

Известия РАН. Серия биологическая, 2020, № 1, стр. 96-104
Восстановительные сукцессии почвенной микрофауны в загрязненных нефтью экосистемах европейской субарктики
Е. Н. Мелехина *
Институт биологии КомиНЦ УрО РАН
167982 Сыктывкар, ул. Коммунистическая, 28, Россия
* E-mail: melekhina@ib.komisc.ru
Поступила в редакцию 07.11.2018
После доработки 04.12.2018
Принята к публикации 04.12.2018
Аннотация
Определена зависимость восстановления почвенного зооценоза от сукцессии растительного сообщества после рекультивации. Зарегистрировано повышение плотности населения беспозвоночных в ряду сукцессии фитоценоза “без растений”–“низкое проективное покрытие”– “высокое проективное покрытие” (5100 ± 1340, 12 400 ± 2823, 22 040 ± 3008 экз./м2 соответственно на площадке “агрохимические методы”). Выявлены разные тенденции изменения плотности населения микроартропод с течением времени: снижение параметра для личинок двукрылых и мезостигматических клещей и повышение для коллембол и панцирных клещей, что определяло динамику структуры группировок и смену доминирующих таксонов. Демутация зооценоза подразделена на три этапа. Определены таксономические группы микроартропод – биомаркеров этапов сукцессии: для первого этапа это личинки двукрылых и мезостигматические клещи, для второго – коллемболы, для третьего – панцирные клещи.
Нефтяное загрязнение – значимый для Субарктики европейской части России фактор нарушения природных сообществ. Аварийные разливы нефти, которые происходят в зоне деятельности предприятий нефтедобывающей и нефтеперерабатывающей промышленности, оказывают отрицательное воздействие на биоту почвы. Для восстановления нарушенных экосистем применяют различные способы рекультивации (Природоохранные…, 2006). Изучение особенностей демутации загрязненных нефтью почвенных сообществ имеет практическое значение для диагностики их состояния и оценки эффективности рекультивационных мероприятий. Комплексы почвенных беспозвоночных быстро реагируют на нарушения условий окружающей среды изменением численности, перестройкой состава и структуры группировок, что определяет их значение как биоиндикаторов.
В качестве модельных систем для исследования закономерностей восстановления почвенной фауны после нарушений природного характера рассматривали хронологические ряды участков, освободившихся после таяния ледников (Kaufmann et al., 2002). Выявлена определенная последовательность колонизации разными таксонами беспозвоночных послеледниковых субстратов (Skubała, Gulvik, 2005; Hågvar et al., 2009; Hågvar, 2010), шлаковых и пепловых полей, образованных после извержения вулканов (Ryabinin, Pan’kov, 2009). Предложены модели первичных сукцессий беспозвоночных (Vater, Matthews, 2015). Изучались особенности восстановления и формирования новых почвенных зооценозов в антропогенно-измененных почвах (Ryabinin, Pan’kov, 2009; Coulson et al., 2015). Сукцессии микроартропод исследованы на постиндустриальных субстратах (Skubała, Gulvik, 2005; Андриевский, Якутин, 2012), рыхлых отвалах каменных карьеров (Ryabinin, Pan’kov, 2009), в зарастающих пахотных полях (Zaitsev et al., 2006), агроценозах (Лящев, 2004). Выявлено, что на посттехногенных субстратах протекают сукцессии беспозвоночных, сходные с сукцессиями природного генеза.
Влияние нефтяного загрязнения на почвенную фауну исследовано еще неполно. Особенности восстановительных сукцессий беспозвоночных обсуждаются лишь в некоторых работах (Артемьева и др., 1988; Артемьева, 1989; Мелехина, 2007а, б; Михеева, 2008). Совершенно недостаточно изучены закономерности восстановления почвенных сообществ в загрязненных нефтью экосистемах Субарктики. Мало изучено значение рекультивационных мероприятий для полноценного восстановления почвенного зооценоза – одного из основных звеньев процесса гумусообразования.
Цель работы – обобщить результаты долговременного комплексного мониторинга экосистем крайнесеверной тайги, выявить закономерности восстановления почвенной микрофауны после нефтяного загрязнения в условиях Европейской Субарктики.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проведены в условиях Субарктики европейской части России, в Усинском районе Республики Коми. Район исследований расположен в подзонах лесотундры и крайнесеверной тайги (Юдин, 1954; Леса…, 1999). Тип климата умеренно-континентальный. Среднегодовая температура –4°C; средняя температура января –18…–20°C, июля +14°C. Снежный покров удерживается 210 дней в году, с третьей декады октября до начала июня (Атлас…, 1997). Основная часть территории района относится к Печоро-Усинскому округу болотно-подзолистых, глееподзолистых, тундрово-болотных и болотных торфяных почв (Атлас почв…, 2010).
Результаты обсуждаются на примере участка рекультивации № 20, который расположен в зоне деятельности нефтедобывающего предприятия “ЛУКОЙЛ-Усинскнефтегаз”, на территории Возейского нефтяного месторождения (66°37′40′′ с.ш., 57°07′56′′ в.д.). Почвы участка – торфяно-болотные верховые (Fibric Histosols (Dystric), по классификации FAO) (Мировая…, 2018) с водозастойным режимом техногенно-нарушенного ландшафта (Природоохранные…, 2006; Мелехина и др., 2016). Аварийный разлив нефти произошел в 1996 г.; слой торфа был пропитан нефтью на глубину 1.0–1.5 м. В 2002 г. был заложен полевой эксперимент по оценке эффективности различных методов рекультивации в условиях Субарктики.
Анализируются данные по четырем опытным площадкам (“техническая рекультивация”), “агрохимическая обработка”, “биопрепарат Родер”, “биопрепарат Универсал”, а также фоновому сообществу. Содержание нефти в почве опытных площадок перед началом рекультивации составляло от 87 до 465 мг/г (Природоохранные…, 2006). На всех опытных площадках была проведена техническая рекультивация: частичное осушение с помощью дренажных каналов и сбор нефти, фрезерование почвы на глубину 30–40 см. В почву двух площадок были внесены также биопрепараты (“Универсал” и “Родер”) и минеральные удобрения. Биопрепарат “Универсал” разработан в Институте биологии КомиНЦ УрО РАН (Маркарова, 1999, 2004); биопрепарат “Родер” – на химическом факультете МГУ (Мурыгина и др., 2001). На одной площадке была проведена агрохимическая обработка почвы: внесение минеральных удобрений и доломитовой муки одновременно с фрезерованием. На всех площадках биорекультивации применяли минеральное удобрение нитроаммофоску по 350 кг/га (Природоохранные…, 2006). На опытных площадках был проведен посев многолетних трав: на площадке “Универсал” – щучки дернистой Deschampsia caespitosa, на площадке “Родер” – двукисточника тростниковидного Phalaroides arundinacea и тимофеевки луговой Phleum pratense, на площадке “агрохимические методы” – многолетней травосмеси и овса посевного Avena sativa (Природоохранные…, 2006). В качестве контрольной выступала площадка технической рекультивации, на которой посев трав не проводили, биопрепараты и удобрения не вносили. Фоновое растительное сообщество – ивово-ерниковое осоково-хвощевое болото с торфяно-болотной почвой.
Пробы на микрофауну отбирали в августе в 12‑кратной повторности на каждой опытной площадке размером 5 × 5 см (площадью 0.0025 м2), глубиной 10 см каждая (Методы…, 1975). Сбор материала был проведен через 4 года (2006 г.) и 7 лет (2009 г.) после рекультивации. Извлечение беспозвоночных из почвенных проб осуществляли стандартным методом – с помощью термоэклекторов Берлезе–Тульгрена. Выгонку проводили от 7 до 10 дней в зависимости от влажности субстрата. Учитывали всех представителей размерно-функциональной группы микрофауны (животных, размеры которых составляют от 0.1 до 2–3 мм); определяли плотность населения микроартропод (экз./м2) (Методы…, 1975); рассчитывали относительное обилие таксонов (%) (Песенко, 1982). Трофические группы беспозвоночных названы в соответствии с опубликованной ранее классификацией (Стриганова, 1980). Достоверность различий плотности населения и относительного обилия микроартропод определяли с помощью теста Манна–Уитни при уровне значимости p < 0.05 в программе PAST V 3.0 (Hammer et al., 2001).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Связь с сукцессией растительного сообщества. Восстановление комплексов беспозвоночных наблюдали в зависимости от степени развития растительного покрова. На площадках рекультивации происходило формирование фитоценозов из сеяных злаков; общее проективное покрытие травянистого яруса постепенно увеличивалось в ходе восстановительной сукцессии (Мелехина и др., 2016). Были выделены стадии сукцессии фитоценоза “без растений”, “низкое проективное покрытие” и “высокое проективное покрытие”. Наши исследования показали, что восстановление зооценоза начиналось еще в отсутствие растительного покрова. Численность, состав и структура населения почвенных беспозвоночных изменялись с определенной закономерностью по мере развития растительного сообщества. Спустя 4 года после биорекультивации (2006 г.) была выявлена тенденция повышения плотности населения микрофауны в ряду “без растений”–“низкое проективное покрытие”–“высокое проективное покрытие”. Значения параметра в почве с растительным покровом “низкое проективное покрытие” и “высокое проективное покрытие” на площадке “агрохимические методы” статистически отличались от значений в контроле и почве без растений; на площадке “Родер” – от контрольных, а также достоверно различались между собой (p < 0.05) (рис. 1). При этом абсолютное значение данного параметра на стадии “высокое проективное покрытие” было статистически выше (p < 0.05) на площадке с биопрепаратом “Родер” по сравнению с другими опытными площадками.
Рис. 1.
Плотность населения микрофауны в ряду сукцессии фитоценоза (К – контроль, БР – без растений, НПП – низкое проективное покрытие, ВПП – высокое проективное покрытие; для рис. 1 и 2) на разных опытных площадках: а – “агрохимическая обработка”, б – “Универсал”, в – “Родер”, 2006 г. (±стандартная ошибка, n = 12; a, b, c, d – достоверность различий плотности населения, тест Манна–Уитни при p < 0.05).

На ранней стадии сукцессии фитоценоза (“без растений”) группировки микроартропод были представлены небольшим числом таксонов, по обилию преобладали двукрылые (на личиночной стадии жизненного цикла; в некоторых выборках они были единственным компонентом животного населения), а также мезостигматические клещи. На следующей стадии сукцессии (“низкое проективное покрытие”) разнообразие микроартропод повышалось: увеличивалось число таксономических групп и групп-доминантов; доминирующими были клещи (мезостигматические, акаридиевые, простигматические) и личинки двукрылых. На более поздней стадии сукцессии растительного сообщества (“высокое проективное покрытие”) появлялись коллемболы и панцирные клещи. На площадке с биопрепаратом “Родер” коллемболы дополняли группу доминантов.
Таким образом, по мере развития фитоценоза происходило усложнение группировок микрофауны: увеличивалось число таксонов, повышалась плотность населения животных. Ранее (Артемьева, 1989) было установлено, что восстановление комплекса почвенных беспозвоночных при нефтяном загрязнении коррелирует с направлением техногенной сукцессии растительности наряду со скоростью распада нефти в почве, что вполне согласуется с нашими данными. Связь сукцессий почвенной фауны с сукцессиями растительного сообщества была установлена при постпирогенном восстановлении (Kudryasheva, Laskova, 2002), зарастании рыхлых отвалов каменных карьеров (Ryabinin, Pan’kov, 2009), зарастании пахотных угодий (Zaitsev et al., 2006). Отмечалось, что в антропогенно-измененных экосистемах высоких широт распределение микроартропод по местообитаниям зависит от состава и структуры растительного сообщества (Coulson et al., 2003, 2015).
Динамика плотности населения. Для разных таксономических групп микроартропод были выявлены разные тенденции изменения плотности населения с течением времени. Через 7 лет после рекультивации плотность населения личинок двукрылых значительно снизилась на всех опытных площадках. На площадке “Универсал” различия параметра были достоверны для стадии сукцессии фитоценоза “низкое проективное покрытие” (рис. 2). Плотность населения личинок двукрылых статистически снизилась и на контрольной площадке.
Рис. 2.
Динамика плотности населения таксономических групп микрофауны на площадке рекультивации “Универсал”. а – личинки двукрылых, б – мезостигматические клещи, в – коллемболы (±стандартная ошибка, n = 12; a и b – достоверность различий плотности населения при попарном сравнении значений для каждой стадии сукцессии фитоценоза в 2006 (а–в слева) и 2009 гг. (а–в справа), тест Манна–Уитни при p < 0.05).
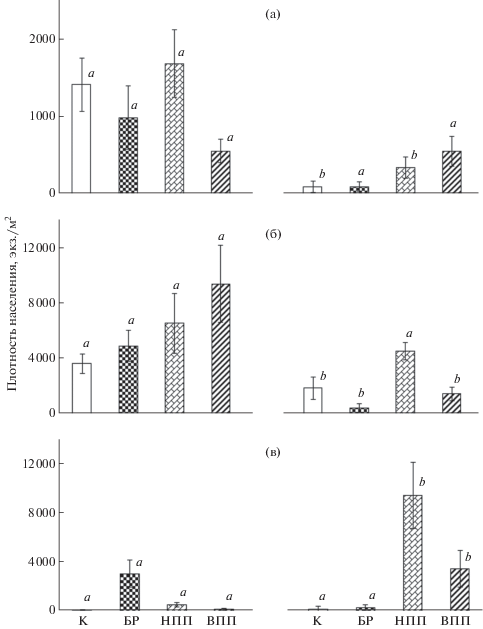
Данный параметр для мезостигматических клещей также заметно снижался; его значения статистически отличались на стадиях “без растений” и “высокое проективное покрытие” опытной площадки в разные годы, а также на контрольной площадке. Через 7 лет после рекультивации было зарегистрировано повышение плотности населения коллембол на стадиях сукцессии фитоценоза “высокое проективное покрытие” и “низкое проективное покрытие” (p < 0.05), в то время как в почве без растений и в контроле этот параметр оставался весьма низким. Таким образом, было зарегистрировано снижение плотности населения личинок двукрылых и мезостигматических клещей с течением времени и, напротив, повышение этого параметра для коллембол.
Восстановительная сукцессия. Сильное нефтяное загрязнение на участке наблюдения можно диагностировать как острое нарушение. Слой торфа был пропитан нефтью до глубины 1–1.5 м; рекультивация была проведена через 6 лет после загрязнения. Кроме того, при уборке нефти и фрезеровании происходило механическое нарушение верхнего (гумусового) горизонта почвы. Были уничтожены как местообитания беспозвоночных, так и сами животные. Восстановление зооценоза, по-видимому, шло по типу первичной сукцессии.
Сукцессия была подразделена на три этапа. Для первого этапа характерно низкое разнообразие группировок микрофауны, которые состояли из небольшого числа таксономических групп и были представлены главным образом или исключительно личинками двукрылых и мезостигматическими клещами при небольшой плотности населения. Первому этапу демутации соответствовала площадка технической рекультивации через 4 года после начала опыта. Здесь доминировали мезостигматические клещи (>65%), вторыми по обилию были личинки двукрылых (~27%, тест Манна–Уитни, p < 0.05) (рис. 3).
Рис. 3.
Динамика относительного обилия (%) таксонов микрофауны в ходе восстановительной сукцессии (I этап: техническая рекультивация, 2006 г.; II этап: площадка “Родер”, 2009 г.; III этап: техническая рекультивация, 2009 г.; фоновый участок: ивово-ерниковое осоково-хвощевое болото).
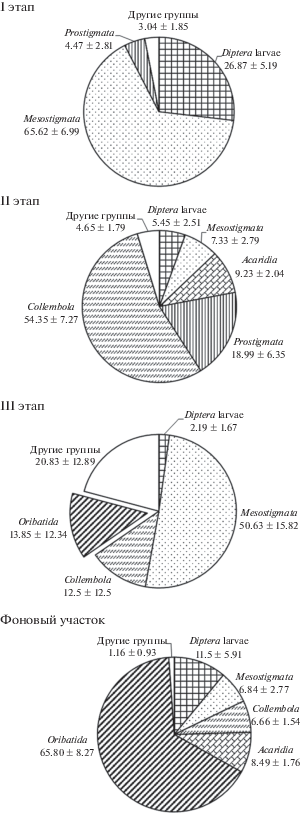
Второй этап связан с появлением коллембол; на ранних стадиях этого этапа доля коллембол в структуре населения незначительна, на более поздних стадиях они становятся доминирующей группой микроартропод. Доминирование по обилию коллембол (54%, тест Манна–Уитни, p < 0.05) наблюдалось на площадке “Родер” через 7 лет после рекультивации (рис. 3). Их наиболее высокое относительное обилие отмечалось на поздней стадии сукцессии фитоценоза (“высокое проективное покрытие”).
На третий этап демутации указывало появление и постепенное повышение обилия панцирных клещей (орибатид). Так, на площадке технической рекультивации через 7 лет доля панцирных клещей составляла ~14% (рис. 3).
Таким образом, в ходе сукцессии менялся вклад разных таксонов в сложение группировок микрофауны. Наиболее высокое относительное обилие личинок двукрылых и мезостигматических клещей наблюдалось на первом этапе восстановления. На более поздних этапах сукцессии двукрылые уходят из числа доминантов вследствие перераспределения обилий между другими таксонами. Уже на втором этапе доля двукрылых в структуре группировок была статистически ниже таковой на первом этапе (p < 0.05). Эта закономерность сохранялась и на третьем этапе. Доля мезостигматических клещей снижалась на втором этапе сукцессии (p < 0.05). В фоновом сообществе они входили в число субдоминантов.
Коллембол на первом этапе демутации не было совсем. На втором этапе они доминировали по обилию. На третьем этапе восстановления их доля в структуре населения статистически снижалась (p < 0.05). В фоновом сообществе коллемболы входили в группу субдоминантов наряду с акаридиевыми и мезостигматическими клещами.
Панцирные клещи отсутствовали на первом и втором этапах сукцессии. Они были отмечены только на третьем этапе. Спустя 4 года после рекультивации орибатиды были единичны, через 7 лет достаточно стабильно присутствовали в почве опытных площадок при невысоком обилии. Их участие в структуре населения начинало приближать площадки рекультивации к фоновому растительному сообществу, где наблюдалось доминирование по обилию панцирных клещей (>65%, тест Манна–Уитни, p < 0.05) и достаточно равномерное распределение обилий между другими группами микроартропод.
В ходе сукцессии увеличивалось разнообразие группировок, в их составе появлялись новые таксономические группы. На площадке “Родер” через 7 лет после рекультивации (второй этап сукцессии) был заметен вклад простигмататических и акаридиевых клещей (18.99 и 9.23% соответственно). Доля Prostigmata статистически повышалась на втором этапе (p < 0.05). Для Acaridia не было отмечено статистических различий обилия между вторым этапом сукцессии и фоновым сообществом, в котором они выступали в роли субдоминантов (8.49%). На третьем этапе восстановления в структуре группировок были представлены личинки жуков и перепончатокрылых при невысоком обилии. В фоновом сообществе разнообразие группировок было выше, чем на опытных площадках за счет присутствия имаго жуков, имаго перепончатокрылых, энхитреид. Ранее были отмечены сходные тенденции динамики крупных беспозвоночных после нефтяного загрязнения: личинки жуков размерной группы мезофауны отмечались на более поздних стадиях восстановления (Мелехина, 2007б).
Таким образом, в ходе сукцессии менялось соотношение по обилию таксонов микрофауны, вследствие чего наблюдались изменения в трофической структуре группировок. На втором этапе более чем в 2 раза увеличивалась доля сапрофагов по сравнению с таковой на первом этапе, соответственно уменьшалась доля хищников (мезостигматических и простигматических клещей). Происходила смена сапрофагов-доминантов: на ранних стадиях восстановления наибольшим обилием отличались личинки двукрылых, затем ведущую роль играли коллемболы, на более поздних стадиях сукцессии повышалось обилие панцирных клещей. Сходную картину наблюдали при восстановлении загрязненной нефтью торфяно-глеевой почвы (Мелехина, 2007а). Доминирование по обилию трофической группы сапрофагов, в основном панцирных клещей, характерно для группировок микрофауны в ненарушенных сообществах крайнесеверной тайги (Мелехина, 2009).
Стадийность процесса восстановления почвенной фауны после нефтяного загрязнения была установлена для черноземных почв лесостепи и дерново-подзолистых почв подзоны южной тайги (Артемьева, 1989). Пионерными группами были микроартроподы: коллемболы и клещи (гамазовые, акароидные и тарсонемоидные). Нами была определена важная роль личинок двукрылых на ранних стадиях восстановления биоценозов Субарктики. Ранее (Артемьева, 1989) отмечалось, что быстро увеличивалась численность хищных (гамазовых) клещей, в то время как формирование комплекса типичных сапрофагов – панцирных клещей – происходило очень медленно. Наши результаты согласуются с этими данными: панцирные клещи заселяли торфяную почву после сильного нефтяного загрязнения гораздо позже, чем другие группы животных, и постепенно увеличивали разнообразие и численность. Низкие темпы восстановления сообществ панцирных клещей были показаны для рыхлых отвалов каменных карьеров (Ryabinin, Pan’kov, 2009), песчаных карьеров (Андриевский, Якутин, 2012), почв, выведенных из сельскохозяйственного пользования (Zaitsev et al., 2006). Отмечалось, что процесс демутации протекает в течение нескольких десятилетий. В условиях Арктики (пос. Пирамида, арх. Шпицберген) в черноземной почве, привезенной с материка для обустройства поселка, через 40 лет наблюдалось крайне низкое разнообразие орибатид, в то время как разнообразие коллембол и мезостигматических клещей было сопоставимо с таковым в естественных сообществах (Coulson et al., 2015). Авторами был сделан вывод о более медленной колонизации панцирными клещами импортированной почвы по сравнению с другими группами беспозвоночных.
Функциональные связи. Очевидно, что последовательность заселения рекультивированной почвы разными группами микроартропод зависит от особенностей их пищевых режимов и смены пищевых субстратов в ходе восстановления почвенного сообщества. Как известно, беспозвоночные трофически связаны с почвенной микробиотой (Стриганова, 1980; Бызов, 2005; Kurakov et al., 2006). Личинки двукрылых, коллемболы, акаридиевые и панцирные клещи относятся к трофической группе сапрофагов; в их рационе важное место занимает сапротрофная микрофлора (бактерии, водоросли, грибы) наряду с органическими остатками растительного и животного происхождения (Гиляров, Стриганова, 1978; Стриганова, 1980). Показано, что водоросли (зеленые и сине-зеленые) наряду со мхами первыми поселяются на почвах после нефтяного загрязнения (Штина, Некрасова, 1988). Вероятно, они и составляют основу рациона пионерных групп беспозвоночных. Для панцирных клещей характерна микофагия; гифы и споры грибов – обязательный компонент их рациона при достаточно широком спектре потребляемых пищевых субстратов (Стриганова, 1980). В сукцессии микрофлоры после нефтяного загрязнения “грибная” стадия следует за “бактериально-грибной” (Маркарова, 1999). Можно предположить, что возрастание обилия панцирных клещей на более позднем этапе сукцессии связано с повышением разнообразия грибных сообществ.
Активное развитие фитоценозов на площадках биорекультивации способствовало формированию структуры почвы и ее обогащению растительными остатками, что создавало благоприятные условия для повышения плотности населения беспозвоночных-деструкторов растительного опада. В наших наблюдениях с этапом развития фитоценоза из сеяных злаков был связан всплеск численности коллембол. Кроме того, злаки-рекультиванты принимали участие в деструкции нефтепродуктов, тем самым ускоряя процессы биологического очищения почвы (Мелехина и др., 2016).
Известно, что микроорганизмы вносят существенный вклад в процессы утилизации нефти и нефтепродуктов в загрязненной почве (Киреева, 1994; Маркарова, 1999, 2004). Через 4 года после рекультивации численность трофических групп микроорганизмов, участвующих в деструкции нефтепродуктов, была выше на площадках с применением биопрепаратов, чем на площадках технической и агрохимической обработки почвы. Соответственно на площадках “Родер” и “Универсал” разрушение нефти и нефтепродуктов протекало более интенсивно (Melekhina et al., 2015).
Восстановление зооценоза на опытных площадках с разными методами биорекультивации шло разными темпами. Так, уже через 4 года после обработки почвы наиболее высокое разнообразие беспозвоночных отмечалось на площадках с биопрепаратами. Через 7 лет на площадках “Родерˮ и “Универсалˮ зооценоз соответствовал более позднему этапу сукцессии, чем на других опытных площадках. Сделан вывод, что методы рекультивации с применением биопрепаратов оказались более эффективными для восстановления зооценоза по сравнению с агрохимическими методами и механической уборкой нефти (Melekhina et al., 2015; Мелехина и др., 2016).
Можно заключить, что прогресс микробоценоза и фитоценоза в ходе сукцессии, с одной стороны, приводит к снижению содержания нефти в почве, с другой – к повышению разнообразия пищевых субстратов микроартропод. Все это создает условия для прогрессирования сукцессии зооценоза. Подготовка субстрата для заселения беспозвоночными и растениями деятельностью микроорганизмов показана для загрязненных нефтью черноземных и дерново-подзолистых почв (Артемьева, 1989), для шлаковых и пепловых полей, образованных после извержения вулканов (Ryabinin, Pan’kov, 2009).
Экологические стратегии. Очевидно, что колонизация рекультивированной почвы пионерными микроартроподами определяется наряду с другими причинами разными типами их экологических стратегий. Двукрылые, акаридиевые и мезостигматические клещи, а также коллемболы – это животные-оппортунисты, r-стратеги. Широкие возможности расселения и высокий потенциал размножения этих групп животных позволяют им первыми занимать рекультивированную почву и увеличивать свою численность на ранних этапах сукцессии. Панцирные клещи – К-стратеги. Их жизненный цикл относительно продолжительный (от нескольких месяцев до года, в условиях холодного климата – до 2–3 лет), а плодовитость низкая (Криволуцкий и др., 1995). Орибатиды малоподвижны, они медленно перемещаются в пространстве, не имеют специальных приспособлений для расселения в новые местообитания. После рекультивации панцирные клещи появлялись в составе группировок микрофауны позже других животных, что определило их значение как индикаторов поздних стадий сукцессии зооценоза.
Для некоторых видов пионерных микроартропод – мезостигматических клещей, коллембол и панцирных клещей – известна способность к партеногенезу, что способствует успешной колонизации новых субстратов (Гиляров, 1982; Рябинин, Паньков, 1987).
ЗАКЛЮЧЕНИЕ
Показана зависимость восстановления почвенного зооценоза от сукцессии растительного сообщества. По мере развития фитоценоза наблюдалось последовательное включение в состав группировок микрофауны новых таксономических групп, повышение плотности населения животных.
Выявлены разные тенденции изменения плотности населения ведущих таксономических групп с течением времени. Зарегистрировано снижение этого параметра для личинок двукрылых и мезостигматических клещей и его повышение для коллембол и панцирных клещей. Характер изменения плотности населения микроартропод определил закономерности динамики структуры группировок и смену доминирующих таксонов.
На основании выявленной последовательности восстановления предложена схема демутационной сукцессии почвенной микрофауны, в которой выделены три этапа. Разные таксономические группы микроартропод представлены в качестве биомаркеров этапов сукцессии; мезостигматические клещи и личинки двукрылых (первый этап), коллемболы (второй), панцирные клещи (третий).
Полученные результаты могут быть рекомендованы для практического применения в биодиагностике состояния почв с нефтяным загрязнением. Определение соответствия группировок микрофауны тому или иному этапу сукцессии позволит судить об успешности восстановления зооценоза при разных методах биорекультивации. Применение характеристик зооценоза на разных этапах восстановления в комплексе с данными о состоянии других компонентов экосистем, таких как фитоценоз и микробоценоз, представляется перспективным для оценки эффективности рекультивационных мероприятий в определенных климатогеографических условиях.
Работа выполнена в рамках Проекта РФФИ № 18-29-05028 мк “Биологические способы ликвидации аварийных разливов нефти в условиях европейской Субарктики: роль микробиологических препаратов, биорекультивация, оценка эффективности, динамика экосистемˮ.
Список литературы
Андриевский В.С., Якутин М.В. Сравнительная характеристика сукцессий панцирных клещей (Oribatei) и почвенной микрофлоры на песчаных карьерах в подзоне северной тайги Западной Сибири // Евразиат. энтомол. журн. 2012. № 11 (1). С. 13–18.
Артемьева Т.И. Комплексы почвенных животных и вопросы рекультивации техногенных территорий. М.: Наука, 1989. 111 с.
Артемьева Т.И., Жеребцов А.К., Борисович Т.М. Влияние загрязнения почвы нефтью и нефтепромысловыми сточными водами на комплекс почвенных животных // Восстановление нефтезагрязненных почвенных экосистем: Сб. науч. трудов. М.: Наука, 1988. С. 82–98.
Атлас почв Республики Коми / Под ред. Добровольского Г.В., Таскаева А.И., Забоевой И.В. Сыктывкар, 2010. 356 с.
Атлас Республики Коми по климату и гидрологии / Под ред. Таскаева А.И. М.: Дрофа; ДиК, 1997. 116 с.
Бызов Б.А. Зоомикробные взаимодействия в почве. М.: ГЕОС, 2005. 213 с.
Гиляров М.С. Экологическое значение партеногенеза // Успехи соврем. биологии. 1982. Т. 93. Вып. 1. С. 10–22.
Гиляров М.С., Стриганова Б.Р. Роль почвенных беспозвоночных в разложении растительных остатков и круговороте веществ // Итоги науки и техники ВИНИТИ. Сер. Зоология беспозвоночных. 1978. Т. 5. С. 8–69.
Киреева Н.А. Микробиологические процессы в нефтезагрязненных почвах. Уфа: Изд-во Башк. гос. ун-та, 1994. 172 с.
Криволуцкий Д.А., Лебрен Ф., Кунст М., Акимов И.А., Баяртогтох Б., Василиу Н., Голосова Л.Д., Гришина Л.Г., Карппинен Э., Крамной В.Я., Ласкова Л.М., Лакстэн М., Маршалл В., Матвеенко А.А., Нетужилин И.А., Нортон Р., Ситникова Л.Г., Смрж Я., Стари Й., Тарба З.М., Шалдыбина Е.С., Эйтминавичюте И.С. Панцирные клещи: Морфология, развитие, филогения, экология, методы исследования, характеристика модельного вида Nothrus palustris C.L. Koch, 1839. М.: Наука, 1995. 224 с.
Леса Республики Коми / Под ред. Козубова Г.М., Таскаева А.И. / Козубов Г.М., Таскаев А.И. Дегтева С.В., Мартыненко В.А., Забоева И.В., Бобкова К.С., Галенко Э.П. М.: Издательско-продюсерский центр “Дизайн. Информация. Картография”, 1999. 332 с.
Лящев А.А. Почвенная биота и плодородие почвы в условиях юга Западной Сибири. Тюмень: Изд-во Тюм. гос. с.-х. акад., 2004. 252 с.
Маркарова М.Ю. Использование углеводородокисляющих бактерий для восстановления нефтезагрязненных земель в условиях Крайнего Севера: Автореф. дис. канд. биол. наук. Сыктывкар: Изд-во Ин-та биологии КомиНЦ УрО РАН, 1999. 22 с.
Маркарова М.Ю. Опыт применения биопрепарата “Универсалˮ для рекультивации нефтезагрязненных земель // Вестн. Ин-та биологии КомиНЦ УрО РАН. 2004. № 10(84). С. 21–23.
Мелехина Е.Н. Влияние нефтяных загрязнений на почвенную микрофауну тундровых сообществ крайнесеверной тайги // Экология человека. 2007а. № 1. С. 16–23.
Мелехина Е.Н. Почвенная мезофауна под влиянием нефтяных загрязнений в условиях крайнесеверной тайги европейского Северо-Востока России // Экология человека. 2007б. № 8. С. 3–9.
Мелехина Е.Н. Трофическая структура населения микроартропод в диагностике состояния загрязненных нефтью почв // Питание и пищевые связи в сообществах животных на европейском Севере: Тр. КомиНЦ УрО РАН, 2009. № 185. С. 39–50.
Мелехина Е.Н., Маркарова М.Ю., Анчугова Е.М., Щемелинина Т.Н., Канев В.А. Определение эффективности методов рекультивации загрязненных нефтью почв // Изв. КомиНЦ УрО РАН. 2016. Вып. 3(27). С. 61–70.
Методы почвенно-зоологических исследований. М.: Наука, 1975. 275 с.
Мировая реферативная база почвенных ресурсов 2014. Международная система почвенной классификации для диагностики почв и создания легенд почвенных карт. Исправленная и дополненная версия 2015. Рим: Изд-во ФАО и МГУ им. М.В. Ломоносова, 2018. 216 с.
Михеева В.Л. Сукцессии панцирных клещей (Oribatei) в почвах верховых болот Нижневартовского района // Вестн. Томск. гос. ун-та. 2008. № 317. С. 246–249.
Мурыгина В.П., Войшвилло Н.Е., Калюжный С.В. Биопрепарат “Родерˮ для очистки почв, почвогрунтов, пресных и минерализованных вод от нефти и нефтепродуктов. Патент RU 2174496. Заявка: 99110890/13. Опубл. 2001.10.10.
Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М., 1982. 287 с.
Природоохранные работы на предприятиях нефтегазового комплекса. Ч. 1. Рекультивация загрязненных нефтью земель в Усинском районе Республики Коми / Ред. Черешнев В.А., Таскаев А.И. / Маганов Р.В., Маркарова М.Ю., Муляк В.В., Загвоздкин В.К., Заикин И.А. Сыктывкар: Изд-во Ин-та биологии КомиНЦ УрО РАН, 2006. 208 с.
Рябинин Н.А., Паньков А.Н. Роль партеногенеза в биологии панцирных клещей // Экология. 1987. № 4. С. 62–64.
Стриганова Б.Р. Питание почвенных сапрофагов. М.: Наука, 1980. 294 с.
Штина Э.А., Некрасова К.А. Водоросли загрязненных нефтью почв // Восстановление нефтезагрязненных почвенных экосистем. М.: Наука, 1988. С. 57–81.
Юдин Ю.П. Геоботаническое районирование // Производительные силы Коми АССР. М., Л.: Изд-во АН СССР, 1954. Т. III. Ч. 1. С. 323–369.
Hågvar S. Primary succession of springtails (Collembola) in a Norwegian Glacier Foreland // Arct. Antarct. Alp. Res. 2010. V. 42. № 4. P. 422–429.
Hågvar S., Solhøy T., Mong C.E. Primary succession of soil mites (Acari) in a Norwegian Glacier Foreland, with emphasis on oribatid species // Arct. Antarct. Alp. Res. 2009. V. 41. № 2. P. 219–227.
Hammer O., Harper D.A.T., Ryan P.D. PAST: Paleontological Statistics software package for education and data analysis // Palaeontol. Electron. 2001. № 4 (1). 9 p.
Coulson S.J., Hodkinson I.D., Webb N.R. Microscale distribution patterns in high Arctic soil microarthropod communities: the influence of plant species within the vegetation mosaic // Ecography. 2003. V. 26. P. 801–809.
Coulson S.J., Fjellberg A., Melekhina E.N., Taskaeva A.A., Lebedeva N.V., Belkina O.A., Seniczak S., Seniczak A., Gwiazdowicz D.J. Microarthropod communities of industrially disturbed or imported soils in the High Arctic; the abandoned coal mining town of Pyramiden, Svalbard // Biodivers. Conserv. 2015. V. 24. № 7. P. 1671–1690. https://doi.org/10.1007/s10531-015-0885-9
Kaufmann R., Fuchs M., Gosterxeier N. The soil fauna of an alpine glacier foreland colonization and succession // Arct. Antarct. Alp. Res. 2002. V. 34. № 3. P. 242–250.
Kudriasheva I.V., Laskova L.M. Oribatid mites (Acariformes, Oribatei) as indicators of postpyrogenous changes in podzol and peat soils of boreal forests // Biol. Bull. 2002. V. 29. № 1. P. 92–99.
Kurakov A.V., Davydova M.A., Byzov B.A. Microarthropods as regulators of the communities of microscopic fungi and biological activity in the litter of a mixed forest // Euras. Soil Sci. 2006. V. 39. № 8. P. 838–847.
Melekhina E.N. Markarova M.Yu., Shchemelinina T.N., Anchugova E.M., Kanev V.A. Secondary successions of biota in oil-polluted peat soil upon different biological remediation methods // Euras. Soil Sci. 2015. V. 48. № 6. P. 643–653.
Ryabinin N.A., Pan’kov A.N. Successions of Oribatid Mites (Acariformes: Oribatida) on Disturbed areas // Biol. Bull. 2009. V. 36. № 5. P. 510–515.
Skubała P., Gulvik M. Pioneer oribatid mite communities (Acari, Oribatida) in newly exposed natural (glacier foreland) and anthropogenic (post-industrial dump) habitats // Polish J. Ecol. 2005. V. 53. P. 105–111.
Vater A.E., Matthews J.A. Succession of pitfall-trapped insects and arachnids on eight Norwegian glacier forelands along an altitudinal gradient: Patterns and models // The Holocene. 2015. V. 25. № 1. P. 108–129.
Zaitsev A.S., Wolters V., Waldhardt R., Daube J. Long-term succession of oribatid mites after conversion of croplands to grasslands // Appl. Soil Ecol. 2006. https://doi.org/10.1016/j.apsoil.2006.01.005
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая