На территории вольерного комплекса в местах пастьбы верблюдов исследовали следующие фитоценозы: однолетниково-злаковый с разнотравьем на каштановых почвах, лерхополынный на солонцах каштановых средних, разнотравно-злаковый на лугово-каштановых почвах и однолетниковую залежь на каштановых почвах. На основании этих данных с учетом доли площади выделенных фитоценозов была определена общая надземная масса растительности и ее основных видов на пастбище в разные сезоны года.
Питание животных исследовали в те же сроки: весной, летом, осенью и в конце зимнего (снежного) периода (март). Наблюдение за пастьбой животных вели в светлое время в течение 3 сут. Состав потребляемых растений и их соотношение в рационе определяли методом микрогистологического кутикулярно-копрологического анализа (Stewart, 1967; Розенфельд, Ларионов, 2006; Розенфельд, 2011; и др.). Образцы фекалий для анализов брали непосредственно во время наблюдения за пастьбой верблюдов и лошадей Пржевальского (от трех взрослых особей каждого вида). Свежеотложенные еще непросохшие фекалии сайгаков собирали в местах пастьбы животного. Соотношение видов растений в рационе оценивали под микроскопом по доле фрагментов кутикулы каждого вида растений в собранных образцах фекалий. Для видовой идентификации кутикулярных фрагментов в фекалиях готовили атлас фотографий клеточной структуры кутикулы массовых степных растений этого пастбища.
Для оценки предпочтения в выборе видов кормовых растений на пастбище использовали индекс избирательности (ИИ), который в нашем случае представляет собой отношение средней доли вида растения в составе рациона (%) к его доле (%) в составе растений на пастбище. Понятно, что чем меньше полученное значение ИИ, тем ниже предпочтение в выборе данного вида (избирательность). Очевидно, при равных долях вида в фитоценозе и в рационе, т.е. при ИИ = 1, избирательность отсутствовала, при ИИ > 1 растение поедалось избирательно, при ИИ < 1 животные избегали его потребление. Отметим, что этот способ расчета ИИ, примененный ранее Шорыгиным (1939), отличен от расчетов избирательности, использованных в других случаях (Ивлев, 1955; Jacobs, 1974).
Определение переваримости выполняли непрямым методом по соотношению инертных (непереваримых) веществ в корме и фекалиях (Абатуров и др., 2003). В качестве инертного вещества использовали органогенный кремний, содержащийся в тканях поедаемых растений (Ely et al., 1953; Streeter, 1969; Колесников, Абатуров, 1997). Для оценки концентрации кремния в съеденном корме использовали его средневзвешенное содержание в рационе путем учета доли потребленных видов растений и концентрации кремния в каждом из них. Коэффициент переваримости сухого вещества корма (D, %) рассчитывали по формуле
где ${v}$ и f – концентрации кремния соответственно в корме и в фекалиях (Абатуров и др., 2003).
Суточное потребление корма (С, г/особь, сухая масса) свободнопасущимися верблюдами и лошадьми Пржевальского рассчитывали из количества выделенных фекалий и переваримости по формуле
где F – суточное выделение фекалий (г/особь, сухая масса), D – коэффициент переваримости (%) (Абатуров и др., 2003). Для определения суточного выделения фекалий устанавливали средний интервал между дефекациями у исследуемых животных и среднюю массу фекалий за одну дефекацию. Интервал между дефекациями определяли путем хронометража при непрерывных визуальных наблюдениях за отдельным животным. Определяли массу экскрементов за одну дефекацию, для чего фекалии собирали сразу же после каждой дефекации у животных, находившихся под наблюдением. Образцы фекалий высушивали при 90°С до постоянной массы для пересчета взвешенных фекалий на сухую массу. Суточное выделение фекалий рассчитывали по числу дефекаций за 1 сут и их средней массе за одну дефекацию. Оценка количества фекалий, откладываемых свободнопасущимися сайгаками, невозможна, поэтому суточное потребление корма сайгаками не определяли. Ранее такие определения были проведены с использованием калосборных мешков на прирученных сайгаках (Абатуров и др., 1997, 1998; Abaturov, Subbotin, 2011). Свежие образцы экскрементов использовали для последующего химического анализа.
Определяли количество переваримой и обменной энергии в потребленном корме. Для оценки содержания энергии в корме были использованы результаты зоотехнического анализа (жиры, протеин, углеводы) и расчета энергетической ценности пастбищной растительности, полученные ранее для растительности степного пастбища Северного Прикаспия (Абатуров, 1999). Среднее содержание энергии в пастбищной растительности в разные месяцы года (с апреля по декабрь) по этим данным составило 17.32 ± 0.06 кДж/г. Для перевода переваримой энергии в обменную использовали для верблюдов коэффициент 0.8, применяемый для крупного рогатого скота и овец, а для лошадей – 0.9 (Мак-Дональд и др., 1970; Нормы…, 2003).
Химический анализ содержания питательных веществ в корме и фекалиях был выполнен стандартными методами (Инструкция…, 1968). Анализ содержания кремния проведен на основе кислотного (HNO3) гидролиза образцов растений и фекалий с использованием колориметрического метода кремнемолибденовой сини (Колесников, Абатуров, 1997; Колесников, 2001; Гинс и др., 2012). Этот метод позволяет исключить из результатов анализа кристаллический кремний, поступающий в корм из почвы и пыли и искажающий показатель переваримости.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Характеристика растительного покрова. Степное пастбище, на котором вольно выпасалось стадо верблюдов (~20 особей), расположено на водоразделе между двумя водоемами (озера Маныч и Грузское) с различающимися по структуре растительными сообществами на вершине водораздела, его южном и северном склонах (табл. 1). Пастбище включает в себя большой участок (~214 га) с залежной растительностью на бывшей пашне.
Таблица 1.
Видовой состав растений и средняя фитомасса на пастбище верблюдов в 2015–2016 гг. (г/м2 сухой массы, M ± m, n = 28)
| Растения | Апрель | Июнь–июль | Сентябрь–октябрь | |||
|---|---|---|---|---|---|---|
| г/м2 | % | г/м2 | % | г/м2 | % | |
| Злаки, осоки | ||||||
| Stipa lessingiana | 7 ± 3.1 | 2.8 ± 1.1 | 7.6 ± 3 | 3.1 ±1.2 | 2.6 ± 1 | 1.0 ± 0.3 |
| Stipa capillata | 0 | 0 | 15 ± 6.7 | 6.1 ±2.7 | 12.5 ± 4.2 | 4.7 ± 1.5 |
| Festuca valesiaca | 7.6 ± 2.8 | 2.9 ± 1.1 | 6 ± 2 | 2.4 ±0.8 | 2.9 ± 1 | 1.1 ± 0.3 |
| Agropyron desertorum | 51 ± 14.2 | 19.5 ± 5.4 | 43.8 ± 16.9 | 17.8 ±6.8 | 31 ± 10.4 | 11.5 ± 3.9 |
| Elytrigia repens | 2.4 ± 2.2 | 0.9 ± 0.8 | 1.7 ± 1.7 | 0.7 ±0.7 | 0.3 ± 0.3 | 0.1 ± 0.1 |
| Anisantha tectorum | 15.9 ± 5 | 6.1 ± 1.9 | 1.5 ± 1.1 | 0.6 ±0.4 | 3.4 ± 1.4 | 1.3 ± 0.5 |
| Poa bulbosa | 5.4 ± 3.5 | 2.1 ± 1.3 | 8.3 ± 2.7 | 3.4 ±1.1 | 1.2 ± 1.2 | 0.4 ± 0.4 |
| Carex stenophylla | 2.9 ± 2.9 | 1.1 ± 1.1 | 1.4 ± 1.4 | 0.6 ±0.5 | 0.1 ± 0.1 | 0.1 ±0.1 |
| Прочие виды злаков | 10.5 ± 5 | 4 ± 1.9 | 0 | 0 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| Всего (злаки, осоки) | 102.7 ± 17.2 | 39.4 ± 6.5 | 85.3 ± 18.9 | 34.7 ± 7.6 | 54.1 ± 11.4 | 20.1 ± 4.2 |
| Разнотравье | ||||||
| Medicago romanica | 0.4 ± 0.4 | 0.2 ± 0.1 | 0 | 0 | 0 | 0 |
| Bassia sedoides | 0 | 0 | 0.3 ± 0.1 | 0.1 ±0.1 | 52.1 ± 21.3 | 19.4 ± 8.1 |
| Camphorosma monspeliaca | 0 | 0 | 0.6 ± 0.6 | 0.2 ±0.2 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| Kochia prostrata | 7.5 ± 1.2 | 2.9 ± 0.4 | 6.8 ± 3.0 | 2.8 ±1.2 | 7.5 ± 2.9 | 2.8 ± 1.1 |
| Atriplex tatarica | 18.0 ± 7.6 | 6.9 ± .9 | 34 ± 15.2 | 14 ±6.1 | 67.3 ± 27.3 | 25 ± 10.4 |
| Chenopodium album | 0.2 ± 0.1 | 0.1 ± 0.1 | 0 | 0 | 0 | 0 |
| Erophyla verna | 3.8 ± 0.3 | 1.5 ± 0.1 | 0.2 ± 0.2 | 0.1 ±0.1 | 0 | 0 |
| Descurainia sophia | 18.6 ± 4.8 | 7.1 ± 1.8 | 13.8 ± 6.7 | 5.6 ±2.7 | 4.9 ± 2.0 | 1.8 ± 0.7 |
| Lamium amplexicaule | 1.5 ± 0.6 | 0.6 ± 0.2 | 0.1 ± 0.1 | 0.1 ±0.1 | 0 | 0 |
| Carduus acanthoides | 5.7 ± 2.2 | 2.2 ± 0.8 | 3.9 ± 1.6 | 1.6 ±0.6 | 0.4 ± 0.4 | 0.1 ± 0.1 |
| Sisymbrium loeselii | 3.7 ± 1.4 | 1.4 ± 0.5 | 0.7 ± 0.3 | 0.3 ±0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| Centaurea diffussa | 3 ± 1.8 | 1.4 ± 0.7 | 3.6 ± 1.8 | 1.5 ±0.7 | 0 | 0 |
| Artemisia austriaca | 5.3 ± 3.4 | 2.0 ± 1.3 | 3.3 ± 2.1 | 1.3 ±0.8 | 46.9 ± 7.9 | 17.4 ± 3.0 |
| Artemisia lerchiana | 6.9 ± 5.5 | 2.6 ± 2.1 | 12.7 ± 9.1 | 5.2 ±3.7 | 8.3 ± 7.8 | 3.1 ± 2.9 |
| Tanacetum millefolium | 1.9 ± 0.9 | 0.7 ± 0.3 | 1.7 ± 0.8 | 0.7 ±0.3 | 0 | 0 |
| Holosteum umbellatum | 0.5 ± 0.2 | 0.2 ± 0.1 | 0.1 ± 0.1 | 0.1 ±0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| Galatella villosa | 0.4 ± 0.4 | 0.2 ± 0.1 | 10.3 ± 4.2 | 4.2 ±1.7 | 24.7 ± 9.0 | 9.2 ± 3.4 |
| Lappula squarrosa | 0.1 ± 0.1 | 0.1 ± 0.1 | 2.9 ± 1.1 | 1.2 ±0.4 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| Lepidium perfoliatum | 32 ± 15.5 | 12.2 ± 5.9 | 0.8 ± 0.3 | 0.3 ± 0.1 | 1.9 ± 0.7 | 0.7 ± 0.2 |
| Polygonum patulum | 0 | 0 | 0.3 ± 0.1 | 0.1 ± 0.1 | 0.6 ± 0.2 | 0.2 ± 0.2 |
| Galium verum | 0.5 ± 0.5 | 0.2 ± 0.2 | 0.2 ± 0.2 | 0.1 ± 0.1 | 0 | 0 |
| Galium aparine | 9.0 ± 4.2 | 3.4 ± 1.6 | 0 | 0 | 0 | 0 |
| Senecio vernalis | 14.3 ± 6.8 | 5.4 ± 2.6 | 0 | 0 | 0 | 0 |
| Matricaria recutita | 0.3 ± 0.1 | 0.1 ± 0.1 | 2.1 ± 1.0 | 0.8 ± 0.4 | 0 | 0 |
| Camelina sylvestris | 20.6 ± 10.1 | 7.9 ± 3.8 | 0 | 0 | 0 | 0 |
| Tripleurospermum perforatum | 0 | 0 | 35.2 ± 17.2 | 14.4 ± 7.0 | 0 | 0 |
| Caucalis lappula | 0 | 0 | 10.8 ± 5.3 | 4.4 ± 2.1 | 0 | 0 |
| Consolida regalis | 0 | 0 | 13.2 ± 6.4 | 5.4 ± 2.6 | 0 | 0 |
| Convolvulus arvensis | 0 | 0 | 1.3 ± 0.6 | 0.5 ± 0.2 | 0 | 0 |
| Прочие виды разнотравья | 2.9 ± 1.2 | 1.1 ± 0.4 | 1.1 ± 0.5 | 0.4 ± 0.1 | 0 | 0 |
| всего (разнотравье) | 165.2 ± 23.2 | 60.6 ± 8.8 | 160 ± 27.0 | 65.4 ±11.1 | 215 ± 37.6 | 79.9 ± 14.3 |
| Всего | 268 ± 28.8 | 100 | 246 ± 33.4 | 100 | 269 ± 39.2 | 100 |
Примечание. M – среднее значение совокупности данных, m – стандартная ошибка среднего значения, n – суммарная численность каждой совокупности; для табл. 1–5.
Растительный покров залежи представлен преимущественно бурьянистыми однолетниковыми растительными сообществами. В апреле в травостое доминируют неравноцветник кровельный Anisantha tectorum, дескурайния Софьи Descurainia Sophia, клоповник пронзеннолистный Lepidium perfoliatum, гулявник Лезеля Sisymbrium loeselii, крестовник весенний Senecio vernalis. В июне–июле и в сентябре–октябре активно вегетируют, цветут и плодоносят лебеда татарская Atriplex tatarica, вьюнок полевой Convolvulus arvensis, трехреберник продырявленный Tripleurospermum perforatum, сокирки полевые Consolida regalis, бассия очитковидная Bassia sedoides, горец отклоненный Polygonum patulum. Надземная масса однолетниковых сообществ 300–400 г/м2.
Целинная растительность вершины и склонов водораздела сложена растительным комплексом, состоящим из трех компонентов с разной долей площади каждого: однолетниково-дерновиннозлакового с разнотравьем на каштановых почвах (65–70%), лерхополынного на солонцах средних (10–15%), разнотравно-злакового на лугово-каштановых почвах (15–20%). В однолетниково-дерновинно-злаковом с разнотравьем фитоценозе доминируют злаки: житняк пустынный Agropyron desertorum, овсяница валисская Festuca valesiaca, ковыль Лессинга Stipa lessingiana, среди эфемеров – неравноцветник кровельный A. tectorum, веснянка весенняя Erophyla verna, крестовник весенний Senecio vernalis. Разнотравье представлено преимущественно видами семейства зонтичных (малабайла душистая Malabaila graveolens, прангос противозубной Prangos odontalgica). Надземная масса однолетниково-дерновинно-злакового с разнотравьем фитоценоза 160–310 г/м2. В лерхополынном фитоценозе доминирует полынь Лерха Artemisia lerchiana, весной обилен эфемероид мятлик луковичный Poa bulbosa, летом и осенью – камфоросма монспелийская Camphorosma monspeliaca, кохия стелющаяся Kochia prostrata. Надземная масса лерхополынного фитоценоза 53–80 г/м2. В разнотравно-злаковом фитоценозе на лугово-каштановых почвах доминируют многолетние злаки: ковыль Лессинга S. lessingiana, ковыль волосовидный S. capillata, житняк гребневидный Agropyron pectinatum, типчак F. valesiaca, пырей ползучий Elytrigia repens. Разнотравье представлено тысячелистником обыкновенным Achillea millefolium, чертополохом колючим Carduus acanthoides, полынью австрийской A. austriaca. Надземная фитомасса 150–390 г/м2.
Участок вольерного комплекса размером 80 га, огороженный забором и выделенный для лошади Пржевальского (10 взрослых животных) и сайгака (15 особей), имеет небольшой уклон к центру огороженной территории, где находится русло соленой речки. Растительность большей части этого участка занята сообществами на бывшей залежи с комплексным растительным покровом, который включает в себя однолетниково-безостокостровый и однолетниково-житняковый фитоценоз (50%), однолетниковый (40%), злаково-полынный (полынь австрийская, полынь таврическая) на каштановых почвах, полынно-бескильницевый полынковый на лугово-каштановыхых почвах (10%). На небольшой нераспахивамой части территории растительность представлена полынково-типчаковым фитоценозом.
Наблюдения и учеты проведены в однолетниково-безостокостровом, однолетниково-житняковом, однолетниковом фитоценозах на каштановых почвах, полынковом и полынно-злаковом на лугово-каштановых почвах (табл. 2). Надземная масса однолетниково-безостокострового фитоценоза на каштановых почвах 150–275 г/м2. В структуре укосов преобладает костер безостый, дескурайния Софьи, мятлик луковичный и клоповник пронзеннолистный, вероника весенняя, резушка Таля, бурачок пустынный, пастушья сумка, тысячелистник обыкновенный, горчак розовый, мышиный горошек, которые осенью сменяются лебедой татарской, горцом отклоненным.
Таблица 2.
Видовой состав и средняя надземная фитомасса на пастбище сайгаков и лошадей Пржевальского в 2015–2016 гг. (г/м2 сухой массы, M ± m, n = 32)
| Растения | Апрель | Июнь–июль | Сентябрь–октябрь | |||
|---|---|---|---|---|---|---|
| г/м2 | % | г/м2 | % | г/м2 | % | |
| Злаки, осоки | ||||||
| Stipa sp.sp. | 9.4 ± 6.9 | 6.6 ± 4.8 | 36.8 ± 17.8 | 18.1 ± 10.2 | 24.1 ±9.8 | 12.9 ± 6.11 |
| Festuca valesiaca | 10.0 ± 3.5 | 7.0 ± 2.4 | 7.9 ± 2.1 | 3.9 ± 1.2 | 3.8 ± 1.5 | 2.0 ± 0.9 |
| Bromus inermis | 19.5 ± 19.5 | 12.1 ± 12.1 | 27.9 ±27.9 | 13.7 ± 13.7 | 26.2 ± 26.2 | 14.1 ± 14.1 |
| Agropyron desertorum | 31.5 ± 19.9 | 22.2 ± 13.4 | 40.7 ± 30.1 | 20.0 ± 17.2 | 46.5 ± 19.0 | 25 ± 15.8 |
| Elytrigia repens | 6.1 ± 4.8 | 4.3 ± 3.4 | 4.6 ± 3.8 | 2.2 ± 2.1 | 1.0 ± 0.8 | 0.5 ± 0.5 |
| Poa bulbosa | 13.1 ± 6.6 | 9.2 ± 4.6 | 11.1 ± 2.3 | 5.5 ± 1.3 | 4.0 ± 3.2 | 2.2 ± 2.0 |
| Carex stenophylla | 7.8 ± 6.4 | 5.5 ± 3.2 | 3.7 ± 3.0 | 1.8 ± 1.7 | 0.3 ± 0.3 | 0.2 ± 0.2 |
| Прочие злаки | 6.9 ± 3.2 | 4.8 ± 2.2 | 1.1 ± 0.8 | 0.5 ± 0.4 | 0.4 ± 0.2 | 0.3 ± 0.1 |
| Всего (злаки, осоки) | 104 ± 23.9 | 64.5 ± 16.8 | 133.8 ± 35.4 | 65.9 ± 20.2 | 106 ± 21.8 | 57 ± 13.6 |
| Разнотравье | ||||||
| Lepídium perfoliatum | 0.2 ± 0.2 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.05 ± 0.05 | 0 | 0 |
| Kochia prostrata | 0 | 0 | 1.3 ± 1.0 | 0.7 ± 0.5 | 0 | 0 |
| Camphorosma monspeliaca | 0 | 0 | 1.6 ± 1.3 | 0.9 ± 0.7 | 0.2 ± 0.2 | 0.1 ± 0.1 |
| Lamium amplexicaule | 1.0 ± 0.7 | 0.7 ± 0.5 | 0.1 ± 0.1 | 0.05 ± 0.05 | 0 | 0 |
| Atriplex tatarica | 1.0 ± 0.8 | 0.7 ± 0.6 | 2.4 ± 2.0 | 1.2 ± 1.1 | 0.6 ± 0.5 | 0.3 ± 0.3 |
| Bassia sedoides | 0 | 0 | 0.9 ± 0.7 | 0.5 ± 0.4 | 0 | 0 |
| Medicago romanica | 1.0 ± 0.8 | 0.7 ± 0.5 | 0.1 ± 0.1 | 0.05 ± 0.05 | 0 | 0 |
| Lapulla squarrosa | 0.2 ± 0.1 | 0.1 ± 0.07 | 0.6 ± 0.2 | 0.3 ± 0.1 | 0.3 ± 0.2 | 0.1 ± 0.1 |
| Polygonum patulum | 0 | 0 | 0.4 ± 0.1 | 0.2 ± 0.05 | 0.47 ± 0.4 | 0.3 ± 0.2 |
| Descurainia sophia | 4.1 ± 3.3 | 2.9 ± 2.3 | 0 | 0 | 0 | 0 |
| Erophila verna | 2.2 ± 1.2 | 1.5 ± 0.8 | 0.4 ± 0.3 | 0.2 ± 0.1 | 0 | 0 |
| Achillea millefolium | 0.3 ± 0.2 | 0.2 ± 0.1 | 0 | 0 | 0 | 0 |
| Sisymbrium loeselii | 2.2 ± 1.7 | 1.5 ± 1.2 | 0.2 ± 0.1 | 0.1 ± 0.05 | 0.43 ± 0.35 | 0.2 ± 0.2 |
| Artemisia austriaca | 10.9 ± 7.0 | 7.7 ± 4.9 | 6.8± 4.4 | 3.4 ± 2.5 | 24.3 ± 9.9 | 13.1 ± 6.1 |
| Artemisia lerchiana | 12.2 ± 10.0 | 8.6 ± 7.0 | 28.5 ± 19.4 | 14.0 ± 11.1 | 27.6 ± 20.6 | 14.8 ± 12.8 |
| Galatella villosa | 1.2 ± 0.9 | 0.8 ± 0.6 | 11.4 ± 5.6 | 5.6 ± 3.2 | 24.9 ± 13.8 | 13.4 ± 8.6 |
| Carduus acanthoides | 8.4 ± 3.6 | 5.9 ± 2.5 | 6.1 ± 2.85 | 3.0 ± 1.6 | 1.2 ± 0.95 | 0.6 ± 0.6 |
| Прочее разнотравье | 14.9 ± 3.6 | 10.5 ± 2.5 | 8.0 ± 3.0 | 3.9 ± 1.7 | 0.06 ± 0.05 | 0 |
| Всего (разнотравье) | 57.3 ± 13.4 | 40.4 ± 9.4 | 68.9 ± 21.1 | 33.4 ± 12.1 | 80.1 ± 33.8 | 43 ± 21.0 |
| Всего | 142 ± 27.4 | 100 | 203 ± 41.2 | 100 | 186 ± 40.2 | 100 |
В однолетниково-житняковом фитоценозе доминант травостоя – житняк пустынный. Весной и летом вегетируют подмаренник цепкий, вероника весенняя, мышиный горошек, клоповник широколистный и пронзеннолистный. Летом и осенью цветут и плодоносят лебеда татарская, горец отклоненный, полынь австрийская, бассия очитковидная. Надземная масса однолетниково-житнякового фитоценоза на каштановых почвах 220–310 г/м2.
Однолетниковый фитоценоз – компонент всех растительных комплексов огороженного участка. Весной в травостое доминируют дескурайния Софьи, гулявник Лезеля, костер кровельный; летом их сменяют лебеда татарская, бассия очитковидная, горец отклоненный, которые до поздней осени цветут и плодоносят. Надземная масса однолетниковых фитоценозов составляет 240–270 г/м2.
Полынково-злаковые, злаково-полынковые и полынковые фитоценозы на лугово-каштановых почвах представляют собой разные стадии восстановления залежной растительности. Их участие в травостое на участке сайгаков и лошадей Пржевальского не превышает 20%. Кроме многолетних злаков – овсяницы валисской, пырея ползучего, житняка гребневидного – развиваются эфемер веснянка весенняя, эфемероид мятлик луковичный, многолетнее разнотравье – ромашник тысячелистниковый, тысячелистник обыкновенный, чертополох курчавый. Полынь австрийская, полынь Лерха максимального развития достигают летом и осенью. Надземная масса фитоценозов 220–310 г/м2.
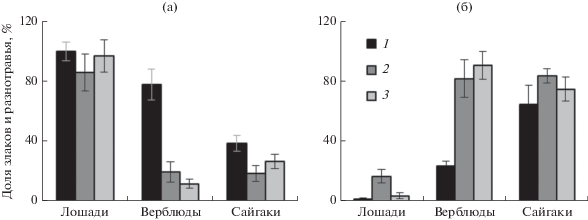
Состав рациона. Сравниваемые виды копытных во все периоды исследований кардинально различались по составу потребляемых растений. В питании лошади Пржевальского преобладали злаки, доля которых в составе рациона варьировала от 85 до 100%, при этом наиболее активно поедались овсяница валисская и ковыли с долей в рационе соответственно до 63 и 30% (табл. 3). Преимущество злаков в питании сочеталось с их избирательным поеданием во все сезоны (во все сезоны ИИ = 1.3–2.2) (табл. 4). Верблюды и сайгаки кардинально отличаются от лошадей по отношению к злакам. Участие злаков в составе рациона верблюдов летом и осенью не превышало 11–19%, причем верблюды в это время явно избегали их потребление (ИИ = 0.5–0.6). Лишь весной в начальный период вегетации молодые растения злаков с успехом поедались верблюдами: участие злаков в питании в это время достигало 77% при их отчетливом предпочтении (ИИ = 1.9). Явное отрицательное отношение к злакам свойственно сайгакам во все сезоны. Доля злаков в питании варьировала от 18 до 38%, при этом животные явно избегали их выбор (ИИ = 0.3–0.5). Лишь типчаку среди злаков на данном пастбище отдавалось безусловное предпочтение (ИИ = 3.3–6.0). Хорошо известно, что для сайгаков злаки – грубый, трудно переваримый корм; они явно предпочитают разнотравье (Абатуров, 1999). Тем не менее на исследованном пастбище, где в растительных сообществах летом и осенью злаки господствуют (57–65%), а разнотравье уступает им по обилию (35–40%), сайгаки вынужденно потребляют злаки, поэтому доля злаков в составе рациона достаточно ощутима (табл. 3).
Таблица 3.
Состав рациона верблюдов, лошади Пржевальского и сайгаков в разные сезоны 2015–2016 гг. (%, M ± m)
| Растение | Апрель | Июнь–июль | Сентябрь–октябрь | ||||||
|---|---|---|---|---|---|---|---|---|---|
| верблюды (n = 7) |
лошади (n = 4) |
сайгаки (n = 4) |
верблюды (n = 7) | лошади (n = 7) | сайгаки (n = 4) |
верблюды (n = 6) | лошади (n = 6) | сайгаки (n = 4) |
|
| Злаки осоки | |||||||||
| Stipa sр.sp. | 28 ± 8.1 | 12 ± 3.5 | 1 ± 0.65 | 0.3 ± 0.3 | 28 ± 8.9 | 0 | 0 | 30 ± 7.6 | 0 |
| Festuca valesiaca | 21 ± 4.9 | 63 ± 3.5 | 22 ± 4.4 | 0.8 ± 0.5 | 27 ± 5.8 | 15 ± 5.1 | 3 ± 2.0 | 36 ± 1.8 | 14 ± 4.1 |
| Bromus inermis | 11 ± 1.9 | 12 ± 2.6 | 0 | 9 ± 5.8. | 6 ± 2.7 | 0 | 6 ± 2.25 | 3 ± 1.7 | 6 ± 0.9 |
| Agropyron sp. | 7 ± 3.3 | 10 ± 2.3 | 8 ± 0.96 | 5 ± 2.7 | 7 ± 3.7 | 3 ± 1.4 | 0 | 11 ± 6.1 | 6 ± 2.2 |
| Koeleria sp. | 1 ± 1.10 | 0 | 1 ± 0.65 | 3 ± 1.9 | 5 ± 2.6 | 0 | 1 ± 0.96 | 2 ± 1.5 | 0 |
| Elytrigia sp. | 1 ± 0.7 | 1 ± 1.0 | 0 | 0.3 ± 0.3 | 2 ± 0.8 | 0 | 1 ± 0.6 | 0 | 0 |
| Leymus sp. | 2 ± 1.0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Poa sp. | 1 ± 0.61 | 1 ± 1.0 | 0 | 0 | 1 ± 1.0 | 0 | 0 | 1 ± 1.3 | 0 |
| Carex sp. | 5 ± 1.4 | 1 ± 1.1 | 6 ± 2.5 | 1 ± 0.3 | 9 ± 2.6 | 0 | 1 ± 0.72 | 13 ± 3.7 | 0 |
| Всего (злаки, осоки) | 77 ± 10.4 | 100 ± 6.3 | 38 ± 5.2 | 19 ± 6.7 | 85 ± 12.2 | 18 ± 5.3 | 11 ± 3.1 | 96 ± 10.9 | 26 ± 4.7 |
| Разнотравье | |||||||||
| Galium sp. | 1 ± 0.5 | 0 | 0 | 2 ± 1.2 | 1 ± 1.0 | 5 ± 1.5 | 3 ± 1.1 | 0 | 7 ± 0.6 |
| Lepídium sp. | 0 | 0 | 0 | 0.4 ± 0.4 | 0 | 2 ± 1.7 | 0 | 0 | 0 |
| Kochia prostrata | 0 | 0 | 0 | 31 ± 10.3 | 12 ± 4.2 | 63 ± 3.4 | 29 ± 12.2 | 1 ± 1.3 | 48 ± 6.6 |
| Chenopodium sp. | 2 ± 1.6 | 0 | 0 | 4 ± 2.4 | 0.3 ± 0.3 | 0 | 4 ± 2.2 | 0 | 0 |
| Camphorosma sp. | 7 ± 1.3 | 0.75 ± 0.65 | 2 ± 1.3 | 0 | 0 | 1 ± 0.86 | 1 ± 0.8 | 0 | 11 ± 3.8 |
| Lamium sp. | 8 ± 3.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Atriplex sp. | 2 ± 1.3 | 0 | 1 ± 0.65 | 33 ± 7.3 | 0 | 5 ± 1.5 | 29 ± 11.7 | 0 | 0 |
| Limonium sp. | 0 | 0 | 0 | 0.3 ± 0.3 | 0 | 0 | 0 | 0 | 3 ± 1.8 |
| Bassia sp. | 0 | 0 | 0 | 1 ± 0.7 | 0 | 1 ± 0.6 | 2 ± 1.1 | 0 | 0 |
| Medicago sp. | 0 | 0 | 0 | 1 ± 1.0 | 0 | 0 | 0 | 0 | 0 |
| Lapulla sp. | 0 | 0 | 0 | 4 ± 2.6 | 0 | 0 | 0 | 0 | 0 |
| Salsola sp. | 3 ± 2.1 | 0 | 0 | 3 ± 2.0 | 2 ± 1.1 | 0 | 10 ± 6.0 | 2 ± 1.2 | 0 |
| Suaeda sp. | 0 | 0 | 0 | 0 | 0 | 0 | 5 ± 2.3 | 0 | 5 ± 1.8 |
| Polygonum sp. | 0 | 0 | 0 | 0.5 ± 0.5 | 0 | 0 | 4 ± 2.0 | 0 | 0 |
| Descurainia sp. | 0 | 0 | 25 ± 9.6 | 0 | 0 | 0 | 0 | 0 | 0 |
| Serrufula sp. | 0 | 0 | 0 | 0 | 0 | 0 | 2 ± 2.0 | 0 | 0 |
| Erophila sp. | 0 | 0 | 27 ± 8.2 | 0 | 0 | 0 | 0 | 0 | 0 |
| Achillea sp. | 0 | 0 | 9 ± 1.2 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sisymbrium sp. | 0 | 0 | 0 | 0 | 0 | 6 ± 1.8 | 0 | 0 | 0 |
| Всего (разнотравье) | 23 ± 3.0 | 0.75 ± 0.65 | 64 ± 12.8 | 81 ± 12.5 | 16 ±4.4 | 83 ± 4.8 | 89 ± 9.4 | 3 ± 1.8 | 74 ± 8.0 |
| Всего | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
Таблица 4.
Индекс избирательности (ИИ) в питании верблюдов, лошади Пржевальского и сайгаков по сезонам 2015–2016 гг.
| Растения | Апрель | Июнь–июль | Сентябрь–октябрь | ||||||
|---|---|---|---|---|---|---|---|---|---|
| верблюды | лошади | сайгаки, | верблюды | лошади | сайгаки, | верблюды | лошади | сайгаки | |
| Злаки, осоки | |||||||||
| Stipa sр.sp. | 10.0 | 1.8 | 0.15 | 0.1 | 1.5 | 0 | 0 | 2.3 | 0 |
| Festuca valesiaca | 7.2 | 9 | 3.14 | 0.8 | 6.9 | 3.3 | 2.7 | 9.2 | 6.0 |
| Bromus inermis | 0 | 1 | 0 | 0 | 0.4 | 0 | 0 | 0.2 | 0.59 |
| Agropyron sp. | 0.4 | 0.5 | 0.36 | 0.3 | 0.3 | 0.18 | 0 | 0.6 | 0.24 |
| Koeleria sp. | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 2 | 0 |
| Elytrigia sp. | 1.1 | 0.2 | 0 | 0.4 | 0.9 | 0 | 10 | 0 | 0 |
| Poa sp. | 0.5 | 0.1 | 0 | 0 | 0.2 | 0 | 0 | 0.5 | 0 |
| Carex sp. | 4.5 | 0.2 | 1.1 | 1.7 | 5.0 | 0 | 10 | 7.2 | 0 |
| Всего (злаки, осоки) | 1.9 | 1.5 | 0.5 | 0.5 | 1.3 | 0.27 | 0.6 | 2.2 | 0.4 |
| Разнотравье | |||||||||
| Galium sp. | 0.3 | 0 | 0 | 20 | 1 | 0 | 3 | 0 | 0 |
| Lepídium sp. | 0 | 0 | 0 | 1.3 | 0 | 40 | 0 | 0 | 0 |
| Kochia prostrata | 0 | 0 | 0 | 11.1 | 17 | 90 | 10 | 1 | 48 |
| Camphorosma sp. | 0 | 0 | 2 | 0 | 0 | 1.1 | 5 | 0 | 110 |
| Lamium sp. | 13.3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Atriplex sp. | 0.3 | 0 | 1.42 | 2.3 | 0 | 3.5 | 1.2 | 0 | 7.5 |
| Limonium sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bassia sp. | 0 | 0 | 0 | 10 | 0 | 2.0 | 9.7 | 0 | 0 |
| Lapulla sp. | 0 | 0 | 0 | 3.3 | 0 | 0 | 0 | 0 | 0 |
| Salsola sp. | 3 | 0 | 0 | 0 | 2 | 0 | 0 | 2 | 0 |
| Polygonum sp. | 0 | 0 | 0 | 5 | 0 | 0 | 20 | 0 | 0 |
| Descurainia sp. | 0 | 0 | 8.6 | 0 | 0 | 0 | 0 | 0 | 0 |
| Erophila sp. | 0 | 0 | 18 | 0 | 0 | 0.4 | 0 | 0 | 0 |
| Achillea sp. | 0 | 0 | 45 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sisymbrium sp. | 0 | 0 | 0 | 0 | 0 | 60 | 0 | 0 | 2.15 |
| Всего (разнотравье) | 0.4 | 0.02 | 1.6 | 1.3 | 0.5 | 2.5 | 1.1 | 0.1 | 1. 8 |
По отношению к разнотравью реакция животных диаметрально противоположна. Если лошади практически исключают виды разнотравья из питания (их суммарная доля в рационе не превышает 16%), то у верблюдов и сайгаков разнотравье во все сезоны составляет основу питания (81–89%), причем доля разнотравья у сайгаков не ниже 64% (табл. 3). Преобладание разнотравья в рационе сочетается с его повышенной избирательностью, суммарный ИИ у сайгаков не ниже 1.6, у верблюдов лишь весной в начале вегетации ИИ < 1 и уступает ИИ злаков (табл. 4). Среди разнотравья в питании верблюдов и сайгаков летом и осенью преобладают поздневегетирующие кохия стелющаяся (30–60%) и лебеда татарская (у верблюдов до 30%). Выбор этих растений объясняется их высокой привлекательностью (у сайгаков ИИ кохии >48, у верблюдов >10).
Таким образом, сравнение участия злаков и разнотравья в питании трех сравниваемых видов животных свидетельствует об их разном отношении к этим группам кормовых растений (рис. 1). Для лошадей, отличающихся заднекишечным типом пищеварения, питание почти во все сезоны года ограничивается преимущественно злаками, доля участия которых в питании >80%. Разнотравье, за небольшим исключением, практически неощутимо в их питании (рис. 1). Примечательно, что злаки преобладают в питании лошадей Пржевальского в разных зональных растительных сообществах от пустынь до лесных лугов (Slivinska, Kopij, 2011; Sen et al., 2017). У сайгаков – типичных жвачных со сложным многокамерным желудком – доля злаков в рационе заметно ограничена и питание во все сезоны обеспечивается в основном разнотравьем. Верблюды более разнообразны в выборе корма. Злаки доминируют в их питании только весной, но летом и особенно осенью полное господство в питании, как и у сайгаков, занимает разнотравье, участие злаков резко снижается (рис. 1). Отметим, что среди разнотравья в питании верблюдов (в отличие от питания сайгаков) большую долю составляют сорные бурьянистые однолетние растения.
Рис. 1.
Соотношение злаков (а) и разнотравья (б) в рационах свободнопасущихся лошадей Пржевальского, верблюдов, сайгаков на естественном степном пастбище в разные месяцы 2015–2016 гг. 1–3 – апрель, июнь–июль, сентябрь–октябрь соответственно.
Переваримость кормов. Обеспеченность растительноядных животных кормом зависит прежде всего от такого важного показателя качества корма, как его переваримость. Не так важно количество съеденного корма, как его усвоенная масса, которая определяется его переваримостью. В настоящее время хорошо известно, что злаковые травы обладают более низкой переваримостью по сравнению с разнотравьем (Holechek, 1984; Абатуров, 2005; Clauss et al., 2008). Доля злаков в рационе животных отражается на переваримости потребляемого корма. Сравнение переваримости корма, потребленного свободнопасущимися животными в разные сезоны, показало, что более низкие и относительно постоянные значения переваримости (55–57%) во все сезоны были у лошадей Пржевальского (табл. 5). Именно у лошадей во вcе сезоны, как показано выше, в питании абсолютно преобладали злаки, причем преимущество злаков объясняется не только их обилием в составе пастбищной растительности, а прежде всего их предпочтением (ИИ = 1.3–1.5), несмотря на их пониженную переваримость. Более высокая переваримость была у верблюдов (67–69%) и у сайгаков (60–64%) – избирательных потребителей разнотравья (ИИ = 1.1–2.1). Лишь в апреле значения переваримости у того и другого вида, как и у лошади Пржевальского, оказались заметно сниженными (56–58%), что вызвано повышенным в этот период потреблением ими злаков.
Таблица 5.
Переваримость потребленного корма верблюдами, лошадьми Пржевальского и сайгаками при свободной пастьбе в разные сезоны (M ± m) за 2015–2016 гг.
| Животные | Месяцы | Содержание органогенного кремния, % | Переваримость съеденного корма, % | |
|---|---|---|---|---|
| в рационе (n = 7) |
в фекалиях (n = 6) |
|||
| Верблюды | Апрель | 1.50 ± 0.20 | 3.43 ± 0.019 | 56.2 ± 5.8 |
| Июнь–июль | 1.15 ± 0.18 | 3.49 ± 0.01 | 67.0 ± 5.1 | |
| Сентябрь–октябрь | 1.09 ± 0.19 | 3.53 ± 0.019 | 69.1 ± 5.34 | |
| Лошади Пржевальского | Апрель | 1.64 ± 0.10 | 3.61 ± 0.03 | 54.6 ± 2.8 |
| Июнь–июль | 1.55 ± 0.21 | 3.61 ± 0.03 | 57.1 ± 5.7 | |
| Сентябрь–октябрь | 1.59 ± 0.23 | 3.67 ± 0.01 | 56.7 ± 6.21 | |
| Сайгаки | Апрель | 1.39 ± 0.18 | 3.31 ± 0.02 | 58.0 ± 5.4 |
| Июнь–июль | 1.19 ± 0.1 | 3.37 ± 0.01 | 64.7 ± 2.9 | |
| Сентябрь–октябрь | 1.27 ± 0.12 | 3.22 ± 0.02 | 60.5 ± 3.7 | |
Ранее на примере питания сайгаков было показано, что переваримость растительного корма отрицательно коррелирует с содержанием кремния в съеденной растительности (Абатуров, 1999, 2005). Эта особенность свойственна не только сайгакaм, но и другим растительноядным млекопитающим. Концентрации кремния в разных пастбищных растениях неодинаковы. Более высока она в злаках, что отражается на их низкой переваримости (Абатуров, 1999, 2005). Очевидно, что показатели переваримости должны меняться в соответствии с изменением обилия злаковых трав в питании животных. Оценка зависимости переваримости корма от количества злаков в потребленном корме показала тесную отрицательную связь между этими показателями (рис. 2). Эта связь у исследованных животных описывается следующими уравнениями регрессии: у верблюдов y = –0.1678 x + 71.126, у сайгаков y = –0.1613 x + + 65.587, у лошадей y = –0.1143 x + 66.141. Из уравнений следует, что при изменении доли злаков в составе рациона всех сравниваемых животных пропорционально и в равной мере меняется переваримость съеденного корма. При равном увеличении доли злаков в питании (например, на 10%) переваримость рациона у жвачных (верблюдов и сайгаков) снижается одинаково (на 1.6–1.7%), у нежвачных (лошадей) меньше (на 1.1%).
Рис. 2.
Связь переваримости рациона с долей злаков в рационе верблюда (а), сайгака (б), лошади Пржевальского (в) на степном пастбище.
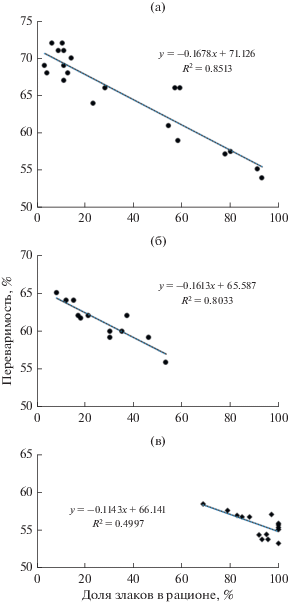
Так или иначе, увеличение злаков в питании животных вызывает сходное или близкое к этому снижение переваримости у всех трех видов. Тем не менее отношение к злакам у них кардинально различается. При пастьбе на одинаковых пастбищах лошади Пржевальского выбирают для питания именно злаки (90–100%), несмотря на их низкую переваримость (<60%) (рис. 2). На графике связи питания с долей злаков в рационе все метки, отражающие набор кормов в рационе, сгущаются в зоне с наивысшей долей злаков (до 100%), что объясняет более глубокое снижение переваримости их рациона. У видов со жвачным типом питания – сайгаков и верблюдов – большая часть кривой, отражающей связь переваримости с количеством злаков, располагается в зоне с низкой долей злаков в питании, что сочетается с более высокими (>60%) значениями переваримости (рис. 2). Характерно, что у верблюдов хорошо заметное сгущение точек располагается в зоне с почти полным исключением злаков (<10%). Это сгущение отражает свойственную верблюдам пастьбу на залежи в сообществах однолетних сорных растений с лебедой татарской и отсутствием злаков.
Таким образом, если сравнить разные виды исследованных копытных по их отношению к потреблению злаков и разнотравья, то явное преимущество в потреблении злаков принадлежит лошади Пржевальского: доля злаков в рационе во все сезоны максимальна (85–100%). При этом преимущество злаков объясняется не только их обилием в составе пастбищной растительности, а прежде всего их предпочтением во все сезоны (ИИ = 1.3–2.2), несмотря на их низкую переваримость (55–57%). Участие злаков в составе рациона верблюдов летом и осенью составляло 11–19%, причем верблюды в это время явно избегали их потребление (ИИ = 0.5–06). Лишь весной молодые растения злаков с успехом поедались верблюдами: участие в питании 77% при их явном предпочтении (ИИ = 1.9) независимо от их пониженной переваримости в это время (56%). Сайгаки избегали потребление злаков во все сезоны. Доля злаков в питании в разные сезоны менялась у них от 18 до 38%, при этом животные явно избегали их выбор (ИИ = 0.3–0.5). Пониженное участие злаков в питании обеспечило сайгакам более высокую переваримость, которая достигала 65%.
Суточное потребление корма. Одновременный учет отложения фекалий и переваримости корма позволил определить суточное потребление корма (табл. 6). Количество потребляемого корма было различным в разные сезоны года, при этом максимальное количество сухой массы корма верблюды и лошади Пржевальского потребляли в летне-осеннее время (соответственно 20–26 и 7–13 кг/особь в сутки) и значительно меньше (соответственно 8 и 5 кг) в зимне-весеннее (апрель), что для лошади Пржевальского было отмечено ранее (Kuntz et al., 2006). В данном случае зимнее снижение интенсивности питания верблюдов и лошадей отражает характерную для растительноядных млекопитающих умеренных широт (северный олень, лось, вапити, сайгак и др.) зимнюю гипофагию. Характерно, что в Аскании-Нова суточное потребление пищи лошадьми Пржевальского на естественном пастбище изменялось в зависимости от качества корма от 7 до 11 кг/особь (сухая масса) (Позднякова и др., 2011). В пересчете на энергетические показатели по нашим данным свободнопасущиеся верблюды потребляли в расчете на обменную массу тела >1.661 МДж/кгW 0.75 обменной энергии в сутки, лошади Пржевальского – практически столько же (1.421) (табл. 6). Известно, что большинство диких жвачных (Capreolus capreolus, Cervus elaphus, Alces alces, Rangifer tarandus, Antilocarpa americana и др.) расходуют на жизнедеятельность в состоянии покоя летом от 0.51 до 0.9 МДж/кгW 0.75 энергии (Hudson, Christopherson, 1985). У верблюдов и лошадей Пржевальского в нашем случае в теплое время года потребляемая обменная энергия достигала 1.4–1.6 МДж/кгW 0.75 в сутки, т.е. существенно превышала отмеченные значения потребной энергии, свойственные большинству копытных.
Таблица 6.
Суточное потребление корма свободнопасущимися верблюдами и лошадьми Пржевальского на естественном пастбище в разные сезоны 2016 г.
| Показатель | Верблюды | Лошади Пржевальского | ||||
|---|---|---|---|---|---|---|
| апрель | июнь–июль | сентябрь–октябрь | апрель | июнь–июль | сентябрь–октябрь | |
| Переваримость рациона, % | 56.2 ± 5.8 | 67.0 ± 5.1 | 69 ± 5.34 | 54.6 ± 2.8 | 57.1 ± 5.7 | 56.7 ± 6.2 |
| Суточное отложение фекалий, кг (сухая масса) | 3.5 ± 0.2 | 6.5 ± 0.6 | 8.0 ± 1.0 | 2.1 ± 0.2 | 3.1 ± 0.2 | 5.6 ± 0.6 |
| Суточное потребление корма (сухая масса) | ||||||
| кг/особь | 7.99 ± 0.9 | 19.7 ± 2.35 | 25.8 ± 3.79 | 4.6 ± 0.50 | 7.2 ± 0.85 | 12.9 ± 2.0 |
| г/кгW 0.75 | 53.2 ± 6.0 | 131.0 ± 15.6 | 171.5 ± 25.2 | 56.8 ± 6.1 | 88.9 ± 10.5 | 159.2 ± 24.7 |
| Суточное потребление энергии, МДж/кгW 0.75 | ||||||
| валовой | 0.91 ± 0.1 | 2.24 ± 0.3 | 2.94 ± 0.4 | 0.97 ± 0.1 | 1.521 ± 0.2 | 2.725 ± 0.4 |
| переваримой | 0.51 ± 0.05 | 0.150 ± 0.2 | 2.03 ± 0.3 | 0.53 ± 0.05 | 0.869 ± 0.1 | 1.545 ± 0.2 |
| обменной | 0.42 ± 0.04 | 1.232 ± 0.16 | 1.66 ± 0.24 | 0.49 ± 0.05 | 0.80 ± 0.09 | 1.421 ± 0.18 |
Таким образом, свободнопасущиеся верблюды и лошади Пржевальского на исследуемом степном пастбище потребляют больше корма, чем им требуется для покрытия текущих расходов энергии, в том числе больше обычных величин питания диких жвачных в естественных природных условиях. Очевидно, превосходство потребления корма верблюдами при свободной пастьбе над известными кормовыми потребностями жвачных – важное свойство верблюдов, позволяющее им накапливать страховочные жировые запасы в горбах (Абатуров и др., 2018). Безусловно, подобное страховочное накопление жировых запасов в естественных условиях обитания – важная адаптация этих животных, обеспечивающая их обитание в суровых природных условиях с крайне неустойчивыми кормовыми запасами (сухие пустыни, суровые зимы) и компенсирующая низкий уровень питания в неблагоприятный период со скудными кормовыми запасами.
Лошади Пржевальского на исследованном пастбище также в полной мере обеспечены полноценным кормом, несмотря на преимущественное питание низкопереваримым злаковым кормом. Безусловно, заднекишечный тип пищеварения позволил лошади Пржевальского компенсировать низкую переваримость этого корма ростом его валового потребления и способствовал специализации лошадей именно на этой группе растительных кормов.
Обращает на себя внимание неустойчивая динамика переваримости кормов, потребляемых на рассматриваемом пастбище сайгаками. Сайгаки остро реагируют на состав растительности на пастбище и связанный с этим состав потребляемого корма. Особенно негативно влияет на их питание обилие в корме злаковых растений. Этим обусловлены относительно невысокие и непостоянные показатели переваримости корма, варьирующие от 58% весной и 61% осенью до 65% летом. Эти значения лишь немного превышают пороги переваримости, необходимые для поддержания жизни (56%), роста (61%) и не достигают уровня, необходимого для лактации (68%) (Abaturov, Subbotin, 2011). Можно полагать, что данное пастбище по обеспеченности полноценным кормом находится в близком к критическому для сайгаков состоянии. Положительное влияние на кормовую обеспеченность сайгаков безусловно оказывают совместно обитающие на этом же пастбище лошади Пржевальского. Специализация лошадей на питании злаками уменьшает обилие злаков на пастбище, способствует тем самым росту разнотравья и оказывает этим положительное влияние на обеспеченность сайгаков полноценным кормом.
ЗАКЛЮЧЕНИЕ
Степная разнотравно-дерновинно-злаковая растительность долины Западного Маныча обладает сложной структурой и богатым разнообразием растительных сообществ. Две группы растений, слагающие кормовую фитомассу степных пастбищ, – злаки и разнотравье – относительно равны по участию в растительном покрове исследуемого пастбища. Обе группы разделяются по важнейшему кормовому качеству – переваримости. Злаки менее переваримы, в связи с чем для всех трех сравниваемых видов копытных закономерный характер приобретает равная по значениям отрицательная связь между обилием злаков в потребляемом корме и его переваримостью. Тем не менее отношение к злакам и разнотравью, к их участию в питании у всех видов коренным образом различается, что выражается в разном предпочтении злаков и разнотравья и отражается на избирательности питания, на разделении животных по кормовой специализации. Возникает адаптивная консортивная связь между каждым видом животного и определенной группой растений, от которой зависит полноценное питание этого вида и его благополучие.
Лошади Пржевальского – облигатные потребители злаковых растений – во время пастьбы целенаправленно выбирают злаки. Несмотря на низкую переваримость последних, лошади Пржевальского в полной мере обеспечены полноценным питанием, поскольку их пищеварительный аппарат способен компенсировать низкую переваримость увеличением валового потребления низкопереваримого злакового корма.
Остро и отрицательно реагируют на обилие злаков сайгаки. В отличие от лошадей они избегают потребление злаков (18–38% в составе рациона). Нестабильность обилия злаков в растительности пастбища служит причиной неустойчивой доли злаков в питании и непостоянства переваримости потребляемого сайгаками корма (58–64%). В случаях периодического увеличения количества злаков на пастбище и в питании животных переваримость уменьшается до критических значений, не обеспечивающих физиологические потребности животных (поддержание жизни, рост, лактация). Это указывает на близкое к критическому состояние современных условий обитания группировки сайгаков на данном пастбище. Положительное влияние на кормовую обеспеченность сайгаков оказывают лошади Пржевальского, питающиеся исключительно злаками и избегающие потребление разнотравья. Тем самым они не только не оказывают конкурентного давления на кормовую обеспеченность сайгаков, но и, наоборот, поддерживают необходимый для сайгаков состав кормовой растительности.
Питание верблюдов в той же степени отрицательно связано с обилием злаков в питании, однако в отличие от других пастбищных животных верблюды с успехом используют в качестве пастбища многочисленные в настоящее время залежные участки, возникшие на месте бывшей пашни. Господствующие на залежах однолетние бурьянистые растения обеспечивают верблюдов высокопитательными (с переваримостью до 68%) видами разнотравья, особенно многочисленной и доминирующей в этих сообществах лебедой татарской.
Разнообразие пастбищных растений по их кормовому качеству требует такого же разнообразия видов животных, способных в полной мере освоить разнородный кормовой ресурс. На примере трех видов пастбищных млекопитающих и двух групп степных пастбищных растений (злаков и разнотравья) хорошо видно, что даже такой небольшой набор взаимодействующих компонентов необходим для сбалансированного функционирования пастбищной экосистемы. Лошади способны ограничиваться питанием грубых кормов, т.е. слабопереваримым злаковым компонентом пастбищной растительности, непригодным для многих растительноядных, в данном случае для сайгаков и в значительной мере для верблюдов. В свою очередь сайгаки приспособлены к питанию преимущественно разнотравьем. Верблюды дополнительно к этому в отличие от большинства растительноядных млекопитающих обладают способностью осваивать растительные ресурсы бурьянистых растительных сообществ, формирующихся на нарушенных по тем или иным причинам почвах.
Особую важность различная специализация (избирательность) питания приобретает в условиях совместной пастьбы видов животных с разными типами пищеварения. С ней связаны важнейшие экологические последствия такой пастьбы как для самих популяций животных, так и для пастбищной экосистемы в целом. С одной стороны, она отражает разделение животных по экологическим нишам и ограничивает конкуренцию между совместно обитающими видами, с другой стороны, выступает важным фактором сохранения видового разнообразия пастбищной растительности. Монодоминантное стадо пастбищных животных вызывает угнетение и даже утрату на пастбищах наиболее поедаемых видов растений, ведет к господству менее поедаемых, низкопитательных и даже ядовитых растений, что резко снижает кормовое качество пастбищ и кардинально меняет видовой состав растительного покрова. В многовидовых сообществах пастбищных животных этот отрицательный эффект исключается за счет различий в выборе разных по качеству кормовых видов, воздействие на состав растений выравнивается, сохраняется и даже улучшается видовое разнообразие пастбищной растительности.
Количество потребляемого корма различно в разные сезоны года, при этом максимальную сухую массу корма верблюды и лошади Пржевальского потребляют в летне-осеннее время и значительно меньше в зимне-весеннее (апрель), что отражает характерную для крупных растительноядных млекопитающих умеренных широт зимнюю гипофагию. У верблюдов и лошадей Пржевальского в нашем случае в теплое время года потребляемая обменная энергия превышала стандартные значения потребной энергии, свойственные большинству диких копытных. Таким образом, свободнопасущиеся верблюды и лошади Пржевальского на исследуемом степном пастбище потребляют больше корма, чем им требуется для удовлетворения текущих энергетических потребностей, свойственных диким жвачным в естественных природных условиях. У сайгаков относительно невысокие и непостоянные показатели переваримости корма лишь незначительно превышают пороговые значения переваримости, обеспечивающие жизнеспособность популяции. Это указывает на то, что данное пастбище в настоящее время находится на близком к критическому для сайгаков состоянию по обеспеченности их полноценным питанием.
Авторы признательны И.Н. Медянникову, С.Н. Олейникову, В.И. Данькову, Е.Ю. Кузнецову, Л.В. Клец, А.Д. Липковичу за организацию и материально-техническую поддержку исследований на землях степного вольерного комплекса Центра редких животных европейских степей “Ассоциации "Живая природа степей” и Ростовского биосферного заповедника.
Работа выполнена при финансовой поддержке РФФИ (грант 18-04-00172).
Список литературы
Абатуров Б.Д. Критические параметры качества растительных кормов для сайгаков (Saiga tatariса) на естественном пастбище в полупустыне // Зоол. журн. 1999. Т. 78. № 8. С. 999–1010.
Абатуров Б.Д. Кормовые ресурсы, обеспеченность пищей и жизнеспособность популяций растительноядных млекопитающих // Зоол. журн. 2005. Т. 84. № 10. С. 1251–1271.
Абатуров Б.Д., Холодова М.В., Субботин А.Е. Интенсивность питания и переваримость кормов у сайгаков // Зоол. журн. 1982. Т. 61. Вып. 12. С. 1870–1881.
Абатуров Б.Д., Колесников М.П., Никонова О.А., Позднякова М.К. Опыт количественной оценки питания свободнопасущихся млекопитающих в естественной среде обитания // Зоол. журн. 2003. Т. 82. № 1. С. 104–114.
Абатуров Б.Д., Ларионов К.О., Колесников М.К., Никонова О.А. Состояние и обеспеченность сайгаков кормом на пастбищах с растительностью разных типов // Зоол. журн. 2005. Т. 84. № 3. С. 377–390.
Абатуров Б.Д., Петрищев Б.И., Колесников М.П., Субботин А.Е. Сезонная динамика кормовых ресурсов и питание сайгака на естественном пастбище в полупустыне // Успехи соврем. биологии. 1998. Т. 118. Вып. 5. С. 564–584.
Абатуров Б.Д., Колесников М.П., Лихнова О.П., Петрищев Б.И., Никонова О.А. Использование лигнина и кремния кормовой растительности в качестве индикаторов переваримости для количественной оценки потребления пищи свободнопасущимися сайгаками // Зоол. журн. 1997. Т. 76. № 1. С. 104–113.
Абатуров Б.Д., Кузнецов Г.В., Магомедов М.-Р.Д., Петелин Д.А., Фекаду Кассайе. Оценка кормовой обеспеченности диких копытных в сухой сезон на луговых пастбищах Эфиопии // Зоол. журн. 1996. Т. 75. Вып.3. С. 439–450.
Абатуров Б.Д., Казьмин В.Д., Джапова Р.Р., Аюшева Е.Ч., Джапова В.В., Нохаева Д.В., Колесников М.П., Миноранский В.А., Кузнецов Ю.Е. Кормовые ресурсы, питание и обеспеченность пищей свободнопасущихся верблюдов (Camelus bactrianus) на пастбищах степной природной зоны // Зоол. журн. 2018. Т. 97. № 3. С. 348–361.
Гинс М.С., Колесников М.П., Гинс В.К., Кононков П.Ф. Методика анализа органической и минеральных (растворимой и полимерной) форм кремния в овощных культурах. М.: РУДН, 2012. 37 с.
Ивлев В.С. Экспериментальная экология питания рыб. М.: Пищепромиздат, 1955. 253 с.
Инструкция для зональных агрохимических лабораторий по анализу кормов и растений. М.: Колос, 1968. 55 с.
Колесников М.П. Формы кремния в растениях // Успехи биол. химии. 2001. Т. 41. С. 301–332.
Колесников М.П., Абатуров Б.Д. Формы кремния в растительном материале и их количественное определение // Успехи соврем. биологии. 1997. Т. 117. Вып. 5. С. 534–548.
Ларин И.В., Агагабян Ш.М., Работнов Т.А., Любская А.Ф., Ларина В.К., Касименко М.А., Говорухин В.С., Зафрен С.Я. Кормовые растения сенокосов и пастбищ СССР / Ред. Ларин И.В. Т. I. М.; Л.: Гос. изд-во с.-х. лит-ры, 1950. 688 с.
Мак-Доналъд П., Эдвардc Р., Гринхалдж Дж. Питание животных. М.: Колос, 1970. 504 с.
Нормы и рационы кормления сельскохозяйственных животных / Ред. Калашников А.П., Фисинин И.В., Щеглов В.В., Клейменов Н.И. М.: Россельхозакадемия, 2003. 456 с.
Позднякова М.К., Жарких Т.Л., Ясинецкая Н.И., Колесников М.П. Количественная оценка питания полувольной группировки лошади Пржевальского (Equus przewalskii) в степном местообитании (заповедник “Аскания-Нова”) // Зоол. журн. 2011. Т. 90. № 3. С. 368–376.
Розенфельд С.Б. Атлас микрофотографий кутикулярной структуры эпидермиса кормовых растений позвоночных фитофагов тундровой и степной зон Евразии. М.: КМК, 2011, CD-диск.
Розенфельд С.Б., Ларионов К.О. Применение кутикулярного копрологического анализа в изучении состава кормов копытных в Калмыкии и Монголии // Актуальные проблемы экологии и эволюции в исследованиях молодых ученых. М.: КМК, 2006. С. 117–141.
Скопин А.Е. Значение компонентов качественного состава кормовых растений диких животных. Киров: ВНИИОЗ им. Б.М. Житкова, 2003. 204 с.
Станков С.С., Талиев В.И. Определитель высших растений европейской части СССР. М.: Современная наука, 1957. 740 с.
Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья, 1995. 995 с.
Шорыгин А.А. Питание, избирательная способность и пищевые взаимоотношения некоторых Gobiidae Каспийского моря // Зоол. журн. 1939. Т. 18. Вып. 1. С. 27–53.
Abaturov B.D., Subbotin A.E. Forage quality thresholds for Saiga antelope in a semideserts rangeland // Rus. J. Theriol. 2011. V. 10. № 2. P. 71–81.
Clauss M., Kaiser T., Hummel J. The morphophysiological adaptations of browsing and grazing mammals // The Ecology of Browsing and Grazing. Ecolugical Studies 195 / Eds Gordon I.J., Prins H.H.T. Berlin: Springer-Verlag, 2008. P. 47–88.
Ely R.E., Kane EA., Jacobson W.C., Moore LA. Studies on the composition of lignin isolated from orchard grass hay cut at stages of maturity and from corresponding feces // J. Dairy Sci. 1953. V. 36. P. 346–355.
Hofmann R.R. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system // Oecologia. 1989. V. 78. P. 443–457.
Hofmann R.R. Die Wiederkauer // Biol. Unserer Zeit. 1991. Bd 21. № 2. S. 73–80.
Holechek J.L. Comparative contribution of Grasses, Forbs, and Shrubs to the nutrition of range ungulates // Rangelands. 1984. V.6. P. 261–263.
Hudson R.J., Christopherson R.J. Maintenance metabolism // Bioenergetics of wild herbivores / Eds Hudson R.Y., White R.G. Florida: CRC Press, Inc. Boca Raton, 1985. P. 121–142.
Jacobs J. Quantitative measurement of food selection // Oecologia. 1974. V. 14. № 2. P. 413–417.
Kuntz R., Kubalek C., Ruf T., Tataruch F., Arnold W. Seasonal adjustment of energy budget in a large wild mammal, the Przewalski horse (Equus ferus przewalskii). I. Energy intake // J. Experim. Biol. 2006. № 209. P. 4557–4565.
Massey F.P., Ennos A.R., Hartley S.E. Grasses and the resource availability hypothesis: the importance of silica-based defenses // J. Ecol. 2007. V. 95. № 3. P. 414–424.
McNaughton S.J., Tarrants J.L., McNaughton M.M., Davis R.H. Silica as a defense against herbivory and a growth promotor in African grasses // Ecology. 1985. V. 66. P. 528–535.
Menard C., Duncan P., Fleurance G., Georges J.-Y., Lila M. Comparative foraging and nutrition of horses and cattle in European wetlands // J. Appl. Ecol. 2002. V. 39. P. 120–133.
Salter R.E., Hudson R.J. Feeding ecology of feral horses in western Alberta // J. Range Manage. 1979. V. 32. № 3. P. 221–225.
Sen Z., Keremu A., Shanhui L., Yongjun Z., Def H. Food patch particularity and forging strategy of reintroduced Przewalski’s horse in North Xinjiang, China // Turkish J. Zool. 2017. V. 41. P. 924–930.
Slivinska K., Kopij G. Diet of the Przewalski’s horse Equus Przewalskii in the Chernobyl exclusion zone // Pol. J. Ecol. 2011. V. 59. № 4. P. 841–847.
Stewart D.R.M. Analysis of plant epidermis in faeces: a technique for studying the food preferences of grazing herbivores // J. Appl. Ecol. 1967. V. 4. P. 83–111.
Streeter C.L. A review of techniques used to estimate the in vivo digestibility of grazed forage // J. Anim. Sci. 1969. V. 29. № 5. P. 757–768.
The Ecology of Browsing and Grazing / Ecological Studies 195 / Eds Gordon I.J., Prins H.H.T. Berlin: Springer-Verlag, 2008. 330 p.
Van Soest P.J., Jones L.H.P. Effect of silica in forages upon digestibility // J. Dairy Sci. 1968. V. 51. № 10. P. 1644–1648.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая