

Известия РАН. Серия биологическая, 2020, № 2, стр. 224-232
Непреднамеренно поставленный эксперимент – заселение вновь созданного городского парка видом-синурбистом обыкновенным хомяком Cricetus cricetus L., 1758
Н. Ю. Феоктистова 1, *, И. Г. Мещерский 1, П. Л. Богомолов 1, С. И. Мещерский 1, Е. А. Кацман 1, Л. А. Пельгунова 1, Е. В. Поташникова 1, А. В. Суров 1
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
* E-mail: feoktistovanyu@gmail.com
Поступила в редакцию 26.08.2019
После доработки 25.10.2019
Принята к публикации 25.10.2019
Аннотация
Описаны условия обитания и генетическая структура популяции обыкновенного хомяка на начальной стадии формирования городской среды. Отмечено, что за довольно непродолжительный срок (не более 12 лет) в искусственно созданном Президентском парке в столице Казахстана г. Нур-Султане благодаря комплексу условий (плодоносящие деревья и кустарники, мощный почвенный слой, искусственный рельеф) сформировалась многочисленная популяция этого вида. Установлено, что все исследованные особи принадлежат к единственной митохондриальной линии, что можно рассматривать как эффект основателя.
В первом десятилетии XXI в. население Земли стало преимущественно городским – к 2018 г. уже 55% населения Земли проживало в городах (4.22 млрд человек), а к 2050 г. в городах будет проживать 68% населения мира (на 2.5 млрд больше, чем в 2018 г.) (World…, 2019). Но высокая концентрация населения и возможность трудиться на территории городов – это лишь внешнее проявление процесса урбанизации. Прямое и косвенное социоэкологическое влияние городской среды распространяется далеко за границы городов. Некоторые из самых глобальных экологических проблем – результат урбанизации (например, нарушение биогеохимических циклов, климата, утрата биоразнообразия, биологические инвазии (von der Lippe et al., 2005)). Более того, стремительное распространение городских экосистем на нашей планете можно рассматривать как качественно новый этап развития жизни на Земле (Schilthuizen, 2018). Из этого следует несомненная значимость городов в формировании условий жизни не только для большей части человечества, но и для всех живых существ на планете. В начале 1990-х гг. возникла самостоятельная дисциплина – городская экология (Collins et al., 2000), которая в силу все возрастающих темпов экономической и культурной глобализации приобрела статус науки (O’Brien, 2012). Сегодня экологи решают фундаментальные и прикладные задачи, связанные с проектированием городов, их устойчивым развитием, повышением качества окружающей среды и благосостояния людей в городах.
Городская среда отличается особым микроклиматом, структурой почв, она настолько своеобразна, что приобретает азональные черты. В городах, расположенных в зоне умеренного климата, наблюдаются признаки опустынивания. Это связано с тем, что среднегодовая и среднесуточная температуры воздуха городских агломераций, как правило, выше, а влажность ниже, чем на окружающих территориях, из-за загазованности и запыленности воздуха, многоэтажной застройки и стока воды. В городах, расположенных в засушливых регионах, напротив, прослеживается тенденция к мезофилизации в результате высаживания деревьев и кустарников, их регулярного полива, создания искусственных водоемов и водотоков, фонтанов и т.д. Иначе говоря, города, несмотря на разную инфраструктуру, географическое положение, культурные традиции, в экологическом плане в какой-то степени сближаются; соответственно, в них могут прослеживаться общие биологические закономерности, касающиеся, в частности, формирования и функционирования биоты.
Радикальное воздействие урбанизации на природно-климатические условия отражается на растительности и животном населении территорий, на которых создаются и развиваются города. Одни виды, наиболее уязвимые, исчезают, не выдерживая антропогенного пресса, другие существуют на грани вымирания. Для некоторых видов измененная в процессе урбанизации среда оказывается не только приемлемой, но даже более благоприятной по сравнению с природной. Яркий пример – сокол сапсан Falco peregrinus, который за последние 30 лет освоил города Северной Америки и Европы (Германии, Польши, России) (Сорокин, 2002). Фактически города стали местом сохранения этого краснокнижного вида при его быстром исчезновении из естественных биотопов. Еще один пример – ястреб Купера Accipiter cooperii, обитающий, в частности, в г. Тусоне (США) (Morinha et al., 2016), где отмечена самая высокая плотность данного вида в Северной Америке (Boggie, Mannan, 2014). Обилие добычи (городские голуби, в частности) и подходящие условия для жизни и гнездостроения (крыши и карнизы небоскребов вместо природных скал) делают города благоприятными для обитания этих хищных птиц.
Подобные случаи сходства между городскими и природными ландшафтами привлекают в крупные города все больше видов дикой флоры и фауны, в том числе экзотические инвазивные виды. Кроме того, формирование городской среды, благоприятной для жизни человека, предполагает включение в ее состав природных или квазиприродных элементов ландшафта (зеленые зоны, парки, скверы, городские сады), что создает подходящие условия и для обитания видов-синурбистов. Урбоценозы лучше осваивают эврибионты и эврифаги с высокой плодовитостью и обширными ареалами (Карасева и др., 1999; Суров и др., 2015). Их лучшему выживанию в городских условиях способствуют дополнительные убежища, кормовые ресурсы (пищевые отходы, плоды и семена декоративных растений), отсутствие естественных хищников и т.д. (Čanady 2013; Feoktistova et al., 2013).
Хотя долгое время городская среда считалась недостойной научных исследований в силу своей “искусственности”, сейчас ученые рассматривают города как естественные лаборатории, где можно и нужно решать не только прикладные сугубо градостроительные, но и фундаментальные экологические задачи. Это, например, влияние фрагментации местообитаний (что типично для городской среды) на генетическую структуру видов, ночного освещения на циркадные ритмы людей и животных, зеленых насаждений на здоровье человека, функционирование новых “сугубо городских” сообществ животных и растений, не имеющих естественных аналогов, и многие другие (Dornelas et al., 2014). Более того, урбанизацию можно назвать одним из самых глобальных и непреднамеренных экспериментов, который поставило (и продолжает ставить) человечество и который позволяет предсказать, как виды будут реагировать на изменения антропогенного характера в масштабах не только городов, но и всей планеты (включая изменения климата) (Lahr et al., 2018).
Однако построение любых моделей возможно только на основе изучения конкретных реальных ситуаций, возникших в рамках локальных экспериментов. Такой непреднамеренный, но совершенно уникальный экологический эксперимент был поставлен в столице Казахстана – Астане (теперь г. Нур-Султан). В 1998 г. столица Казахстана была переведена из Алма-Аты (Алматы) в Акмолу (предыдущее название Астаны), население которой тогда составляло 0.3 млн человек. С этого момента началось интенсивное строительство, население города увеличилось почти в 4 раза (до 1.1 млн человек), значительно увеличилась территория города, в первую очередь в южном направлении.
Город расположен в регионе с резко-континентальным климатом, с теплым засушливым летом и с холодной снежной зимой. Среднегодовая температура +3.5°C, среднегодовое количество осадков 318 мм. Наиболее теплый месяц – июль, его средняя температура +20.8°C. Наиболее холодный месяц – январь со средней температурой –14.2°C. Из-за далекого расположения от океанов зима здесь холоднее, чем в городах, находящихся западнее ее на той же широте (Воронеж, Киев, Лейпциг, Лондон). Нур-Султан – одна из самых холодных столиц в мире (вторая после Улан-Батора).
Почвы в городе каштановые, характерные для сухостепной зоны, обедненные гуминовыми веществами. Этим обусловлены известные трудности в создании зон зеленых насаждений, которые должны обеспечивать комфортную среду, смягчать жесткие климатические условия.
К югу от уже существующего города на значительной территории, ранее занятой садовыми товариществами, был построен новый центр столицы. При этом прежний ландшафт был полностью уничтожен: грунт снят и заменен на новый, выстроены многоэтажные современные кварталы, разбиты парки. Таким образом, был создан совершенно новый городской ландшафт. В самом центре новой столицы разбит Президентский парк, с запада на него выходят окна зданий Президентского дворца, Сената и Правительства Казахстана, а с востока – крупнейшей мечети региона Казахстана и Средней Азии (Хазрет Султан), Университета искусств, Дворца независимости и Национального музея Казахстана. В центре парка возвышается пирамида Дворца мира и согласия (рис. 1). Здесь в 2017 г. мы обнаружили поселение самого крупного из всех современных видов хомяков – хомяка обыкновенного Cricetus cricetus. В последние 50 лет этот вид демонстрирует явную тенденцию к заселению городов (Feoktistova et al., 2013; Суров и др., 2015) на фоне резкого сокращения численности в природных биотопах (Surov et al., 2016). Ранее мы исследовали ситуации, когда обыкновенный хомяк уже десятки лет существовал в конкретном городе, как правило, с уже сложившейся городской структурой (Симферополь, Нальчик, Кисловодск). Нами были выявлены некоторые закономерности пространственного распределения поселений хомяка в этих городах, особенности их генетической структуры (Феоктистова и др., 2016, 2019). В случае с Нур-Султаном процесс заселения видом урбоценоза города должен был начаться “с чистого листа” приблизительно 12 лет назад, когда был сформирован современный ландшафт парка.
Рис. 1.
Распределение жилых нор обыкновенного хомяка в Президентском парке Нур-Султана. Точки – норы, штриховые линии – границы экспериментальных площадок № 1 и 2. Светло-серый фон – озелененные участки (газоны, клумбы, древесные и кустарниковые насаждения) на территории парка. Темно-серый фон – река Ишим (искусственное русло) и искусственные водоемы в Президентском парке. Черный фон – основные здания на территории Президентского парка или примыкающие к ней: 1 – Пирамида (Дворец мира и согласия), 2 – Президентский дворец (Акорда), 3 – мечеть Хазрет Султан, 4 – Университет искусств, 5 – Дворец независимости, 6 – Национальный музей.
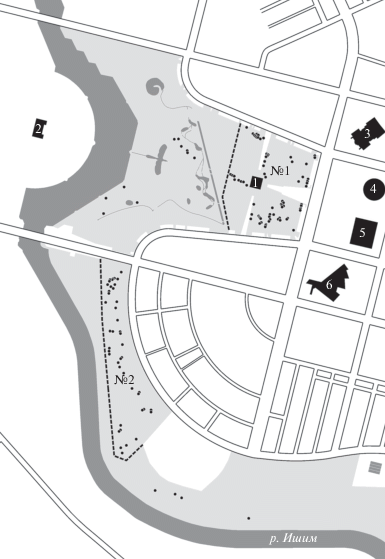
Цель данного исследования – описание условий обитания и генетической структуры популяции обыкновенного хомяка на начальной стадии формирования городской среды.
МАТЕРИАЛЫ И МЕТОДЫ
Материал был собран на территории Президентского парка в Нур-Султане в 2017–2018 гг. В 2017 г. при посещении парка было отмечено присутствие там обыкновенного хомяка – встречены сами зверьки и обнаружены жилые норы. В августе 2018 г. мы провели пешее визуальное обследование территории в центральной части Нур-Султана (включая и Президентский парк). Два или три исследователя, двигаясь челноком, отмечали выходы жилых нор хомяка и фиксировали их местоположение с помощью GPS. Выходы, расположенные ближе 1 м один от другого, учитывались как одна нора.
Помимо обследования проводили отловы животных в местах наибольшей концентрации нор: в восточной части парка вокруг пирамиды Дворца мира и согласия (площадка № 1 (20 га)), занимающая всю территорию первой очереди парка) и в южной части (площадка № 2, занимающая часть территории третьей очереди парка). Площадки отстоят одна от другой на расстоянии 1.5 км и разделены широкой автомагистралью (рис. 1).
Для отловов использовали сетчатые живоловушки размером 40 × 12 × 12 см, которые экспонировали около жилых нор в вечернее время и проверяли утром после рассвета. В качестве приманки использовали свежие корнеплоды (морковь, свеклу), смоченные нерафинированным подсолнечным маслом. Ловушку накрывали листовым опадом и травой во избежание обнаружения посетителями парка. Всего было отработано 48 ловушко-ночей и отловлено 13 особей обыкновенного хомяка.
Пойманных животных взвешивали на электронном безмене с точностью до 1 г. Затем проводили стандартные зоологические измерения, определяли пол, репродуктивный статус животных. Также отрезали небольшие кусочки ушной раковины для генетического анализа и выстригали небольшой участок шерсти для определения содержания в организме тяжелых металлов. Для определения уровня загрязнения тяжелыми металлами мест обитания хомяков было собрано 10 образцов почвы в разных точках Президентского парка.
Пойманных животных выпускали вечером того же дня на месте поимки. Одна взрослая беременная самка (площадка № 2) погибла в ловушке. Образцы ткани от ее 13 эмбрионов примерно двухнедельного возраста были индивидуально зафиксированы в 96%-ном этаноле для дальнейшего генетического анализа.
В качестве маркера филогенетических линий были использованы два локуса митохондриальной ДНК (мтДНК): контрольный регион и ген цитохрома b (cytb). Также проводили определение аллельного состава 10 микросателлитных локусов (Ccrμ3, Ccrμ4, Ccrμ10, Ccrμ11, Ccrμ12, Ccrμ13, Ccrμ15, Ccrμ17, Ccrμ19, Ccrμ20 (по: Neumann et al., 2004)). Анализ был проведен для 12 взрослых особей (шести с площадки № 1 и шести с площадки № 2) и 13 эмбрионов.
Метод выделения ДНК, условия амплификации и методики определения нуклеотидных последовательностей мтДНК и аллельного состава микросателлитных локусов были описаны ранее (Феоктистова и др., 2016, 2019; Surov et al., 2019). Математическая обработка полученных данных сводилась:
к определению различий между выявленными индивидуальными последовательностями мтДНК животных, пойманных в Нур-Султане и других районах Казахстана;
к анализу обобщенной матрицы генотипов 12 взрослых животных из Нур-Султана методом кластеризации (с указанием принадлежности каждой особи к выборке с конкретной площадки – модель admixture locprior) в программе Structure v.2.3.4. (Pritchard et al., 2000);
к сравнению частот аллелей микросателлитных локусов в выборках взрослых животных с площадок № 1 и 2 (Fst-критерий) с использованием программы Arlequin v.3.11 (Excoffier et al., 2005). В обоих случаях генотипы эмбрионов учтены не были. Параметры расчетов были аналогичны использованным ранее (Феоктистова и др., 2016, 2019).
Аллельный состав каждого из 10 микросателлитных локусов, определенный для каждого из 13 эмбрионов, извлеченных из погибшей самки, сравнивали с генотипом самой самки с определением аллелей, которые могли быть получены от матери и одного (не более двух дополнительных аллелей на локус в выводке) или нескольких потенциальных отцов.
Качественное и количественное определение содержания тяжелых металлов и химически токсичных элементов в образцах почвы (объединенная проба десяти образцов) и в шерсти хомяков (объединенная проба от 12 взрослых животных) проводилось методом рентгенофлуоресцентного анализа, основанном на эффекте полного отражения (TXRF) на спектрометре S2 PICOFOX (Bruker AXS, Германия; Свидетельство об утверждении типа средств измерений DE.C.31.076.A № 44359). Образцы почвы и биологического материала анализировались по методике ISO/TS 18705:2015 (Surface chemical analysis use of Total Reflection X‑ray Fluorescence spectroscopy in biological and environmental analysis).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
При обследовании территории всего Президентского парка была обнаружена 131 жилая нора обыкновенного хомяка – средняя плотность ~1 нора/га. На площадке № 1, расположенной в более старой части парка, были отмечены 73, а на площадке № 2 – 44 норы. Распределялись норы неравномерно, с максимальной локальной плотностью до 14 нор/га. Наибольшая плотность нор была в частях парка с плодовыми деревьями и кустарниками, которые могли обеспечить зверьков кормами и хорошими убежищами. Кроме того, норы отмечались вдоль фундаментов, парапетов и насыпей.
Определять достоверно возраст пойманных зверьков не представлялось возможным, но, судя по морфометрическим признакам, в основном они были сеголетками.
Анализ аллельной композиции 10 микросателлитных локусов 13 эмбрионов и самки-матери не выявил “избыточных” аллелей, свидетельствовавших о факте множественного отцовства. С высокой вероятностью все детеныши выводка происходили от одного отца.
Анализ методом кластеризации по 10 микросателлитным локусам обобщенной выборки от 12 взрослых животных, пойманных на двух площадках, не выявил наличия какой-либо популяционной структуры. Наиболее вероятным стал результат анализа для K = 1 (где K – параметр, соответствующий предполагаемому числу генетически обособленных групп в общей выборке), из чего следует, что население хомяков на обеих площадках представляет собой единую, генетически однородную популяцию. Генетическая дистанция между двумя выборками взрослых животных из поселений, разделенных расстоянием 1.5–2 км (площадки № 1 и 2), оказалась крайне незначительной (Fst = 1.26%), а разница между этими выборками – статистически недостоверной (P = 0.381).
Для всех пойманных в Нур-Султане животных был отмечен единый митохондриальный гаплотип – последовательность, депонированная в базе данных GenBank NCBI под номером MK202466 (ген cytb) в сочетании с последовательностью GenBank KY795998 (контрольный регион). Из других известных вариантов последовательности данного участка наиболее близким (одна нуклеотидная замена на 1803 пары нуклеотидов) оказывается гаплотип, отмеченный нами у особи обыкновенного хомяка, пойманного в районе с. Тургай Акмолинской обл. Казахстана (GenBank KF271761 + GenBank KY795998) примерно в 100 км к востоку от Нур-Султана. Суммарное число аллелей в 10 исследованных локусах в выборке хомяков из Нур-Султана (для n = 25, с учетом генотипов эмбрионов) составило 46.
Данные о содержании химических веществ, включая тяжелые металлы в почве и в шерсти животных, обитающих в городских парках Симферополя и Нур-Султана, приведены в табл. 1.
Таблица 1.
Концентрации (мг/кг воздушно-сухой массы) ряда химических элементов (в том числе тяжелых металлов) в пробах почвы и в шерсти обыкновенного хомяка в Президентском парке в Нур-Султане и в Парке им. Ю.А. Гагарина в Симферополе
| Химический элемент | Симферополь | Нур-Султан | ||
|---|---|---|---|---|
| почва | шерсть | почва | шерсть | |
| Na | 5.73 | 6.23 | 0 | 0 |
| Mg | 0.48 | 0.454 | 0.058 | 0.108 |
| Al | 0.224 | 0.218 | 0 | 0 |
| Si | 5.1 | 26.75 | 0 | 0 |
| P | 1.0 | 8.95 | 0 | 0 |
| S | 19.87 | 3.345 | 0 | 0 |
| Cl | 0 | 0.290 | 0 | 0 |
| K | 2.283 | 3.61 | 0 | 0 |
| Ca | 24.14 | 10.46 | 0 | 0 |
| Ti | 0.68 | 0.26 | 0 | 0.00017 |
| Cr | 0.92 | 0.017 | 0 | 0 |
| Mn | 0.3 | 0.397 | 0 | 0 |
| Fe | 135.81 | 238.25 | 21507 | 35.02 |
| Ni | 0.057 | 0.0012 | 0.0001 | 0.0007 |
| Cu | 0.59 | 1.99 | 15.04 | 1.36 |
| Zn | 28.82 | 58.125 | 104.8 | 28.5 |
| As | 0.081 | 0.166 | 0.0006 | 0.0013 |
| Se | 0.01 | 0.010 | 0 | 0 |
| Sr | 0.074 | 26.3 | 20.34 | 0.133 |
| Ba | 0.32 | 152.4 | 30.06 | 0.003 |
| Re | 0.226 | 0.538 | 0 | 0 |
| Pb | 1.93 | 1.995 | 0.738 | 0.118 |
| Mo | 0.002 | 0.00645 | 0 | 0 |
| Hg | 0 | 0.000240 | 0.004 | 0 |
| Co | 0 | 0.013 | 0 | 0 |
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Согласно анализу материалов космической съемки средствами ресурса Google Earth Pro Президентский парк, в настоящее время расположенный в современном центре города, был сооружен в три очереди на территории, большая часть которой еще в 2004 г. была занята садовыми участками. В 2006 г. было закончено строительство восточной части парка (первой очереди) площадью 20 га вокруг пирамиды Дворца мира и согласия. На территории западной части парка (второй очереди площадью ~60 га) в 2006 г. проводились активные устроительные работы, завершившиеся в 2008 г. На части территории третьей очереди парка (его южной части площадью ~135 га) в 2011–2012 гг. еще сохранялись садовые участки. Завершенный вид парк приобрел в 2014 г.
Почва для создания искусственного рельефа в Президентском парке (серозем) завозилась из внегородских источников, в качестве естественных удобрений вносились торф и навоз. Согласно имеющимся сведениям о ходе устройства парка на его территории исходно было высажено 43 тыс. декоративных деревьев и кустарников: боярышник, яблони, груши, терн и т.д. Примечательно, что для посадки использовали только взрослые растения (так называемые крупномеры), корневая система которых была помещена в специальные металлические сетчатые корзины. Кроме того, в парке разбили цветочные клумбы и установили систему автоматического орошения. В парке много каналов, фонтанов и малых архитектурных форм, напоминающих гроты.
Архитекторы, создавшие парк, руководствовались идеей искусственного, но максимально совершенного ландшафта, напоминающего природный. Для этого высаживались в основном взрослые уже плодоносящие растения, создавались искусственный всхолмленный рельеф, а также система каналов и фонтанов, которые должны были создавать комфортную атмосферу для посетителей в период летнего зноя. Однако условия, комфортные для человека, оказались также благоприятной средой для обыкновенного хомяка. Мягкий грунт, удобный для устройства нор, кормовая база в виде плодов и семян, пересеченный рельеф и практически полное отсутствие хищников (выгул собак в парке запрещен) обеспечили весьма благополучное существование для этого вида-синурбиста. Кроме того, преимущественно ночная активность, а также разветвленная система нор позволяют хомякам быть практически невидимыми для людей.
Вероятнее всего вновь созданный городской парк был заселен хомяком с территории существовавших на этом месте и на смежных землях садовых участков. Заселение восточной части парка в связи с интенсивными работами по переустройству этой территории не могло начаться раньше 2006 г. Таким образом, возраст существующих здесь поселений хомяка не превышает 12 лет. Обустройство западной части парка завершилось двумя годами позже, однако условия на этой территории, видимо, оказались менее подходящими для хомяка и число обнаруживаемых здесь нор остается небольшим. В заселении южной части парка (возраст существующих здесь поселений хомяков не более 5 лет) могли участвовать как особи-мигранты из соседних районов, так и зверьки из местной популяции, сохранившиеся в каком-то количестве в ходе работ по перестройке территории.
Наибольшая концентрация нор в Президентском парке в Нур-Султане наблюдается в наиболее старой части парка и в местах с более плотными посадками плодовых культур (рис. 1). Ранее мы показали, что норы обыкновенного хомяка в Парке им. Ю.А. Гагарина в Симферополе также располагаются неравномерно и достигают наибольшей концентрации в частях парка с более густой древесной и кустарниковой растительностью (Katzman et al., 2018). Условия в таких местах облегчают устройство нор, обеспечивают защиту от хищников и отличаются хорошей кормовой базой. Однако максимальная плотность нор в парке Симферополя (>40 нор/га в летний период) намного превышает отмеченную для Президентского парка в Нур-Султане (до 14 нор/га).
Все особи, пойманные нами на территории Президентского парка как в более старой восточной, так и в более молодой южной его частях, оказались носителями одного гаплотипа мтДНК. Поскольку закрепление в популяциях грызунов вновь возникающих в результате мутаций вариантов последовательностей происходит крайне редко (раз в тысячелетия), возможностью появления новых гаплотипов непосредственно в городских популяциях можно пренебречь. Отмечаемое в городах число митохондриальных линий свидетельствует о разнообразии особей-основателей, проникавших на городскую территорию с момента начала ее заселения видом. Так, в поселениях обыкновенного хомяка в Симферополе присутствуют три, а в Кисловодске – пять митохондриальных линий (Феоктистова и др., 2016, 2019). Сведений о разнообразии гаплотипов в популяциях обыкновенного хомяка в окрестностях Нур-Султана у нас нет, однако в выборках из соседних районов и областей (Темиртау, Щучинск, Тургай) обнаружено не менее трех разных гаплотипов исследованного участка мтДНК, а всего на территории Северного Казахстана в настоящее время известно девять гаплотипов (Feoktistova et al., 2018). Присутствие во всей городской популяции обыкновенного хомяка в Нур-Султане единственной митохондриальной линии можно рассматривать как эффект основателя – “митохондриальную Еву” локального масштаба.
Число аллелей микросателлитных локусов, отмеченное у хомяков из Нур-Султана – 46 при значении показателя allelic richness (AR) – 44.3, меньше такового, известного для Кисловодска (64 аллеля, AR = 56.0) и Симферополя (55 аллелей, AR = 47.3), на территории которых хомяк обитает давно. В отсутствие сведений о генетическом разнообразии популяций хомяка в окрестностях города этот факт не может трактоваться однозначно, но и не противоречит предположению об относительно небольшом числе особей-основателей формирующейся городской популяции Нур-Султана.
Для выводков обыкновенного хомяка из Симферополя и его окрестностей нами был установлен факт множественного отцовства – в одновременном оплодотворении яйцеклеток самки принимала участие сперма двух и более самцов (Surov et al., 2019; неопубликованные данные). Для выводка из нурсултанской популяции обыкновенного хомяка множественное отцовство подтверждено не было. Отсутствие множественного отцовства в популяциях вида, которому это явление свойственно, можно трактовать как эффект низкой плотности (недостаток зрелых самцов). Однако, чтобы сделать окончательные заключения, необходимо проанализировать большее число выводков и большее число генотипов особей – потенциальных отцов.
Следует отметить, что число эмбрионов у исследованной в Нур-Султане самки (13 жизнеспособных и два резорбированных) демонстрирует весьма высокий уровень плодовитости в изучаемой популяции. Среднее число эмбрионов для обыкновенного хомяка в период между 1976 и 1986 гг. (т.е. до начала массовой депрессии численности) оценивается как 12.61 ± 1.12 (Surov et al., 2016).
Поселения обыкновенного хомяка в Симферополе и Кисловодске представляют собой обособленные субпопуляции (демы). Уровень генетических отличий (оцененный по частотам аллелей того же набора локусов) между субпопуляциями, населяющими расположенные в 1.5–4 км один от другого районах собственно урбанизированной территории, может составлять 12–33%. На окраинах городов, представляющих собой зеленую зону и вплотную примыкающих к пригородной (неурбанизированной) территории, различия между локальными группировками снижаются до 5–9% (Феоктистова и др., 2016, 2019). Ситуация, наблюдаемая нами в настоящий момент в Нур-Султане, аналогична той, что характерна для категории зеленой зоны, примыкающей к неурбанизированной территории. Однако в Нур-Султане генетические различия между двумя поселениями хомяков, разделенными расстоянием 1.5–2 км, оказываются еще ниже (практически отсутствуют). Этот факт можно объяснить продолжающимся становлением структуры населения хомяков в парке, сопровождающимся активной миграцией и расселением особей по территории. Однако даже если принимать во внимание определенную территориальную разобщенность групп, населяющих восточную и южную части парка, не следует забывать, что произошли все зверьки, скорее всего, от одного и того же относительно небольшого пула особей-основателей. Нескольких лет (соответствующих нескольким поколениям) разницы между заселением хомяками разных частей парка, очевидно, недостаточно для заметного изменения частот аллелей под влиянием генетического дрейфа. Поэтому крайне интересно продолжить наблюдения с регистрацией динамики отдельных поселений и уровня генетических отличий между ними в последующие годы.
Тяжелые металлы, содержащиеся в окружающей среде, обладают способностью накапливаться в организме. В дальнейшем их высокие концентрации могут быть одним из факторов сокращения продолжительности жизни животных. Попавшие в организм тяжелые металлы полностью сохраняются в волосах, что позволяет проводить анализ их накопления в течение всей жизни вне зависимости от состояния условий в момент поимки животного. Ранее нами были исследованы образцы почвы из Парка им. Ю.А. Гагарина в Симферополе и шерсти обитающих там обыкновенных хомяков. Как показывают результаты сравнения исследованных выборок из двух городов (табл. 1), в Симферополе содержание исследуемых веществ, в том числе тяжелых металлов, как в почве, так и в шерсти животных существенно выше, чем в Нур-Султане, где большая часть исследованных веществ вообще не была обнаружена либо присутствовала в дозах, близких к порогу чувствительности прибора. Это относится и к такому типичному загрязнителю города, как свинец. Очевидно, городская среда в столице Казахстана пока слабо загрязнена промышленными и автотранспортными отходами.
ЗАКЛЮЧЕНИЕ
Правительство Республики Казахстан уделяет большое внимание проблемам экологии, а вопросы озеленения городских территорий возведены в ранг государственной кампании. Под руководством первого президента Казахстана Нурсултана Назарбаева в столице были начаты работы по созданию зеленого пояса (лесопосадки кольцом окружают город и смыкаются с 10 парками и множеством скверов). В 2017 г. в городе был заложен Ботанический сад. По имеющимся планам число деревьев в черте города должно достичь 1 млн, а число городских парков предполагается довести в скором времени до 17.
Непреднамеренно поставленный эксперимент – заселение вновь созданного городского парка видом-синурбистом обыкновенным хомяком – предоставляет уникальную возможность проследить не только за развитием популяции этого вида в городе, но и за становлением всего урбоценоза. Ранее мы имели возможность оценить генетическую структуру поселений обыкновенного хомяка в давно сложившихся городах, не имея информации о том, когда и сколько раз хомяки могли вселяться на эту территорию. В Нур-Султане такая точка отсчета есть. Данная работа лишь первый шаг в изучении процесса первичного формирования и дальнейшего становления городского населения обыкновенного хомяка. Будущие исследования должны показать, как с течением времени и на фоне развития всего урбоценоза будут меняться численность этого вида в городе, пространственная, биотопическая и генетическая структура его поселений.
Работа выполнена при финансовой поддержке Программы фундаментальных исследований Президиума РАН “Биоразнообразие природных систем и биологические ресурсы России” по теме “Мониторинг биоразнообразия природных систем и биологические ресурсы России”, а также РФФИ (грант 17-04-01061).
Список литературы
Карасёва Е.В., Телицына А.Ю., Самойлов Б.Л. Млекопитающие Москвы в прошлом и настоящем. М.: Наука, 1999. 244 с.
Сорокин А.Г. Современные проблемы сохранения и восстановления популяций редких птиц // Сохранение и восстановление биоразнообразия. Разд. V. М.: Изд-во Научного и учебно-методического центра, 2002. С. 131–147.
Суров А.В., Поплавская Н.C., Богомолов П.Л., Кропоткина М.В., Товпинец Н.Н., Кацман Е.А., Феоктистова Н.Ю. Синурбанизация обыкновенного хомяка (Cricetus cricetus L., 1758) // Рос. журн. биол. инвазий. 2015. № 4. С. 105–117.
Феоктистова Н.Ю., Мещерский И.Г., Суров А.В., Богомолов П.Л., Товпинец Н.Н., Поплавская Н.С. Генетическая структура городской популяции обыкновенного хомяка (Cricetus cricetus) // Генетика. 2016. Т. 52. № 2. С. 221–230.
Феоктистова Н.Ю., Мещерский И.Г., Богомолов П.Л., Мещерский С.И., Поплавская Н.С., Чунков М.М., Юферева В.В., Тельпов В.А., Суров А.В. Обыкновенный хомяк (Cricetus cricetus) в Предкавказье: генетическая структура городских и пригородных популяций // Генетика. 2019. Т. 55. № 3. С. 337–348.
Boggie M.A., Mannan R.W. Examining seasonal patterns of space use to gauge how an accipiter responds to urbanization // Landscape Urban Planning. 2014. V. 124. P. 34–42.
Čanady A. New site of the European hamster (Cricetus cricetus) in the urban environment of Košice city (Slovakia) // Zool. Ecol. 2013. V. 23. № 1. P. 61–65.
Collins J.P., Kinzig A., Grimm N.B., Fagan W.F., Hope D., Wu J., Borer E.T. A new urban ecology. Modeling human communities as integral parts of ecosystems poses special problems for the development and testing of ecological theory // Am. Sci. 2000. V. 88. P. 416–425.
Dornelas M., Gotelli N.J., McGill B., Shimadzu H., Moyes F., Sievers C., Magurran A.E. Assemblage time series reveal biodiversity change but not systematic loss // Science. 2014. V. 344. P. 296–299. https://doi.org/10.1126/science.1248484
Excoffier L., Laval G., Schneider S. Arlequin ver. 3.0: An integrated software package for population genetics data analysis // Evol. Bioinform. Online. 2005. V. 1. P. 47–50.
Feoktistova N.Yu., Meschersky I.G., Bogomolov P.L., Meschersky S.I., Sayan A.S., Sitnikova E.F., Vlasov A.A., Surov A.V. About the border between two phylogroups of the common hamster (Cricetus cricetus) (Cricetinae, Rodentia) // Povolzh. J. Ecol. 2018. № 4. P. 485–494. https://doi.org/10.18500/1684-7318-2018-4-485-494
Feoktistova N.Y., Surov A.V., Tovpinetz N.N., Kropotkina M.V., Bogomolov P.L., Siutz C., Haberl W. The common hamster as a sinurbist a history of settlement in European cities // Zool. Polon. Ed. Kotusz J. Pol. Towarzystwo Zool. 2013. V. 58. № 3–4. P. 113–126.
Katzman E.A., Zaytseva E.A., Feoktistova N.Yu., Tovpinetz N.N., Bogomolov P. L., Potashnikova E.V., Surov A.V. Seasonal changes in burrowing of the Common hamster (Cricetus crictus L., 1978) (Rodentia: Cricetidae) in the city // Povolzh. J. Ecol. 2018. № 3. P. 251–258.
Lahr E.C., Dunn R.R., Frank S.D. Getting ahead of the curve: cities as surrogates for global change // Proc. Roy Soc. B: Biol. Sci. 2018. https://doi.org/10.1098/rspb.2018.0643
Morinha F., Ramos P.S., Gomes S., Mannan R.W., Guedes-Pinto H., Bastos E. Microsatellite markers suggest high genetic diversity in an urban population of Cooper’s hawks (Accipiter cooperii) // J. Genet. 2016. V. 95. P. 19–24.
O’Brien K. Global environmental change II: from adaptation to deliberate transformation // Prog. Human Geogr. 2012. V. 36. P. 667–676.
Pritchard J.K., Stephens M., Donnelly P.J. Inference of population structure using multilocus genotype data // Genetics. 2000. V. 155. P. 945–959.
Schilthuizen M. Darwin comes to town: How the urban jungle drives evolution. N.Y.: Macmillan Publishing Group, LLC., 2018. 304 p.
Surov A., Banaszek A., Bogomolov P., Feoktistova N., Monecke S. Dramatic global decrease in the range and reproduction rate of the European hamster Cricetus cricetus // Endangered Species Res. 2016. P. 119–145.
Surov A.V., Zaytseva E.A., Kuptsov A.V., Katzman E.A., Bogomolov P.L., Sayan A.S., Potashnikova E.V., Tovpinetz N.N., Kuznetsova E.V., Tsellarius A.Y., Feoktistova N.Y. Circle of life: The common hamster (Cricetus cricetus l., 1758) adaptations to the urban environment // Integrat. Zool. 2019. V. 14. № 4. P. 383–395.
von der Lippe M., Säumel I., Kowarik I. Cities as drivers for biological invasions – the role of urban climate and traffic // Die Erde. 2005. V. 136. № 2. P. 123–143.
World Urbanization Prospects. The 2018 Revision. N.Y.: United Nations, Department of Economic and Social Affairs, Population Division, 2019. 103 p.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая