

Известия РАН. Серия биологическая, 2020, № 2, стр. 144-152
Онтогенетический спектр как индикатор состояния ценопопуляций растений
Г. О. Османова 1, *, Л. А. Животовский 2
1 Марийский государственный университет
пл. Ленина, 1, Йошкар-Ола, 424000 Россия
2 Институт общей генетики им. Н.И. Вавилова РАН
ул. Губкина, 3, Москва, 119991 Россия
* E-mail: gyosmanova@yandex.ru
Поступила в редакцию 26.08.2019
После доработки 25.10.2019
Принята к публикации 25.10.2019
Аннотация
Отмечено, что онтогенетический спектр – чувствительный популяционный индикатор изменения условий среды – регистрирует возрастное состояние растений. Дан обзор популяционно-онтогенетических методов для исследования популяций (ценопопуляций) растений, основанных на данных об их онтогенетических спектрах. Введены новые возрастные индексы: молодости и зрелости. Для индекса восстановления предложены границы для степени самоподдержания ценопопуляций.
В настоящее время популяционно-онтогенетический метод исследований в ботанике и экологии растений получил широкое распространение, так как он не только основывается на визуальных оценках, но и учитывает разные показатели, характеризующие развитие растений в условиях конкретного растительного сообщества. В частности, важная особенность каждой ценопопуляции – ее онтогенетический спектр (распределение особей по онтогенетическим состояниям).
Цель работы – обзор показателей, характеризующих онтогенетический спектр (Османова, Животовский, 2017).
Онтогенетические состояния растений. Онтогенез особи начинается с зиготы (изредка – с гаметы), споры, или вегетативного зачатка, откуда разворачивается программа развития организма, и заканчивается его угасанием и смертью. Развитие и рост растения критически зависят от условий обитания и могут значительно варьировать как в пределах одной ценопопуляции, так в разных местообитаниях. Онтогенетические состояния – это последовательные этапы онтогенеза особей данного вида, которые отличаются один от другого набором диагностических качественных признаков.
Темпы развития растений могут быть разными. Длительность пребывания растений в том или ином онтогенетическом состоянии прежде всего зависит от жизненной формы, а в пределах одной жизненной формы – от внутри- и межвидовой конкуренции, влияния факторов среды. Например, на богатых и влажных почвах темпы развития отдельных состояний древесных растений несколько ускоряются, причем общая продолжительность жизни сокращается. На бедных же почвах развитие замедляется, растения медленно переходят из одного состояния в другое, а общая продолжительность жизни увеличивается. У травянистых растений продолжительность отдельных состояний также определяет общую продолжительность онтогенеза растения. Многолетние наблюдения за особями ежи сборной Dactylis glomerata L. и овсяницы луговой Festuca pratensis Huds. в агроценозах показали, что из-за уменьшения продолжительности жизни прегенеративного периода некоторые особи этих видов завершили свой полный онтогенез за 4–5 вместо 10–20 лет (Ермакова, Жукова, 1985). При изучении луговика дернистого Deschampsia cespitosa (L.) Beauv. на пойменных лугах Северной Двины (Жукова, 1987) были выявлены 7–8-летние ювенильные растения, хотя в этом онтогенетическом состоянии растение находится не более одного вегетационного периода. Таким образом, можно отметить, что каждый этап онтогенеза растения соответствует его онтогенетическому состоянию, но не его календарному возрасту.
Помимо календарного возраста каждое растение характеризуется своим онтогенетическим возрастом, который определяется качественными преобразованиями организма во времени. В частности, онтогенез цветковых растений (исключая предзародышевые состояния) – это “последовательность сменяющих друг друга морфологических состояний и изменений растений от прорастания семени до отмирания особи и – в случае вегетативного размножения – всего вегетативно возникшего потомства” (Уранов, 1967).
Классификация онтогенетических состояний. Онтогенетические состояния определяют с учетом способа питания, способности к семенному размножению, наличия зародышевых, ювенильных и взрослых структур, побегов, листьев и корневых систем, а также соотношения процессов новообразования органов и их отмирания, целостности особей или этапов их разделения на дочерние. Различают 12 онтогенетических состояний: se – семя, p – проросток, j – ювенильное, im – имматурное, v – виргинильное, g0 – скрытогенеративное, g1 – молодое генеративное, g2 – средневозрастное генеративное, g3 – старое генеративное, ss – субсенильное, s – сенильное, sc – отмирающее (Работнов, 1950; Уранов, 1975; Жукова и др., 1976; Шестакова, 1991; Нухимовский, 1997). Скрытогенеративное состояние это одно из онтогенетических состояний особи, присутствующее у всех растений, только не всегда явно различимое, поэтому в подавляющем большинстве популяционных исследований оно не выделяется.
Онтогенетический спектр (основные понятия). Растения, находящиеся в данном онтогенетическом состоянии, называют онтогенетической группой. Распределение особей ценопопуляции по онтогенетическим группам называют спектром онтогенетических состояний, или кратко онтогенетическим спектром (иногда возрастными состояниями и возрастным спектром). Онтогенетический спектр – демографическая характеристика ценопопуляции. Его можно представить в табличной или графической форме в виде или абсолютных численностей, или в долях единицы, или как процентные соотношения разных онтогенетических групп. При изучении онтогенетической структуры популяций в большинстве случаев проростки не учитывают в связи с их лабильностью; также обычно не учитывают отмершие растения ввиду сложности их учета. Эти формы и числовые представления одинаково отражают соотношение разных онтогенетических групп.
Характерный и базовый онтогенетические спектры. Онтогенетический спектр – лабильная характеристика популяционного состава: разные ценопопуляции могут иметь разные онтогенетические спектры, тип спектра в одной и той же ценопопуляции может меняться со временем, зависеть от естественных средовых влияний, антропогенных воздействий и прочих факторов. Тем не менее спектры онтогенетических состояний нормальных ценопопуляций разных видов растений в определенном диапазоне условий могут иметь сходный характер. Это позволяет выделить четыре теоретических типа онтогенетических спектров, характерных для данного вида, типа онтогенеза или жизненной формы и определяемых по соотношению частот встречаемости онтогенетических групп и положением их максимумов по фактическим данным о разных ценопопуляциях с учетом теоретических поправок и экспертных оценок. Они названы характерными онтогенетическими спектрами (Заугольнова, 1994) (рис. 1).
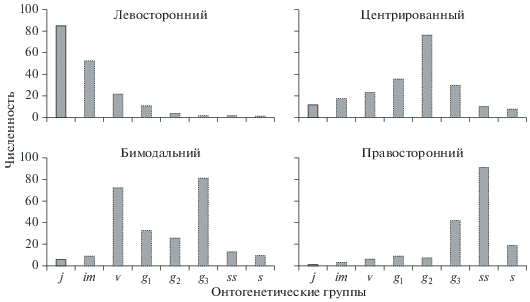
Левосторонний спектр – это спектр, в котором доминируют особи прегенеративного периода с максимумом на одной из этих онтогенетических групп. Центрированный спектр – это спектр, в котором максимум приходится на средневозрастные генеративные растения (g2). Бимодальный спектр – это спектр, в котором отмечены два пика: на молодых (включая g1) и на старых растениях (начиная с g3-растений). Правосторонний спектр – это спектр с преобладанием особей постгенеративного периода.
Наряду с характерным спектром можно рассмотреть базовый онтогенетический спектр, т.е. модальную оценку или усреднение по многим ценопопуляциям данного вида или ряда видов данной жизненной формы (Заугольнова, 1976). Базовый онтогенетический спектр может не совпадать с теоретически вводимым характерным спектром, так как он является чисто эмпирической оценкой и определяется биологическими свойствами вида: общей продолжительностью онтогенеза и отдельных состояний, темпами развития особей, способами самоподдержания, интенсивностью и периодичностью инспермации, почвенным запасом диаспор, размерами площади поглощения ресурсов особями разных онтогенетических состояний.
Можно предложить количественную меру для выделения спектров, представленных на рис. 1. Для этого обратимся к типизации форм базовых спектров (Заугольнова, 1976), в которой учитываются соотношения численностей молодых и старых растений и расположение максимума спектра, и применим эту типизацию для характерных спектров (Животовский, Османова, 2019). В этом случае U – сумма численностей особей онтогенетических состояний v, g1 и g2, а D – сумма численностей особей g3, ss и s. Спектр относится к левостороннему типу, если U > D и максимум приходится на одну из онтогенетических групп в молодой части ценопопуляции (v и младше), и к центрированному типу, если U > D и максимум приходится на группу особей в g2-состоянии. Спектр относится к правостороннему типу, если U < D и максимум приходится на одну из онтогенетических групп постгенеративного периода (ss и старше). В случае бимодального спектра можно выделить два подтипа: бимодальный левосторонний (если U > D) и бимодальный правосторонний (если U < D).
Классификация ценопопуляций по онтогенетическим спектрам. Исследование онтогенетической структуры ценопопуляций растений разных жизненных форм, произрастающих в разных географических условиях, привело к необходимости создания целого ряда классификаций, каждая из которых охватывает разные типы онтогенетических спектров ценопопуляций растений. Однонаправленное развитие сукцессивных ценопопуляций растений было описано ранее (Работнов, 1950). В частности, были выделены три основных типа ценопопуляций по характеру онтогенетического спектра: инвазионный, нормальный и регрессивный, которые соответствуют возникновению, полному развитию и угасанию популяций. Инвазионная ценопопуляция – это ценопопуляция, целиком состоящая из особей прегенеративного периода (подроста), а потому зависящая от заноса зачатков извне и не способная в данный момент к самоподдержанию (рис. 2а). Нормальная ценопопуляция – это ценопопуляция, включающая в себя растения всех онтогенетических состояний (рис. 2б); регрессивная – это ценопопуляция, которая содержит в основном особей постгенеративного периода и потому потеряла способность к самоподдержанию (рис. 2в).
Рис. 2.
Спектр онтогенетических состояний инвазионной (а), нормальной (б), регрессивной (в), инвазионно-регрессивной (г) ценопопуляций. se – почвенный банк семян; его размер, как правило, неизвестен.
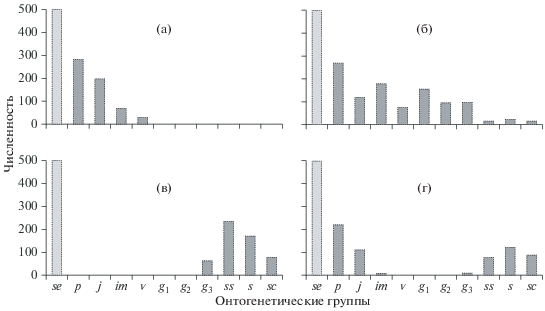
Кроме этих трех основных типов ценопопуляций был выделен переходный тип – инвазионно-регрессивный (Рысин, Казанцева, 1975). Инвазионно-регрессивная ценопопуляция – это ценопопуляция, состоящая из угнетенных особей прегенеративного и постгенеративного периодов (рис. 2г).
Инвазионные ценопопуляции. Каждый из указанных трех основных типов ценопопуляций подразделяется далее на классы, в частности среди инвазионных выделяют три класса ценопопуляций – I1, I2, I3 (Рысин, Казанцева, 1975) (рис. 3).
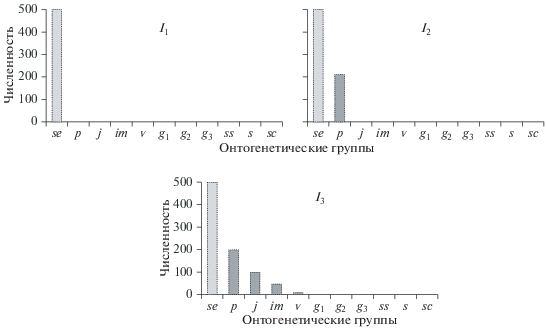
Ценопопуляции I1 состоят только из почвенного банка семян и/или покоящихся вегетативных диаспор. По наблюдениям Г.А. Богданова (устное сообщение), в окрестностях горы Карман-Курык Моркинского р-на Республики Марий Эл произрастает небольшая ценопопуляция лука Вальдштейна Allium waldsteinii G. Don. fil. Вследствие аномально жаркого лета 2010 г. в дальнейшем эта ценопопуляция исчезла. Результаты почвенных исследований свидетельствуют о наличии в почве большого числа вегетативных диаспор – луковичек (>500 штук на площадке 0.25 м2), благодаря которым ценопопуляция восстановилась через несколько лет.
Ценопопуляции I2 также включают в себя проростки. Иными словами, такая ситуация свидетельствует о начале развития ценопопуляции в данном местообитании. В ценопопуляциях I3 появляются также особи прегенеративного периода (тенденция к возобновлению данной ценопопуляции).
Нормальные ценопопуляции. Среди нормальных популяций выделяют следующие шесть классов (рис. 4, 5): молодые, зрелые, старые (Жукова, 1967), стареющие (Уранов, Смирнова, 1969), зреющие и переходные (Животовский, 2001). Критерий молодых ценопопуляций – преобладание в онтогенетическом спектре особей прегенеративного периода (подроста), а среди генеративных – доминирование группы g1. К зрелым относят ценопопуляции с наличием подроста и преобладанием среди генеративных особей группы g2. Старые – это ценопопуляции с небольшой долей подроста и преобладанием среди генеративных группы g3-особей. Стареющие ценопопуляции – это промежуточные между зрелыми и старыми ценопопуляциями, с небольшой долей подроста (в основном виргинильных особей) и сопоставимыми долями особей в g2- и g3-состояниях.
Рис. 4.
Онтогенетические спектры нормальных ценопопуляций – молодой (а), зрелой (б), стареющей (в), старой (г).
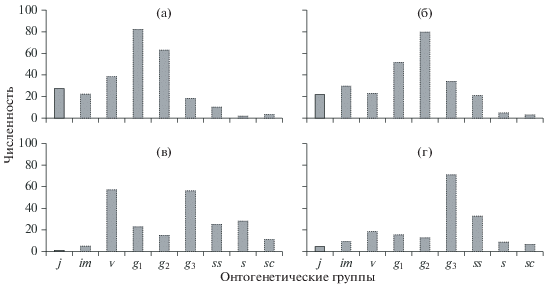
Рис. 5.
Классификация “дельта–омега”. Цифры – значения Δ и ω, разделяющие классы ценопопуляций; точки – примеры ценопопуляций.
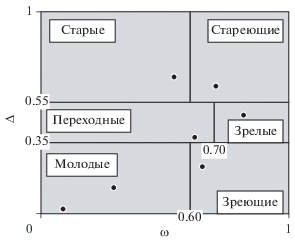
Перечисленные три основных класса нормальных ценопопуляций (молодой, зрелый и старый) предполагают, что онтогенетический спектр имеет один максимум (унимодальность) (рис. 4). Однако в небольших выборках этот максимум может быть смещен. Кроме того, нередко встречаются спектры с двумя и более максимумами, поэтому критерий максимума не всегда объективен. Дело в том, что критерий максимума принимает во внимание доминирование только особей одного онтогенетического состояния и не учитывает другие онтогенетические состояния. Поэтому была предложена классификация “дельта–омега”, основанная на совместном использовании индексов возрастности Δ (Уранов, 1975) и средней эффективности ω (Животовский, 2001), учитывающая особи всех онтогенетических состояний (рис. 5).
Классификация “дельта–омега” позволила дополнительно выделить ценопопуляции “переходного” типа, а из “молодых” ценопопуляций удалось выделить “зреющие”, т.е. ценопопуляции с высокой долей генеративных особей. Указанные классификации основаны на разных критериях, поэтому могут по-разному оценивать возрастной статус ценопопуляций.
Регрессивные ценопопуляции. Среди регреcсивных выделяют четыре класса ценопопуляций: R1–R4 (Жукова и др., 1976) (рис. 6). Ценопопуляции R1 состоят из старых генеративных особей (цветущих, но не плодоносящих или образующих некачественные семена, и особей постгенеративного периода. Ценопопуляции R2 включают в себя только особей постгенеративного периода. В популяциях R3 имеются только сенильные и отмирающие растения. В ценопопуляциях R4 полностью отсутствуют живые особи.
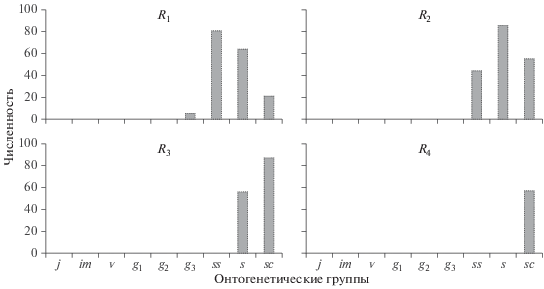
Хотя регрессивные ценопопуляции считаются погибщими, нельзя полностью исключить их восстановление в ближайшем или отдаленном будущем. Начнем с того, что даже в угасающих ценопопуляциях R4 некоторые отмирающие растения несут покоящиеся почки, из которых могут развиваться побеги. Более того, во все предыдущие годы существования ценопопуляции в почве шел процесс формирования банка семян этой ценопопуляции. Кроме покоящихся семян в почве могут накапливаться и вегетативные диаспоры. И наконец, возможно появление слабоомоложенных вегетативных потомков за счет старческой партикуляции.
Количественные оценки состояния ценопопуляций. Популяционные исследования базируются не только на визуальных оценках. Успешность анализа полученных данных определяется также использованием количественных показателей, оценивающих численность, плотность, приспособленность, онтогенетические спектры и другие характеристики ценопопуляций.
Численность (N) – это число всех особей ценопопуляции или определенных возрастных, половых или иных ее когорт. Иногда используют термин “размер ценопопуляции” как синоним численности. Плотность (p) – это численность, деленная на площадь (S) занимаемого ценопопуляцией пространства (p = N/S) и интерпретируемая как число особей на единицу площади пространства. Например, плотность ценопопуляции березы повислой Betula pendula Roth. – 250 деревьев на 1 га, а плотность подорожника ланцетолистного Plantago lanceolata L. – 35 особей на 1 м2.
Плотность, рассчитанную по общей площади без учета пятнистости ареала, называют средней плотностью. Плотность, отнесенную только к участкам произрастания растений данного вида, называют экологической плотностью ценопопуляции. Средняя плотность ценопопуляции всегда меньше ее экологической плотности во столько раз, во сколько площадь ее местообитаний меньше общей учетной площади. Методы определения плотности – тотальный учет (для редких видов растений), закладка пробных площадей, дистанционные оценки с помощью аэрокосмических методов анализа ландшафтов.
Оценка эффективности самоподдержания ценопопуляций. Для нее можно использовать индексы восстановления (Iв) и замещения (Iз). Индекс восстановления оценивает число потомков, приходящихся на одно растение генеративного периода (Жукова, 1987):
Если Iв > 1, то это означает, что на одно генеративное растение приходится более одного потомка.
Можно предложить условные границы между возможностями самоподдержания ценопопуляции (Животовский, Османова, 2019): если Iв > 2, то ценопопуляция самоподдерживается эффективно; при 1 < Iв < 2 ценопопуляция самоподдерживается умеренно; при Iв < 1 ценопопуляция самоподдерживается слабо.
Индекс замещения оценивает число потомков, приходящихся на одно взрослое растение генеративного или постгенеративного периодов (Жукова, 1987):
Если Iз > 1, то это означает, что на одно взрослое растение приходится более одного потомка.
Показатель сходства онтогенетических спектров. В разных эколого-ценотических условиях онтогенетические спектры ценопопуляций одного вида могут меняться, т.е. изменяются частоты онтогенетических состояний. Поэтому для оценки отличий ценопопуляции по онтогенетическим спектрам можно использовать различные меры сходства или расстояния, в частности показатель сходства r (Животовский, 1979, 1982), который оценивает частоту общих морф (в данном случае – частоту общих онтогенетических состояний) в двух сравниваемых ценопопуляциях (онтогенетическое состояние считается общим, если в обеих сравниваемых выборках присутствуют особи в этом состоянии):
где p1, p2, …, pk – частоты особей состояний 1, 2, …, k в выборке из первой ценопопуляции; q1, q2, …, qk – частоты особей состояний 1, 2, …, k в выборке из второй ценопопуляции; k – число онтогенетических состояний, обнаруженных хотя бы в одной из сравниваемых ценопопуляций. Эти частоты равны:Минимально возможное значение показателя сходства r равно 0 (общие онтогенетические состояния отсутствуют), максимальное – 1 (спектры частот онтогенетических состояний идентичны в сравниваемых выборках). Его стандартная ошибка (обусловленная объемом выборок) следующая:
где p0 – сумма частот тех онтогенетических состояний в первой выборке, которые не представлены во второй выборке, а соответственно, q0 – сумма частот тех онтогенетических состояний во второй выборке, которые не представлены в первой выборке.
Индексы возрастных периодов. Иногда полезно вычислить популяционные демографические показатели (пропорции молодых, зрелых и старых растений) по отношению к общей численности (плотности) ценопопуляции (Животовский, Османова, 2019):
где Iv, Ig, Is – индексы молодости, зрелости, старения (индекс старения был введен ранее (Глотов, 1998) и обозначен как I2).
Эти индексы применимы ко всем типам ценопопуляций. Они не заменяют выделения молодых, зрелых и старых классов нормальных ценопопуляций, поскольку те основаны на другом принципе. При мониторинге вида на большой территории индексы позволяют провести простую качественную оценку возрастных категорий ценопопуляций по большому числу пробных участков и дать обзор возрастного состояния вида в просто интерпретируемых демографических статистиках.
Индекс возрастности. Ранее (Уранов, 1975) были введены понятие “возрастность особи” (mi) как доля энергии, потребленной ею к данному онтогенетическому состоянию i, по отношению ко всей энергии, доступной особи в течение полного онтогенеза, и понятие “индекс возрастности ценопопуляции” (∆) как средневзвешенные значения mi, где вес – доля растений i-го состояния:
где mi – возрастность особей i-го онтогенетического состояния (табл. 1), ni – число особей i-го состояния в ценопопуляции, N – общее число особей в ценопопуляции.
Таблица 1.
Энергетические характеристики онтогенетических состояний
| Показатель | Онтогенетические состояния | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| se | pl | j | im | v | g1 | g2 | g3 | ss | s | sc | |
| Возрастность, mi | 0.0025 | 0.0067 | 0.0180 | 0.0474 | 0.1192 | 0.2689 | 0.5 | 0.7311 | 0.8808 | 0.9526 | 0.9820 |
| Эффективность, ei | 0.0099 | 0.0266 | 0.0707 | 0.1807 | 0.4200 | 0.7864 | 1.0 | 0.7864 | 0.4200 | 0.1807 | 0.0707 |
Индекс эффективности ценопопуляции. В дополнение к предыдущему индексу А.А. Уранова Л.А. Животовский (Животовский, 2001) ввел понятие “энергетическая эффективность”, или “эффективность растений i-го онтогенетического состояния” (ei), которая оценивает относительную величину его “нагрузки” на энергетические ресурсы среды (значения ei приведены в табл. 1), и соответственно ввел понятие “индекс эффективности ценопопуляции” (ω) – средневзвешенные значения mi, где вес – доля растений i-го онтогенетического состояния:
где ω – индекс эффективности ценопопуляции, ei – относительная эффективность потребления энергии растениями i-го онтогенетического состояния, ni – число особей i-го состояния в ценопопуляции, N – общее число особей в ценопопуляции.
Индекс эффективности можно интерпретировать как среднюю энергетическую нагрузку на среду, вызываемую данной ценопопуляцией.
Скорость развития ценопопуляции. Показатель был введен ранее (Жукова, 1995):
где VΔ – скорость развития ценопопуляции, ∆1 – начальная возрастность, ∆2 – конечная возрастность, (t2 – t1) – интервал времени.
Этот индекс показывает скорость изменения онтогенетического спектра за единицу времени. Если ее измерять относительно начальной возрастности ∆1, то получим специфическую скорость развития ценопопуляции (Жукова, 1995):
${{r}_{{\Delta }}} = {{{{V}_{{\Delta }}}} \mathord{\left/ {\vphantom {{{{V}_{{\Delta }}}} {{{\Delta }_{1}}}}} \right. \kern-0em} {{{\Delta }_{1}}}}.$
Эффективная экологическая плотность ценопопуляции. Растения разных онтогенетических состояний потребляют энергию среды разными темпами, определяемыми значениями ωi. Следовательно, нагрузка данной популяции на энергетические ресурсы среды зависит не только от ее физической численности (плотности), но и от соотношения растений разных онтогенетических состояний в данной популяции. Поэтому вклад растений разных онтогенетических состояний в популяционную плотность должен быть взвешен соответственно их энергетической эффективности.
На этой основе было предложено понятие “эффективная экологическая плотность ценопопуляции” (Животовский, 2001):
где Me – эффективная экологическая плотность ценопопуляции, ω – индекс эффективности ценопопуляции, M – численность (плотность) ценопопуляции.
Эффективная плотность всегда меньше обычной, физической плотности ценопопуляции. Значение Me близко к значению физической плотности для зрелых ценопопуляций, однако для старых и молодых ценопопуляций значения эффективной плотности могут быть гораздо меньше значений обычной плотности.
Таким образом, приведенные показатели позволяют дать количественную оценку состояния ценопопуляций растений по данным об онтогенетических спектрах и выявить их временнýю и пространственную динамику в разных условиях среды.
Работа выполнена в рамках программы гостемы ГЗ 0112-2019-2 (подтема “Эколого-генетическая структура вида”).
Список литературы
Глотов Н.В. Об оценке параметров возрастной структуры популяций растений // Жизнь популяций в гетерогенной среде. Ч. 1. Йошкар-Ола: Периодика Марий Эл, 1998. С. 146–149.
Ермакова И.М., Жукова Л.А. Типы функционирования ежи сборной и овсяницы луговой в луговых агрофитоценозах // Динамика ценопопуляций растений. М.: Наука, 1985. С. 110–126.
Животовский Л.А. Показатель сходства популяций по полиморфным признакам // Журн. общ. биологии. 1979. № 4. С. 587–602.
Животовский Л.А. Показатели популяционной изменчивости по полиморфным признакам // Фенетика популяций / Под ред. Яблокова А.В. М.: Наука, 1982. С. 38–44.
Животовский Л.А. Онтогенетические состояния, эффективная плотность и классификация популяций растений // Экология. 2001. № 21. С. 3–7.
Животовский Л.А., Османова Г.О. Популяционная биогеография растений. Йошкар-Ола: Вертикаль, 2019. 128 с.
Жукова Л.А. Изменение возрастного состава популяций луговика дернистого на окских лугах при разной продолжительности выпаса // Онтогенез и возрастной состав популяций цветковых растений. М.: Наука, 1967. С. 114–131.
Жукова Л.А. Динамика популяций луговых растений: Автореф. дис. д-ра биол. наук. Новосибирск: ЦСБС РАН, 1987. 32 с.
Жукова Л.А. Популяционная жизнь луговых растений. Йошкар-Ола: РИИК “Ланар”, 1995. 224 с.
Жукова Л.А., Заугольнова Л.Б., Смирнова О.В. Введение // Ценопопуляции растений (основные понятия и структура). М.: Наука, 1976. С. 5–12.
Заугольнова Л.Б. Типы возрастных спектров нормальных ценопопуляций растений // Ценопопуляции растений (основные понятия и структура). М.: Наука, 1976. С. 81–92.
Заугольнова Л.Б. Структура популяций семенных растений и проблемы их мониторинга: Автореф. дис. д-ра биол. наук в форме науч. докл. СПб.: БИН, 1994. 70 с.
Нухимовский Е.Л. Основы биоморфологии семенных растений. М.: Недра, 1997. 629 с.
Османова Г.О., Животовский Л.А. Принципы и методы изучения ценопопуляций растений разных жизненных форм // Проблемы популяционной биологии: Матер. XII Всерос. популяционного семинара памяти Николая Васильевича Глотова (1939–2016). Йошкар-Ола, 11–14 апреля 2017 г. Йошкар-Ола: СТРИНГ, 2017. С. 164–166.
Работнов Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах // Тр. БИН АН СССР. Сер. 3, Геоботаника. М.; Л.: 1950. Вып. 6. С. 7–204.
Рысин Л.П., Казанцева Т.Н. Метод ценопопуляционного анализа в геоботанических исследованиях // Бот. журн. 1975. Т. 60. № 2. С. 199–209.
Уранов А.А. Онтогенез и возрастной состав популяций // Онтогенез и возрастной состав популяций цветковых растений. М.: Наука, 1967. С. 3–8.
Уранов А.А. Возрастной состав фитоценопопуляций как функции времени и энергетических волновых процессов // Биол. науки. 1975. № 2. С. 17–29.
Уранов А.А., Смирнова О.В. Классификация и основные черты развития популяций многолетних растений // Бюл. МОИП. Отд. биол. 1969. Т. 74. Вып. 1. С. 119–134.
Шестакова Э.В. Тмин обыкновенный // Изучение проблем популяционной экологии растений. Популяционно-онтогенетические аспекты экологического мониторинга: Отчет о НИР; № ГР 01910056055. Йошкар-Ола: МарГУ, 1991. С. 13–15.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая