

Известия РАН. Серия биологическая, 2020, № 2, стр. 177-185
Использование разных систем признаков в фенетике популяций
П. Н. Кораблев 1, М. П. Кораблев 2, *, Н. П. Кораблев 3, 4, И. Л. Туманов 5
1 Центрально-Лесной государственный природный биосферный заповедник
пос. Заповедный, Нелидовский р-н, Тверская обл., 172513 Россия
2 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
Ленинский просп., 33, Москва, 119991 Россия
3 Великолукская государственная сельскохозяйственная академия
просп. Ленина, 2, Великие Луки, Псковская обл., 182112 Россия
4 Псковский государственный университет
пл. Ленина, 2, Псков, 180000 Россия
5 Всероссийский научно-исследовательский институт охотничьего хозяйства и звероводства
им. Б.М. Житкова, Преображенская ул., 79, Киров, 610000 Россия
* E-mail: mir-kor@yandex.ru
Поступила в редакцию 26.08.2019
После доработки 25.10.2019
Принята к публикации 25.10.2019
Аннотация
Проведен сравнительный анализ фенофонда популяций млекопитающих на основе двух групп признаков: одонтологических и собственно краниологических. Всего изучено 1637 черепов семи видов. Показано, что значения популяционных характеристик (внутрипопуляционное разнообразие, доля редких фенов, флуктуирующая асимметрия) для двух групп признаков достоверно различаются. Предположены существование большей степени генного контроля в формировании одонтологических признаков и их меньшая реактивность в отношении средовых факторов по сравнению с краниологическими. Выявлены достоверные корреляции между значениями флуктуирующей асимметрии для разных групп признаков и достоверная положительная связь между флуктуирующей асимметрией и разнообразием. Предложено комплексное использование двух систем признаков для выявления начальных стадий нарушений индивидуального развития и оценки глубины этих нарушений при изучении стабильности онтогенеза по величине флуктуирующей асимметрии в природных популяциях млекопитающих.
Фенетический подход как самостоятельное направление изучения природных популяций сформировался в 1980-х гг. во многом благодаря А.В. Яблокову (Яблоков, 1980, 1987). Однако столь же широкого распространения, как классические морфологические методы, изучение неметрических вариаций признаков организмов не получило. Одна из вероятных причин этого – начало массового использования молекулярно-генетических методов параллельно с развитием фенетики. Когда методы, позволяющие изучать генетический полиморфизм, стали практически общедоступны, могло показаться, что фенетические подходы устарели и потеряли свою актуальность. В некоторой степени это справедливо, если фенетику считать частью менделевской генетики. Однако если рассматривать фенетику как часть морфологии, то можно ожидать, что она будет вновь востребована при осмыслении результатов сравнительного изучения популяционного разнообразия. К этому имеется ряд предпосылок. Прежде всего, не следует забывать, что фенотипическая изменчивость отражает важные адаптивные свойства живой материи и отбор в локальных популяциях идет именно по фенотипам. Причинно-следственные связи генотипа и фенотипа столь сложны, что их обсуждение выходит за рамки биологии и нередко принимает философские формы, являясь частью фундаментальной дискуссии в рамках теории эволюции. В последние десятилетия внесено немало нового в развитие эволюционных идей. Обобщив и осмыслив научные достижения в популяционной генетике, Назаров (Назаров, 2007, стр. 450) сделал важное применительно к нашим рассуждениям о важности изучения фенотипа заключение, что виды осуществляют свою адаптивную стратегию “осмысленно” через адаптациогенез, а “… термин “микроэволюция” утрачивает свою легитимность”, поскольку популяционные процессы не ведут к видообразованию. Фактически признается, “что генетические изменения не диктуют организму, что ему делать, а лишь фиксируют в своей генетической памяти его правильные эволюционные шаги” (Назаров, 2007, стр. 420). Пока молекулярно-генетические исследования и изучение фенетической структуры популяций проводятся параллельно и, как правило, независимо одно от другого, однако они должны пересечься, когда появятся корректные подходы к их сравнительному анализу.
Егоров, ссылаясь, на работы Берри (Berry, 1963), Яблокова (Яблоков, 1980) и Тимофеева-Ресовского с соавт. (Тимофеев-Ресовский и др., 1973), считает, что “признаки, относимые к разряду фенетических, более точно отражают генотип популяции по сравнению с признаками, имеющими непрерывную изменчивость – линейными, весовыми и т.д.” (Егоров, 1983, стр. 99). Изучение фенетических признаков, имеющих эпигенетическую природу, позволяет проводить сравнительный анализ фенофонда популяций на гораздо меньшей географической шкале, чем использование пластических признаков. Последние, безусловно, интересны при изучении морфологических адаптаций видов в спектре параметров среды обитания в границах ареалов, в то время как анализ частот проявления неметрических вариаций позволяет изучать внутрипопуляционную структуру и дифференциацию, о которой мы знаем гораздо меньше, чем о внутривидовой.
Явления полигении и плейотропии делают практически невозможным определение составляющей наследственности или внешних факторов в изменчивости большинства признаков. Вместе с тем при изучении фенофонда популяций млекопитающих близких и отдаленных систематических групп мы столкнулись с тем, что разные группы признаков дают достоверно различающиеся значения популяционных характеристик. Это позволяет хотя бы на качественном уровне судить о степени их генетической детерминированности или, напротив, о реактивности на изменения экзогенных факторов.
Цель работы – анализ использования разных групп признаков в популяционно-фенетических исследованиях.
МАТЕРИАЛЫ И МЕТОДЫ
Нами обобщены и осмыслены ранее полученные данные о фенофонде популяций лося Alces alces L. 1758 (Кораблев, 2003), европейской норки Mustela lutreola L. 1758 (Кораблев, 2013), американской норки Neovison vison Baryshnikov et Abramov 1997 (Schreber, 1777) (Кораблев и др., 2012а; 2016б), лесного хоря Mustela putorius L. 1758 (Кораблев и др., 2015), лесной куницы Martes martes L. 1758 (Кораблев и др., 2016а), лисицы Vulpes vulpes L. 1758 и енотовидной собаки Nyctereutes procyonoides Gray, 1834 (Кораблев и др., 2012а) в центре европейской части России (Тверская обл. и прилегающие к ней территории Вологодской, Новгородской, Псковской и Смоленской областей). Эти работы объединяет то, что при характеристике фенофонда популяций использовались две группы признаков: одонтологические и собственно краниологические. Для хищных млекопитающих признаки зубов были одинаковые и представляли собой описание дополнительных элементов (бугорков или корешков) между основными корнями премоляров и моляров (Кораблев и др., 2005). Для лося одонтологические признаки характеризовали строение лингвальной и вестибулярной поверхности коронок коренных зубов (Кораблев, 1997).
На основе частот проявления вариаций признаков были получены показатели, характеризующие степень и структуру внутрипопуляционного разнообразия, сходства выборок (Животовский, 1982), а также уровень флуктуирующей асимметрии (Захаров, 1987). Фены регистрировали на правой и левой сторонах черепа, для расчетов использовали среднюю частоту между сторонами. Для статистического анализа были сформированы выборки одинакового объема по 50 черепов. Если объем материала значительно превышал 50 черепов, он разбивался на выборки, близкие к стандартной (50 экземпляров), для каждой из которых были получены значения популяционных характеристик. В сравнительном межвидовом анализе были использованы средние значения для выборок по видам. Всего изучили 1637 черепов, в том числе: черепа лося (A.a.) – 176, европейской норки (M.l.) – 103, американской норки (N.v.) – 367, лесного хоря (M.p.) – 250, лесной куницы (M.m.) – 250, лисицы (V.v.) – 103, енотовидной собаки (N.p.) – 379.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В табл. 1–3 приведены диапазоны значений параметров, характеризующих степень и структуру внутрипопуляционного разнообразия, фенетические дистанции (показатель сходства) и стабильность онтогенеза семи видов млекопитающих. Данные, характеризующие диапазон значений популяционных параметров выборок каждого вида и усредненные данные по видам (рис. 1) свидетельствуют, что значения показателей существенно различаются для групп признаков.
Таблица 1.
Диапазоны значений параметров, характеризующих степень (µ) и структуру (h) внутрипопуляционного разнообразия млекопитающих (min–max)
| Вид | µ | h | ||
|---|---|---|---|---|
| одонтологические | краниологические | одонтологические | краниологические | |
| M.l. | 1.58 | 2.52 | 0.29 | 0.21 |
| N.v. | 1.69–1.97 | 2.41–2.83 | 0.28–0.37 | 0.18–0.26 |
| M.p. | 1.64–1.86 | 2.55–2.98 | 0.16–0.30 | 0.16–0.1 |
| M.m. | 1.14–1.26 | 1.84–2.28 | 0.12–0.24 | 0.20–0.24 |
| V.v. | 1.13–1.16 | 2.18–2.47 | 0.17–0.26 | 0.22–0.28 |
| N.p. | 1.56–1.87 | 2.50–2.74 | 0.40–0.44 | 0.24–0.31 |
| A.a. | 1.66–1.93 | 2.03–2.20 | 0.19–0.27 | 0.17–0.26 |
Примечание. M.l. – европейская норка, N.v. – американская норка, M.p. – лесной хорь, M.m. – лесная куница, V.v. – лисица, N.p. – енотовидная собака, A.a. – лось; для табл. 1–3.
Таблица 2.
Значения показателя сходства (r) между выборками на основе попарного сравнения (min–max/среднее)
| Вид | Одонтологические | Краниологические |
|---|---|---|
| N.v. | $\frac{{0.971{\kern 1pt} - {\kern 1pt} 0.990}}{{0.982 \pm 0.002}}$ | $\frac{{0.974{\kern 1pt} - {\kern 1pt} 0.995}}{{0.985 \pm 0.002}}$ |
| M.p. | $\frac{{0.985{\kern 1pt} - {\kern 1pt} 0.993}}{{0.992 \pm 0.001}}$ | $\frac{{0.977{\kern 1pt} - {\kern 1pt} 0.992}}{{0.986 \pm 0.002}}$ |
| M.m. | $\frac{{0.995{\kern 1pt} - {\kern 1pt} 0.997}}{{0.996 \pm 0.001}}$ | $\frac{{0.989{\kern 1pt} - {\kern 1pt} 0.997}}{{0.993 \pm 0.001}}$ |
| V.v. | 0.997 ± 0.0003 | 0.973 ± 0.003 |
| N.p. | $\frac{{0.976{\kern 1pt} - {\kern 1pt} 0.981}}{{0.979 \pm 0.002}}$ | $\frac{{0.919{\kern 1pt} - {\kern 1pt} 0.954}}{{0.939 \pm 0.007}}$ |
| A.a. | $\frac{{0.982{\kern 1pt} - {\kern 1pt} 0.988}}{{0.984 \pm 0.001}}$ | $\frac{{0.975{\kern 1pt} - {\kern 1pt} 0.989}}{{0.984 \pm 0.001}}$ |
Таблица 3.
Диапазоны значений показателя флуктуирующей асимметрии разных групп признаков млекопитающих, % (min–max)
| Вид | Одонтологические | Краниологические |
|---|---|---|
| M.l. | 10.4–12.7 | 26.4–29.2 |
| N.v. | 7.8–12.6 | 23.9–37.4 |
| M.p. | 10.2–13.5 | 29.2–41.8 |
| M.m. | 2.3–4.8 | 23.0–30.2 |
| V.v. | 1.2–1.7 | 22.6–27.0 |
| N.p. | 3.8–6.7 | 25.6–31.2 |
| A.a. | 1.8–2.9 | 7.0–10.3 |
Рис. 1.
Сравнительная характеристика значений популяционных показателей для групп признаков. а – степень внутрипопуляционного разнообразия; б – структура внутрипопуляционного разнообразия; в – стабильность онтогенеза. M.l. – европейская норка, N.v. – американская норка, M.p. – лесной хорь, M.m. – лесная куница, V.v. – лисица, N.p. – енотовидная собака, A.a. – лось; для рис. 1–3. 1 – одонтологические признаки, 2 – краниологические признаки.
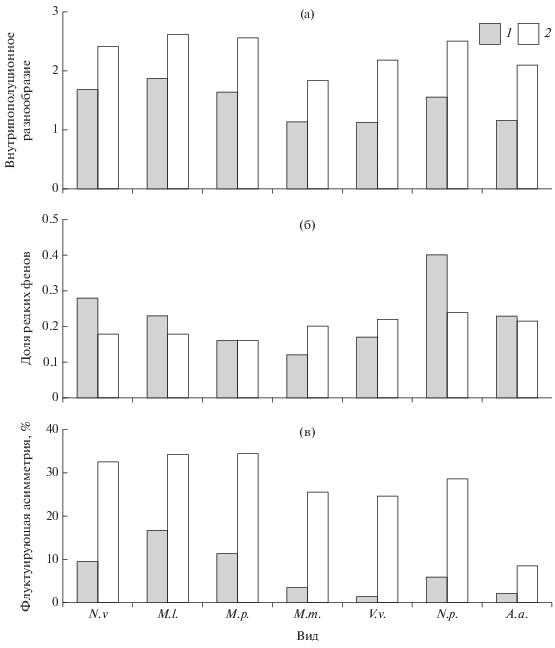
Одонтологические признаки характеризуются достоверно меньшими значениями показателей разнообразия и флуктуирующей асимметрии, что может объясняться как меньшей вариабельностью, допускаемой естественным отбором, так и бо́льшим удельным весом наследственных факторов в их формировании по сравнению с собственно краниологическими признаками. При анализе частот проявления фенов для каждого вида обнаруживаются более высокие значения частот доминирующих вариаций и нередкие случаи мономорфизма, что не характерно для краниологических признаков.
Вместе с тем многими антропологами отмечено совпадение в общих чертах, но не в деталях, при сопоставлении одонтологических признаков с классическими расово-диагностическими признаками антропометрии и антропоскопии (Зубов, Халдеева, 1989). Тенденция синхронного колебания значений популяционных характеристик прослеживается и в нашем случае как для внутрипопуляционного разнообразия, так и для флуктуирующей асимметрии. В первом случае она достоверно не подтверждается коэффициентом корреляции Спирмена (R = 0.75, p > 0.05), а во втором случае положительная корреляция достоверна (R = 0.81, p < 0.05). Еще более очевидна связь между асимметрией и разнообразием, одинаково ярко проявляющаяся для одонтологических (R = 0.94, p < 0.05) и краниологических (R = 0.79, p < 0.05) признаков (рис. 2, 3).
Рис. 2.
Взаимосвязь уровней флуктуирующей асимметрии (штриховые кривые) и внутрипопуляционного разнообразия (сплошные кривые) одонтологических (а) и краниологических (б) признаков у семи видов млекопитающих.
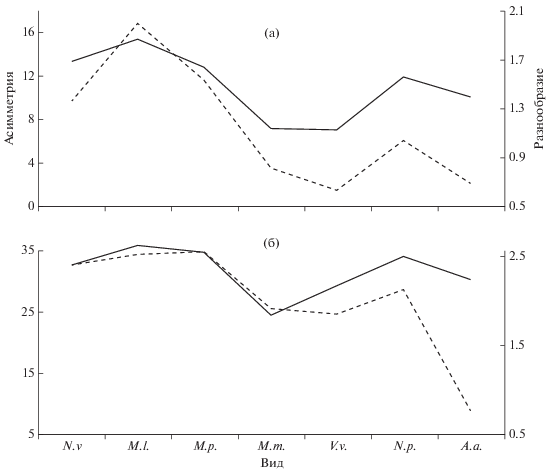
Рис. 3.
Дендрограммы фенетических дистанций и значения показателей сходства трех видов куньих. а – краниологические признаки, б – одонтологические признаки; W – уровень статистических различий в парных сравнениях.

Однако в структуре разнообразия (рис. 1б) подобной согласованности нет. Очевидно, это следствие разной реактивности признаков на воздействие различных факторов, хотя и в структуре разнообразия видов тоже прослеживается определенная логика. Показатель “доля редких фенов” у американской и европейской норок, а также у енотовидной собаки имеет бо́льшие значения для одонтологических признаков, чем для краниологических. Это виды, в популяциях которых можно ожидать большую интенсивность генетических и эпигенетических процессов, определяемую демографическими тенденциями, причем для интродуцированных видов она положительная, а для аборигенной европейской норки – отрицательная.
Особенно контрастны различия между группами признаков по уровню флуктуирующей асимметрии (табл. 4 , рис. 1в). При этом отмеченная тенденция характерна как для признаков корневой системы зубов хищных млекопитающих, так и для особенностей строения коронок зубов лося. Есть основания считать, что низкие уровни вариабельности и асимметричности признаков зубной системы определяются относительно большим удельным весом генетических факторов в их формировании. Антропологами накоплено достаточно данных, свидетельствующих, что морфологические особенности строения зубов, которые могут быть сведены к дискретным характеристикам, имеют высокую степень генного контроля, хотя и они подвержены влиянию средовых факторов и действию различных изменений онтогенеза (Зубов, Халдеева, 1989). Показано, например, что флуктуирующая асимметрия одонтологических признаков у человека отражает уровень стресса в популяции и более выражена у древних представителей гоминид и в этнических группах с архаичным типом хозяйствования.
Основываясь на этих рассуждениях, можно заключить, что факторы внешней среды и внутрипопуляционные процессы оказывают большее влияние на формирование собственно краниологических признаков. Следовательно, они более реактивны в отношении этих факторов и процессов и отражают менее значительные “события” в онтогенезе. Одонтологические признаки, таким образом, в большей степени отражают генетические и эпигенетические процессы, проходящие в популяциях.
Четко выраженная тенденция в различиях значений популяционных показателей для двух совокупностей признаков позволяет сделать вывод, что они контролируются разными группами генов и их анализ целесообразно проводить отдельно, хотя и средние значения параметров тоже информативны. Возможное разное соотношение генетических и средовых факторов при реализации одонтологических и собственно краниологических признаков может быть весьма важным качеством групп признаков при рассмотрении изменчивости популяций. Поскольку отбор действует на фенотипическом уровне, всегда важно и интересно знать, какая часть наблюдаемой изменчивости генетическая, а какая определяется совокупностью других факторов. В подобных ситуациях генетики старались решить этот вопрос изучением признаков с простой наследственной основой (предпочтительно контролируемых одним геном), у которых альтернативными аллелями обусловливаются ярко выраженные фенотипические эффекты, легко поддающиеся измерению или описанию (Солбриг, Солбриг, 1982). При изучении признаков, имеющих полигенную и плейотропную природу, эта задача представляется невыполнимой. Однако использование двух групп признаков, не давая количественную характеристику соотношения факторов при формировании фенотипа, позволяет обсуждать этот вопрос хотя бы на качественном уровне.
Отмеченная особенность групп признаков делает их удобным инструментом для изучения роли экологической ниши в формировании фенотипических особенностей гомологичных структур у таксономически близких симпатрических видов (европейская норка, лесной хорь и американская норка, которая в результате непродуманного эксперимента (ложной идеи насильственного повышения разнообразия животного мира) вынуждена делить с ними экологическую нишу).
Крайняя актуальность эволюционно-экологического анализа закономерностей феногенетической изменчивости гомологичных морфоструктур была отмечена ранее (Васильев и др., 2010), причем на основе изучения 46 внутривидовых форм и видов с разной экологической специализацией в пределах семейства Cricetidae было показано, что сходные экологические требования среды приводят к историческим однонаправленным преобразованиям гомологичных структур у разных видов.
Сравнительный анализ генетического и морфологического разнообразия популяций куньих позволил сделать принципиально важное обобщение: у экологически близких видов при разной степени генетического разнообразия может формироваться сходный уровень фенетического полиморфизма в соответствии с требованиями среды (Кораблев, 2018).
Еще раньше этой проблемой занимался Егоров (Егоров, 1983), проводя сравнительное изучение краниометрических признаков у пяти видов представителей семейства куньих (M. lutreola, M. putorius, M. eversmanni, M. erminea, N. vison). Используя коэффициент дивергенции как показатель филогенетической близости, он показал, что европейская норка морфологически наиболее близка к американской, а горностай – наиболее удаленный среди рассматриваемых видов, что и следовало ожидать. Эти выводы подтвердились и при сравнительном анализе размерных признаков посткраниального скелета. Иные результаты были получены при изучении изменчивости корневой системы премоляров. По числу корней Р1 два вида норок различались между собой в 100% случаев. Таким образом, M.l. имела всегда один корень, N.v. – два. Оба однокорневых премоляра были отмечены у 84.4% лесных хорей, а у 12.2% – с одной из сторон. Два корня у нижнего первого премоляра было у 94.4% европейской норки при 100% у американской. У лесного хоря оба нижних первых премоляра имели по два корня в 53.2% случаях, а в 37.6% случаях – только слева или справа. Автор констатирует, что по вариациям зубных признаков американская норка демонстрирует большее сходство с колонком и горностаем, а европейская – с хорем, что свидетельствует о параллельном развитии некоторых морфологических структур.
На рис. 3 приведены дендрограммы фенетических дистанций (1 – r, метод построения UPGMA) и значения показателей сходства, полученные как среднее для внутривидовых выборок (r ± Sr) между тремя видами куньих.
Между европейской норкой и лесным хорем сходство по одонтологическим признакам оказалось бо́льшим, чем по краниологическим. Иными словами, морфологические различия оказались больше условно генетических. Это вполне объяснимо с экологических позиций. Будучи генетически очень близкими видами (вплоть до единичных, но достоверных случаев гибридизации в естественных условиях), они занимают разные экологические ниши. Европейская норка среди трех видов – наибольший стенобионт, привязанный к водоемам. Добыча ее жертв в водной среде происходит в 60–65% случаев (Сидорович и др., 1997). Основу ее питания при ограниченном обилии или отсутствии широкопалого рака составляют рыбы и амфибии. На их долю, по данным разных авторов, приходится 53–70% всего рациона (Данилов, Туманов, 1976; Сидорович и др., 1997). Лесной хорь, напротив, обладает наибольшей эвритопностью. Он равномерно заселяет различные биотопы и охотно селится вблизи животноводческих помещений и жилья человека и в основном потребляет мелких млекопитающих, амфибий и птиц, причем встречаемость мышевидных грызунов в пище может достигать 95.8% (Данилов, Туманов, 1976). Подобная экологическая разобщенность европейской норки и лесного хоря способствует формированию в целом схожих, но различных в мелких деталях строения морфологических черт, что и нашло отражение в значении фенетической дистанции по комплексу краниологических признаков.
Значение показателя сходства двух видов норок по одонтологическим признакам с учетом статистической ошибки оказалось практически таким же, как при сравнении европейской норки и лесного хоря, хотя генетические различия между норками, казалось бы, очевидны. В этом нет ничего удивительного, поскольку мы анализируем не генетические признаки, а лишь группы признаков с разным удельным весом генетических факторов в их формировании. Напомним, что многие консервативные признаки – побочный продукт хорошо интегрированного генного комплекса – могут не реагировать даже на глубокие перестройки генотипа (Майр, 1974). Два вида норок чрезвычайно далеки в генетическом отношении (Терновский, Терновская, 1994). Это убеждение основывается на сравнительных иммунологических исследованиях сывороточных белков, изучении повторов ДНК и кариотипа.
Однако в данном случае примечательно то, что сходство норок по краниологическим признакам больше, чем сходство между европейской норкой и хорем. Это отражение их большей экологической близости. Американская норка одинаково хорошо адаптирована к полуводному и сухопутному образу жизни. В ее рационе относительно равномерно представлены мелкие млекопитающие, земноводные, рыбы и в меньшем количестве птицы. Тем самым американская норка осваивает значительную часть экологической емкости, используемой европейской норкой и хорем. Именно поэтому сходство американской норки по краниологическим признакам с лесным хорем и аборигенной норкой одинаково высокое. Различия американской норки и хоря по комплексу одонтологических признаков самые большие, что, вероятно, указывает на их бо́льшие генетические различия.
Таким образом, перекрывание экологических ниш трех симпатрических видов куньих способствует поддержанию сходного фенотипа, внутри которого наблюдается слабая дивергенция гомологичных фенотипических признаков вследствие незначительных, но жизненно важных биотопических и трофических различий. Использование двух групп признаков делает эколого-эволюционный анализ более информативным и аргументированным.
Отмеченная зависимость обсуждаемых показателей в определенной степени зависит от особенностей методического подхода при изучении частот неметрических вариаций. В фенетических исследованиях вариации фиксируются, как правило, на одной из сторон. Поскольку в нашем случае фены регистрировались на правой и левой сторонах черепа, асимметричное проявление вариаций соответственно тоже фиксировалось на обеих сторонах, что способствовало увеличению степени реализации фенофонда. Вместе с тем влияние методического подхода не носит абсолютный характер.
На рис. 4 показана изменчивость популяционных показателей для европейской норки в разные периоды сбора черепов. Период сбора краниологического материала составляет >20 лет (с 1983 по 2006 г.). Этот промежуток времени можно разделить на два периода жизни популяционной группировки. Первый период (с 1983 по 1992 г.) характеризуется демографической стабильностью и высокой плотностью, второй (с 1994 по 2006 г.) – стремительным сокращением численности и угасанием очага обитания вида на данной территории. Хотя дискуссия о причинах повсеместного исчезновения европейской норки продолжается, очевидно, что появление ее экологического двойника (американской норки) делает этот процесс быстрым и необратимым. Появление сильного конкурента и быстрый рост численности интродуцированного вида вызвали стресс у аборигенной норки, который отразился в том числе на изменении популяционных показателей.
Рис. 4.
Изменчивость популяционных показателей для европейской норки в разные периоды сбора черепов. 1 – одонтологические признаки, 2 – краниологические признаки.

На диаграммах, иллюстрирующих этот процесс, видно, что во второй период заметно выросла флуктуирующая асимметрии одонтологических (50%) и краниологических (38%) признаков, в то время как разнообразие одонтологических признаков увеличилось на 22%, а краниологических снизилось на 1%. Разнонаправленность векторов изменчивости показателей для краниологических признаков, таким образом, является нелогичной и лишь подчеркивает силу влияния негативных факторов (стресса) на состояние фенофонда группировки европейской норки. Второй период сбора черепов европейской норки в большей степени соответствует сроку сбора основной массы материала для других видов, поэтому на рис. 1 и 2 приводятся данные для европейской норки именно второго периода.
Закономерно, что в конкурентные отношения между норками (американской и европейской) вовлечен лесной хорь, асимметрия признаков у которого сопоставима с таковой у европейской норки. Несмотря на то что хорь по сравнению с аборигенной норкой гораздо больший эврибионт, его излюбленные стации – околоводные биотопы. Именно эти территории активно заселила американская норка, которая благодаря своей биотопической пластичности освоила экологическую емкость, принадлежащую европейской норке и лесному хорю. Проявление американской норкой выраженной синантропности, нарушившее монополию лесного хоря на эту нишу, еще более усугубило трофические проблемы последнего.
Факт, что разные системы признаков достоверно различаются по значениям популяционных характеристик, необходимо учитывать при сравнительном анализе структуры фенофонда группировок млекопитающих, что особенно актуально при использовании данных, полученных разными авторами.
При оценке состояния популяций на основе стабильности онтогенеза (флуктуирующая асимметрия) комплексное использование двух систем признаков позволяет как выявлять начальные стадии нарушений индивидуального развития (краниологические признаки), так и оценивать глубину этих нарушений (одонтологические признаки).
Список литературы
Васильев А.Г., Васильева И.А., Большаков В.Н. Эволюционно-экологический анализ закономерностей феногенетической изменчивости гомологичных морфоструктур: от популяций до экологических рядов видов // Экология. 2010. № 5. С. 323–329.
Данилов П.И., Туманов И.Л. Куньи Северо-Запада СССР. Л.: Наука, 1976. С. 8–10.
Егоров Ю.Е. Механизмы дивергенции. М.: Наука, 1983. 172 с.
Животовский Л.А. Показатель популяционной изменчивости по полиморфным признакам // Фенетика популяций. М.: Наука, 1982. С. 38–44.
Захаров В.М. Асимметрия животных (популяционно-феногенетический подход). М.: Наука, 1987. 216 с.
Зубов А.А., Халдеева Н.И. Одонтология в современной антропологии. М.: Наука, 1989. 231 с.
Кораблев М.П. Внутрипопуляционная изменчивость хищных млекопитающих в центре европейской части России: Афтореф. дис. канд. биол. наук. М.: ИПЭЭ РАН, 2018. 26 с.
Кораблев М.П., Кораблев Н.П., Кораблев П.Н. Морфо-фенетический анализ популяций американской норки (Neovison vison) Каспийско-Балтийского водораздела // Рос. журн. биол. инвазий. 2012а. № 4. С. 36–56.
Кораблев М.П., Кораблев Н.П., Кораблев П.Н., Туманов И.Л. Внутрипопуляционный полиморфизм лесной куницы (Martes martes, Carnivora, Mustelidae) Тверской области // Зоол. журн. 2016а. Т. 95. № 1. С. 80–93.
Кораблев М.П., Кораблев П.Н., Кораблев Н.П., Туманов И.Л. Характеристика полиморфизма исчезающей популяции европейской норки (Mustela lutreola, Carnivora, Mustelidae) в районе Центрально-Лесного заповедника // Зоол. журн. 2013. Т. 9. № 10. С. 1259–1268.
Кораблев М.П., Кораблев П.Н., Кораблев Н.П., Туманов И.Л. Внутрипопуляционный полиморфизм лесного хоря (Mustela putorius, Carnivora, Mustelidae) // Зоол. журн. 2015. Т. 94. № 5 С. 580–592.
Кораблев Н.П., Кораблев М.П., Кораблев П.Н. Краниометрическая изменчивость енотовидной собаки (Nyctereutes procyonoides Grey, Carnivora, Canidae) Тверской области: от интродуцентов до современных популяций // Бюл. МОИП. Отд. биол. 2012б. Т. 117. Вып. 1. С. 16–25.
Кораблев Н.П., Кораблев М.П., Кораблев П.Н., Туманов И.Л. Эпигенетическая изменчивость американской норки Neovison vison Восточной Европы: поиск факторов полиморфизма // Экология. 2016б. № 3. С. 221–228.
Кораблев П.Н. Лось (Alces alces) // Популяционная фенетика. Каталог основных неметрических вариаций краниологических признаков млекопитающих. М.: Наука, 1997. С. 247–254.
Кораблев П.Н. Динамика фенофонда популяции лося (Alces alces) Центрально-Лесного заповедника и факторы ее определяющие // Заповедное дело. 2003. Вып. 11. С. 44–61.
Кораблев П.Н., Кораблев Н.П., Кораблева В.Н., Кораблев М.П. Методические рекомендации по изучению фенофонда популяций хищных млекопитающих // Методические рекомендации по ведению мониторинга на особо охраняемых природных территориях (на примере Центрально-Лесного государственного природного биосферного заповедника). М.: Экотерра, 2005. С. 185–231.
Майр Э. Популяции, виды и эволюция. М.: Мир, 1974. 460 с.
Назаров В.И. Эволюция не по Дарвину: Смена эволюционной модели. Изд. 2-е, испр. М.: ЛКИ, 2007. 520 с.
Сидорович В.Е., Крук Х., Макдональд Д.В. Тестирование конкуренции в гильдии куньих // Куньи в Беларуси. Эволюционная биология, демография и биоценотические связи. Минск: Золотой улей, 1997. С. 107–114.
Солбриг О., Солбриг Д. Популяционная биология и эволюция. М.: Мир, 1982. 488 с.
Терновский Д.В., Терновская Ю.Г. Экология куницеобразных. Новосибирск, 1994. 200 с.
Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В. Очерк учения о популяции. М.: Наука, 1973. 278 с.
Яблоков А.В. Фенетика. М.: Наука, 1980. 135 с.
Яблоков А.В. Популяционная биология. М.: Высш. шк., 1987. 303 с.
Berry R.J. Epigenetic polymorphism in wild populations of Mus musculus // Genet. Res. Cambridge, 1963. V. 4. P. 193–220.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая